

Abstract
AIM: To investigate the effectiveness of antioxidant compounds in modulating mitochondrial oxidative alterations and lipids accumulation in fatty hepatocytes.
METHODS: Silybin-phospholipid complex containing vitamin E (Realsil®) was daily administered by gavage (one pouch diluted in 3 mL of water and containing 15 mg vitamin E and 47 mg silybin complexed with phospholipids) to rats fed a choline-deprived (CD) or a high fat diet [20% fat, containing 71% total calories as fat, 11% as carbohydrate, and 18% as protein, high fat diet (HFD)] for 30 d and 60 d, respectively. The control group was fed a normal semi-purified diet containing adequate levels of choline (35% total calories as fat, 47% as carbohydrate, and 18% as protein). Circulating and hepatic redox active and nitrogen regulating molecules (thioredoxin, glutathione, glutathione peroxidase), NO metabolites (nitrosothiols, nitrotyrosine), lipid peroxides [malondialdehyde-thiobarbituric (MDA-TBA)], and pro-inflammatory keratins (K-18) were measured on days 0, 7, 14, 30, and 60. Mitochondrial respiratory chain proteins and the extent of hepatic fatty infiltration were evaluated.
RESULTS: Both diet regimens produced liver steatosis (50% and 25% of liver slices with CD and HFD, respectively) with no signs of necro-inflammation: fat infiltration ranged from large droplets at day 14 to disseminated and confluent vacuoles resulting in microvesicular steatosis at day 30 (CD) and day 60 (HFD). In plasma, thioredoxin and nitrosothiols were not significantly changed, while MDA-TBA, nitrotyrosine (from 6 ± 1 nmol/L to 14 ± 3 nmol/L day 30 CD, P < 0.001, and 12 ± 2 nmol/L day 60 HFD, P < 0.001), and K-18 (from 198 ± 20 to 289 ± 21 U/L day 30 CD, P < 0.001, and 242 ± 23 U/L day 60 HFD, P < 0.001) levels increased significantly with ongoing steatosis. In the liver, glutathione was decreased (from 34.0 ± 1.3 to 25.3 ± 1.2 nmol/mg prot day 30 CD, P < 0.001, and 22.4 ± 2.4 nmol/mg prot day 60 HFD, P < 0.001), while thioredoxin and glutathione peroxidase were initially increased and then decreased. Nitrosothiols were constantly increased. MDA-TBA levels were five-fold increased from 9.1 ± 1.2 nmol/g to 75.6 ± 5.4 nmol/g on day 30, P < 0.001 (CD) and doubled with HFD on day 60. Realsil administration significantly lowered the extent of fat infiltration, maintained liver glutathione levels during the first half period, and halved its decrease during the second half. Also, Realsil modulated thioredoxin changes and the production of NO derivatives and significantly lowered MDA-TBA levels both in liver (from 73.6 ± 5.4 to 57.2 ± 6.3 nmol/g day 30 CD, P < 0.01 and from 27.3 ± 2.1 nmol/g to 20.5 ± 2.2 nmol/g day 60 HFD, P < 0.01) and in plasma. Changes in mitochondrial respiratory complexes were also attenuated by Realsil in HFD rats with a major protective effect on Complex II subunit CII-30.
CONCLUSION: Realsil administration effectively contrasts hepatocyte fat deposition, NO derivatives formation, and mitochondrial alterations, allowing the liver to maintain a better glutathione and thioredoxin antioxidant activity.
Keywords: Fatty liver, Glutathione, Lipid peroxidation, Nitrosothiols, Nitrotyrosine, Thioredoxin
INTRODUCTION
Simple steatosis of the liver without inflammation is considered a rather benign condition, although it represents a favoring substrate for the potentially damaging effects of a second hit (i.e., ischemia-reperfusion, starvation)[1]. However, severe fatty degeneration represents a leading factor of hepatocyte dysfunction (mitochondrial respiration, microsomal metabolism, biliary secretion)[2] and is associated with excess delivery of nitrosative and oxidative stress molecules[3], thus potentially rendering the liver a major source of systemic alterations in patients with metabolic syndrome[4]. Also, while several adaptive metabolic mechanisms have been described during the early phase of fatty infiltration[1,5] including expression of intracellular sensors and signaling molecules for lipid metabolism and oxidative stress pathways[6,7], the threshold above which fat infiltration becomes dangerous is not clear, so far. Indeed, it has been observed that transient hepatocellular triglycerides accumulation is essential for normal liver regeneration[8] and represents a mechanism of liver protection from lipotoxicity. Buffering free fatty acids might, therefore, prevent the formation of liver steatosis[9]. By contrast, several observations suggest that ongoing fatty degeneration indeed exposes hepatocytes to higher risk of oxidative damages[10].
Experimental rat models of liver steatosis are characterized by accumulation of triglycerides, decreased mitochondrial function, and increased activity of microsomal enzymes[2]. The altered functions of these subcellular organelles favor the enhanced production of reactive oxygen species (ROS) and NO derivatives with consequent morphological and functional modifications of crucial structures, thus rendering fatty hepatocytes particularly susceptible to additional injuring factors[3].
A number of natural or chemical compounds are able to counteract the damages induced by oxidative and nitrosative stress and therefore are claimed to have antioxidant properties. Vitamin E efforts a remarkable protection against lipid oxidation[11] and, if vehicolated within a phospholipid complex together with silybin, a silybum marianum extract, protects against pro-fibrotic oxidative injury[12]. Less is known about the effectiveness of such a compound to block or modulate ROS/NO production and their pro-oxidant effects. Also, it would be of interest to know if the administration of such an antioxidant complex may contribute to break off the intracellular mechanisms leading to a progressive accumulation of neutral lipids in fatty hepatocytes and in particular, mitochondrial dysfunction.
Therefore, this study aimed to evaluate the effect of a silybin-phospholipid complex containing vitamin E (Realsil®) on hepatocyte fatty degeneration and nitrosative/oxidative stress in two different rat dietary models [choline deficiency (CD) and high fat diet (HFD)] of fatty liver. Both diets induce fatty degeneration without major inflammation and fibrosis, representing therefore ideal models for assessing changes associated with simple steatosis without the metabolic consequences depending on inflammation. The results of this study contribute to clarify both pathophysiologic mechanisms of damage in fatty hepatocytes and of pharmacological protection.
MATERIALS AND METHODS
Male Wistar rats (b.w. 250-270 g, Harlan, S. Pietro al Natisone, Italy) were kept under controlled conditions of temperature and humidity and on a 12 h dark/light cycle. Animals were divided into five groups: rats fed a CD diet (Dyets, Bethlem, PA); rats fed a CD diet plus daily administration of Realsil by gavage (one pouch diluted in 3 mL of water and containing 15 mg vitamin E and 47 mg silybin complexed with phospholipids); rats fed a HFD (20% fat content), containing 71% total calories as fat, 11% as carbohydrate, and 18% as protein (Altromin Rieper, Vandoies, Italy); rats fed a HFD supplemented by daily administration of Realsil by gavage; control group fed a normal semi-purified diet containing adequate levels of choline, 35% total calories as fat, 47% as carbohydrate, and 18% as protein. The control diet, CD, and HFD were nutritionally adequate, calorically equivalent (1 kcal/mL), and contained equal amounts of fat as olive and safflower oil with excess corn oil added to the HFD.
Following an overnight fast, five rats per group were anesthetized with xylazine/ketamine (5.45 mg/36.4 mg/kg im) and then sacrificed by decapitation on days 0, 7, 14, and 30 for the CD and 0, 14, 30 and 60 for the HFD groups. Blood was collected into heparinized tubes and centrifuged at 4000 × g for 5 min to obtain serum. Livers were removed immediately and homogenized in an ice-cold potassium-phosphate buffer containing 5 mmol/L EDTA (pH 7.4). General and liver parameters are reported in Table 1.
Table 1.
Characteristics and fat infiltration of rats fed a choline-deficient diet or a high fat diet with or without Realsil or a normal choline-supplemented diet (controls)
| Characteristics | Controls | CD | CD + Realsil | HFD | HFD + Realsil | Controls | HFD | HFD + Realsil |
| day 30 | day 30 | day 30 | day 30 | day 30 | day 60 | day 60 | day 30 | |
| Body weight (g) | 305 ± 8 | 311 ± 12 | 304 ± 11 | 302 ± 10 | 308 ± 12 | 355 ± 11 | 364 ± 13 | 351 ± 12 |
| Liver weight (g) | 12.7 ± 0.5 | 19.7 ± 1.3a | 16.7 ± 0.7a | 15.7 ± 1.7a | 13.1 ± 1.3 | 12.7 ± 0.5 | 18.8 ± 1.1a | 15.1 ± 0.6ac |
| ALT (IU/L) | 30 ± 8 | 92 ± 14a | 68 ± 12ac | 42 ± 10a | 31 ± 7c | 30 ± 8 | 69 ± 11a | 52 ± 8ac |
| % area | 3 ± 2 | 311 ± 12 | 304 ± 11 | 16 ± 4 | 8 ± 3 | 4 ± 1 | 20 ± 4 | 15 ± 3ac |
| No. of droplets | 11 ± 5 | 19.7 ± 1.3a | 16.7 ± 0.7a | 570 ± 47a | 356 ± 23 | 12 ± 6 | 389 ± 31a | 519 ± 26ac |
| Mean droplet diameter | 1.1 ± 0.3 | 9.2 ± 1.4a | 6.8 ± 1.2ac | 4.2 ± 1.0a | 3.2 ± 0.7c | 0.9 ± 0.5 | 4.4 ± 1.1a | 3.4 ± 0.9ac |
Data represent percentage of fat in the whole area, number of fat droplets and mean diameter of the droplets. Results are mean ± SD of n = 5 different slides per rat per group at each time point.
P < 0.05 vs control rats;
P < 0.05 vs choline-deficient diet (CD) or high fat diet (HFD) only. ALT: Alanine transaminase.
The protocol was conducted according to the Guide Principles for the care and use of laboratory animals and was approved by the local committee for animal experimentation.
Biochemical determinations
Total glutathione (GSH) determination was performed by precipitating tissue homogenates with 15% sulfosalicylic acid and processing the supernatant by the oxidized glutathione recycling procedure[13]. Protein thiols were measured with an Elmann’s procedure modification[14]. The hepatic and serum levels of thiobarbituric acid malondialdehyde complex (MDA-TBA) were first separated by high-performance liquid chromatography using an analytical column Spherisorb ODS 5 μmol/L (250 mm × 4.6 mm) eluted with 60% (v/v) potassium phosphate buffer 50 mmol/L, pH 6.8 and 40% (v/v) methanol at a flow rate of 1 mL/min. Next, spectrophotometric detection of the MDA-TBA adducts occurred at 532 nm[15]. Glutathione Peroxidase (GPx) activity was assessed by use of the method described by Flohè et al[16]. Calculations were made with 1 unit enzyme considered as the amount consuming 1.15 μmol of nicotinamide adenine dinucleotide phosphate reduced (NADPH) in 1 min at 37 °C (pH 7.0). Thioredoxin levels were quantitated by a standardized ELISA method in serum and liver homogenates. Procedure followed the manufacturer’s instructions (Histo-line Laboratories S.r.l., Milan, Italy); samples were located in micro-wells previously coated with a polyclonal antibody (LF-PA0002) and successively with a primary (LF-MA0077) and a secondary HRP-conjugated anti-mouse antibody (81-6720). After the addition of a chromogenic system, the reaction was stopped and absorbance read at 492 nm. Nitrosothiols were measured according to the method of Cook et al[17] using a mixture of sulfanilamide/N-1-naphty-lethylendiamine dihydrochloride, neutral Griess as reagents. Nitrotyrosine in the serum was quantified using an ELISA Kit containing Streptavidin-peroxidase conjugate which reacts with the substrate tetramethylbenzidine as per manufacturer’s instructions (HyCult Biotechnology b.v., UDEN, The Netherlands) with absorbance read at 450 nm. Keratin 18 fragments (K-18) level was quantified in serum using the ELISA Kit as per manufacturer’s instructions (Cusabio Biotech Co., Ltd): samples were located in microwells previously coated with a human-specific antibody and successively with one substrate and a chromogenic system. The reaction was stopped and absorbance read at 450 nm. Protein concentration was measured by using a Bio-Rad kit for the assay of proteins (Bio-Rad GmbH, Munich, Germany).
Western blotting analysis
The expression of mitochondrial oxidative phosphorylation (OXPHOS) system components was assessed by using an antibody cocktail targeting specific subunits from complexes I (NADH-ubiquinone oxidoreductase), II (succinate dehydrogenase), III [ubiquinone-cytochrome c oxidoreductase (COX)], IV (cytochrome c oxidase), and V (ATP synthase). β-actin was used as a loading control. Frozen tissues were homogenized in a cold Ripa buffer (50 mmol/L Tris-HCl, pH 8.8; 150 mmol/L NaCl; 1% Igepal; 0.5% sodium deoxycholate; 0.1% SDS) supplemented with a protease inhibitor cocktail (Sigma) and ruptured by 30 passages through a needle. Homogenized tissues were centrifuged at 14000 rpm (4 °C, 10 min). The supernatant was collected and kept at -80 °C until used. Protein concentration of each sample was measured using BCA Protein Assay kit according to the manufacturer’s protocol. After denaturation at 100 °C during 5 min in Laemmli buffer (BioRad), proteins (30 μg) were separated by electrophoresis on 10% SDS-polyacrylamide gels (SDS-PAGE) and transferred to a polyvinylidene difluoride (PVDF) membrane. After blocking with 2% of milk in TBST (50 mmol/L Tris-HCl, pH 8; 154 mmol/L NaCl, and 0.1% Tween 20) for 1 h at room temperature, membranes were incubated overnight at 4 °C with antibodies against OXPHOS components (1:2000) and β-actin (1:2500). Membranes were incubated with secondary alkaline phosphatase-conjugated antibodies: goat anti-mouse IgG (1:5000) for 1 h at room temperature. Membranes were incubated with ECF detection system (Amersham, GE Health-Care, Piscataway, NJ) and read with the Versa Doc imaging system (Bio-Rad, Barcelona, Spain).
Histology
Liver specimens were fixed in 10% neutral buffered formalin and paraffin embedded. Five sections of 4 μm thickness from each sample were cut and stained with toluidin blue-periodic acid-schiff stain. Histologic features were examined on five low-power fields per specimen, and a semi-quantitative estimation of the empty vacuoles (fat) was performed by measuring the fat percentage distribution on the surface areas.
Chemicals
Total OXPHOS Rodent Antibody Cocktail was purchased from Mitosciences Inc. (Cambridge, MA). Secondary alkaline phosphatase-conjugated antibodies were purchased from Jackson ImmunoResearch Laboratories, Inc. (Cambridgeshire, United Kingdom). All other chemicals used were of the highest purity available and were purchased from Sigma-Aldrich Chemical Co. (Barcelona, Spain or Milan, Italy).
Statistical analysis
All data are expressed as mean ± SD. The Mann-Whitney rank sum test was used to compare groups. For multiple comparisons, the ANOVA on ranks analysis of variance followed by Dunn’s method was used. The nonparametric Spearman rank order correlation was used to relate biochemical parameters. P < 0.05 defined significance.
RESULTS
Effect of CD and HFD on general parameters and liver histology
Feeding CD or HFD was associated with a progressive and significant increase of liver weight but not of body weight at days 30 and 60, respectively, as compared with control rats (Table 1). In rats with fatty liver, ALT levels increased progressively. At days 30 and 60, both CD and HFD rats showed a two-to-threefold increase.
Rats on CD and HFD showed remarkable liver steatosis (Figure 1 and Table 1). There was a progressive hepatic fat infiltration ranging from large droplets at day 14 to disseminated and confluent vacuoles resulting in microvesicular steatosis at day 30 (CD) and day 60 (HFD). Single cell necrosis was rarely noted. No evidence for inflammation and/or fibrosis was present.
Figure 1.
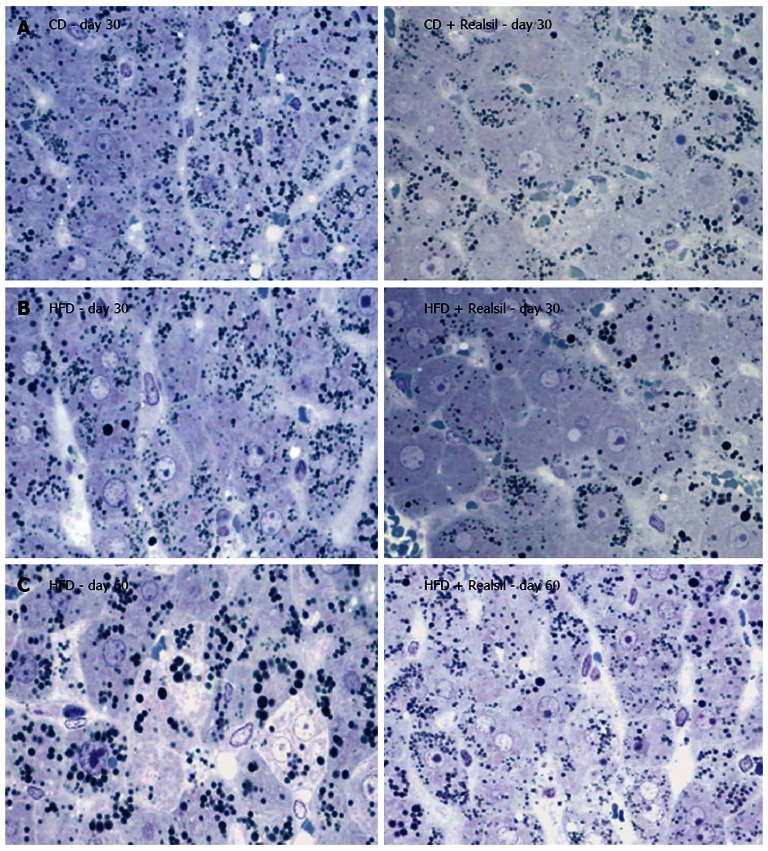
Light micrographs of rat liver stained with toluidin blue-periodic acid-schiff stain. A: Liver from rat fed a choline deficient diet (day 30); B: Liver from rat fed a high fat diet (day 30); C: Liver from rat fed a high fat diet (day 60). Right column without and left column with administration of Realsil (Magnification: 200 ×). CD: Choline deficiency; HFD: High fat diet.
Effect of CD on oxidative and nitrosative stress parameters
In the liver, CD was associated with a higher content of GSH, thioredoxin, and nitrosothiols, while GPx activity was higher at day 7 and then lower at day 30 (Table 2). MDA-TBA levels were five-fold higher from 9.1 ± 1.2 nmol/g to 75.6 ± 5.4 nmol/g at day 30 (Figure 2). Serum MDA-TBA and nitrosothiols levels as well as nitrotyrosine and K-18 (Table 2) were higher in CD rats.
Table 2.
Concentrations of total glutathione, thioredoxin, nitrosothiols, and activity of glutathione peroxidase, malondialdehyde-thiobarbituric acid reactive substances, nitrosothiols, nitrotyrosine, and keratin-18 in the liver of rats
| Day 7 | Day 14 | Day 30 | ||
| Glutathione | S | 34.3 ± 3.1 | 33.2 ± 1.4 | 25.3 ± 1.2a |
| 34.0 ± 1.3 nmol/mg prt | R | 35.1 ± 1.2 | 33.7 ± 1.0 | 30.9 ± 1.1ac |
| Thioredoxin | S | 14.2 ± 1.0a | 16.3 ± 2.4a | 5.2 ± 0.7 |
| 5.6 ± 0.9 nmol/mg prt | R | 11.3 ± 1.0a | 14.4 ± 1.0a | 6.2 ± 0.7c |
| Nitrosothiols | S | 32.9 ± 3.5a | 30.0 ± 2.2a | 13.0 ± 1.2a |
| 16.8 ± 4.7 pmol/mg prt | R | 35.4 ± 2.7a | 27.3 ± 2.2ac | 15.8 ± 1.5ac |
| MDA-TBA | S | 17 ± 4a | 39 ± 8a | 92 ± 10a |
| 12 ± 3 nmol/L | R | 12 ± 3 | 19 ± 4ac | 41 ± 9ac |
| Nitrosothiols | S | 73 ± 15 | 79 ± 12a | 88 ± 10a |
| 62 ± 10 nmol/L | R | 67 ± 10 | 69 ± 10 | 74 ± 11ac |
| Nitrotyrosine | S | 6 ± 2 | 10 ± 3a | 14 ± 3a |
| 6 ± 1 nmol/L | R | 5 ± 1 | 5 ± 2c | 10 ± 2ac |
| K-18 | S | 222 ± 21 | 233 ± 16a | 289 ± 20a |
| 198 ± 20 U/L | R | 210 ± 10 | 213 ± 13c | 240 ± 17ac |
| GPx activity | S | 9.3 ± 1.2a | 21.1 ± 3.7a | 13.7 ± 1.8a |
| 4.5 ± 0.4 nmol NADH/min/mg prt | R | 6.3 ± 0.9a,b | 20.8 ± 1.4a | 16.6 ± 2.6a,b |
The liver of rats fed a choline-deficient (CD) diet without (S)/with (R) Realsil or a normal choline-supplemented diet (controls). Values from controls are reported in the first column under the parameter. Data are mean ± SD of n = 5 rats per group at each time point.
P < 0.05 vs control rats;
P < 0.05 vs rats on CD diet without Realsil at the same time point. GPx: Glutathione peroxidase; MDA-TBA: Malondialdehyde-thiobarbituric acid reactive substances; K-18: Keratin-18; prt: Protein.
Figure 2.
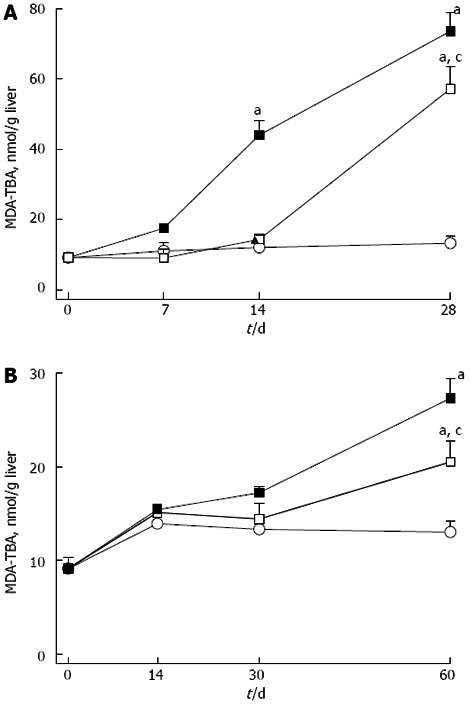
Concentrations of malondialdehyde-thiobarbituric acid reactive substances. A: Liver of rat fed a choline-deficient diet; B: Liver of rat fed a high fat diet. Closed square: Without; open square: With Realsil administration; open circle: Control rats fed a standard chow-diet with adequate content of choline. Data are mean ± SD of n = 5 rats per group at each time point. aP < 0.05 vs control rats; cP < 0.05 vs untreated rats at the same time points. MDA-TBA: Malondialdehyde-thiobarbituric.
Effect of HFD on oxidative and nitrosative stress parameters
In the liver, HFD determined a progressive decrease in the content of GSH, thioredoxin and nitrosothiols (Table 3). GPx activity was initially unchanged, then increased and next decreased (Table 3); MDA-TBA were doubled at day 60 (Figure 2). Serum levels of MDA-TBA, K-18, nitrotyrosine and nitrosothiols were significantly higher in HFD rats (Figure 3).
Table 3.
Concentrations of total glutathione, thioredoxin, nitrosothiols, and activity of glutathione peroxidase in the liver of rat
| Day 14 | Day 30 | Day 60 | ||
| Glutathione | S | 23.7 ± 1.9a | 20.8 ± 2a | 22.4 ± 2.4a |
| 33.4 ± 0.8 nmol/mg prt | R | 22.3 ± 1.8a | 23.9 ± 1.7a | 23 ± 1.6a |
| Thioredoxin | S | 9.2 ± 0.9a | 8.9 ± 0.8a | 4.3 ± 1.0 |
| 5.4 ± 0.5 nmol/mg prt | R | 7.6 ± 0.7ac | 7.8 ± 0.7a | 7.4 ± 1.7ac |
| Nitrosothiols | S | 18.1 ± 1.5 | 19.6 ± 2.5 | 10 ± 2.3a |
| 20.4 ± 3.1 pmol/mg prt | R | 14.4 ± 1.7ac | 16.1 ± 2.3a | 14.3 ± 1.9ac |
| GPx | S | 4.7 ± 0.6a | 6.5 ± 1.3a | 4.6 ± 1.6 |
| 3.9 ± 0.3 nmol NADH/min/mg prt | R | 3.4 ± 0.5c | 4.3 ± 1.0c | 4.1 ± 1.0 |
The liver of rats fed a high fat diet (HFD) diet without (S)/with (R) Realsil or a standard control diet (controls). Values from controls are reported in the first column under the parameter. Data are mean ± SD of n = 5 rats per group at each time point.
P < 0.05 vs control rats;
P < 0.05 vs rats on HFD diet without Realsil at the same time point. GPx: Glutathione peroxidase; NADH: Nicotinamide adenine dinucleotide; prt: Protein.
Figure 3.
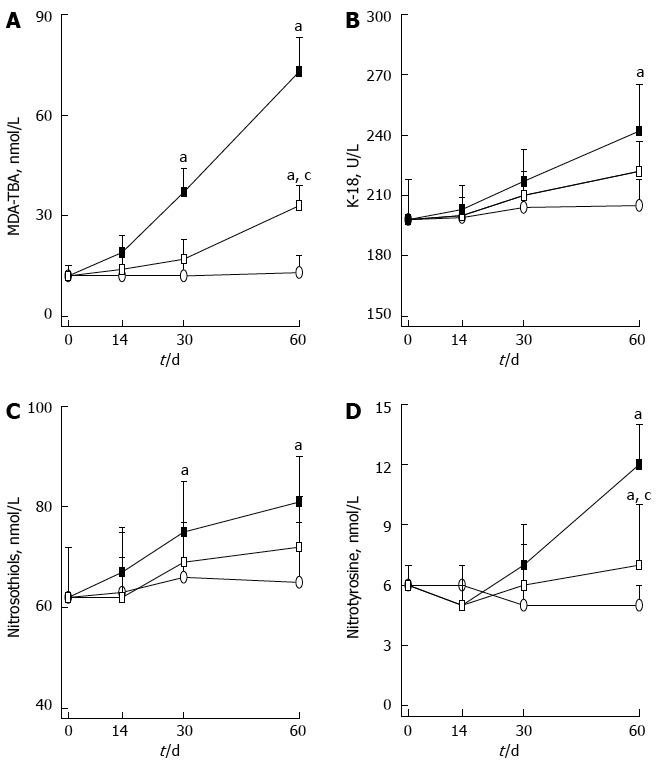
Plasma concentration of redox active and nitrogen regulating molecules. A: Malondialdehyde-thiobarbituric acid reactive substances (MDA-TBA); B: Keratine 18 (K-18); C: Nitrosothiols; D: Nitrotyrosine. Rats were fed a high fat diet without (closed square) or with (open square) Realsil administration; control rats fed a standard chow-diet (open circle). Data are mean ± SD of n = 5 rats per group at each time point. aP < 0.05 vs control rats; cP < 0.05 vs untreated rats at the same time points.
Effect of HFD on mitochondrial oxidative phosphorylation complexes
Samples from liver, heart and skeletal muscle were compared regarding the content in subunits of the mitochondrial respiratory chain. Western blotting yielded bands to all OXPHOS subunits studied, although the band corresponding to complex III was very difficult to detect in the three tissues and hence was not quantified by densitometry. Band density for each protein was normalized for the corresponding β-actin band.
In the liver (Figure 4), the HFD caused an increase in the amount of the Complex I subunit NDUFB8 (at day 60) and Complex IV subunit I, COX I (at day 30) and a decrease of Complex II subunit 30kDa, CII-30 (days 14 and 60) and ATP synthase subunit α, CV-α (day 14). Minor changes in the heart and skeletal muscle proteins were observed (data not shown).
Figure 4.
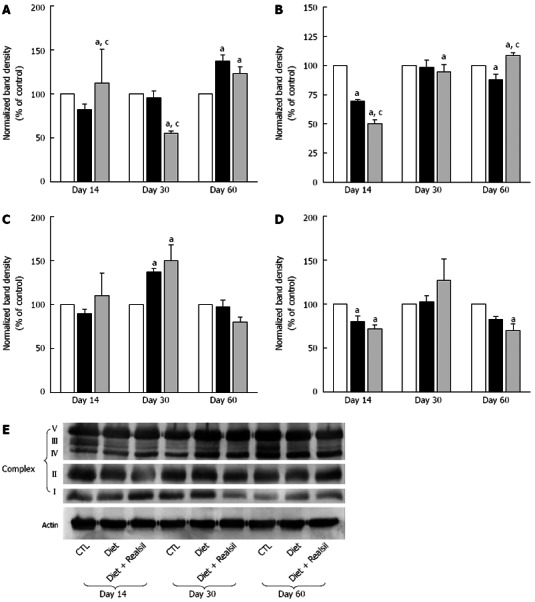
Western blotting analysis of the components of liver mitochondrial oxidative phosphorylation system in rats receiving high fat diet with (grey bar) or without (black bar) Realsil and in control (white bar) rats fed a standard chow-diet. Band density for the target protein normalized for the corresponding β-actin for A: Respiratory complex I; B: Respiratory complex II; C: Respiratory complex IV; D: Respiratory complex V; E: Picture representative of Western blotting analysis. Data are means ± SE of n = 3-4 independent experiments. aP < 0.05 vs control rats; cP < 0.05 vs untreated rats at the same time points.
Effect of Realsil on general parameters, liver histology, stress markers, and mitochondrial proteins
Realsil administration was associated with a lower increase of liver weight and a less pronounced increase of serum transaminase levels in rats (Table 1). As shown (Figure 1 and Table 2), Realsil resulted in a lower extent of fat infiltration in both CD and HFD rats. In CD rats, Realsil determined an improvement in the number and diameter of fat droplets. In HFD rats, Realsil was associated with a lower diameter of fat droplets; the number of droplets per histological section was lower at day 30 but higher at day 60, as a likely consequence of a less confluent phenomenon.
Realsil was protective against CD and HFD induced oxidative and nitrosative changes both in the liver and in plasma. The improvement in such parameters was more evident for CD rats. In particular, the decrease of liver GSH and nitrosothiols was kept to 50% compared with untreated rats, while the changes in MDA-TBA, thioredoxin, and GPx were less marked although significant (Table 2). The same parameters were less affected by Realsil in HFD rats (Table 3 and Figure 3). In fact, the hepatic concentrations of GSH, MDA-TBA, GPx, and nitrosothiols did not differ significantly compared to untreated HFD rats. Only thioredoxin levels were maintained to a higher level by Realsil administration. In these rats, Realsil halved the variations of serum MDA-TBA and nitrotyrosine levels, while it was less effective on K-18 (222 ± 15 IU/L vs 242 ± 23 IU/L at day 60 in treated and untreated HFD rats, respectively) and nitrosothiols (72 ± 10 nmol/L vs 81 ± 9 nmol/L at day 60 in treated and untreated HFD rats, respectively) levels.
In HFD animals receiving also Realsil, several changes occurred in liver mitochondria (Figure 4). At day 14, administration of Realsil resulted in a significant increase in the protein levels of NDUFB8 subunit, which were slightly decreased (although not statistically significant) by the diet alone. On the other hand, Realsil in HFD animals further decreased the amount of CII-30 (day 14), NDUFB8 (day 30), and COX I (day 60). At day 60, Realsil had protective effect on the Complex II subunit CII-30 by increasing the amount of the protein that was decreased by the diet alone. Interestingly, in the muscle, at day 60 Realsil reversed the effect induced by HFD in CII-30 subunit and in COX I subunit and, at day 30, decreased CV-α subunit protein amount.
DISCUSSION
Obesity is a health problem in developed countries[18-21] and is related to insulin resistance, dyslipidemia, type 2 diabetes, hypertension, and liver steatosis (NAFLD)[22]. Simple steatosis is considered a benign condition with low risk of evolution, while its inflammatory form (NASH) is considered a risk factor for liver cirrhosis.
Mechanisms governing hepatocellular fat deposition involve metabolic pathways partly depending on the up-regulation of peroxisome proliferator-activated receptor and on the consequent activation of adipocyte differentiation programs[23]. The relationship between intracellular metabolic processes and systemic changes occurring in patients with fatty liver are still under debate. Also, the effect of modulating compounds and antioxidant molecules on such lipid effectors and on NO metabolic changes has been poorly investigated. Mitochondria, coordinators of energy metabolism, are actively involved in such processes[18,21]. In NASH patients, mitochondria show morphological alterations and functional impairment[7,24]. Ultrastructural modifications were also observed in rats fed a steatogenic diet (HFD)[25]. Mitochondria are the most relevant source of ROS in most cells and especially in fatty hepatocytes[26,27]. ROS alter the activity of JNK enzymes, disturb insulin signaling, and enhance potassium transport across the inner mitochondrial membrane[28] leading to mitochondrial uncoupling and triggering as well adaptive response[29-31].
In a recent study[3], we observed that hepatocytes react to fat deposition by a very early increase of GSH and thioredoxin both in the cytosol and in the mitochondria to likely prevent lipid and protein oxidation. Prolongation of steatogenic diet, however, led to major mitochondrial redox changes, i.e., increased MDA-TBA, progressive decrease of GSH and thioredoxin, and increase of mixed disulfides, nitrates, and nitrosothiols, all consistent with both oxidative damage and increased NO synthesis. In fatty livers, NO synthase (iNOS) is induced by enhanced inflow of gut-derived toxins[32] and tumor necrosis factor-α expression[33] and generates hyper reactive species such as NO with accumulation of nitrotyrosine[33]. This likely reflects an increased peroxynitrite formation and suggests potential NO participation to liver injury[34]. Thioredoxin, a redox-active protein induced by oxidative stress[35], is actively involved in NO regulation through nitrosothiols cleavage[36,37]. Nitrosothiols, formed by conjugation of NO with free thiols, oppose peroxynitrite formation and act as intracellular messenger controlling cellular and mitochondrial functions[38,39].
In the fatty liver, intracellular redox changes and protein nitrosation may represent a major factor stimulating the progression from simple steatosis to NASH[40]. Also, a critical role for such hepatic variations in the pathogenesis of systemic chronic inflammatory conditions has been recently proposed[41,42]. In our study, we show that liver steatosis is associated with high levels of circulating NO derivatives (nitrosothiols and nitrotyrosine) and with high levels of K-18, the major keratin expressed in the liver and one of the most prominent substrates of caspases during hepatocytes apoptotic death[43]. These alterations were closely dependent on the changes occurring in the liver in which GSH content and thioredoxin activity declined with ongoing steatosis. Also, the close relation observed between the circulatory NO derivatives and K-18 levels clearly links NO with the hepatic inflammatory processes occurring under marked liver steatosis. Previous studies have demonstrated elevation of these molecules in NAFLD[44] as a consequence of the increased apoptotic rate due to hepatic inflammation[45].
In the present work, the HFD promoted alterations in mitochondrial complexes, although not deep enough to result in bioenergetic changes, as reported for other diet models including CD[46,47]. Interestingly, the limited array of alterations observed in the heart and skeletal muscle points to the conclusion that this model selectively affects the liver.
In this scenario, the administration of antioxidant molecules able to contrast oxidative and nitrosative phenomena improved most of the investigated stress parameters both in the liver and in serum. In fact, in our study the administration of Realsil was effective in reducing the extent of fatty infiltration of the liver and in modulating the changes in mitochondrial function and oxidative and nitrosative stress both in the liver and in the systemic circulation. This would suggest that Realsil is effective in contrasting the metabolic alterations resulting in excess fat deposition and at the same time in counteracting the increased formation of ROS and NO species.
Silybin, the major constituent of milk thistle extract, affords hepatoprotection in vitro and in vivo[48,49] by inhibiting the production of pro-inflammatory and pro-fibrogenic factors[12,50]. The conjugation with phospholipids greatly increases its intestinal absorption and the systemic bioavailability[51]. However, the mechanisms of the hepatocyte protection have not been completely defined. Some hypotheses point to a potential antioxidant cytoprotective effect of Realsil by including the modulation of protein oxidation/denitrosation and the maintenance of membrane lipid composition and function[52]. To demonstrate these effects, we investigated the protective effects of Realsil on oxidative/nitrosative changes both in the liver and in circulation.
Indeed, although the subtle mechanisms regulating the protein nitrosation/denitrosation process have not been completely identified, it has been shown that thioredoxin-deficient cells denitrosate nitrosothiols less efficiently[37] and that nitrosative stress is critically important in promoting S-nitrosylation and nitration of various mitochondrial proteins, leading to mitochondrial dysfunction, decreased energy supply, and increased hepatic injury[53]. With ongoing steatosis, the decrement in hepatic thioredoxin and GSH levels we observed may be due also to a down-regulation process associated with excess deposition of fat and toxic molecules and may indirectly contribute to the progressive appearance of other oxidative changes.
In our study, Realsil was able to counteract most of the oxidative biochemical alterations during the early phases of steatosis while it resulted less effective later (30 and 60 d) when the extent of fat infiltration was massive. The protection promoted by Realsil is certainly exerted at different levels and particularly at mitochondrial level[54]. The changes (both morphological and biochemical) observed in the liver of rats receiving Realsil were also evident at systemic level and this more explicitly relates systemic with hepatic changes in animals with fatty liver.
COMMENTS
Background
Fatty degeneration represents a leading factor of hepatocyte dysfunction and is associated with excess delivery of stress molecules thus rendering the liver a major source of systemic alterations in patients with metabolic syndrome. Adaptive metabolic mechanisms have been described during the early phase of fatty infiltration including expression of intracellular sensors and signaling molecules for lipid metabolism and oxidative stress pathways. Vitamin E efforts a remarkable protection against lipid oxidation and, if vehicoled within a phospholipids complex together with silybin, a silybum marianum extract, protects against pro-fibrotic oxidative injury.
Research frontiers
The threshold above which fat infiltration becomes dangerous is not clear, so far. Several observations suggest that ongoing fatty degeneration indeed exposes hepatocytes to higher risk of oxidative damages.
Innovations and breakthroughs
Little is known about the effectiveness of vitamin E-silybin-phospholipid complex in blocking or modulating reactive oxygen species/NO production and their oxidant effects. This study also gives answer to the question whether administration of an antioxidant complex is able to break off the intracellular mechanisms leading to a progressive accumulation of neutral lipids in fatty hepatocytes and mitochondrial dysfunction.
Applications
Realsil was able to counteract most of the oxidative hepatic changes during the early phases of steatosis while it resulted less effective later when the extent of fat infiltration was massive. The changes observed in the rat fatty liver were also evident at systemic level pointing to a relationship between systemic and hepatic alterations.
Terminology
Liver steatosis occurs when the amount of neutral fat exceeds 5% of organ weight. Realsil is a compound constituted by vitamin E and silybin complexed with phospholipids. Silybin is a silybum marianum extract; it is known to protect against pro-fibrotic oxidative injury.
Peer review
In this descriptive study, the authors show that Realsil is able to reduce liver injury in two different animal models of steatosis. Realsil seems to protect by decreasing toxic free radicals species as highly delivered by fatty hepatocytes.
Footnotes
Supported by Grants from MIUR (Ministero Università e Ricerca Scientifica COFIN2006) and “Fondi Ateneo Ricerca Scientifica 2005/2006” from the University of Bari, Italy
P- Reviewers Cai SY, Wisse E S- Editor Gou SX L- Editor A E- Editor Zhang DN
References
- 1.Humphreys D. Careers extraordinary: “what’s on the programme today, sister”? Nurs Times. 1972;68:308–310. [PubMed] [Google Scholar]
- 2.Portincasa P, Grattagliano I, Palmieri VO, Palasciano G. Nonalcoholic steatohepatitis: recent advances from experimental models to clinical management. Clin Biochem. 2005;38:203–217. doi: 10.1016/j.clinbiochem.2004.10.014. [DOI] [PubMed] [Google Scholar]
- 3.Grattagliano I, Caraceni P, Calamita G, Ferri D, Gargano I, Palasciano G, Portincasa P. Severe liver steatosis correlates with nitrosative and oxidative stress in rats. Eur J Clin Invest. 2008;38:523–530. doi: 10.1111/j.1365-2362.2008.01963.x. [DOI] [PubMed] [Google Scholar]
- 4.Palasciano G, Moschetta A, Palmieri VO, Grattagliano I, Iacobellis G, Portincasa P. Non-alcoholic fatty liver disease in the metabolic syndrome. Curr Pharm Des. 2007;13:2193–2198. doi: 10.2174/138161207781039652. [DOI] [PubMed] [Google Scholar]
- 5.Yang S, Zhu H, Li Y, Lin H, Gabrielson K, Trush MA, Diehl AM. Mitochondrial adaptations to obesity-related oxidant stress. Arch Biochem Biophys. 2000;378:259–268. doi: 10.1006/abbi.2000.1829. [DOI] [PubMed] [Google Scholar]
- 6.Merriman RB, Aouizerat BE, Bass NM. Genetic influences in nonalcoholic fatty liver disease. J Clin Gastroenterol. 2006;40 Suppl 1:S30–S33. doi: 10.1097/01.mcg.0000168643.16074.19. [DOI] [PubMed] [Google Scholar]
- 7.Sanyal AJ, Campbell-Sargent C, Mirshahi F, Rizzo WB, Contos MJ, Sterling RK, Luketic VA, Shiffman ML, Clore JN. Nonalcoholic steatohepatitis: association of insulin resistance and mitochondrial abnormalities. Gastroenterology. 2001;120:1183–1192. doi: 10.1053/gast.2001.23256. [DOI] [PubMed] [Google Scholar]
- 8.Shteyer E, Liao Y, Muglia LJ, Hruz PW, Rudnick DA. Disruption of hepatic adipogenesis is associated with impaired liver regeneration in mice. Hepatology. 2004;40:1322–1332. doi: 10.1002/hep.20462. [DOI] [PubMed] [Google Scholar]
- 9.Yamaguchi K, Yang L, McCall S, Huang J, Yu XX, Pandey SK, Bhanot S, Monia BP, Li YX, Diehl AM. Inhibiting triglyceride synthesis improves hepatic steatosis but exacerbates liver damage and fibrosis in obese mice with nonalcoholic steatohepatitis. Hepatology. 2007;45:1366–1374. doi: 10.1002/hep.21655. [DOI] [PubMed] [Google Scholar]
- 10.Trevisani F, Colantoni A, Caraceni P, Van Thiel DH. The use of donor fatty liver for liver transplantation: a challenge or a quagmire? J Hepatol. 1996;24:114–121. doi: 10.1016/s0168-8278(96)80195-4. [DOI] [PubMed] [Google Scholar]
- 11.Hu JJ, Roush GC, Berwick M, Dubin N, Mahabir S, Chandiramani M, Boorstein R. Effects of dietary supplementation of alpha-tocopherol on plasma glutathione and DNA repair activities. Cancer Epidemiol Biomarkers Prev. 1996;5:263–270. [PubMed] [Google Scholar]
- 12.Trappoliere M, Caligiuri A, Schmid M, Bertolani C, Failli P, Vizzutti F, Novo E, di Manzano C, Marra F, Loguercio C, et al. Silybin, a component of sylimarin, exerts anti-inflammatory and anti-fibrogenic effects on human hepatic stellate cells. J Hepatol. 2009;50:1102–1111. doi: 10.1016/j.jhep.2009.02.023. [DOI] [PubMed] [Google Scholar]
- 13.Tietze F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: applications to mammalian blood and other tissues. Anal Biochem. 1969;27:502–522. doi: 10.1016/0003-2697(69)90064-5. [DOI] [PubMed] [Google Scholar]
- 14.Grattagliano I, Russmann S, Palmieri VO, Portincasa P, Palasciano G, Lauterburg BH. Glutathione peroxidase, thioredoxin, and membrane protein changes in erythrocytes predict ribavirin-induced anemia. Clin Pharmacol Ther. 2005;78:422–432. doi: 10.1016/j.clpt.2005.07.002. [DOI] [PubMed] [Google Scholar]
- 15.Slater TF, Sawyer BC. The stimulatory effects of carbon tetrachloride on peroxidative reactions in rat liver fractions in vitro. Inhibitory effects of free-radical scavengers and other agents. Biochem J. 1971;123:823–828. doi: 10.1042/bj1230823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Flohé L, Günzler WA. Assays of glutathione peroxidase. Methods Enzymol. 1984;105:114–121. doi: 10.1016/s0076-6879(84)05015-1. [DOI] [PubMed] [Google Scholar]
- 17.Cook JA, Kim SY, Teague D, Krishna MC, Pacelli R, Mitchell JB, Vodovotz Y, Nims RW, Christodoulou D, Miles AM, et al. Convenient colorimetric and fluorometric assays for S-nitrosothiols. Anal Biochem. 1996;238:150–158. doi: 10.1006/abio.1996.0268. [DOI] [PubMed] [Google Scholar]
- 18.Begriche K, Igoudjil A, Pessayre D, Fromenty B. Mitochondrial dysfunction in NASH: causes, consequences and possible means to prevent it. Mitochondrion. 2006;6:1–28. doi: 10.1016/j.mito.2005.10.004. [DOI] [PubMed] [Google Scholar]
- 19.Bray GA, Nielsen SJ, Popkin BM. Consumption of high-fructose corn syrup in beverages may play a role in the epidemic of obesity. Am J Clin Nutr. 2004;79:537–543. doi: 10.1093/ajcn/79.4.537. [DOI] [PubMed] [Google Scholar]
- 20.Jew S, AbuMweis SS, Jones PJ. Evolution of the human diet: linking our ancestral diet to modern functional foods as a means of chronic disease prevention. J Med Food. 2009;12:925–934. doi: 10.1089/jmf.2008.0268. [DOI] [PubMed] [Google Scholar]
- 21.Nicolson GL. Metabolic syndrome and mitochondrial function: molecular replacement and antioxidant supplements to prevent membrane peroxidation and restore mitochondrial function. J Cell Biochem. 2007;100:1352–1369. doi: 10.1002/jcb.21247. [DOI] [PubMed] [Google Scholar]
- 22.Federspil G, Nisoli E, Vettor R. A critical reflection on the definition of metabolic syndrome. Pharmacol Res. 2006;53:449–456. doi: 10.1016/j.phrs.2006.03.008. [DOI] [PubMed] [Google Scholar]
- 23.Yu S, Matsusue K, Kashireddy P, Cao WQ, Yeldandi V, Yeldandi AV, Rao MS, Gonzalez FJ, Reddy JK. Adipocyte-specific gene expression and adipogenic steatosis in the mouse liver due to peroxisome proliferator-activated receptor gamma1 (PPARgamma1) overexpression. J Biol Chem. 2003;278:498–505. doi: 10.1074/jbc.M210062200. [DOI] [PubMed] [Google Scholar]
- 24.Le TH, Caldwell SH, Redick JA, Sheppard BL, Davis CA, Arseneau KO, Iezzoni JC, Hespenheide EE, Al-Osaimi A, Peterson TC. The zonal distribution of megamitochondria with crystalline inclusions in nonalcoholic steatohepatitis. Hepatology. 2004;39:1423–1429. doi: 10.1002/hep.20202. [DOI] [PubMed] [Google Scholar]
- 25.Altunkaynak BZ, Ozbek E. Overweight and structural alterations of the liver in female rats fed a high-fat diet: a stereological and histological study. Turk J Gastroenterol. 2009;20:93–103. [PubMed] [Google Scholar]
- 26.Kowaltowski AJ, de Souza-Pinto NC, Castilho RF, Vercesi AE. Mitochondria and reactive oxygen species. Free Radic Biol Med. 2009;47:333–343. doi: 10.1016/j.freeradbiomed.2009.05.004. [DOI] [PubMed] [Google Scholar]
- 27.Tahara EB, Navarete FD, Kowaltowski AJ. Tissue-, substrate-, and site-specific characteristics of mitochondrial reactive oxygen species generation. Free Radic Biol Med. 2009;46:1283–1297. doi: 10.1016/j.freeradbiomed.2009.02.008. [DOI] [PubMed] [Google Scholar]
- 28.Nishikawa T, Kukidome D, Sonoda K, Fujisawa K, Matsuhisa T, Motoshima H, Matsumura T, Araki E. Impact of mitochondrial ROS production in the pathogenesis of insulin resistance. Diabetes Res Clin Pract. 2007;77 Suppl 1:S161–S164. doi: 10.1016/j.diabres.2007.01.071. [DOI] [PubMed] [Google Scholar]
- 29.Facundo HT, Fornazari M, Kowaltowski AJ. Tissue protection mediated by mitochondrial K+ channels. Biochim Biophys Acta. 2006;1762:202–212. doi: 10.1016/j.bbadis.2005.06.003. [DOI] [PubMed] [Google Scholar]
- 30.Ferranti R, da Silva MM, Kowaltowski AJ. Mitochondrial ATP-sensitive K+ channel opening decreases reactive oxygen species generation. FEBS Lett. 2003;536:51–55. doi: 10.1016/s0014-5793(03)00007-3. [DOI] [PubMed] [Google Scholar]
- 31.Fornazari M, de Paula JG, Castilho RF, Kowaltowski AJ. Redox properties of the adenoside triphosphate-sensitive K+ channel in brain mitochondria. J Neurosci Res. 2008;86:1548–1556. doi: 10.1002/jnr.21614. [DOI] [PubMed] [Google Scholar]
- 32.Yang S, Lin H, Diehl AM. Fatty liver vulnerability to endotoxin-induced damage despite NF-kappaB induction and inhibited caspase 3 activation. Am J Physiol Gastrointest Liver Physiol. 2001;281:G382–G392. doi: 10.1152/ajpgi.2001.281.2.G382. [DOI] [PubMed] [Google Scholar]
- 33.García-Ruiz I, Rodríguez-Juan C, Díaz-Sanjuan T, del Hoyo P, Colina F, Muñoz-Yagüe T, Solís-Herruzo JA. Uric acid and anti-TNF antibody improve mitochondrial dysfunction in ob/ob mice. Hepatology. 2006;44:581–591. doi: 10.1002/hep.21313. [DOI] [PubMed] [Google Scholar]
- 34.Ottesen LH, Harry D, Frost M, Davies S, Khan K, Halliwell B, Moore K. Increased formation of S-nitrothiols and nitrotyrosine in cirrhotic rats during endotoxemia. Free Radic Biol Med. 2001;31:790–798. doi: 10.1016/s0891-5849(01)00647-5. [DOI] [PubMed] [Google Scholar]
- 35.Nakamura H, Nakamura K, Yodoi J. Redox regulation of cellular activation. Annu Rev Immunol. 1997;15:351–369. doi: 10.1146/annurev.immunol.15.1.351. [DOI] [PubMed] [Google Scholar]
- 36.Nikitovic D, Holmgren A. S-nitrosoglutathione is cleaved by the thioredoxin system with liberation of glutathione and redox regulating nitric oxide. J Biol Chem. 1996;271:19180–19185. doi: 10.1074/jbc.271.32.19180. [DOI] [PubMed] [Google Scholar]
- 37.Stoyanovsky DA, Tyurina YY, Tyurin VA, Anand D, Mandavia DN, Gius D, Ivanova J, Pitt B, Billiar TR, Kagan VE. Thioredoxin and lipoic acid catalyze the denitrosation of low molecular weight and protein S-nitrosothiols. J Am Chem Soc. 2005;127:15815–15823. doi: 10.1021/ja0529135. [DOI] [PubMed] [Google Scholar]
- 38.Arnelle DR, Stamler JS. NO+, NO, and NO- donation by S-nitrosothiols: implications for regulation of physiological functions by S-nitrosylation and acceleration of disulfide formation. Arch Biochem Biophys. 1995;318:279–285. doi: 10.1006/abbi.1995.1231. [DOI] [PubMed] [Google Scholar]
- 39.Grattagliano I, Portincasa P, Palmieri VO, Palasciano G. Mutual changes of thioredoxin and nitrosothiols during biliary cirrhosis: results from humans and cholestatic rats. Hepatology. 2007;45:331–339. doi: 10.1002/hep.21519. [DOI] [PubMed] [Google Scholar]
- 40.Klatt P, Lamas S. Regulation of protein function by S-glutathiolation in response to oxidative and nitrosative stress. Eur J Biochem. 2000;267:4928–4944. doi: 10.1046/j.1432-1327.2000.01601.x. [DOI] [PubMed] [Google Scholar]
- 41.Dunn W, Xu R, Wingard DL, Rogers C, Angulo P, Younossi ZM, Schwimmer JB. Suspected nonalcoholic fatty liver disease and mortality risk in a population-based cohort study. Am J Gastroenterol. 2008;103:2263–2271. doi: 10.1111/j.1572-0241.2008.02034.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Grattagliano I, Palmieri VO, Portincasa P, Moschetta A, Palasciano G. Oxidative stress-induced risk factors associated with the metabolic syndrome: a unifying hypothesis. J Nutr Biochem. 2008;19:491–504. doi: 10.1016/j.jnutbio.2007.06.011. [DOI] [PubMed] [Google Scholar]
- 43.Linder S. Cytokeratin markers come of age. Tumour Biol. 2007;28:189–195. doi: 10.1159/000107582. [DOI] [PubMed] [Google Scholar]
- 44.Wieckowska A, Zein NN, Yerian LM, Lopez AR, McCullough AJ, Feldstein AE. In vivo assessment of liver cell apoptosis as a novel biomarker of disease severity in nonalcoholic fatty liver disease. Hepatology. 2006;44:27–33. doi: 10.1002/hep.21223. [DOI] [PubMed] [Google Scholar]
- 45.Yilmaz Y, Dolar E, Ulukaya E, Akgoz S, Keskin M, Kiyici M, Aker S, Yilmaztepe A, Gurel S, Gulten M, et al. Soluble forms of extracellular cytokeratin 18 may differentiate simple steatosis from nonalcoholic steatohepatitis. World J Gastroenterol. 2007;13:837–844. doi: 10.3748/wjg.v13.i6.837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Caraceni P, Bianchi C, Domenicali M, Maria Pertosa A, Maiolini E, Parenti Castelli G, Nardo B, Trevisani F, Lenaz G, Bernardi M. Impairment of mitochondrial oxidative phosphorylation in rat fatty liver exposed to preservation-reperfusion injury. J Hepatol. 2004;41:82–88. doi: 10.1016/j.jhep.2004.03.022. [DOI] [PubMed] [Google Scholar]
- 47.Cardoso AR, Cabral-Costa JV, Kowaltowski AJ. Effects of a high fat diet on liver mitochondria: increased ATP-sensitive K+ channel activity and reactive oxygen species generation. J Bioenerg Biomembr. 2010;42:245–253. doi: 10.1007/s10863-010-9284-9. [DOI] [PubMed] [Google Scholar]
- 48.Di Sario A, Bendia E, Taffetani S, Omenetti A, Candelaresi C, Marzioni M, De Minicis S, Benedetti A. Hepatoprotective and antifibrotic effect of a new silybin-phosphatidylcholine-Vitamin E complex in rats. Dig Liver Dis. 2005;37:869–876. doi: 10.1016/j.dld.2005.05.011. [DOI] [PubMed] [Google Scholar]
- 49.Wellington K, Jarvis B. Silymarin: a review of its clinical properties in the management of hepatic disorders. BioDrugs. 2001;15:465–489. doi: 10.2165/00063030-200115070-00005. [DOI] [PubMed] [Google Scholar]
- 50.Pietrangelo A, Montosi G, Garuti C, Contri M, Giovannini F, Ceccarelli D, Masini A. Iron-induced oxidant stress in nonparenchymal liver cells: mitochondrial derangement and fibrosis in acutely iron-dosed gerbils and its prevention by silybin. J Bioenerg Biomembr. 2002;34:67–79. doi: 10.1023/a:1013874804911. [DOI] [PubMed] [Google Scholar]
- 51.Morazzoni P, Montalbetti A, Malandrino S, Pifferi G. Comparative pharmacokinetics of silipide and silymarin in rats. Eur J Drug Metab Pharmacokinet. 1993;18:289–297. doi: 10.1007/BF03188811. [DOI] [PubMed] [Google Scholar]
- 52.Pietrangelo A, Borella F, Casalgrandi G, Montosi G, Ceccarelli D, Gallesi D, Giovannini F, Gasparetto A, Masini A. Antioxidant activity of silybin in vivo during long-term iron overload in rats. Gastroenterology. 1995;109:1941–1949. doi: 10.1016/0016-5085(95)90762-9. [DOI] [PubMed] [Google Scholar]
- 53.Moon KH, Hood BL, Mukhopadhyay P, Rajesh M, Abdelmegeed MA, Kwon YI, Conrads TP, Veenstra TD, Song BJ, Pacher P. Oxidative inactivation of key mitochondrial proteins leads to dysfunction and injury in hepatic ischemia reperfusion. Gastroenterology. 2008;135:1344–1357. doi: 10.1053/j.gastro.2008.06.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Serviddio G, Bellanti F, Giudetti AM, Gnoni GV, Petrella A, Tamborra R, Romano AD, Rollo T, Vendemiale G, Altomare E. A silybin-phospholipid complex prevents mitochondrial dysfunction in a rodent model of nonalcoholic steatohepatitis. J Pharmacol Exp Ther. 2010;332:922–932. doi: 10.1124/jpet.109.161612. [DOI] [PubMed] [Google Scholar]