

To the Editor
Germline aberrations in the CDKN2A gene are observed in some melanoma-prone families and represent high penetrance mutations (Hussussian et al., 1994; Kamb et al., 1994). INK4A (p16) and ARF (p14) are two distinct proteins encoded by the CDKN2A locus. Loss of INK4A has been associated with unrestricted cell cycle progression through retinoblastoma (RB) protein inactivation, while loss of ARF has been linked to p53 inactivation with subsequent malfunction in cell cycle regulation, apoptosis and DNA repair (Chin et al., 2006).
The NRAS and BRAF genes are commonly mutated in sporadic primary cutaneous melanomas, with mutation frequencies between 4–50% (Platz et al., 2008) and 25–80% (Platz et al., 2008), respectively. Codon 61 is the most common position of NRAS alterations in melanoma, with frequent glutamine changes to either lysine, Q61K (c.181C>A), or arginine, Q61R (c.182A>G) (Omholt et al., 2002). Mutations in this residue lock the Ras protein in the GTP-bound state with subsequent continuous activation of its downstream effectors (Platz et al., 2008) through the Ras-Raf-MEK-ERK and Ras-PI3K-Akt pathways. Approximately 90% of reported BRAF mutations occur at residue 600, which is located in the activation domain of this kinase (Thomas, 2006). Current results from melanoma cohorts show that mutations in these genes are almost always mutually exclusive (Edlundh-Rose et al., 2006; Omholt et al., 2003; Platz et al., 2008). Moreover, a high rate of BRAF mutation is found also in nevi, suggesting a role in early stages of the neoplastic process (Pollock et al., 2003). Previously, a high frequency of NRAS mutations (95%) has been reported in Swedish familial melanoma cases with germline CDKN2A alterations (Eskandarpour et al., 2003). The association of BRAF somatic mutations with MC1R germline variants indicates an influence of constitutive genotype on the preferential acquisition of specific mutations during melanoma development (Landi et al., 2006). In line with this, cooperation between RAS and CDKN2A has been shown in animal models of melanoma (Chin et al., 1997).
Since there is limited information on NRAS and BRAF mutations in familial melanoma we sought to assess their mutation frequency in melanomas from patients with different CDKN2A germline alterations.
The study was performed on formalin-fixed, paraffin-embedded (FFPE) primary familial cutaneous melanomas originating from Brisbane, Australia (16 samples from 15 patients) and Genoa, Italy (3 patients/samples). Clinical and pathological characteristics are shown in Table 1. One patient with a CDKN2A L32P mutation had two melanomas originating from the trunk and upper extremity, respectively, while a single melanoma was analyzed from each of the other patients. Eight different germline CDKN2A mutations were present in the patients from whom the melanomas were analyzed (Table 2). Laser capture microdissection, DNA extraction, polymerase chain reaction (PCR) amplification, single strand conformation polymorphism (SSCP) and nucleotide sequence analyses of NRAS exon 2 and BRAF exon 15 were carried out as previously described (Jovanovic et al., 2008; Omholt et al., 2002; Omholt et al., 2003). Each mutation was confirmed by two independent PCR/SSCP analyses followed by sequence analysis performed in both directions. The study was approved by the Ethics Committees of the Queensland Institute of Medical Research, University of Genoa and Karolinska Institutet.
Table 1.
Patient and tumor characteristics
| Brisbane | Genoa | Total | ||
|---|---|---|---|---|
| N | N | N | ||
| Number of patients | 15 | 3 | 18 | |
| Number of tumors | 16 | 3 | 19 | |
| Gender | ||||
| Male | 8 | 2 | 10 | |
| Female | 7 | 1 | 8 | |
| Median age at diagnosis (range) | 36 (19–56) | 37 (20–41) | 36 (19–56) | |
| Site | ||||
| Head and neck | 0 | 1 | 1 | |
| Trunk | 6 | 1 | 7 | |
| Upper extremities | 6 | 1 | 7 | |
| Lower extremities | 4 | 0 | 4 | |
| Histology | ||||
| Superficial spreading melanoma | 10 | 2 | 12 | |
| Nodular melanoma | 0 | 1 | 1 | |
| Unclassifiable melanoma | 6 | 0 | 6 | |
| Clark | ||||
| I | 6 | 1 | 7 | |
| II | 4 | 1 | 5 | |
| III | 2 | 1 | 3 | |
| Unknown | 4 | 0 | 4 | |
| Mean tumor thickness in mm (SD*) | 0.61 (0.63) | 1.71 | 0.81 (0.92) | |
| Ulceration | ||||
| Present | 0 | 0 | 0 | |
| Absent | 3 | 3 | 6 | |
| Unknown | 13 | 0 | 13 | |
| Median survival in months (range) | 372+ (72–942+) | 389+ (257–451+) | 373+ (72–942+) | |
| Alive | 12 | 3 | 15 | |
| Dead | 3 | 0 | 3 | |
Standard deviation
Table 2.
NRAS and BRAF genotypes in familial melanomas harboring CDKN2A germline mutations
| Tumor number |
CDKN2A mutation | NRAS mutation | BRAF mutation | |||
|---|---|---|---|---|---|---|
| Protein change | Base change | Protein change | Base change | Protein change | Base change | |
| 1 | p.A4_P11del | c.9_32del24 | WT‡ | none | p.V600E | c.1799T>A |
| 2 | p.A4_P11dup | c.32_33dup9-32 | WT | none | WT | none |
| 3 | p.A4_P11 dup | c.32_33 dup9-32 | WT | none | WT | none |
| 4 | p.A4_P11 dup | c.32_33 dup9-32 | WT | none | WT | none |
| 5 | p.L16P | c.47T>C | WT | none | WT | none |
| 6 | p.L16P | c.47T>C | WT | none | WT | none |
| 7 | p.R24P | c.71G>C | WT | none | WT | none |
| 8 | p.R24P | c.71G>C | WT | none | p.V600E | c.1799T>A |
| 9 | p.R24P | c.71G>C | WT | none | p.V600E | c.1799T>A |
| 10 | p.L32P | c.95T>C | p.Q61K | c.181C>A | p.V600E | c.1799T>A |
| 11 | p.L32P | c.95T>C | p.Q61R | c.182A>G | p.V600E | c.1799T>A |
| 12 | p.L32P* | c.95T>C | p.Q61K | c.181C>A | p.V600E | c.1799T>A |
| 13 | p.L32P* | c.95T>C | WT | none | WT | none |
| 14 | p.G35A | c.104G>C | WT | none | WT | none |
| 15 | p.G35A | c.104G>C | WT | none | p.V600E | c.1799T>A |
| 16 | p.M53I | c.159G>C | WT | none | WT | none |
| 17 | p.G101W† | c.301G>T | WT | none | WT | none |
| 18 | p.G101W† | c.301G>T | WT | none | WT | none |
| 19 | p.G101W† | c.301G>T | WT | none | WT | none |
| Total: 19 | 3 (16%) | 7 (37%) | ||||
Samples originating from the same patient
Samples originating from Genoa
Wild type
Three (16%) samples had NRAS residue 61 alterations (one Q61R and two Q61K substitutions) while 7 (37%) samples had valine to glutamic acid changes in amino acid 600 of BRAF (V600E; Table 2).
The NRAS mutation frequency in this study was lower than we reported previously in melanomas from Swedish families with germline CDKN2A mutations (Eskandarpour et al., 2003). The reason for this is unclear. It could possibly be attributed to different origins of studied cohorts (Sweden versus Australia/Italy) and to different CDKN2A germline alterations in these two studies (the 112Argdup founder mutation is predominant in Swedish families, which is in contrast to genotypes reported here). However, we cannot exclude that technical factors, possibly related to the fragmented nature of DNA extracted from FFPE samples, may play a role in different rates of mutation detection.
Intriguingly, all 3 tumors with NRAS mutations also had BRAF V600E mutations. The presence of both NRAS and BRAF V600E mutations in the same lesions is contrary to the current consensus that such mutations are almost always mutually exclusive in melanomas and other tumor types (Davies et al., 2002; Omholt et al., 2003; Thomas et al., 2007). However, Pollock et al (2003) observed concomitant NRAS and BRAF V600E mutations in 9% of nevi and suggested this might be due to different clonal nests of cells within these tumors carrying distinct mutations, a possibility that could also explain our findings. Although intriguingly, the joint presence of both NRAS and BRAF mutations was found only in tumors from patients with CDKN2A L32P mutations. In two of these tumors, NRAS and BRAF mutations (NRAS/BRAF: Q61K/V600E and Q61R/V600E) were found in two different DNA extracts while in one case, alterations of both genes (NRAS/BRAF: Q61K/V600E) were identified in the same DNA extract. Thus far, it is recognized that a mutation in either NRAS or BRAF is sufficient for activation of the Ras-Raf-MEK-ERK pathway, with mutant RAS having a 50-fold higher activation effect than mutant BRAF (Davies et al., 2002). Although we do not have any evidence that the NRAS and BRAF mutations found in the same DNA extract were coexisting in the same cells, it is possible that the L32P mutation in CDKN2A somehow permits cellular tolerance of these dual mutations.
In conclusion, the NRAS and BRAF mutation rates we observed in familial melanomas were generally lower than most previous reports in sporadic melanoma but equal to those reported for primary melanomas of similar thickness (Goel et al., 2006; Shinozaki et al., 2004). Samples that harbored INK4A L32P substitutions also had high frequency of coexisting mutations in both NRAS and BRAF. This suggests that in some instances constitutional CDKN2A mutations affect the occurrence of somatic mutations in NRAS and BRAF, although further work is needed to substantiate this hypothesis.
Supplementary Material
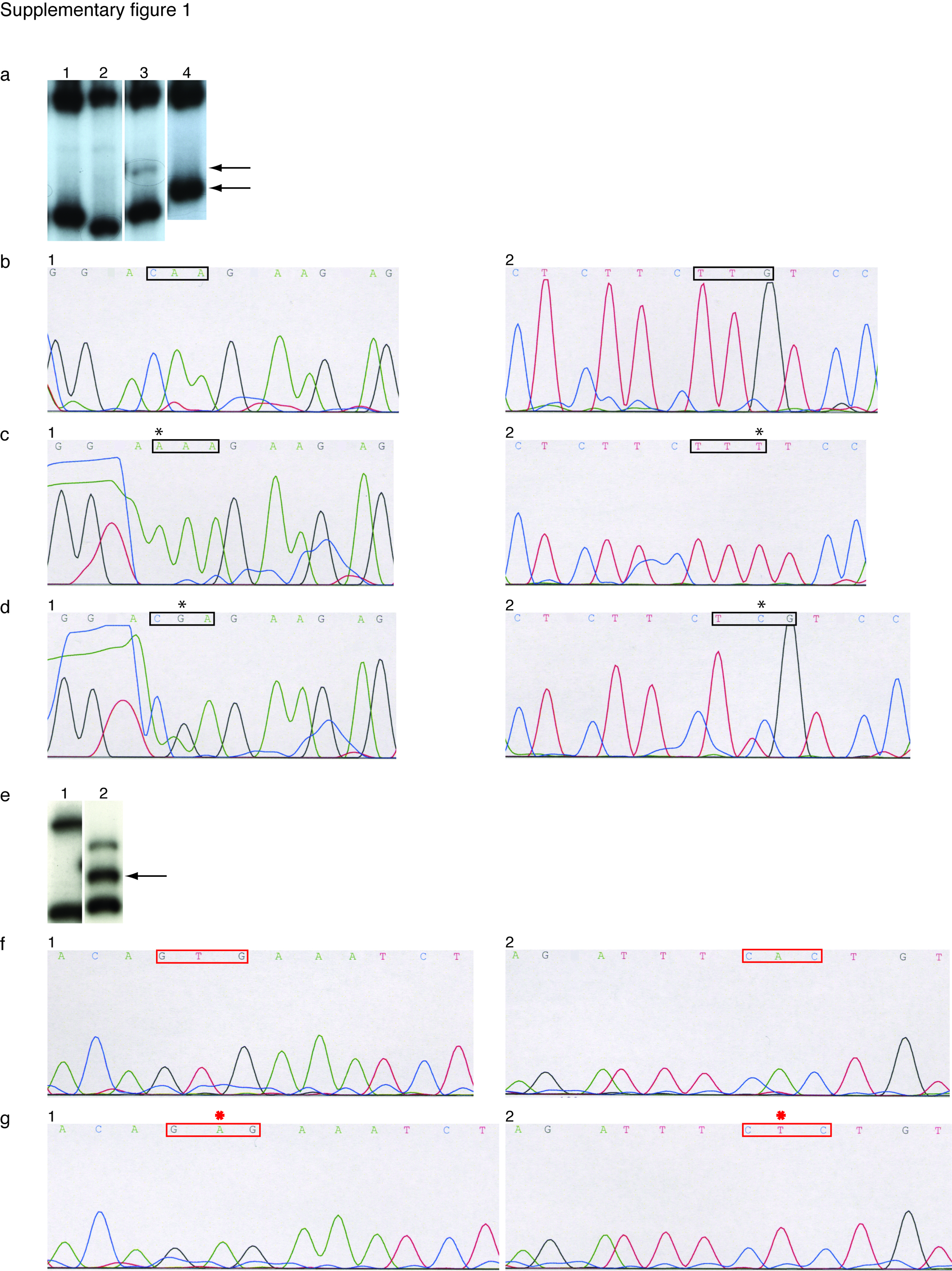
Supplementary figure 1. NRAS exon 2 (a–d) and BRAF exon 15 (e–g) mutation analyses. Panel a: SSCP lanes 1–2, wild type samples; lane 3, Q61K (c.181C>A) mutation sample; lane 4, Q61R (c.182A>G) mutation sample. Panel b, wild type sequencing graphs (codons 59–62; 1-forward sequence, 2-reverse sequence); panel c, sequencing graphs of Q61K (c.181C>A) mutation (1-forward, 2-reverse); panel d, sequencing graphs of Q61R (c.182A>G) mutation (1-forward, 2-reverse). Panel e: SSCP lane 1, wild type sample; lane 2: V600E mutation sample. Panel f, wild type sequencing graphs (codons 599–602; 1-forward, 2-reverse); panel g, sequencing graphs of V600E (c.1799T>A) mutation (1-forward, 2-reverse). Arrows indicate bands with abnormal mobility, signifying mutations. Black rectangulares indicate localization of the NRAS codon 61, with black asterisk showing base change. Red colored rectangulares indicate localization of the BRAF codon 600, with red asterisk showing base change.
{kind=link}
Acknowledgements
The Cancer Society of Stockholm, the King Gustav V Jubilee Fund, the Karolinska Institute Research Fund, the Swedish Cancer Society, the National Health and Medical Research Council of Australia and Italian Regional Grant supported this work.
The data on cell lines were obtained through web sites from the Wellcome Trust Sanger Institute Cancer Genome Project (http://www.sanger.ac.uk/genetics/CGP/) and the European Searchable Tumor Cell Line Data Base (ESTDAB) project (http://www.ebi.ac.uk/ipd/estdab/). The authors wish to thank Per Guldberg for melanoma cell line information as well as Joseph Lawrence for help with sequencing.
Footnotes
Conflict of Interest
The Authors state no conflict of interest.
References
- Chin L, Garraway LA, Fisher DE. Malignant melanoma: genetics and therapeutics in the genomic era. Genes Dev. 2006;20:2149–2182. doi: 10.1101/gad.1437206. [DOI] [PubMed] [Google Scholar]
- Chin L, Pomerantz J, Polsky D, Jacobson M, Cohen C, Cordon-Cardo C, et al. Cooperative effects of INK4a and ras in melanoma susceptibility in vivo. Genes Dev. 1997;11:2822–2834. doi: 10.1101/gad.11.21.2822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chong H, Lee J, Guan KL. Positive and negative regulation of Raf kinase activity and function by phosphorylation. EMBO J. 2001;20:3716–3727. doi: 10.1093/emboj/20.14.3716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies H, Bignell GR, Cox C, Stephens P, Edkins S, Clegg S, et al. Mutations of the BRAF gene in human cancer. Nature. 2002;417:949–954. doi: 10.1038/nature00766. [DOI] [PubMed] [Google Scholar]
- Edlundh-Rose E, Egyhazi S, Omholt K, Mansson-Brahme E, Platz A, Hansson J, et al. NRAS and BRAF mutations in melanoma tumours in relation to clinical characteristics: a study based on mutation screening by pyrosequencing. Melanoma Res. 2006;16:471–478. doi: 10.1097/01.cmr.0000232300.22032.86. [DOI] [PubMed] [Google Scholar]
- Eskandarpour M, Hashemi J, Kanter L, Ringborg U, Platz A, Hansson J. Frequency of UV-inducible NRAS mutations in melanomas of patients with germline CDKN2A mutations. J Natl Cancer Inst. 2003;95:790–798. doi: 10.1093/jnci/95.11.790. [DOI] [PubMed] [Google Scholar]
- Goel VK, Lazar AJ, Warneke CL, Redston MS, Haluska FG. Examination of mutations in BRAF, NRAS, and PTEN in primary cutaneous melanoma. J Invest Dermatol. 2006;126:154–160. doi: 10.1038/sj.jid.5700026. [DOI] [PubMed] [Google Scholar]
- Houben R, Becker JC, Kappel A, Terheyden P, Brocker EB, Goetz R, et al. Constitutive activation of the Ras-Raf signaling pathway in metastatic melanoma is associated with poor prognosis. J Carcinog. 2004;3:6. doi: 10.1186/1477-3163-3-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hussussian CJ, Struewing JP, Goldstein AM, Higgins PA, Ally DS, Sheahan MD, et al. Germline p16 mutations in familial melanoma. Nat Genet. 1994;8:15–21. doi: 10.1038/ng0994-15. [DOI] [PubMed] [Google Scholar]
- Jovanovic B, Krockel D, Linden D, Nilsson B, Egyhazi S, Hansson J. Lack of cytoplasmic ERK activation is an independent adverse prognostic factor in primary cutaneous melanoma. J Invest Dermatol. 2008;128:2696–2704. doi: 10.1038/jid.2008.134. [DOI] [PubMed] [Google Scholar]
- Kamb A, Shattuck-Eidens D, Eeles R, Liu Q, Gruis NA, Ding W, et al. Analysis of the p16 gene (CDKN2) as a candidate for the chromosome 9p melanoma susceptibility locus. Nat Genet. 1994;8:23–26. doi: 10.1038/ng0994-22. [DOI] [PubMed] [Google Scholar]
- Landi MT, Bauer J, Pfeiffer RM, Elder DE, Hulley B, Minghetti P, et al. MC1R germline variants confer risk for BRAF-mutant melanoma. Science. 2006;313:521–522. doi: 10.1126/science.1127515. [DOI] [PubMed] [Google Scholar]
- Lorenzi PL, Reinhold WC, Varma S, Hutchinson AA, Pommier Y, Chanock SJ, et al. DNA fingerprinting of the NCI-60 cell line panel. Mol Cancer Ther. 2009;8:713–724. doi: 10.1158/1535-7163.MCT-08-0921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Omholt K, Karsberg S, Platz A, Kanter L, Ringborg U, Hansson J. Screening of N-ras codon 61 mutations in paired primary and metastatic cutaneous melanomas: mutations occur early and persist throughout tumor progression. Clin Cancer Res. 2002;8:3468–3474. [PubMed] [Google Scholar]
- Omholt K, Platz A, Kanter L, Ringborg U, Hansson J. NRAS and BRAF mutations arise early during melanoma pathogenesis and are preserved throughout tumor progression. Clin Cancer Res. 2003;9:6483–6488. [PubMed] [Google Scholar]
- Platz A, Egyhazi S, Ringborg U, Hansson J. Human cutaneous melanoma; a review of NRAS and BRAF mutation frequencies in relation to histogenetic subclass and body site. Molecular Oncology. 2008;1:395–405. doi: 10.1016/j.molonc.2007.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollock PM, Harper UL, Hansen KS, Yudt LM, Stark M, Robbins CM, et al. High frequency of BRAF mutations in nevi. Nat Genet. 2003;33:19–20. doi: 10.1038/ng1054. [DOI] [PubMed] [Google Scholar]
- Robinson J, Roberts CH, Dodi IA, Madrigal JA, Pawelec G, Wedel L, et al. The European searchable tumour line database. Cancer Immunol Immunother. 2009 doi: 10.1007/s00262-008-0656-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shinozaki M, Fujimoto A, Morton DL, Hoon DS. Incidence of BRAF oncogene mutation and clinical relevance for primary cutaneous melanomas. Clin Cancer Res. 2004;10:1753–1757. doi: 10.1158/1078-0432.ccr-1169-3. [DOI] [PubMed] [Google Scholar]
- Thomas NE. BRAF somatic mutations in malignant melanoma and melanocytic naevi. Melanoma Res. 2006;16:97–103. doi: 10.1097/01.cmr.0000215035.38436.87. [DOI] [PubMed] [Google Scholar]
- Thomas NE, Edmiston SN, Alexander A, Millikan RC, Groben PA, Hao H, et al. Number of nevi and early-life ambient UV exposure are associated with BRAF-mutant melanoma. Cancer Epidemiol Biomarkers Prev. 2007;16:991–997. doi: 10.1158/1055-9965.EPI-06-1038. [DOI] [PubMed] [Google Scholar]
- Wan PT, Garnett MJ, Roe SM, Lee S, Niculescu-Duvaz D, Good VM, et al. Mechanism of activation of the RAF-ERK signaling pathway by oncogenic mutations of B-RAF. Cell. 2004;116:855–867. doi: 10.1016/s0092-8674(04)00215-6. [DOI] [PubMed] [Google Scholar]
- Zhang BH, Guan KL. Activation of B-Raf kinase requires phosphorylation of the conserved residues Thr598 and Ser601. EMBO J. 2000;19:5429–5439. doi: 10.1093/emboj/19.20.5429. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary figure 1. NRAS exon 2 (a–d) and BRAF exon 15 (e–g) mutation analyses. Panel a: SSCP lanes 1–2, wild type samples; lane 3, Q61K (c.181C>A) mutation sample; lane 4, Q61R (c.182A>G) mutation sample. Panel b, wild type sequencing graphs (codons 59–62; 1-forward sequence, 2-reverse sequence); panel c, sequencing graphs of Q61K (c.181C>A) mutation (1-forward, 2-reverse); panel d, sequencing graphs of Q61R (c.182A>G) mutation (1-forward, 2-reverse). Panel e: SSCP lane 1, wild type sample; lane 2: V600E mutation sample. Panel f, wild type sequencing graphs (codons 599–602; 1-forward, 2-reverse); panel g, sequencing graphs of V600E (c.1799T>A) mutation (1-forward, 2-reverse). Arrows indicate bands with abnormal mobility, signifying mutations. Black rectangulares indicate localization of the NRAS codon 61, with black asterisk showing base change. Red colored rectangulares indicate localization of the BRAF codon 600, with red asterisk showing base change.