

Abstract
Extensive morphogenetic remodelling takes place during metamorphosis from a larval to an adult insect body plan. These changes are particularly intricate in the generation of the dipteran wing hinge, a complex structure that is derived from an apparently simple region of the wing imaginal disc. Using the characterisation of original outstretched alleles of the unpaired locus as a starting point, we demonstrate the role of JAK/STAT pathway signalling in the process of wing hinge development. We show that differences in JAK/STAT signalling within the proximal most of three lateral folds present in the wing imaginal disc is required for fold morphology and the subsequent differentiation of the first and second auxiliary sclerites as well as the posterior notal wing process. Changes in these domains are consistent with the established fate map of the wing disc. We show that outstretched wing posture phenotypes arise from the loss of a region of Unpaired expression in the proximal wing fold and demonstrate that this results in a decrease in JAK/STAT pathway activity. Finally we show that reduction of JAK/STAT pathway activity within the proximal wing fold is sufficient to phenocopy the outstretched phenotype. Taken together, we suggest that localised Unpaired expression and hence JAK/STAT pathway activity, is required for the morphogenesis of the adult wing hinge, providing new insights into the link between signal transduction pathways, patterning and development.
Introduction
First characterised over 80 years ago, the outstretched (os) phenotype represents the first JAK/STAT pathway-related defect to be genetically described [1]. Originally named on the basis of the distinctive outstretched wing posture of adults (Figure 1A–F), other alleles with small eyes (oss; Figure 1I) or both small eyes and outstretched wings (Figure 1D,F & J) were subsequently identified [2]. Some years later, mutations in the unpaired (upd) locus were independently identified on the basis of their atypical embryonic pair-rule segmentation defects [3] and complementation experiments demonstrated that strong upd alleles fail to complement the original viable os alleles, suggesting that both represent mutations in the same locus [4], [5].
Figure 1. The outstretched wing phenotype.

(A) Wild type (w1118) adult male Drosophila melanogaster showing typical resting wing posture. (B) Adult males hemizygous for the oso allele showing their typical resting wing posture. (C–F) Adult males of the indicated hemizygous genotypes showing resting wing postures typically associated with the outstretched phenotype. Dorsal (C,D) and ventral (E–F) views are shown. The angle of the wing relative to the body axis (dotted lines) is indicated. (G–J) Eyes of the indicated genotypes. Extent of ventral eye field is outlined by dashed line. Minor ventral eye size reduction (arrows) is visible in oss and os1 alleles.
Using embryonic cuticle clonal analysis, genetic characterisation of strong upd alleles demonstrated that the associated gene encodes a protein able to act non-autonomously during development [6]. Consistent with this, molecular cloning showed that upd encodes a secreted and glycosylated protein able to act as a JAK/STAT pathway ligand [7], [8]. More recently, two additional Upd-like ligands termed Upd2 and Upd3 have also been identified. All three ligands are able to stimulate pathway signalling both in vivo and in cell based assays [9] with upd and upd2 being expressed in similar patterns and functioning semi-redundantly during development [10].
As shown both in vivo and in cell culture-based systems, Upd ligands bind to the Domeless cytokine receptor [11], [12], causing the activation of the JAK tyrosine kinase homologue Hopscotch [4], [13] which leads to phosphorylation of the Drosophila STAT92E transcription factor [14], [15]. Once activated, STAT92E translocates into the nucleus where it binds a palindromic DNA sequence present in the promoter of target genes and activates transcription. Functionally, the Drosophila JAK/STAT pathway is involved in multiple developmental processes including haematopoiesis, immune responses, cell proliferation and stem cell maintenance (reviewed in [16]–[18]). In addition, studies into the role of JAK/STAT signalling in the developing Drosophila eye have shown that reduced pathway activity results in cellular under proliferation [19] to generate a phenotype similar to the viable oss and os1 small eye phenotype (Figure 1G–J).
Here we present an analysis of the previously uncharacterised role for upd expression in the developing wing hinge as demonstrated by outstretched alleles. We show that the development of adult structures on the dorsal hinge requires the most proximal of three folds present within the third instar wing imaginal disc. Furthermore, formation of this fold requires localised upd expression, which is lacking in os alleles. Using in vivo RNAi approaches, we also show that reducing JAK/STAT pathway activity can phenocopy the held out wing defect associated with these previously uncharacterised classical os alleles.
Results and Discussion
The Outstretched Phenotype
Wild type adult Drosophila melanogaster generally maintain a characteristic resting wing posture where wings are held almost parallel to the main body axis (Figure 1A). By contrast, males hemizygous for the outstretched (os) alleles oso and os1 hold their wings out at 30–80° from the principal body axis (Figure 1B–F) but have apparently normal wing and body morphology (Figure 1C–F). In addition males hemizygous for os1 and oss also have mildly reduced ventral eye size (arrows in Figure 1I,J) compared to wild type and oso alleles (Figure 1G,H). However, the wing posture of oss flies is normal.
In order to better understand the basis of the os wing posture phenotype we examined the hinge area of adult flies. By comparison to wild type males (Figure 2A), individuals hemizygous for both oso and os1 alleles show distinct dorsal hinge defects. Flies carrying the os1 allele have variable expressivity: those with a relatively ‘weak’ held out phenotype have a reduced/rudimentary first auxiliary sclerite (AS1) (arrow in Figure 2B). Moderately affected individuals lack the AS1 completely (Figure 2C) while individuals with the strongest expressivity lack AS1 and, in addition, the AS2 and AS3 are reduced as is the posterior notal wing process (PNWP) (Figure 2D). However, adjacent structures such as the unnamed plate (UP) and the anterior notal wing process (ANWP) appear normal in all individuals. By contrast to the differences apparent on the dorsal surface of the hinge, ventral regions of both wild type and moderate os1 alleles appear to be similar (Figure 2E–F).
Figure 2. Structure of the os1 hinge and adult musculature.

Scanning electron micrographs of the adult dorsal (A–D) and ventral (E–F) wing hinge regions of wild type (A, E) and os1 (B–D and F) individuals. The relative expressivity of the os1 allele phenotypes is indicated by ‘weak’ ‘moderate’ and ‘strong’ labels. Note how increasing phenotypic strength is associated with the progressive loss of the AS1, AS2 and PNWP. Wing hinge structures are named in [25]. Unnamed plate (UP), first, second and third auxiliary sclerites (AS1, AS2 and AS3), anterior notal wing process (ANWP) and posterior notal wing process (PNWP) are present in the dorsal hinge. In the ventral hinge, auxillary pouch (AP) and proximal ventral radius (PVR) are shown. (G–J) Longitudinal sections through adult males of the indicated genotypes show the structure of the primary flight muscles.
An alternative cause of wing posture phenotypes in Drosophila are defects in the primary flight musculature, one example of which are homozygous parkin mutants (compare Figure 2G and 2H) [20]. However, longitudinal sections through both oso and os1 males (Figure 2I–J) show muscle structures similar to wild type (Figure 2G) suggesting that the wing posture defect in these alleles is not a consequence of a gross defect in flight muscle development.
Taken together, these results suggest that defects in the wing hinge, and the auxiliary sclerites, and AS1 in particular, are the basis of the wing posture defects observed in os1 and oso mutants.
Outstretched Mutant Wing Discs
The adult wing, hinge region and dorsal thoracic body wall are derived from the wing imaginal disc, a sheet of columnar epithelial cells set aside during embryogenesis [21]. During larval life, cells within each wing imaginal disc proliferate and grow and by late third instar the disc contains three prominent transverse folds spanning the presumptive dorsal hinge region. During pupariation the wing imaginal discs undergo a series of complex morphological rearrangements termed eversion that ultimately give rise to the adult structures [22].
Given the correlation between os1 wing posture phenotypes and the loss of dorsal wing hinge structures shown above, we examined the expression of upd mRNA within the third instar wing imaginal disc. In wild type individuals, upd is expressed within three regions of the presumptive dorsal wing hinge as well as in the anterior body wall and, at lower levels, in the ventral body wall (purple regions in Figure 3A and 3E). By contrast, in oso wing discs the most proximal dorsal region of upd expression is no longer detected (arrow in Figure 3B), although other expression domains appear normal.
Figure 3. Expression of upd and activation of JAK/STAT pathway signalling in the wing disc.

Proximal (P), medial (M) and distal (D) folds are labelled. Scale bar is 100 µm, (A & B) Third instar wing imaginal discs showing the expression of upd mRNA in wild type (A) and oso (B) individuals. Expression of upd within the P fold (arrow) is lost in the oso mutant. (C & D) Third instar wing imaginal discs showing DNA (white) and the 10xSTATGFP reporter (green) in wild type (C) and oso (D) individuals. Reporter activity within the P fold (arrow) is greatly reduced in oso mutants suggesting that JAK/STAT pathway ligands are not expressed in this area. (E) Schematic representation of the third instar wing imaginal disc fate map showing the regions labelled in Figure 2, the P, M and D lateral folds and the approximate regions of wild type upd expression (purple). Image based on fate map originally published in [25].
In vitro reporters containing multiple STAT92E binding sites upstream of a GFP coding region allow the spatial and temporal pattern of JAK/STAT pathway activity elicited by Upd-like ligands to be visualised during development [9], [23]. We used one such 10xSTATGFP reporter to identify regions of pathway activation within the third instar wing imaginal disc. Consistent with upd mRNA expression, 10xSTATGFP reporter activity in wild type wing discs is detected in regions centred on the areas of ligand expression (Figure 3C). Furthermore, these regions are larger than the area of detectable upd expression, a finding that is consistent with Upd being a diffusible molecule able to trigger pathway activity non-autonomously [7], [8]. In oso mutant wing discs, 10xSTATGFP activity is not detected in the proximal fold (arrow in Figure 3D). This result is consistent with the upd in situ data and also suggests that no other upd-like genes able to stimulate the JAK/STAT pathway are expressed in this domain in oso mutants.
One of the more noticeable features of the third instar wing imaginal disc are three prominent lateral folds, which we term the proximal (P), medial (M) and distal (D) folds (Figure 3 and 4). These folds are present within the presumptive dorsal hinge between the wing pouch and the future notum, and run parallel to one another across both anterior and posterior compartments of the disc (Figure 3E). Expression domains of upd during late third instar development coincide with the M and P folds (Figure 3A), as does 10xSTATGFP reporter expression (Figure 3C) while upd is also expressed in this general domain at earlier stages before folds are formed [24]. Consistent with the change in upd expression and JAK/STAT pathway activity in oso mutants described above, fate mapping experiments have previously identified the AS1 as being derived from a region of the disc located in the middle of the P fold (Figure 3E) [25]. This correlation is both consistent with our own findings and also supports the accuracy of the original fate mapping experiments.
Figure 4. JAK/STAT signalling and fold formation.
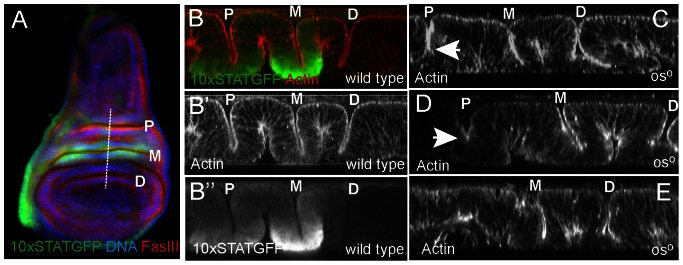
Proximal (P), medial (M) and distal (D) folds are labelled. (A) Wild type third instar wing imaginal discs showing the pattern of 10xSTATGFP reporter activity (green), FasIII (red) and DNA (blue). The approximate position of the XZ sections shown in B–E is indicated by the dotted line. (B) An XZ section through the lateral wing folds of wild type third instar wing imaginal disc showing F-actin (red in B and white in B’) and the 10xSTATGFP reporter (green in B and white in B”). (C–E) XZ sections through third instar wing imaginal discs of males hemizygous for the oso allele stained to visualise F-actin (white), and hence disc structure. The P fold is reduced in depth in C and D (arrows) and is missing completely in E.
Taken together, our results suggest a link between the loss of the AS1 and loss of an upd expression domain in oso mutants.
Wing Disc Folds
The P, M and D folds present within the third instar wing imaginal disc can be readily visualised both in plan (XY; Figure 4A) and cross section (XZ; Figure 4B). Viewed in cross section (Figure 4B), folds can be seen to result from a concertina-type arrangement of the columnar epithelium in this area, a morphology that also results in two additional ‘opposite’ folds on the basal surface. Within this structure, 10xSTATGFP reporter activity is highest in the cells making up the basal portion of each fold with overall levels of pathway activation higher in the M than the P fold (Wells et al submitted; Figure 4B”). Given the diffusible nature of the Upd ligand, this localised JAK/STAT pathway activation may be a result of ligand trapping within the fold. By contrast, and consistent with the lack of upd expression in this region, no reporter activity is detected in the D fold.
While all three folds are consistently of comparable depth in wild type discs (Figure 4B) the P fold in oso individuals is often disrupted (Figure 4C–E). Although somewhat variable, the P fold phenotype in this background can include a reduction in fold depth (Figure 4C), a reduced depth combined with an apical widening of the fold (Figure 4D) or even complete loss of the structure (Figure 4E). It seems likely that variations in P fold morphology and the variable loss of structures within the adult wing hinge derived from this region of the disc are related, with more severe fold phenotypes possibly resulting in more extensive loss of hinge structures.
Modulation of STAT92E can Reproduce the Outstretched Phenotype
The link between the oso phenotype, upd expression and the morphology of the AS1 in the adult suggests that the os wing posture phenotype results from a defect in JAK/STAT signalling. In order to test this we used the 10xSTATGFP pathway reporter recombined with a patched (ptc) transgene expressing Gal4 in a stripe of cells running through the middle of presumptive dorsal hinge region. Using this driver, we expressed an in vivo RNAi hairpin targeting the single stat92E transcript (Figure 5A) and find that activity of the 10xSTATGFP pathway reporter is down regulated in a domain that intersects both the M and P folds (arrow in Figure 5A’). This demonstrates the extent of the ptc expression domain, the efficiency of knockdown and, as previously shown in Wells et al (submitted), also results in disruption of P fold morphology. Having established the extent of JAK/STAT pathway knockdown we allowed individuals to develop to adulthood. While a control RNAi targeting Rhodopsin4 (Rh4), a gene not expressed in the wing, shows no adult wing posture phenotype (Figure 5B), knockdown of stat92E results in a held out wing phenotype reminiscent of os1 and oso (Figure 5C). When examined by SEM, ptc>stat92E RNAi flies show defects in the dorsal hinge with a reduction in both the AS1 and AS2 (Figure 5D). These defects are comparable to the original os1 allele (Figure 2) and suggest that the wing posture phenotype in both genetic backgrounds are caused by the same fundamental mechanism.
Figure 5. Modulation of JAK/STAT activity phenocopies os defects.
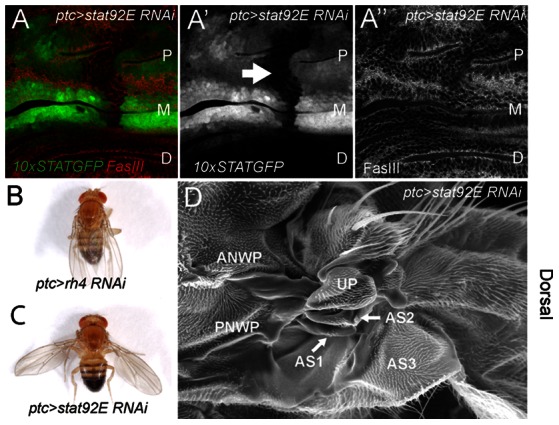
(A) Dorsal hinge region of a third instar wing imaginal disc from a 10xSTATGFP, ptc>Gal4/UAS-stat92E RNAi individual showing the extent of 10xSTATGFP reporter activity (green in A, white in A’) and FasciclinIII (red in A, white in A”) to outline cell shape. P, M and D folds are labeled and the domain in which reporter activity is ablated following knockdown of stat92E is shown (arrow in A’), (B–C) Adult male flies of the indicated genotypes showing resting wing posture. Knockdown of stat92E mRNA in the ptc domain produces a clear and highly penetrant wings held out defect. (D) SEM of the dorsal wing hinge of a ptc>stat92E RNAi male with a wings held out phenotype. Compared to a wild type hinge (Figure 2A) both the first and second auxiliary sclerites (AS1 and AS2) are reduced.
In summary, we present the first detailed morphological and gene expression data of the classical group of outstretched (os) alleles – viable alleles of the unpaired locus. We go on to show that pathway activity in the proximal (P) lateral fold normally present in the third instar wing disc, is required for the normal development of this fold, and that a reduction of JAK/STAT pathway signalling in this fold is sufficient to phenocopy the os wing posture defect.
Materials and Methods
Drosophila Stocks and Genetics
Drosophila stocks were maintained on standard food at 25°C unless otherwise stated. Stocks used were white1118 (w1118) as a wild type control, w,os1 and f,oso,car (gift of the Perrimon lab, Harvard Medical School) and oss (Bloomington Stock Centre – stock 79). 10xSTATGFP reporter lines were a gift of Erika Bach (NYU Langone Medical Centre). Recombinants between 10xSTATGFP [23] and ptc>Gal4 [26] were generated using standard techniques. The in vivo RNAi constructs targeting stat92E mRNA (ID 106980) and Rh4 mRNA (ID 101708) [27] were obtained from the Vienna Drosophila RNAi Centre.
Scanning Electron Microscopy
Scanning electron micrographs were prepared from representative males flies of the indicated genotypes. In order to maintain normal live wing posture flies were rapidly killed in a petri dish on dry ice for 90 minutes, before being placed on 12 mm round Leit Adhesive Carbon Tabs (Agar Scientific). Flies were handled via their legs to ensure minimal disturbance of wing position. Samples were mounted onto 0.5 inch aluminium studs and gold coated using an Edwards S1501b sputter coater (Edwards, Crawley). Samples were examined in a Philips SEM XL-20 with an operating voltage of 20 kv. Pictures were taken at a working distance of between 9.8 and 10.2 mm with a magnification of 400x.
Histology and Microscopy
Wing imaginal discs were dissected from wandering third instar larvae of the indicated genotypes. Whole mount in situ hybidisation to wing imaginal discs was undertaken as described [24] using antisense RNA probes generated from Upd cDNA plasmids (gift of Norbert Perrimon, Harvard Medical School). Immunostaining of imaginal discs was undertaken as described in [8]. Discs were counterstained to visualise DNA and filamentous-actin (F-actin) according to standard protocols using DAPI or Phaloidin-Alexa568 respectively. Antibodies used include mouse anti-Fasciclin III (1∶10), (Developmental Studies Hybridoma Bank). Secondary antibodies were from Jackson Labs.
Whole flies were photographed using a Nikon SMZ1500 stereo microscope and Nikon Elements extended depth of focus software package. Imaginal discs with in situ hybridisation patterns and sections through intact flies were imaged using differential interference contrast or bright field techniques on a Zeiss Axioskop microscope with Q-Imaging camera system. Zeiss LSM510 or Leica SP confocals were used to capture XY and XZ sections of imaginal discs containing fluorescent proteins and stains.
Longitudinal sections of adult flight muscles flies were generated from male w1118 and parkin25 mutants (used as controls [20]) and hemizygous os0 and os1 males. Individuals were anesthetised before partial dissection to remove legs, wings, proboscis and the posterior abdomen. Individuals were then fixed in 10% formaldehyde in PBS for 1 hr before dehydration through a series of ethanol dilutions (30%, 50%, 70%, 90% and 100% x 3) for 15 minutes at each step. Flies were then incubated over night in JB4 (Polysciences Inc) solution A combined with 18 mg/ml of catalyst (benzoyl peroxide) before being arranged into embedding moulds containing embedding solution mixture (JB4 solution A, 18 mg/ml of catalyst and 1% JB4 solution B). Block holders were added to exclude oxygen during the polymerisation process. Moulds were incubated at 4°C overnight before 3–5 µm sections were cut using a glass blade on a LKB microtome (Bromma, Sweden) and stained using 1% Tolundine blue/1% Na-Borax for 1 minute before being washed with water and dried. Coverslips were mounted using DPX before imaging.
Acknowledgments
The authors wish to thank Shirui Hou for initial observations and Doug Harrison for sharing information prior to publication. The Developmental Studies Hybridoma Bank for antibodies, Alex Whitworth, Norbert Perrimon and the Bloomington Stock Centre for Drosophila stocks, Chris Hill for assistance with sectioning and electron microscopy and the Sheffield Light Microscopy Facility. MPZ is a Cancer Research UK Senior Cancer Research Fellow.
Funding Statement
Funding has been provided by a Cancer Research UK Senior Cancer Research Fellowship to MPZ. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Müller HJ (1930) Types of visible variations induced by X-rays in Drosophila. J Genet 22: 299–334. [Google Scholar]
- 2. Müller HJ (1935) [New mutants report.]. DIS 3: 29–30. [Google Scholar]
- 3. Wieschaus E, Nusslein-Volhard C, Jurgens G (1984) Mutations affecting the pattern of the larval cuticle in Drosophila melanogaster. III. Zygotic loci on the X-chromosome and fourth chromosome. Rouxs Arch Dev Biol 193: 296–307. [DOI] [PubMed] [Google Scholar]
- 4. Eberl DF, Perkins LA, Engelstein M, Hilliker AJ, Perrimon N (1992) Genetic and developmental analysis of polytene section 17 of the X chromosome of Drosophila melanogaster. Genetics 130: 569–583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Tsai YC, Sun YH (2004) Long-range effect of upd, a ligand for Jak/STAT pathway, on cell cycle in Drosophila eye development. Genesis 39: 141–153. [DOI] [PubMed] [Google Scholar]
- 6. Gergen JP, Wieschaus EF (1986) Localized requirements for gene activity in segmentation of Drosophila embryos: analysis of armadillo, fused, giant and unpaired mutations in mosaic embryos. Rouxs Arch Dev Biol 195: 49–62. [DOI] [PubMed] [Google Scholar]
- 7. Harrison DA, McCoon PE, Binari R, Gilman M, Perrimon N (1998) Drosophila unpaired encodes a secreted protein that activates the JAK signaling pathway. Genes Dev 12: 3252–3263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Zeidler MP, Perrimon N, Strutt DI (1999) Polarity determination in the Drosophila eye: a novel role for unpaired and JAK/STAT signaling. Genes Dev 13: 1342–1353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Wright VM, Vogt KL, Smythe E, Zeidler MP (2011) Differential activities of the Drosophila JAK/STAT pathway ligands Upd, Upd2 and Upd3. Cell Signal 23: 920–927. [DOI] [PubMed] [Google Scholar]
- 10. Hombria JC, Brown S, Hader S, Zeidler MP (2005) Characterisation of Upd2, a Drosophila JAK/STAT pathway ligand. Dev Biol 288: 420–433. [DOI] [PubMed] [Google Scholar]
- 11. Brown S, Hu N, Hombria JC (2001) Identification of the first invertebrate interleukin JAK/STAT receptor, the Drosophila gene domeless. Curr Biol 11: 1700–1705. [DOI] [PubMed] [Google Scholar]
- 12. Chen HW, Chen X, Oh SW, Marinissen MJ, Gutkind JS, et al. (2002) mom identifies a receptor for the Drosophila JAK/STAT signal transduction pathway and encodes a protein distantly related to the mammalian cytokine receptor family. Genes Dev 16: 388–398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Binari R, Perrimon N (1994) Stripe-specific regulation of pair-rule genes by hopscotch, a putative Jak family tyrosine kinase in Drosophila. Genes Dev 8: 300–312. [DOI] [PubMed] [Google Scholar]
- 14. Hou XS, Melnick MB, Perrimon N (1996) Marelle acts downstream of the Drosophila HOP/JAK kinase and encodes a protein similar to the mammalian STATs. Cell 84: 411–419. [DOI] [PubMed] [Google Scholar]
- 15. Yan R, Small S, Desplan C, Dearolf CR, Darnell JEJ (1996) Identification of a Stat gene that functions in Drosophila development. Cell 84: 421–430. [DOI] [PubMed] [Google Scholar]
- 16. Arbouzova NI, Zeidler MP (2006) JAK/STAT signalling in Drosophila: insights into conserved regulatory and cellular functions. Development 133: 2605–2616. [DOI] [PubMed] [Google Scholar]
- 17.Resende LP, Jones DL (2012) Local signaling within stem cell niches: insights from Drosophila. Curr Opin Cell Biol. [DOI] [PMC free article] [PubMed]
- 18. Wang YH, Huang ML (2010) Organogenesis and tumorigenesis: insight from the JAK/STAT pathway in the Drosophila eye. Dev Dyn 239: 2522–2533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Bach EA, Vincent S, Zeidler MP, Perrimon N (2003) A sensitized genetic screen to identify novel regulators and components of the Drosophila janus kinase/signal transducer and activator of transcription pathway. Genetics 165: 1149–1166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Greene JC, Whitworth AJ, Kuo I, Andrews LA, Feany MB, et al. (2003) Mitochondrial pathology and apoptotic muscle degeneration in Drosophila parkin mutants. Proc Natl Acad Sci U S A 100: 4078–4083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Cohen B, Simcox AA, Cohen SM (1993) Allocation of the thoracic imaginal primordia in the Drosophila embryo. Development 117: 597–608. [DOI] [PubMed] [Google Scholar]
- 22. Taylor J, Adler PN (2008) Cell rearrangement and cell division during the tissue level morphogenesis of evaginating Drosophila imaginal discs. Dev Biol 313: 739–751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Bach EA, Ekas LA, Ayala-Camargo A, Flaherty MS, Lee H, et al. (2007) GFP reporters detect the activation of the Drosophila JAK/STAT pathway in vivo. Gene Expr Patterns 7: 323–331. [DOI] [PubMed] [Google Scholar]
- 24. Mukherjee T, Hombria JC, Zeidler MP (2005) Opposing roles for Drosophila JAK/STAT signalling during cellular proliferation. Oncogene 24: 2503–2511. [DOI] [PubMed] [Google Scholar]
- 25. Bryant PJ (1975) Pattern formation in the imaginal wing disc of Drosophila melanogaster . J Exp Zool 193: 49–77. [DOI] [PubMed] [Google Scholar]
- 26. Brand AH, Perrimon N (1993) Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118: 401–415. [DOI] [PubMed] [Google Scholar]
- 27. Dietzl G, Chen D, Schnorrer F, Su KC, Barinova Y, et al. (2007) A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature 448: 151–156. [DOI] [PubMed] [Google Scholar]