

Abstract
Drosophila melanogaster is a valuable model organism to study aging and pathological degenerative processes in the nervous system. The advantages of the fly as an experimental system include its genetic tractability, short life span and the possibility to observe and quantitatively analyze complex behaviors. The expression of disease-linked genes in specific neuronal populations of the Drosophila brain, can be used to model human neurodegenerative diseases such as Parkinson's and Alzheimer's 5.
Dopaminergic (DA) neurons are among the most vulnerable neuronal populations in the aging human brain. In Parkinson's disease (PD), the most common neurodegenerative movement disorder, the accelerated loss of DA neurons leads to a progressive and irreversible decline in locomotor function. In addition to age and exposure to environmental toxins, loss of DA neurons is exacerbated by specific mutations in the coding or promoter regions of several genes. The identification of such PD-associated alleles provides the experimental basis for the use of Drosophila as a model to study neurodegeneration of DA neurons in vivo. For example, the expression of the PD-linked human α-synuclein gene in Drosophila DA neurons recapitulates some features of the human disease, e.g. progressive loss of DA neurons and declining locomotor function 2. Accordingly, this model has been successfully used to identify potential therapeutic targets in PD 8.
Here we describe two assays that have commonly been used to study age-dependent neurodegeneration of DA neurons in Drosophila: a climbing assay based on the startle-induced negative geotaxis response and tyrosine hydroxylase immunostaining of whole adult brain mounts to monitor the number of DA neurons at different ages. In both cases, in vivo expression of UAS transgenes specifically in DA neurons can be achieved by using a tyrosine hydroxylase (TH) promoter-Gal4 driver line 3, 10.
Keywords: Neuroscience, Issue 74, Genetics, Neurobiology, Molecular Biology, Cellular Biology, Biomedical Engineering, Medicine, Developmental Biology, Drosophila melanogaster, neurodegenerative diseases, negative geotaxis, tyrosine hydroxylase, dopaminergic neuron, α-synuclein, neurons, immunostaining, animal model
Introduction
The specificity of the assays described here relies on the use of a Gal4 fly line which exploits the regulatory sequences of the tyrosine hydroxylase gene to achieve specific expression in dopaminergic neurons 3. Tyrosine hydroxylase catalyses the first and rate-limiting step in dopamine synthesis. TH immunoreactivity in the adult fly brain overlaps with that of dopamine, making TH a good candidate to identify DA neurons in vivo (see for example 6). Moreover, the expression pattern of THGal4 is more specific than that of other Gal4 lines such as DdcGal4, which contains regulatory sequences from the dopa decarboxylase gene and drives transgenic expression not only in DA neurons, but also in serotoninergic neurons and subsets of glial cells 3.
Feany and Bender (2000) first observed that pan-neuronal expression of the PD-linked human α-synuclein gene accelerates the progressive loss of startle-induced negative geotaxis behavior in Drosophila. We have observed similar results using the DA-neuron specific THGal4 line to drive the expressionof an α-synuclein transgene 10 and used this functional read-out to study the neuroprotective role of the Nrf2 pathway in this Drosophila model of PD 14. The specific assay described here is adapted from 2 and 3.
The Drosophila brain contains more than 100 DA neurons, arranged in at least 5 different clusters (PPL1, PPL2, PAL, PPM1/2, PPM3) which can be visualized in whole brain mounts by confocal microscopy (see for example 11 and 7). It is still controversial whether or not the number of DA neurons in the Drosophila brain declines with age 9, 11; however, age-dependent neurodegeneration is observed in flies where PD-linked genes have been mutated/overexpressed to model human diseases 5. The expression of human α-synuclein in DA neurons has such an effect and, hence, has been used as a model for PD. Here we describe an assay for DA neurons count in whole brain mounts using TH as a cell marker. This assay has been adapted from 13 and 10.
Protocol
1. Startle-induced Negative Geotaxis Assay
Select flies of the desired genotype, separate them by gender, randomly group them in cohorts of 20-30 animals and flip them into new food vials every 2-3 days; test each set of flies (i.e. age-matched cohorts, one cohort/genotype) at least once a week.
Thirty minutes before the negative geotaxis test, anesthetize the flies with CO2 and place them in pre-labeled clear plastic tubes made with two empty food vials (see Materials Table) joined at their openings with clear tape (chamber dimensions: 19 cm x 2.85 cm). In our assays we typically use 25 flies/tube. Note. The recovery time from anesthesia can be varied from few minutes to several hours (e.g. O/N) and should be optimized for the specific genotypes tested.
Mark different heights (from 1 to 20 cm) on a piece of blotting paper and tape the paper on a vertical surface, perpendicular to the bench.
Place the tubes in front of the paper: the tubes should be all aligned and as close as possible to the paper to allow accurate height measurement.
Place the digital camera (see the "Special Equipment" Table) in front of the tubes, at a distance of 20-30 cm from the paper, on a stand (e.g. a pipette tip box), select the "movie" option and start recording; at this time you should also start a timer to keep track of the time between consecutive trials.
Allow the flies to collect at the bottom of the tube by tapping the tube for few seconds (e.g. 10 sec). This step should be performed consistently (i.e. same duration, same strength, same operator) throughout the whole experiment.
Record flies' movements with the digital camera for 10 sec.
Repeat steps 1.5-1.7 fourteen times at one-min intervals.
- Analyze data:
- Count the number of flies that are above the 2 cm mark after 10 sec by visual inspection of the recorded movies. Note: In our assay conditions the flies' negative geotaxis behavior didn't last beyond the first 10 sec after startling (i.e. flies started moving in all directions 10 sec after startling them). The minimum distance climbed by control flies (i.e. 1 week old flies expressing the THGal4 driver, see legend of Figure 1) in this time window was 2 cm. Thus, we used the 2 cm mark as a reference for analyzing the climbing activities of flies of different ages and genotypes 10 sec after initiating the behavior. These parameters may require adjusting to account for differences in behavior of the flies tested (due for example to different genetic backgrounds or laboratory conditions). Notice that in our assay conditions, all the genotypes tested performed worse or similarly to the flies of the control genotype.
- Take the average of the results from the fifteen trials.
- Express average as a percentage of the total number of flies in the tube (= % climbing activity); the number of repeats (N) is given by the number of independent cohorts tested for each genotype
- Assess statistical significance between different genotypes over time. This can be accomplished with a Student's t test (when comparing two genotypes) or a 2-way ANOVA analysis followed by Bonferroni post-hoc test (when performing multiple comparisons) using commercial softwares such as GraphPad (GraphPad Software, Inc.La Jolla, CA, USA).
Note
We perform all our assays at the same time of the day, at RT and under the same light conditions. Keeping the flies in a 12 hr light/dark cycle environment is advisable to control for the effect of circadian rhythms on the flies' locomotor behavior.
2. Tyrosine Hydroxylase Immunofluorescence Assay of Whole Brain Mounts
Solutions and buffers
Fixative solution: 4% Paraformaldehyde (PFA) in phosphate buffered saline (PBS)
Washing buffer: 0.1% Triton X-100 in PBS
Blocking buffer: 0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, 0.1% Triton X-100 and 0.5% BSA
Mounting media: Mowiol-Dabco (see protocol described in Harlow E, Lane D., Antibodies- A laboratory manual, Cold Spring Harbor Laboratories Press, Cold Spring Harbor, NY, USA, 10:418 (1988))
Procedure
Put flies in a dissection dish filled with 70% EtOH for about 1 min to remove the cuticle wax.
Transfer flies to another dissection dish filled with ice-cold PBS.
- Dissect the brain:
- Place the fly with the ventral side up (optional: remove wings and legs).
- Pull the head away from the body: use one set of forceps to hold on to the body and another one to hold on to the cuticle under the eye to prevent damaging the brain
- Remove the proboscis.
- Hold on to the head cuticle on opposite sides of the slit left open after removing the proboscis and pull in opposite directions until the brain is removed from the head capsule.
- Remove all cuticle from the brain.
- Remove all the air-filled tracheal tissue; this will prevent the brain from floating during the following incubation steps.
Cut a few millimeters off the tip of a P-200 pipette tip and equilibrate it with a 0.1% Triton X-100/PBS solution
Using the prepared P-200 pipette tip, transfer the brains to a 0.5 ml Eppendorf tube filled with 0.5 ml of 4% PFA/PBS and keep on ice until the dissection is complete.
Fix the brains at RT for 20 min: apply a gentle rotation by putting the Eppendorf tubes on a rack and placing the rack on a nutator.
Remove the fixative using a P-1000 micro pipette, add 0.5 ml of 0.1% Triton X-100/PBS, mix by inversion and let incubate for 1 min; repeat this washing step once.
Remove the buffer from the second wash, add 0.5 ml of 0.1% Triton X-100/PBS and let incubate at RT for 20 min; repeat this step twice.
Remove the washing buffer and let the brains incubate in blocking buffer (0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, 0.1% Triton X-100 and 0.5% BSA) for 1 hr at RT.
Incubate the brains with a 1:100 dilution of anti-TH antibody (see the "Table of specific reagents") in blocking solution, for two days, at 4 °C, with gentle rotation (see step 2.6.).
Repeat washing steps 2.7. and 2.8..
Equilibrate the brains with a 1:200 dilution of secondary antibodyin blocking solution, for two days, at 4 °C, with gentle rotation.
Repeat washing steps 2.7. and 2.8..
- Prepare the mounting slides as illustrated in Figure 2A:
- Add 2 X 25 μl drops of mounting media to a cover glass (24 x 60 mm, no. 1.5).
- Place two cover glasses (size 18 x 18 mm, no. 2) on the mounting media leaving a gap in the middle of the slide and let dry.
Remove the washing buffer, add 50 μl of mounting media and allow the brains to equilibrate with it by pipetting up and down.
Using a cut P-200 pipette tip (see step 2.4) transfer the brains to the slide in the space between the two coverslips and cover them with a coverslip no. 1.5 (see Figure 2A; to orient the brains on the slide see 7). Note: the samples are mounted between two cover glasses to allow visualization of both the anterior and the posterior side of the brain.
Fill the whole space between the cover glasses with mounting media; pipette from one corner to remove all the air; let dry and seal with nail polish.
To assess the number of dopaminergic neurons in a specific cluster acquire a series of optical sections along the z-axis with a 1.0 μm step-size using a confocal microscope (for the anatomical identification of dopaminergic clusters in the Drosophila adult brain see 7).
Representative Results
We have used the assays described here to study the role of the stress protective Nrf2 pathway in a Drosophila model of PD 14. This model relies on the expression of the human α-synuclein gene in DA neurons using a TH Gal4 driver 2, 10.
Figure 1 shows a representative result of a startle-induced negative geotaxis (climbing) assay in male flies expressing different transgenes under the control of the TH Gal4 driver. All the genotypes tested exhibit a decline in locomotor activity over time; however, flies expressing the PD-linked, human α-synuclein transgene exhibit an accelerated decline relative to age-matched control flies (TH Gal4 driver alone) or flies co-expressing the Nrf2 DNA binding partner Maf-S. Similar results were observed in female flies.
Figure 2 illustrates a representative result for a TH immunofluorescence-based DA neuron count. The effect of different genetic backgrounds on the number of PPL1 neurons in brains from 4 week old male flies is quantified as described in the figure legend. Notice the small but significant loss of PPL1 neurons in brains from flies expressing α-synuclein.
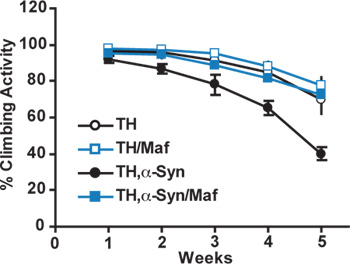 Figure 1. Startle-induced negative geotaxis assay. Cohorts of age-matched flies of the indicated genotypes were tested for locomotor activity at one-week intervals. Climbing activity was calculated by counting the number of flies above the 2 cm mark 10 sec after tapping the vial and expressing this value as a percentage of the total number of flies contained in the vial. Data show means ± s.e.m. of five independent cohorts of 20-30 flies. Significance was assessed with a two-way ANOVA with Bonferroni post-hoc test (P<0.05). The difference between TH,α-Synflies and flies of the other three genotypes tested was significant at 4 and 5 weeks.
Figure 1. Startle-induced negative geotaxis assay. Cohorts of age-matched flies of the indicated genotypes were tested for locomotor activity at one-week intervals. Climbing activity was calculated by counting the number of flies above the 2 cm mark 10 sec after tapping the vial and expressing this value as a percentage of the total number of flies contained in the vial. Data show means ± s.e.m. of five independent cohorts of 20-30 flies. Significance was assessed with a two-way ANOVA with Bonferroni post-hoc test (P<0.05). The difference between TH,α-Synflies and flies of the other three genotypes tested was significant at 4 and 5 weeks.
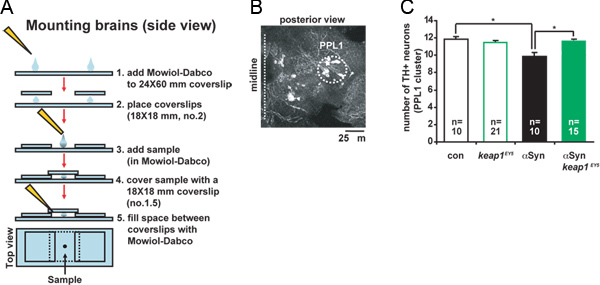 Figure 2. Detection and count of DA neurons by TH immunofluorescence.A. Flow diagram illustrating the protocol for preparing brain slides. For a more detailed description of each step see text. B. Representative micrograph of a whole brain mount (right hemibrain, posterior view) immunostained for TH. A set of confocal z-series images was acquired with a Leica SP2 confocal microscope, 40X oil objective (N/A 1.25), frame average 2, step-size 1 μm, in a 1,024 x 1,024 format and compiled as a maximum projection (shown). The position of the PPL1 DA neuron cluster is highlighted. C. Representative results for DA neuron counts obtained using TH immunostaining. The number of neurons in the PPL1 cluster was assessed by inspecting the individual images of each confocal z-series. N represents the number of independent samples analyzed for each genotype. Data are means ± s.e.m. Significance was assessed with the Student's t test (*P<0.05). Click here to view larger figure.
Figure 2. Detection and count of DA neurons by TH immunofluorescence.A. Flow diagram illustrating the protocol for preparing brain slides. For a more detailed description of each step see text. B. Representative micrograph of a whole brain mount (right hemibrain, posterior view) immunostained for TH. A set of confocal z-series images was acquired with a Leica SP2 confocal microscope, 40X oil objective (N/A 1.25), frame average 2, step-size 1 μm, in a 1,024 x 1,024 format and compiled as a maximum projection (shown). The position of the PPL1 DA neuron cluster is highlighted. C. Representative results for DA neuron counts obtained using TH immunostaining. The number of neurons in the PPL1 cluster was assessed by inspecting the individual images of each confocal z-series. N represents the number of independent samples analyzed for each genotype. Data are means ± s.e.m. Significance was assessed with the Student's t test (*P<0.05). Click here to view larger figure.
Genotypes: 'con', THGal4/+;THGal4/+; 'UAS-Syn', THGal4, UAS -αSynuclein/+;THGal4,UAS -αSynuclein/+; 'UAS-Maf-S', THGal4/UASMaf-S;THGal4/+; 'UAS-αSyn/UAS-Maf-S', THGal4, UAS-αSynuclein/UASMaf-S;THGal4, UAS-αSynuclein/+; 'keap1EY5',THGal4/+;THGal4/keap1EY5; 'UAS-αSyn/keap1EY5', THGal4, UAS-αSynuclein/+;THGal4, UAS-αSynuclein/keap1EY5.
[Reproduced from Barone et al. 2011) in agreement with the "Open access" policy of "Disease models and mechanisms".]
Discussion
The assays described here provide a useful approach to study the role of specific genes, signaling pathways or small compounds in the maintenance of DA neurons in aging as well as in different disease-linked genetic backgrounds (reviewed in 5).
The startle-induced negative geotaxis behavior of Drosophila has been extensively used as a functional read-out for the functionality of DA neurons in different genetic backgrounds, particularly in the presence of PD-linked mutations. Some discrepancies among different studies have been observed (see 12 for a more detailed discussion). These have been attributed at least in part to different assay set-ups and conditions, including the number of flies tested in each tube (single fly versus flies' cohort), the time of recovery after CO2-mediated anesthesia and the number of consecutive trials (see 4 and 11 as examples of different assay set-ups). It might therefore be advisable to control for such parameters and, if necessary optimize them for specific assay requirements.
The quantification of DA neurons by TH immunostaining/confocal microscopy has been used interchangeably with the method based on THGal4-driven GFP expression and identification of DA neurons by GFP fluorescence/confocal microscopy. The two methods give comparable results in the majority of cases; however, in general, the TH immunostaining technique is preferred by us and other investigators as it is sensitive to the functional state of DA neurons. For a more detailed comparison see 11 and 10. This assay could be complemented by measuring the dopamine levels in fly heads homogenates [see 1 as an example].
Disclosures
We declare that we have no competing financial interests.
Acknowledgments
We thank Leo Pallanck for fly stocks, Christine Sommers for technical assistance and Gerasimos P. Sykiotis for helpful discussions. This work was funded by the NIH training grant T32CA009363 to M.C.B.
References
- Bayersdorfer F, Voigt A, Schneuwly , Botella J. Dopamine-dependent neurodegeneration in Drosophila models of familial and sporadic Parkinson's disease. Neurobiol. Dis. 2010;40:113–119. doi: 10.1016/j.nbd.2010.02.012. [DOI] [PubMed] [Google Scholar]
- Feany MB, Bender WW. A Drosophila model of Parkinson's disease. Nature. 2000;404:394–398. doi: 10.1038/35006074. [DOI] [PubMed] [Google Scholar]
- Friggi-Grelin F, Coulom H, Meller M, Gomez D, Hirsh J, Birman S. Targeted gene expression in Drosophila dopaminergic cells using regulatory sequences from tyrosine hydroxylase. J. Neurobiol. 2003;54:618–627. doi: 10.1002/neu.10185. [DOI] [PubMed] [Google Scholar]
- Gargano JW, Martin I, Bandari P, Grotewiel MS. Rapid iterative negative geotaxis (RING): a new method for assessing age-related locomotor decline in Drosophila. Exp. Gerontol. 2005;40:386–395. doi: 10.1016/j.exger.2005.02.005. [DOI] [PubMed] [Google Scholar]
- Hirth F. Drosophila melanogaster in the study of human neurodegeneration. CNS Neurol. Disord. Drug Targets. 2010;9:504–523. doi: 10.2174/187152710791556104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lundell M, Hirsh J. Temporal and spacial development of serotonin and dopamine neurons in the Drosophila CNS. Dev. Biol. 1994;165:385–396. doi: 10.1006/dbio.1994.1261. [DOI] [PubMed] [Google Scholar]
- Mao Z, Davis RL. Eight different types of dopaminergic neurons innervate the Drosophila mushroom body neuropil: anatomical and physiological heterogeneity. Front Neural Circuits. 2009;3:5. doi: 10.3389/neuro.04.005.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mizuno H, Fujikake N, Wada K, Nagai Y. Alpha-Synuclein Transgenic Drosophila As a Model of Parkinson's Disease and Related Synucleinopathies. Parkinsons Dis. 2011;2011:212706. doi: 10.4061/2011/212706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Neckameyer WS, Woodrome S, Holt B, Mayer A. Dopamine and senescence in Drosophila melanogaster. Neurobiol. Aging. 2000;21:145–152. doi: 10.1016/s0197-4580(99)00109-8. [DOI] [PubMed] [Google Scholar]
- Trinh K, Moore K, et al. Induction of the phase II detoxification pathway suppresses neuron loss in Drosophila models of Parkinson's disease. J. Neurosci. 2008;28:465–472. doi: 10.1523/JNEUROSCI.4778-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White KE, Humphrey DM, Hirth F. The dopaminergic system in the aging brain of Drosophila. Front Neurosci. 2010;4:205. doi: 10.3389/fnins.2010.00205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitworth AJ, Wes PD, Pallanck LJ. Drosophila models pioneer a new approach to drug discovery for Parkinson's disease. Drug Discov. Today. 2006;11:119–126. doi: 10.1016/S1359-6446(05)03693-7. [DOI] [PubMed] [Google Scholar]
- Wu JS, Luo L. A protocol for dissecting Drosophila melanogaster brains for live imaging or immunostaining. Nat. Protoc. 2006;1:2110–2115. doi: 10.1038/nprot.2006.336. [DOI] [PubMed] [Google Scholar]
- Barone MC, Sykiotis GP, Bohmann D. Genetic activation of Nrf2 signaling is sufficient to ameliorate neurodegenerative phenotypes in a Drosophila model of Parkinson's disease. Dis. Model Mech. 2011;4(5):701–707. doi: 10.1242/dmm.007575. [DOI] [PMC free article] [PubMed] [Google Scholar]