

Abstract
Rapid technological development is inevitably associated with many environmental problems which primarily include pollution of soil, water and air. In many cases, the presence of contamination is difficult to assess. It is even more difficult to evaluate its potential danger to the environment and humans. Despite the existence of several whole organism-based and cell-based models of sensing pollution and evaluation of toxicity and mutagenicity, there is no ideal system that allows one to make a quick and cheap assessment. In this respect, transgenic organisms that can be intentionally altered to be more sensitive to particular pollutants are especially promising. Transgenic plants represent an ideal system, since they can be grown at the site of pollution or potentially dangerous sites. Plants are ethically more acceptable and esthetically more appealing than animals as sensors of environmental pollution. In this review, we will discuss various transgenic plant-based models that have been successfully used for biomonitoring genotoxic pollutants. We will also discuss the benefits and potential drawbacks of these systems and describe some novel ideas for the future generation of efficient transgenic phytosensors.
Keywords: Transgenic phytosensor, plant biosensor, Arabidopsis thaliana, Nicotiana tabacum, environmental pollution, heavy metal, radioactive contamination
Introduction
Every organism on our planet experiences constant exposure to natural forces. Environmental factors such as fluctuations in temperature and light intensity as well as water availability have a substantial impact on the genetic makeup of organisms [1]. Additional impact on all living organisms comes from humans. Our dramatic economical and technological progress is associated with heavy metal pollution, radiation exposure and influences of such stressors as xenobiotic chemicals or nutrients. A majority of present problems in the environment are due to careless industrial development, mistakes and malfunctions of man-made equipment. Most forms of environmental pollution are chemical in nature, although some of them are physical, like ultraviolet radiation, and others, like ionizing radiation, are both chemical and physical. In this review, we did not intend to cover the whole range of impacts that a broad variety of natural and man-made contaminants have on biological objects. For information on these topics we refer our readers to the comprehensive reviews explaining the influence of radiation (ultraviolet and ionizing), heavy metal salts, and organic chemicals (pesticides, herbicides and insecticides) on living organisms [2-8].
Here, we will describe biomonitoring of environmental genotoxic pollutants using transgenic phytosensors. We will briefly describe the various organisms used as biosensors, and will discuss plant models that have recently been used for monitoring. We will also propose a few models that potentially can be used.
Various biosensors
A majority of pollutants are difficult to detect without specialized equipment. Thus, using wide-area monitoring of the environment for the presence of toxic and mutagenic substances is needed. A variety of chemical and physical methods used for detecting the presence of various pollutants can define an exact amount of a certain chemical. Unfortunately, no monitoring equipment can estimate potential toxicity and/or mutagenicity of toxicants. The release of new chemicals into the environment makes the monitoring process even more problematic. Therefore, the analysis of environmental pollution using biosensors (living organisms which are able to assess the quality of soil, water or air) is absolutely essential. A number of different biosensors were successfully used in the past including bacterial, animal (mammals, fish, and worms) and plants.
Bacterial assays for biomonitoring
Bacterial bioassays were among the first laboratory tests used for biomonitoring. The original Ames test based on Salmonella typhimurium and a variety of its modifications proved to be valuable for assessing mutagenicity [9]. The gain and loss of function of Escherichia coli- based test systems contributed to further elaborating on the Ames test [10-11]. Despite an attempt at making bacterial tests suitable for the analysis of mutagenicity of various compounds for eukaryotes, they appeared to be insensitive to many pollutants. Exposure of higher eukaryotes to environmental mutagens depends on bioavailability of the compound and on the duration of contact between an organism and contamination rather than on the amount of mutagens present. Hence, new assays using eukaryotes should be more physiologically relevant and more precise.
Animal-based bioassays
Animal-based assays include those that use worms, insects, mammals and fish. Some of the general assays provide estimates of mutagen exposure and involve DNA damage analysis. The alkaline single cell-gel electrophoresis assay (a Comet assay) [12] and the sister chromatid exchange assay [13] are good examples.
Whole organism models include the transgenic zebrafish assay, which uses a non-active lacI transgene as a target gene [14], the transgenic mouse mutation and recombination-based assay [15-17], which uses restoration of a lacZ gene in germ cells, the transgenic “fluorescent yellow direct repeat” (FYDR) mouse model, based on restoration of gene coding for the enhanced yellow fluorescent protein [18-19], and a Drosophila melanogaster system, which is based on reversion of the recessive eye color mutation [20].
Unfortunately, these models do not allow the study of impacts of complex patterns of soil pollution, because many living organisms are not suitable for biomonitoring, especially for biomonitoring in the field [21]. Ideally, to be sensitive field biomonitors, test-organisms should absorb and integrate doses of toxicants from polluted air, water or/and soil. Therefore, the use of animal-based biomonitoring models is problematic due to the non-sessile lifestyle of animals.
Plants as biosensors
Plants as biomonitors can be an appealing alternative to animals. Plants are energy producers and a food source for many organisms. Therefore, their role in transferring contaminants to higher trophic levels is difficult to overestimate [22-23]. Plants are the main components of a healthy environment, since they produce oxygen and organic carbon. They are also the basic components of agriculture and forestry. These are just some of the reasons why many plant species are used as indicators of adverse environmental factors.
The use of plants as biosensors has a long history. For decades, they were used as a part of ecological risk assessment of agricultural and industrial chemicals, solid wastes, food additives, and chemically and radioactively polluted soil and water. Phytoremediation or cleaning the environment using plants is yet another area where plants can be used to great advantage.
At the same time, it should be admitted that the use of plants as environmental biomonitors has some drawbacks. Despite being higher eukaryotes, plants have completely different mechanisms of uptake, distribution, storage, compartmentalization and metabolism of various pollutants. The influence of pollutants on an open-air phytosensor depends on the interaction of plants with the environment. Various abiotic and biotic stresses, such as wind, drought, and light intensity, bacterial, fungal and viral infections can potentially influence the response of plants to the pollutant in question and hence, result in false-negative/false-positive outcomes. These are perhaps the main drawbacks that prevent plants from being used in standard genotoxicity assays and being approved without reserve by regulatory organizations.
Classical plant biosensors
The most commonly assays used for studying mutagenicity of various pollutants in plants are based on the detection of chromosomal aberrations in Allium cepa [24, 25], Tradescantia [26], Vicia faba plants [27] or Zea mays [28]. An Allium cepa chromosome aberration test that can serve as a rapid screen for toxic effects of chemicals is among them [29-30]. Due to its sensitivity, the Allium cepa test was the first of nine plant assay systems evaluated by the Gene-Tox Program of the US Environmental Protection Agency [29]. The assay is based on the evaluation of the percentage of aberrant mitotic events and different fractions of chromosomal aberrations [31].
Tradescantia is another important plant for mutagenesis studies. This plant makes it possible to perform chromosome aberration, stamen-hair mutation and micronuclei formation assays [32]. It has been used to analyze the air quality in high traffic areas [33], municipal waste storages [34], and the quality of air from a landfill vent pipe [34]. This plant has also been used for the analysis of water and soil pollution, and even for the evaluation of bioremediation effectiveness at hazardous waste sites [35-37].
Vicia faba is yet another plant commonly used as a biosensor. Various chemicals have scored positive in the Vicia faba-based sister chromatide exchange assay [25, 38].
Among plants that are less commonly used for biomonitoring, tobacco plants heterozygous for the sulfur (Su) nuclear gene have also proved to be useful. Su is a nuclear encoded, semi-dominant aurea mutation in Nicotiana tabacum. The homozygous plants (Su/Su) are pale yellow and non-photosynthetic while the heterozygous (Su/+) are photosynthetically competent and have a yellow-green phenotype which is distinct from that of green wild-type plants (+/+) [39-40]. The Su/+ plants exposed to gamma-radiation and treated with chemicals reveal a strong increase in the number of dark-green spots on light-green leaves in comparison to non-treated plants [41]. The appearance of dark green sectors can serve as an indication of mutagenicity due to various mutations (Figure 1A), whereas the appearance of twin-spots of darker green and albino is an indication of genome instability due to a homologous recombination event (Figure 1B).
Figure 1.
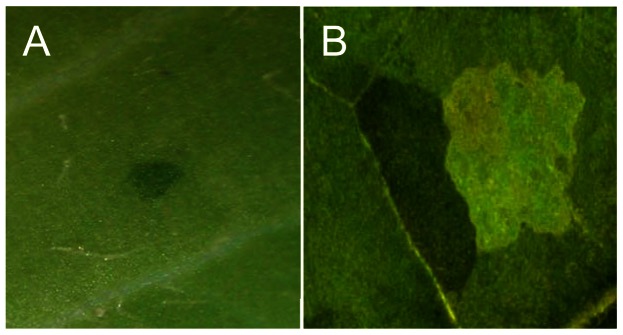
Non-transgenic tobacco-based system for detecting the genotoxicity. The system is based on naturally occurring mutation at the Sulfur (Su) gene. Plants heterozygous for the gene, Su/+ are pale green. Mutations that inactivate the only Su allele are visualized as dark green spots (A). Rearrangements that occur between Su and + allele result in the formation of a “twin” spot visualized as a dual sector of dark green (+/+) and albino (Su/Su) (B).
Non-transgenic biosensors
Genomic regions containing simple sequence repeats, also known as microsatellites, are extremely unstable. The analysis of their expansion/retraction upon exposure to the environmental pollution can be used for monitoring and analyzing germline mutations (the latter is a particularly difficult process). The low mutation rate, around 10-6 per gene [42], does not allow the frequent appearance of new phenotypes in the next generation. In contrast, loci containing microsatellites are several orders of magnitude more unstable, with mutation rate being 10-2-10-4. The assay is used to evaluate the influence of mutagens on the germline. The exposed seeds are germinated, and the extracted DNA is used to PCR amplify various loci containing unstable microsatellites. Using this approach, two genetically identical wheat (Triticum aestivum, L.) populations have been profiled. The growth of plants in soil with the surface radioactive contamination density of 900 Ci/km2 and an external gamma dose rate of 980 μR/h resulted in an over 6-fold increase in germline mutation rate [43]. Further analysis of changes occurred at the molecular level showed an extremely complex pattern of radiation induced instabilities [44].
Transgenic plant biosensors
Much public debate and heated controversy exist surrounding the topic of consumption of genetically modified organisms (GMOs). The alternative use of transgenic plants in horticulture, forestry, and construction seems to be more appealing for the public. In this respect, design and production of transgenic plants for environmental biomonitoring and cleaning up polluted areas can be “a bridge pass” for more favorable public perception of GMOs.
In recent years, substantial progress in generation and exploitation of transgenic plants as biomonitors has been made [45-53]. One of the important advantages of transgenic biosensors is the ability to customize the assay in accordance with monitoring needs. This not only makes transgenic biosensors more sensitive to a particular pollutant, but also allows for easy scoring.
Another big advantage of a customized transgenic assay is the ability to understand changes at the molecular level. Transgenic plant biomonitors used for the evaluation of genotoxicity are relatively cheap and simple in use. The assays used in the past decade were based on the restoration of the transgenes β-glucuronidase (uidA or GUS) and luciferase activity in Arabidopsis thaliana or Nicotiana tabacum plants transformed with non-active forms of these marker genes.
Recombination reporter assay
The very first transgenic biomonitor for the detection of recombination events was produced by Lebel et al. (1993) [45]. The authors generated a construct based on two overlapping, non-functional copies of a kanamycin resistance gene. Recombination events were scored in protoplasts of transgenic tobacco plants that were propagated on kanamycin. Homologous recombination (HR) events restored the gene activity, and protoplasts resistant to kanamycin were regenerated [45]. Protoplasts were shown to be sensitive to X-rays, mitomycin C and heat shock [45]. The system has never been used for biomonitoring due to the laborious tissue culture procedures necessary for the culturing of protoplasts and the detection of recombination events.
Another system for the detection of HR events in transgenic Arabidopsis and tobacco plants utilized the overlapping and truncated versions of a β-glucuronidase [54]. Recombination events at a transgene locus lead to restoration of the uidA (GUS) transgene and synthesis of the β-glucuronidase enzyme (Figure 2). Upon histochemical staining, the enzyme cleaves the substrate X-gluc which results in the formation of a blue precipitate. Upon ethanol treatment and chlorophyll removal, cells in which recombination events occurred can be precisely localized as blue sectors on the transparent plants. This enables developing a quantitative assay (Figure 2). The plants were scored positive in response to a variety of DNA damaging agents such as UV-C irradiation and MMS treatment [55].
Figure 2.

Transgenic “recombination” system for the detection of environmental mutagens. Transgenic plants carry in the genome two non-functional truncated copies of the GUS gene, depicted as “UG” and “US”. The two parts of the truncated, overlapping GUS gene can be in either orientation with respect to each other. Activation of the β-glucuronidase (GUS) gene via homologous recombination (HR) restores the gene activity and is visualized as blue spots after histochemical staining.
Perhaps the biggest success of transgenic recombination plants has been their use for the detection of radioactive pollution of soil and water. Large-scale environmental monitoring experiments employing transgenic Arabidopsis thaliana and Nicotiana tabacum plants showed substantial genotoxicity of soil patterns from different contaminated areas in Ukraine. The plants responded to higher level of contamination with a higher increase in HR frequency [46-47]. The use of plants as bioindicators allowed calculating an absorbed dose of radiation for A. thaliana. This dose consists of external and internal doses. It was possible to correlate recombination frequency not only to the level of soil contamination but also to the absorbed dose. The sensitivity of these plants made it possible to detect the HR frequency difference between plants grown in very low-contaminant “clean” soil (22 Bq/kg) and plants grown in contaminated soil, where the contamination level is as low as 1.5-3.3 Ci/km2 (188-575 Bq/kg) [47].
It should be noted that transgenic systems used as biosensors can be compared in sensitivity with the well-established Allium cepa chromosomal aberration assay. Mitotic root-tips of onion seeds germinated in the same contaminated soils exhibited the chromosomal aberrations level that was significantly correlated with the HR level in transgenic plants (r> 0.90, n=6, P<0.05) [56].
Point mutation-detection assay
Transgenic biomonitoring of HR frequency permits the detection of even minor portions of all possible changes in DNA. The majority of changes are point mutations. In contrast to animals, plants do not have a predetermined germ line, and thus any somatic mutation in the meristem can potentially be inherited.
In one of our previous studies, we developed a system that would allow the detection of somatic point mutations. We have introduced a stop codon sequence at the very 5′ end of the GUS (uidA) gene by the substitution of a single nucleotide [42]. The new stop codon resulted in complete inactivation of the transgene. Transgenic plants carrying such a construct exhibited occasional sectors of blue upon histochemical staining. These sectors represent spontaneous restoration of uidA activity from reversion of stop codons to original codons. These plants responded strongly to various mutagens such as UV-C, X-rays and methyl methanesulfonate (MMS) by increasing the mutation frequency in a transgene [42].
Further studies confirmed the usefulness of these plants for biomonitoring. Plants germinated at various concentrations of heavy metal salts such as Cd 2+, Pb 2+, Ni 2+, Zn 2+, Cu 2+ showed substantial dose- and uptake-dependent increase in the frequency of point mutations. It is noteworthy that the increase in mutation frequency was strongly and positively correlated with the increase in HR frequency in recombination plants grown in the presence of the same concentrations of aforementioned salts [49].
In a recent report, Van der Auwera et al. (2008) [53] described a very valuable model that is an improvement to the one reported by Kovalchuk et al. (2000) [42]. Inactivating nucleotide substitutions in the GUS gene reported by Kovalchuk et al. (2000) [42] were not designed to score for C:G-to-T:A transition events. This particular mutation, however, has been shown to be induced by several mutagenic treatments in E. coli [58]. In Arabidopsis, C:G-to-T:A mutations are typically initiated by deamination of methylated cytosine residues at symmetrical CpG or CpNpG positions. This is believed to be one of the reasons why these sequences are underrepresented in the genome [59]. Many commonly-used chemical mutagens, such as ethyl methanesulfonate (EMS), predominantly induce C:G-to-T:A transitions [60-62]. Thus, generation of plants carrying a substrate for this type of mutation was a logical step. Van der Auwera et al. (2008) [53] reported spontaneous mutation frequency that was substantially higher to the one observed by Kovalchuk et al. (2000) [42]. Perhaps, it can be explained by the fact that C:G-to-T:A transitions are more common in the plant genome. The authors found that ethyl methanesulfonate and ultraviolet-C induced the mutation frequency, whereas heavy metals, methyl jasmonate, salicylic acid, and heat stress did not [53]. This result suggests that several systems for measuring different end-points should be used simultaneously for the analysis of potential mutagenicity, since none of the systems alone is capable of being sensitive to every mutagen.
Comparison of sensitivity of transgenic and non-transgenic systems
Most non-transgenic plant biomonitors rely on the appearance/disappearance of a certain phenotype or/and on the visualization of gross-chromosomal rearrangements. Many point mutations are either synonymous (without any change in the encoded amino acid) or conservative (with some changes in the biochemically similar amino acid). These changes would typically be undetected in many systems that rely on the appearance of a certain phenotype. In contrast, the transgenic “mutation” system, that we developed, senses genotoxic agents that cause point mutations.
We have evaluated the data obtained from our transgenic plant systems as well as reports found in the literature [16, 63-71]. It should be noted that in various systems being used, the tested parameters differ significantly, the data among systems cannot be directly compared. Nonetheless, our transgenic systems appear to be much more sensitive than other available assays on mutagenicity of various heavy metals.
It should also be mentioned that only some of the tested systems, (the E. coli based MetPLATE test is among them), have been able to detect mutagenicity of Cd2+ of less than 0.05 mg/L [64]. Our approach allows us to detect more than a two-fold increase in both, mutation and recombination levels after plants were exposed to the same or lower concentration of Cd2+. Several other systems presented by Kong et al. (1995) [69] were comparably sensitive to the transgenic Arabidopsis system. Exposure of several invertebrate species (Ceriodaphnia dubia, Daphnia magna, Daphnia pulex) to As, Cd, Cu, Ni, Pb and Zn resulted in EC50 (mg/L) of 1.8-6.6, 0.05-0.35, 0.018-0.23, 0.14-7.6, 0.53-4.9, and 0.05-5.1, respectively. Median lethal concentrations of Cd, Cu and Zn after 96 hrs of exposure of Pimephales promelas were 0.01-2.2 mg/l, 0.02-0.1 mg/L and 0.33-1.7 mg/L, respectively [69]. Our system could also detect toxic effects of Cd, Cu, Ni, Pb and As at concentrations as low as 0.001, 0.05, 0.1, 0.5 and 0.05 mg/L, respectively [49].
Moreover, the transgenic Arabidopsis systems were much more sensitive than the non-transgenic Tradescantia MCN assay [70]. It could be explained by differences in time of exposure to mutagenic compounds. Plants used in our assays were germinated in the presence of a mutagen and were subsequently exposed for 25-35 days, whereas in the Tradescantia MCN assay cuttings were exposed for 6 h, and intact plants were exposed for only 72 h.
The other cell culture-, bacterial-, and mammalian-based tests were significantly less sensitive than the transgenic plant-based tests. The bacterial systems were less sensitive than our system, but they are not necessarily less sensitive than the other systems used for genotoxicity studies. The Allium cepa chromosome aberration assay, another frequently used assay, also exhibited higher sensitivity than the bacterial Ames and Microscreen tests [63].
It is difficult to explain why plant-based assays detect heavy metals with higher sensitivity. It is quite possible that plants accumulate higher levels of metal ions than other organisms. Although the hypothesis that active heavy metal concentrations (concentrations in the organism) are higher in plants than in animals is an attractive one, it still remains untested because data comparing heavy metal uptakes in different organisms are not available.
Transgenic systems can be efficiently used to sense other types of environmental mutagens
The transgenic Arabidopsis and tobacco systems described here have also been used for the analysis of mutagenicity of other factors in soil, water or air. In our previous research, we analyzed potential genotoxicity of radioactively polluted water. We sampled water from private wells in the villages from the inhabited areas contaminated after the Chernobyl accident. The radiological analysis of water samples did not reveal any 137Cs or 90Sr activity because concentrations of these radionuclides were below detectable limits. Despite this fact, we found an increase of HR frequency in A. thaliana grown in the media prepared from contaminated water [44]. We speculated that the increase was attributed to low-level contamination in drinking water. It is, however, possible that the observed effects were due to a yet unknown factor (a contaminant).
Recently, we have tested our systems for sensing soil contaminated by herbicides. We found that the herbicides 2,4-D and dicamba increased both the HR frequency and the frequency of point mutations [72]. In contrast, herbicide atrazine increased only the HR frequency but not point mutations [50]. The fact that these herbicides influence the HR frequency and point mutation frequency in a different way is very important; that there is herbicide-specificity in DNA damage mechanisms.
Transgenic recombination lines were also used for the analysis of potential mutagenic influence of UV-B radiation. Sun simulators were used to mimic various levels of UV-B, from slightly elevated levels to those that currently do not exist on earth. The system showed a direct positive correlation between the level of UV-B and recombination [48]. For the first time, this transgenic system was used for the analysis of heritable changes, and an increase in germline recombination was observed. Elevated UV-B levels resulted in an increase in the number of plants that stained totally blue. These plants inherited the restored marker gene [48].
Another stress that frequently accompanies an increase in UV-B levels is a temperature increase. It is definitely a powerful type of stress, although it is not considered to be an environmental pollutant. The mutagenic effects of elevated temperatures were documented as early as in 1993, when Lebel et al. (1993) [45] reported an increase of the homologous recombination frequency in tobacco plants that were briefly exposed to 50 °C. The authors reported the over 5-fold increase in rearrangements of the kanamycin-based homologous recombination [45]. Similar data were obtained by Boyko et al. (2005) [73]. The growth of Arabidopsis plants carrying the GUS-based HR construct at 32 °C resulted in the 5-10 fold increase in the HR rate (HR frequency is related to the number of genomes per plant) [73]. The authors also reported a substantial increase in the recombination rate in plants exposed to 4°C. These publications indeed support the notion that shifts in temperature represent a powerful genotoxic stress.
Lately we have tested the influence of biotic stress factors on the genome stability of plants. In our work, we showed that exposure to viral pathogens resulted in an increase of HR frequencies in both somatic [74] and meiotic cells [75]. Our recent data also suggest that bacterial pathogen infection of plants results in genome destabilization (data not published). These experiments demonstrate that transgenic plants carrying a marker that can detect genome destabilization are able to sense a variety of mutagens in the environment.
Many excellent assays used currently are missing one characteristic feature: they cannot be used for analyzing transgenerational changes in the genome. These changes are of a critical importance, since many mutagens may not exhibit immediate mutagenicity (in somatic cells) but can lead to changes in the next generation. Previously, several publications reported heritable changes in inbred flax in response to specific environmental changes such as nutrient balance and temperature regimes [76-77]. This appeared to originate from the activity of a transposon-like sequence, LIS1, which assembles and inserts itself into the genome of stressed flax plants [77]. A new genotroph is apparently stable, as no further changes occur in the activity of this transposon in plants upon exposure to additional stresses [77]. The report by Ries et al. (2000) [48] also suggests that exposure to stress, specifically to UVB, results in an increase in genomic instability in the progeny.
The experiments in our lab showed that various types of stresses, including water stress, heavy metal stress and pathogen stress, result not only in an increase of the HR frequency in somatic cells but, more importantly, they cause heritable changes in the progeny (data not published). The progeny of stressed plants exhibited elevated levels of the spontaneous HR frequency. This suggests that plants inherit “memory” of stress, as it was recently confirmed by Molinier et al. (2006) [78]. Their studies showed that single exposure of plants to UVC results in an increase of the HR frequency in many generations of plants. Especially curious is the fact that plants remember stress exposure for 5-6 generations, thus maintaining the elevated levels of HR frequencies even without the presence of stress itself. Our recent work supports this finding partially. We showed that memory of stress does not last for more than 1 or 2 generations, unless plants are constantly exposed to stress (data not shown). The discrepancy can be explained by the fact that we used natural stresses, such as drought, flood, heavy metals, etc., whereas Molinier et al. (2006) [78] used exposure to more artificial stress such as UVC.
Marker genes to be used for mutagenesis assays
Marker gene expression is relatively easy to observe in plants. The most suitable marker genes are GUS (β-glucuronidase), LUC (luciferase) and various fluorescent proteins (FPs), including green FP (GFP), yellow FP (YFP), red FP (e.g., DsRed, from the coral Discosoma sp.) [79]. GUS and LUC have been commonly used in plants [80-82], among FPs, only GFP has been used extensively [79, 83, 84].
All the aforementioned reporter genes have different detection sensitivity in plants. To the best of our knowledge, the GFP gene-marker is perhaps one of the easiest and cheapest to visualize. It just requires a UV lamp or a lamp emitting blue light to be seen as green on a red background [84]. The main problem of this marker is that it has the lowest sensitivity among all the GFP, GUS and LUC transgenes applied in biomonitoring; i.e., it requires the highest expression for detection.
The GUS gene-marker is the second cheapest and easiest to use. The GUS protein can be visualized upon histochemical staining by a substrate for the β-glucuronidase enzyme, the chemical X-glu. The reaction results in the formation of a blue precipitate accumulating in cells. The LUC-gene marker is the most difficult and expensive to visualize. Its advantage, however, is that it is, by far, the most sensitive one [80]. As in the case of the GUS gene-marker, a substrate needs to be provided. In this case, the plants are sprayed with the substrate, luciferin. The cleavage of luciferin by luciferase results in the emission of luminescence that is detected by a luminometer. The major advantage of the luciferase reporter gene system is its in vivo detection of luciferase activity. One of the disadvantages of the GUS marker is that it requires the destructive non-vital histochemical staining for the visualization of events, and thus can be done only once, at specific time points during plant growth. In contrast, the detection of the LUC and GFP markers does not kill the plants and can be done at any time points during plant growth. GFP requires no substrate and can be assayed on plants in the field.
The GUS and LUC-based reporter assays for the detection of potential mutagenicity of pollutants has been used before [46-50, 52]. These repair assays were based on the detection of the marker gene activity upon restoration of gene integrity. The luciferase–based recombination assay was similar in design to the GUS-based assay [80]. Frequent recombination events were observed using an in vivo imaging system and visualized before and after the application of mutagens (Figure 3).
Figure 3.

Transgenic plants carrying the luciferase recombination marker allow continuous visualization of recombination events. A). Recombination events in plants before the application of the mutagene. B). Recombination events in the same plants 3 days after the UVC treatment.
We have also tested another transgenic visual marker frequently used in mammalian research, the GFP marker. This marker gene appeared not to be suitable for recombination and mutation assays. While the whole GFP+ plants are easily detectable, single cells with recombination or mutation events leading to restoration of GFP function are more difficult to detect, requiring laborious microscopic analysis (Jan Lucht, personal communication). The FP-based recombination reporter has been, however, successfully used in animals. Transgenic mice carrying a recombination substrate based on the enhanced yellow fluorescent protein have been generated [18]. However, the detection of DNA damage using this system requires a complex procedure that includes sorting of fluorescent cells using a flow cytometer [18].
Other transgenic systems for the detection of environmental mutagens
Although a recombination reporter line detects mutagenicity of various factors efficiently, it still depends on the mutagen's ability to trigger double strand breaks (DSBs) in a marker gene. Presumably, agents that do not cause DSB would not score positive in this assay. We have designed another system based on the stop codon reversion [42]. Here, there is potential for increased efficacy, but there are several drawbacks. First, each transgenic line allows the detection of only one particular type of mutation. Second, the frequency of these mutations is typically very low; 500-1000 plants per each experimental group are needed to obtain statistically significant data. The low frequency of point mutations, which is observed in the plant lines, is due to a small target: a single base pair in the diploid genome. Recently, an improved version of this system has been published [53]. The authors designed the GUS-based marker that scores C:G-to-T:A transitions that are common mutations in a plant genome. The frequency of point mutations detected in these plants was substantially higher than that observed by Kovalchuk et al. (2000) [42]. This system sensitivity to various environmental pollutants remains to be established.
Recently, another transgenic Arabidopsis-based reporter system has been published. These reporter plants carry in their genome the GUS gene inactivated by the introduction of a microsatellite marker in the 5′ of the gene directly after the AUG codon [85]. The gene activity is restored via the gain or loss of one or several repeats (Figure 4). Since microsatellites usually have a high mutation frequency, the number of blue spots in these plants is also high. This allows the use of significantly fewer plants for the analysis of potential mutagenesis.
Figure 4.

Schematic diagram of microsatellite-containing phytosensor. Active copy of the β-glucuronidase (GUS) gene was disrupted by integration of microsatellite containing sixteen G nucleotides (shown in green). This resulted in a complete inactivation of the gene (pale blue). Mutations leading to gain of 2 guanines or loss of 1 guanine restore the frame and result in activation of the transgene (dark blue).
Another way to increase a target size would be to design a marker that functions in the form of a repressor-test gene combination. In this case, an active repressor would bind to a promoter sequence blocking a reporter gene expression (Figure 5). The tetracycline repressor is one of the most commonly used repressors [86]. Inactivation of the tetracycline repressor by any kind of mutation would restore promoter function and activate reporter transgene. Plants that carry such a construct would have a substantial number of sectors expressing the transgene. This transgenic line allows us to use a significantly smaller number of plants and have some advantages with regards to statistical analysis of data. As any system, there are potential disadvantages: it requires repressor binding to be tight enough to prevent any “leakage”, and moreover, plants used for testing have to be heterozygous for such a construct to ensure inactivation of a single copy repressor element by a mutation event.
Figure 5.

Possible tetracycline repressor-based system. A) Marker gene placed under the tetracycline repressor/promoter element is not active since the repressor blocks the promoter. B) Any mutation (point, deletion, insertion etc.) in the repressor region leading to production of inactive repressor or no repressor at all will result in activation of the marker gene. This should result in the appearance of discrete spots representing cells and their progeny where the mutation has occurred.
There are many other ways of designing a good transgenic biosensor. Introducing simple frame shifts might abolish gene activity. Any mutation, deletion or insertion that restores the reading frame would potentially activate the transgene. One more way of making a transgenic biosensor is to generate plants carrying an inactive version of the antibiotic (or herbicide) resistance gene. The progeny of exposed plants can be grown in the presence of a selective agent, and resistant plants are scored. This approach is good for the detection of inherited spontaneous or induced mutation events. Although the approach looks rather appealing, it requires a longer time for the evaluation of environmental influences, and a large number of progeny plants must be scored. We have recently designed plants that carry a dual recombination marker: a visible marker based on the luciferase transgene and a sulfonamide antibiotic-based marker. Whereas the luciferase-based marker allows scoring somatic and meiotic events, the sulfonamide-based marker makes it possible to score only meiotic events (data not published). Preliminary data show a high efficiency of such system.
Other transgenic systems may include promoters that are regulated either by stress or by specific metabolites. Stress-regulated promoters could be specific to a particular chemical or a physical agent, or they could be broad and activated by the most common types of stresses. Initial steps in the identification of stress-regulated promoters would include transcription profiling of plants grown under various stresses or under a specific stress in question. Obviously, only a promoter activated in the presence of stress should be used [87]. Alternatively, the minimal promoter for incorporating specific regulatory elements activated by stress can be used. These regulatory elements can be found in genes such as ATM, HSP70, etc. The approach has been effectively used by Saidi et al. (2007) [88]. Transgenic Physcomitrella patens expressing the GUS reporter under the control of the stress-inducible promoter hsp17.3B have been generated. They have shown to react to chlorophenols, heavy metals and sulphonated anthraquinones [88].
Yet another transgenic system to be generated would detect epimutations. This term is used for the (ir)reversible modification of gene expression that is not associated with a change in a DNA sequence. There are many agents such as DNA methylation and histone modifications that do not cause DNA damage but potentially influence the epigenetic regulation of cells. Methylation changes have indeed been found in the progeny of plants exposed to radiation. Substantial global genome hypermethylation in the progeny of pine trees grown in Chernobyl has been reported [89]. Potential transgenic systems would have a marker gene under the influence of a heavily methylated promoter. This is possible to achieve by generating several hundreds of transgenic lines and by selecting a line that will not have any transgene expression. Such plant has been generated by Amedeo et al. (2000) [90], although it has not been used specifically for biomonitoring. A transgenic line of this type should have an intact but heavily methylated transgene. The promoter can be modified by generating GC-rich clusters. Potential epimutagens can be screened by detecting transgene activation.
Summary and Outlook
Evaluation of environmental contamination levels is a difficult task that requires steady attention and efforts. Patterns of water, soil and air pollution are constantly changing, which require new and improved biosensors. Here, we have described several transgenic plants that have already been used or could potentially be used as phytosensors. These plants can be effectively used for both sensing environmental pollution and monitoring the efficiency of polluted areas decontamination. However, it should be noted that the transgenic phytosensors described above would not be able to differentiate between different genotoxic agents. Novel, mutagen-specific transgenic plant biosensors should be generated on demand, depending on mutagens to be tested.
Acknowledgments
We are grateful to Valentina Titova for proofreading of the manuscript. The original work presented in this review was supported by the NSERC operating grants to O.K. and I.K.
References
- 1.Harwell M., Gentile J., Norton B., Cooper W. Ecological Risk Assessment issue Papers. U.S. Environmental Protection Agency; Washington, D.C.: 1994. Issue paper on ecological significance. EPA/630/R-94/009. [Google Scholar]
- 2.Greenberg B. M., Wilson M. I., Huang X.-D., Duxbury C. L., Gerhardt K. E., Gensemer R. W. The effects of ultraviolet-B radiation on higher plants. In: Wang W., Gorsuch J. W., Hughes J., editors. Plant for Environmental Studies. CRC Press; Boca Raton, Florida: 1997. pp. 1–36. [Google Scholar]
- 3.Holst R., Nagel D. Radiation effects on plants. In: Wang W., Gorsuch J. W., Hughes J., editors. Plants for Environmental Studies. CRC Press; Boca Raton, Florida: 1997. pp. 38–79. [Google Scholar]
- 4.Kovalchuk I., Kovalchuk O. A new use for transgenic plants – environmental biomonitors. Biotechnology and Genetic Engineering Reviews, Annual Review. 2003;20 doi: 10.1080/02648725.2003.10648036. Chapter 1. [DOI] [PubMed] [Google Scholar]
- 5.Clemens S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie. 2006;88:1707–19. doi: 10.1016/j.biochi.2006.07.003. [DOI] [PubMed] [Google Scholar]
- 6.Sharma S. S., Dietz K. J. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot. 2006;57:711–26. doi: 10.1093/jxb/erj073. [DOI] [PubMed] [Google Scholar]
- 7.Wagner G., Nadasy E. Effect of pre-emergence herbicides on growth parameters of green pea. Commun. Agric. Appl. Biol. Sci. 2006;71:809–13. [PubMed] [Google Scholar]
- 8.Vij S., Tyagi A. K. Emerging trends in the functional genomics of the abiotic stress response in crop plants. Plant Biotechnol. J. 2007;5:361–80. doi: 10.1111/j.1467-7652.2007.00239.x. [DOI] [PubMed] [Google Scholar]
- 9.Levin D.E., Ames B.N. Classifying mutagens as to their possible specificity in causing the six possible transitions and transversions: a simple analysis using the Salmonella mutagenicity assay. Environ. Mutagen. 1986;8:9–28. doi: 10.1002/em.2860080103. [DOI] [PubMed] [Google Scholar]
- 10.Miller J.H. Mutagenic specificity of ultraviolet light. J. Mol. Biol. 1985;182:45–68. doi: 10.1016/0022-2836(85)90026-9. [DOI] [PubMed] [Google Scholar]
- 11.Schaaper R.M., Dunn R.L. Spontaneous mutation in the Escherichia coli. Carcinogenesis. 1991;11:1087–1095. [Google Scholar]
- 12.Moller P. The alkaline comet assay: towards validation in biomonitoring of DNA damaging exposures. Basic Clin. Pharmacol. Toxicol. 2006;98:336–45. doi: 10.1111/j.1742-7843.2006.pto_167.x. Review. [DOI] [PubMed] [Google Scholar]
- 13.Simpson . Sale Sister chromatid exchange assay. In: Jean-Marie Buerstedde J.-M., Takeda S., editors. Reviews and Protocols in DT40 Research. Springer; The Netherlands: 2006. pp. 399–403. [DOI] [PubMed] [Google Scholar]
- 14.Amanuma K., Takeda H., Amanuma H., Aoki Y. Transgenic zebrafish for detecting mutations caused by compounds in aquatic environments. Nature Biotechnol. 2000;18:62–65. doi: 10.1038/71938. [DOI] [PubMed] [Google Scholar]
- 15.Murti R., Schimenti K., Schimenti J. A recombination-based transgenic mouse system for genotoxicity testing. Mutat Res. 1994;307:583–595. doi: 10.1016/0027-5107(94)90268-2. 1994. [DOI] [PubMed] [Google Scholar]
- 16.Sacco M.G., Zecca L., Bagnasco L., Chiesa G., Parolini C., Bromley P., Cato E.M., Roncucci R., Clerici L.A., Vezzoni P. A transgenic mouse model for the detection of cellular stress induced by toxic inorganic compounds. Nature Biotechnol. 1997;15:1392–1397. doi: 10.1038/nbt1297-1392. [DOI] [PubMed] [Google Scholar]
- 17.Mayer C., Klein R.G., Wesch H., Schmezer P. Nickel subsulfide is genotoxic in vitro but shows no mutagenic potential in respiratory tract tissues of BigBlue rats and Muta Mouse mice in vivo after inhalation. Mutat Res. 1998;420:85–98. doi: 10.1016/s1383-5718(98)00140-5. [DOI] [PubMed] [Google Scholar]
- 18.Hendricks C.A., Engelward B.P. “Recombomice”: the past, present, and future of recombination-detection in mice. DNA Repair (Amst). 2004;3:1255–61. doi: 10.1016/j.dnarep.2004.04.006. [DOI] [PubMed] [Google Scholar]
- 19.Rugo R.E., Almeida K.H., Hendricks C.A., Jonnalagadda V.S., Engelward B.P. A single acute exposure to a chemotherapeutic agent induces hyper-recombination in distantly descendant cells and in their neighbors. Oncogene. 2005;24:5016–25. doi: 10.1038/sj.onc.1208690. [DOI] [PubMed] [Google Scholar]
- 20.Green M., Todo T., Ryo H., Fujikawa K. Genetic-molecular basis for a simple Drosophila melanogaster somatic system that detects environmental mutagens. Proc. Nat. Acad. Sci. USA. 1986;83:6667–6671. doi: 10.1073/pnas.83.18.6667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sheehan P.J., Loucks O.L. Ecological Risk Assessment issue Papers. U.S. Environmental Protection Agency; Washington, D.C.: 1994. Issue paper on effects characterization. EPA/630/R-94/009. [Google Scholar]
- 22.McVey M., Macler B.A. A Review of Ecological Assessment Case Studies from a Risk Assessment Perspective. U.S. Environmental Protection Agency; Washington, D.C.: 1993. Ecological risk assessment case study: selenium effects at Keterson Reservoir. EPA/630/R-92/005. [Google Scholar]
- 23.Notten M.J., Oosthoek A.J., Rozema J., Aerts R. Heavy metal concentrations in a soil-plant-snail food chain along a terrestrial soil pollution gradient. Environ. Pollut. 2005;138:178–90. doi: 10.1016/j.envpol.2005.01.011. [DOI] [PubMed] [Google Scholar]
- 24.Fiskesjo G. Allium test. Methods Mol. Biol. 1995;43:119–27. doi: 10.1385/0-89603-282-5:119. [DOI] [PubMed] [Google Scholar]
- 25.Ma T.H., Cabrera G.L., Owens E. Genotoxic agents detected by plant bioassays. Rev. Environ. Health. 2005;20:1–13. doi: 10.1515/reveh.2005.20.1.1. [DOI] [PubMed] [Google Scholar]
- 26.Ichikawa S. Tradescantia stamen-hair system as an excellent botanical tester of mutagenicity: its responses to ionizing radiations and chemical mutagens, and some synergistic effects found. Mutat. Res. 1992;270:3–22. doi: 10.1016/0027-5107(92)90096-k. [DOI] [PubMed] [Google Scholar]
- 27.Kanaya N., Gill B., Grover I., Murin A., Osiecka R., Sandhu S., Andersson H. Vicia faba chromosomal aberration assay. Mutat. Res. 1994;310:231–247. doi: 10.1016/0027-5107(94)90116-3. [DOI] [PubMed] [Google Scholar]
- 28.Grant W.F., Owens E.T. Zea mays assays of chemical/radiation genotoxicity for the study of environmental mutagens. Mutat. Res. 2006;613:17–64. doi: 10.1016/j.mrrev.2006.04.002. [DOI] [PubMed] [Google Scholar]
- 29.Grant W. F. The present status of higher plant bioassays for detection of environmental mutagens. Mutat. Res. 1994;310:175–185. doi: 10.1016/0027-5107(94)90112-0. [DOI] [PubMed] [Google Scholar]
- 30.Bolle P., Mastrangelo S., Tucci P., Evandri M.G. Clastogenicity of atrazine assessed with the Allium cepa test. Environ. Mol. Mutagen. 2004;43:137–41. doi: 10.1002/em.20007. [DOI] [PubMed] [Google Scholar]
- 31.Fiskesjo G. The Allium test - an alternative in environmental studies: the relative toxicity of metal ions. Mutat. Res. 1988;197:243–260. doi: 10.1016/0027-5107(88)90096-6. [DOI] [PubMed] [Google Scholar]
- 32.Rodrigues G.S., Ma T.H., Pimentel D., Weinstein L. Tradescantia bioassays as monitoring systems for environmental mutagenesis: a review. Crit. Rev. Plant Sci. 1997;16:325–359. [Google Scholar]
- 33.Ma T.H., Harris M.M. In situ monitoring of environmental mutagens. In: Saxena J., editor. Hazard Assessment of Chemicals, Current Developments. Vol. 4. Academic Press; New York: 1985. pp. 77–106. [Google Scholar]
- 34.Ma T.H., Xu C., Powers L., Liao S. In situ monitoring with the Tradescantia bioassays on the genotoxicity of gaseous emissions from a closed landfill site and an incinerator. Mutat. Res. 1996;359:39–52. doi: 10.1016/s0165-1161(96)90008-4. [DOI] [PubMed] [Google Scholar]
- 35.Sandhu S.S., Ma T.H., Peng Y., Zhou X. Clastogenicity evaluation of seven chemicals commonly found at hazardous industrial waste sites. Mutat. Res. 1989;224:437–446. doi: 10.1016/0165-1218(89)90068-2. [DOI] [PubMed] [Google Scholar]
- 36.Ruiz E.F., Rabago V.M.E., Lecona S.U., Perez A.B., Ma T.H. Tradescantia micronucleus (Trad-MCN) bioassay on clastogenicity of wastewater and in situ monitoring. Mutat. Res. 1992;270:45–51. doi: 10.1016/0027-5107(92)90100-g. [DOI] [PubMed] [Google Scholar]
- 37.Baud-Grasset S., Baud-Grasset F., Bifulco J.M., Meier J.M., Ma T.H. Reduction of genotoxicity of a creosote-contaminated soil after fungal treatment determined by the Tradescantia micronucleus test. Mutat. Res. 1993;303:77–82. doi: 10.1016/0165-7992(93)90098-g. [DOI] [PubMed] [Google Scholar]
- 38.Sang N., Li G. Genotoxicity of municipal landfill leachate on root tips of Vicia faba. Mutat. Res. 2004;560:159–65. doi: 10.1016/j.mrgentox.2004.02.015. [DOI] [PubMed] [Google Scholar]
- 39.Burk L.G., Menser H.A. A dominant aurea mutation in tobacco. Tob. Sci. 1964;8:101–104. [Google Scholar]
- 40.Kawata E. E, Cheung A.Y. Molecular analysis of an aurea photosynthetic mutant (Su/Su) in tobacco: LHCP depletion leads to pleiotropic mutant phenotypes. EMBO J. 1990;9:4197–4203. doi: 10.1002/j.1460-2075.1990.tb07644.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Friedlender M., Lev-Yadun S., Baburek I., Angelis K., Levy A.A. Cell divisions in cotyledons after germination: localization, time course and utilization for a mutagenesis assay. Planta. 1996;199:307–313. [Google Scholar]
- 42.Kovalchuk I., Kovalchuk O., Hohn B. Genome-wide variation of the somatic mutation frequency in transgenic plants. EMBO J. 2000;19:4431–4438. doi: 10.1093/emboj/19.17.4431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kovalchuk O., Dubrova Y., Arkhipov A., Hohn B., Kovalchuk I. Wheat DNA mutation rate after Chernobyl. Nature. 2000;407:583–584. doi: 10.1038/35036692. [DOI] [PubMed] [Google Scholar]
- 44.Kovalchuk O., Telyuk P., Kovalchuk L., Kovalchuk I., Titov V. Novel plant bioassays for monitoring the genotoxicity of drinking water from the inhabited areas of the Ukraine affected by the Chernobyl accident. Bull. Environ. Contam. Toxicol. 2003;70:847–53. doi: 10.1007/s00128-003-0060-y. [DOI] [PubMed] [Google Scholar]
- 45.Lebel E.G., Masson J., Bogucki A., Paszkowski J. Stress-induced intrachromosomal recombination in plant somatic cells. Proc. Natl. Acad. Sci. U S A. 1993;90:422–6. doi: 10.1073/pnas.90.2.422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Kovalchuk I., Kovalchuk O., Arkhipov A., Hohn B. Transgenic plants are sensitive bioindicators of nuclear pollution caused by the Chernobyl accident. Nat. Biotechnol. 1998;16:1054–1057. doi: 10.1038/3505. [DOI] [PubMed] [Google Scholar]
- 47.Kovalchuk O., Kovalchuk I., Titov V., Arkhipov A., Hohn B. Radiation hazard caused by the Chernobyl accident in inhabited areas of Ukraine can be monitored by transgenic plants. Mutat. Res. 1999;446:49–55. doi: 10.1016/s1383-5718(99)00147-3. [DOI] [PubMed] [Google Scholar]
- 48.Ries G., Heller W., Puchta H., Sandermann H., Seidlitz H.K., Hohn B. Elevated UV-B radiation reduces genome stability in plants. Nature. 2000;406:98–101. doi: 10.1038/35017595. [DOI] [PubMed] [Google Scholar]
- 49.Kovalchuk O., Titov V., Hohn B., Kovalchuk I. A sensitive transgenic plant system to detect toxic inorganic compounds in the environment. Nat. Biotechnol. 2001;19:568–72. doi: 10.1038/89327. [DOI] [PubMed] [Google Scholar]
- 50.Besplug J., Filkowski J., Burke P., Kovalchuk I., Kovalchuk O. Atrazine induces homologous recombination but not point mutation in the transgenic plant-based biomonitoring assay. Arch. Environ. Contam. Toxicol. 2004;46:296–300. doi: 10.1007/s00244-003-3075-9. [DOI] [PubMed] [Google Scholar]
- 51.Li L, Jean M, Belzile F. The impact of sequence divergence and DNA mismatch repair on homeologous recombination in Arabidopsis. Plant J. 2006;45:908–16. doi: 10.1111/j.1365-313X.2006.02657.x. [DOI] [PubMed] [Google Scholar]
- 52.Boyko A., Greer M., Kovalchuk I. Acute exposure to UVB has a more profound effect on plant genome stability than chronic exposure. Mutat. Res. 2006;602:100–9. doi: 10.1016/j.mrfmmm.2006.08.008. [DOI] [PubMed] [Google Scholar]
- 53.Van der Auwera G., Baute J., Bauwens M., Peck I., Piette D., Pycke M., Asselman P., Depicker A. Development and application of novel constructs to score C:G-to-T:A transitions and homologous recombination in Arabidopsis. Plant Physiol. 2008;146:22–31. doi: 10.1104/pp.107.105213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Swoboda P., Gal S., Hohn B., Puchta H. Intrachromosomal homologous recombination in whole plants. EMBO J. 1994;13:484–489. doi: 10.1002/j.1460-2075.1994.tb06283.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Puchta H., Swoboda P., Hohn B. Induction of homologous DNA recombination in whole plants. Plant J. 1995;7:203–210. [Google Scholar]
- 56.Kovalchuk O., Kovalchuk I., Arkhipov A., Telyuk P., Hohn B., Kovalchuk L. The Allium cepa chromosome aberration test reliably measures genotoxicity of soils of inhabited areas in the Ukraine contaminated by the Chernobyl accident. Mutat. Res. 1998;415:47–57. doi: 10.1016/s1383-5718(98)00053-9. [DOI] [PubMed] [Google Scholar]
- 57.Kovalchuk O., Arkhipov A., Barylyak I., Karachov I., Titov V., Hohn B., Kovalchuk I. Plants experiencing chronic internal exposure to ionizing radiation exhibit higher frequency of homologous recombination than acutely irradiated plants. Mutat Res. 2000;449:47–56. doi: 10.1016/s0027-5107(00)00029-4. [DOI] [PubMed] [Google Scholar]
- 58.Cupples C.G., Cabrera M., Cruz C., Miller J.H. A set of lacZ mutations in Escherichia coli that allow rapid detection of specific frameshift mutations. Genetics. 1990;125:275–280. doi: 10.1093/genetics/125.2.275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Tran R.K., Henikoff J.G., Zilberman D., Ditt R.F., Jacobsen S.E., Henikoff S. DNA methylation profiling identifies CG methylation clusters in Arabidopsis genes. Curr. Biol. 2005;15:154–159. doi: 10.1016/j.cub.2005.01.008. [DOI] [PubMed] [Google Scholar]
- 60.Koornneef M. Classical mutagenesis in higher plants. In: Gilmartin P.M., Bowler C., editors. Molecular Plant Biology. Vol. 1. Oxford University Press; Oxford, UK: 2002. pp. 1–11. [Google Scholar]
- 61.Greene E.A., Codomo C.A., Taylor N.E., Henikoff J.G., Till B.J., Reynolds S.H., Enns L.C., Burtner C., Johnson J.E., Odden A.R., Comai L., Henikoff S. Spectrum of chemically induced mutations from a large-scale reverse-genetic screen in Arabidopsis. Genetics. 2003;164:731–740. doi: 10.1093/genetics/164.2.731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yoshihara R., Nakane C., Takimoto K. A new system for detecting mutations in Arabidopsis thaliana and the mutational spectra resulting from ethylmethanesulfonate treatment. J. Radiat. Res. (Tokyo) 2006;47:223–228. doi: 10.1269/jrr.0623. [DOI] [PubMed] [Google Scholar]
- 63.Rossman T.G., Molina M., Meyer L., Boone P., Klein C.B., Wang Z., Li F., Lin W.C., Kinney P.L. Performance of 133 compounds in the lambda prophage induction endpoint of the Microscreen assay and a comparison with S. typhimurium mutagenicity and rodent carcino-genicity assays. Mutat. Res. 1991;260:349–367. doi: 10.1016/0165-1218(91)90021-d. [DOI] [PubMed] [Google Scholar]
- 64.Bitton G., Koopman B. Bacterial and enzymatic bioassays for toxicity testing in the environment. Rev. Environ. Con. Tox. 1992;125:1–22. doi: 10.1007/978-1-4612-2890-5_1. [DOI] [PubMed] [Google Scholar]
- 65.Codina J.C., Perez-Garcia A., Romero R., de Vincente A. A comparison of microbial bioassays for the detection of metal toxicity. Arch. Environ. Con. Tox. 1993;25:250–254. doi: 10.1007/BF00212137. [DOI] [PubMed] [Google Scholar]
- 66.Bitton G., Jung K., Koopman B. Evaluation of a microplate assay specific for heavy metal toxicity. Arch. Environ. Contam. Toxicol. 1994;27:25–8. doi: 10.1007/BF00203883. [DOI] [PubMed] [Google Scholar]
- 67.Hartmann A., Speit G. Comparative investigations of the genotoxic effects of metals in the single cells gel (SCG) assay and the sister chromatid exchange (SCE) test. Environ. Mol. Mutagen. 1994;23:299–305. doi: 10.1002/em.2850230407. [DOI] [PubMed] [Google Scholar]
- 68.Rank J., Nielsen M.N. Evaluation of the Allium cepa anaphase-telophase test in relation to genotoxicity screening of industrial wastewater. Mutat. Res. 1994;312:17–24. doi: 10.1016/0165-1161(94)90004-3. [DOI] [PubMed] [Google Scholar]
- 69.Kong I.-C., Bitton G., Koopman B., Jung K.-H. Heavy metal toxicity testing in environmental samples. Rev. Environ. Con. Tox. 1995;125:1–22. doi: 10.1007/978-1-4612-4252-9_5. [DOI] [PubMed] [Google Scholar]
- 70.Steinkellner H., Mun-Sik K., Helma C., Ecker S., Ma T.H., Horak O., Kundi M., Knasmuller S. Genotoxic effects of heavy metals: Comparative investigation with plant bioassays. Environ. Mol. Mutagen. 1998;31:183–191. [PubMed] [Google Scholar]
- 71.Helleday T., Nilsson R., Jenssen D. Arsenic [III] and heavy metal ions induce intrachromosomal homologous recombination in the hprt gene of V79 Chinese Hamster Cells. Environ. Mol. Mutagen. 2000;35:114–122. doi: 10.1002/(sici)1098-2280(2000)35:2<114::aid-em6>3.0.co;2-q. [DOI] [PubMed] [Google Scholar]
- 72.Filkowski J., Besplug J., Burke P., Kovalchuk I., Kovalchuk O. Genotoxicity of 2,4-D and dicamba revealed by transgenic Arabidopsis thaliana plants harboring recombination and point mutation markers. Mutat. Res. 2003;542:23–32. doi: 10.1016/j.mrgentox.2003.07.008. [DOI] [PubMed] [Google Scholar]
- 73.Boyko A., Filkowski J., Kovalchuk I. Homologous recombination in plants is temperature and day-length dependent. Mutat. Res. 2005;572(1-2):73–83. doi: 10.1016/j.mrfmmm.2004.12.011. [DOI] [PubMed] [Google Scholar]
- 74.Kovalchuk I., Kovalchuk O., Kalck V., Boyko V., Filkowski J., Heinlein M., Hohn B. Pathogen-induced systemic plant signal triggers DNA rearrangements. Nature. 2003;423:760–2. doi: 10.1038/nature01683. [DOI] [PubMed] [Google Scholar]
- 75.Boyko A., Kathiria P., Zemp F.J., Yao Y., Pogribny I., Kovalchuk I. Transgenerational changes in the genome stability and methylation in pathogen-infected plants: (virus-induced plant genome instability) Nucleic Acids Res. 2007;35:1714–25. doi: 10.1093/nar/gkm029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Schneeberger R.G., Cullis C.A. Specific DNA alterations associated with the environmental induction of heritable changes in flax. Genetics. 1991;128:619–30. doi: 10.1093/genetics/128.3.619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Cullis C.A., Swami S., Song Y. RAPD polymorphisms detected among the flax genotrophs. Plant Mol. Biol. 1999;41:795–800. doi: 10.1023/a:1006385606163. [DOI] [PubMed] [Google Scholar]
- 78.Molinier J., Ries G., Zipfel C., Hohn B. Transgeneration memory of stress in plants. Nature. 2006;442:1046–9. doi: 10.1038/nature05022. [DOI] [PubMed] [Google Scholar]
- 79.Stewart C.N., Jr. Go with the glow: fluorescent proteins to light transgenic organisms. Trends Biotechnol. 2006;24(4):155–62. doi: 10.1016/j.tibtech.2006.02.002. [DOI] [PubMed] [Google Scholar]
- 80.Ilnytskyy Y., Boyko A., Kovalchuk I. Luciferase-based transgenic recombination assay is more sensitive than beta-glucoronidase-based. Mutat. Res. 2004;559:189–97. doi: 10.1016/j.mrgentox.2004.02.001. [DOI] [PubMed] [Google Scholar]
- 81.Engineer C.B., Fitzsimmons K.C., Schmuke J.J., Dotson S.B., Kranz R.G. Development and evaluation of a Gal4-mediated LUC/GFP/GUS enhancer trap system in Arabidopsis. BMC Plant Biol. 2005;5:9. doi: 10.1186/1471-2229-5-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Koo J., Kim Y., Kim J., Yeom M., Lee I.C., Nam H.G. A GUS/luciferase fusion reporter for plant gene trapping and for assay of promoter activity with luciferin-dependent control of the reporter protein stability. Plant Cell Physiol. 2007;48:1121–31. doi: 10.1093/pcp/pcm081. [DOI] [PubMed] [Google Scholar]
- 83.Stewart C.N., Jr. The utility of green fluorescent protein in transgenic plants. Plant Cell Rep. 2001;20:376–82. doi: 10.1007/s002990100346. [DOI] [PubMed] [Google Scholar]
- 84.Stewart C.N., Jr. Monitoring the presence and expression of transgenes in living plants. Trends Plant Sci. 2005;10:390–6. doi: 10.1016/j.tplants.2005.06.003. [DOI] [PubMed] [Google Scholar]
- 85.Azaiez A., Bouchard E.F., Jean M., Belzile F.J. Length, orientation, and plant host influence the mutation frequency in microsatellites. Genome. 2006;49:1366–73. doi: 10.1139/g06-099. [DOI] [PubMed] [Google Scholar]
- 86.Padidam M. Chemically regulated gene expression in plants. Curr. Opin. Plant Biol. 2003;6:169–77. doi: 10.1016/s1369-5266(03)00005-0. [DOI] [PubMed] [Google Scholar]
- 87.Ma S., Bohnert H.J. Integration of Arabidopsis thaliana stress-related transcript profiles, promoter structures, and cell-specific expression. Genome Biol. 2007;8:R49. doi: 10.1186/gb-2007-8-4-r49. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Saidi Y., Domini M., Choy F., Zryd J.P., Schwitzguebel J.P., Goloubinoff P. Activation of the heat shock response in plants by chlorophenols: transgenic Physcomitrella patens as a sensitive biosensor for organic pollutants. Plant Cell Environ. 2007;30:753–63. doi: 10.1111/j.1365-3040.2007.01664.x. [DOI] [PubMed] [Google Scholar]
- 89.Kovalchuk O., Burke P., Arkhipov A., Kuchma N., James S.J., Kovalchuk I., Pogribny I. Genome hypermethylation in Pinus silvestris of Chernobyl--a mechanism for radiation adaptation? Mutat. Res. 2003;529:13–20. doi: 10.1016/s0027-5107(03)00103-9. [DOI] [PubMed] [Google Scholar]
- 90.Amedeo P., Habu Y., Afsar K., Mittelsten Scheid O., Paszkowski J. Disruption of the plant gene MOM releases transcriptional silencing of methylated genes. Nature. 2000;405:203–206. doi: 10.1038/35012108. [DOI] [PubMed] [Google Scholar]