

Abstract
Wolbachia is a maternally transmitted endosymbiotic bacterium that is estimated to infect up to 65% of insect species. The ability of Wolbachia to both induce pathogen interference and spread into mosquito vector populations makes it possible to develop Wolbachia as a biological control agent for vector-borne disease control. Although Wolbachia induces resistance to dengue virus (DENV), filarial worms, and Plasmodium in mosquitoes, species like Aedes polynesiensis and Aedes albopictus, which carry native Wolbachia infections, are able to transmit dengue and filariasis. In a previous study, the native wPolA in Ae. polynesiensis was replaced with wAlbB from Ae. albopictus, and resulted in the generation of the transinfected “MTB” strain with low susceptibility for filarial worms. In this study, we compare the dynamics of DENV serotype 2 (DENV-2) within the wild type “APM” strain and the MTB strain of Ae. polynesiensis by measuring viral infection in the mosquito whole body, midgut, head, and saliva at different time points post infection. The results show that wAlbB can induce a strong resistance to DENV-2 in the MTB mosquito. Evidence also supports that this resistance is related to a dramatic increase in Wolbachia density in the MTB's somatic tissues, including the midgut and salivary gland. Our results suggests that replacement of a native Wolbachia with a novel infection could serve as a strategy for developing a Wolbachia-based approach to target naturally infected insects for vector-borne disease control.
Author Summary
Aedes polynesiensis is a vector for both dengue and filariasis in the South Pacific. Efforts are ongoing to utilize Wolbachia as a biological control agent targeting this vector through either population suppression via releases of incompatible males or population replacement for spreading disease resistance into a population. Replacing the native Wolbachia with a novel infection from Ae. albopictus has generated the “MTB” strain of Ae. polynesiensis. This MTB mosquito is reproductively-incompatible with the wild type of Ae. polynesiensis and has a low susceptibility for filarial worms. In this work, we show that the MTB mosquito is resistant to dengue virus with a reduced viral infection in the mosquito whole body, midgut, head, and saliva. Our results further show its refractoriness to dengue virus is associated with a dramatic increase in Wolbachia density in those mosquito tissues where dengue virus needs to reside, replicate, and travel in order to be transmitted to humans. These results suggest that the MTB strain has the potential to be used in Wolbachia-based strategies to control both dengue and filariasis in the South Pacific.
Introduction
Wolbachia is a maternally transmitted endosymbiotic bacterium that infects an estimated 65% of insect species [1], [2]. The ability of Wolbachia to both induce viral interference and spread into mosquito vector populations makes it a potential candidate as a biological control agent for dengue control [3], [4], [5], [6], [7]. The primary dengue vector, Aedes aegypti, does not naturally carry Wolbachia. When artificially introduced into Ae. aegypti, three types of Wolbachia (wAlbB, wMelPop-CLA, and wMel) show significant inhibition of dengue virus (DENV) replication and dissemination, resulting in either complete or partial block of viral transmission [4], [6], [7]. Furthermore, this Wolbachia-mediated pathogen interference has a broad spectrum and can inhibit a variety of pathogens in Ae. aegypti, including Chikungunya, Plasmodium, and filarial worms [7], [8].
In addition to Ae. aegypti, dengue is transmitted by secondary mosquito vectors including Ae. albopictus and Ae. polynesiensis [9], [10], [11]. Both species of mosquitoes can transmit other pathogens to humans as well. While Ae. albopictus is a competent vector for at least 22 arboviruses [10], Ae. polynesiensis is the primary vector of Wuchereria bancrofti in the South Pacific [12]. In contrast to Ae. aegypti, both of them naturally carry Wolbachia infections. Ae. albopictus is infected with wAlbA and wAlbB [13], and Ae. polynesiensis with wPolA [14]. These Wolbachia can induce cytoplasmic incompatibility (CI) when infected males mate with females that are uninfected or carry different type of Wolbachia [14], [15]. Although native Wolbachia infections were reported to confer host resistance to pathogen infection in both the Drosophila and Culex mosquitoes [16], [17], neither wAlbA and wAlbB, nor wPolA appear to induce resistance to DENV in Ae. albopictus or Ae. polynesiensis, respectively [6], [9], [18].
Wolbachia-induced viral interference depends on the density of Wolbachia. In the original Ae. albopictus Aa23 cell line, wAlbB reaches a density high enough to completely clear dengue infection [18]. A strong negative linear correlation was observed between the genome copy numbers of wAlbB and DENV [18]. In the mosquito vector, this density-dependent viral inhibition occurs in a tissue-specific manner. A very low density of native Wolbachia in mosquito somatic tissues (e.g., midguts and salivary glands) makes its resistance to DENV undetectable [18], or detected with a weak effect [19]. When the native Wolbachia was replaced with a wMel infection, the transinfected Ae. albopictus becomes strongly resistant to DENV. This resistance is associated with a 7-fold increase in wMel density compared to the native infection [20].
Following ingestion by mosquitoes in a DENV-infected blood meal, virus first enters the midgut epithelial cells and infects the midgut tissue, in which it replicates to produce more viral particles. DENV then escapes from the midgut and disseminates through the hemolymph to other tissues, including the salivary glands, where it replicates and resides until it is injected into a human host [21]. During this journey virus needs to pass a number of physiological barriers, such as the midgut infection/escape barrier and salivary gland infection/escape barrier, and is subjected to a series of attacks by the mosquito immune system [21], [22], [23], [24]. The dynamics and tropism of DENV within a mosquito are dependent on the mosquito strain, the virus genotype, and environmental factors [25]. Recent studies show that endogenous microbial flora, including Wolbachia, also influence the mosquito's tissue- and cell-specific susceptibility to DENV by boosting basal immunity [22], [26].
Due to its importance in transmitting both Lymphatic filariasis and dengue in the South Pacific, Ae. polynesiensis was targeted for control through either population suppression via releases of incompatible males or population replacement to harness CI as a gene-drive mechanism for spreading disease resistance into a population [12], [14]. A strain of Ae. polynesiensis was previously infected with Wolbachia from Ae. riversi through interspecific hybridization and introgression. This strain was used to test a population suppression strategy in semi-field conditions with encouraging results [12]. With successful experiences in Wolbachia transinfection through embryo microinjection, additional studies were conducted to transfer Wolbachia from Ae. albopictus into the aposymbiotic strain of Ae. polynesiensis (APMT), resulting in the “MTB” strain stably infected by wAlbB only [14]. There is a strong bi-directional CI when the MTB strain crosses with the wild type “APM” strain. Furthermore, the number of successfully developing infective stage filarial worms was reduced in the MTB strain [14], which was consistent with previous findings that interaction of wAlbB with mosquito hosts can trigger a process of pathogen interference [26].
The mosquito MTB strain has a decreased ability to regulate oxidative stress [14]. We previously showed that Wolbachia can induce reactive oxygen species (ROS)-dependent activation of the Toll pathway to control DENV in Ae. aegypti [26]. Due to the role of Ae. polynesiensis in dengue transmission, it is important to know how wAlbB will influence MTB's susceptibility for DENV. In this study, we measured the vector competence of the MTB strain for DENV serotype 2 (DENV-2) as compared to the naturally infected APM strain of Ae. polynesiensis. We also compared the Wolbachia density between the MTB and APM strain. These results indicate that wAlbB induces a strong resistance to DENV in the MTB strain, which is associated with an increased Wolbachia density in mosquito somatic tissues.
Methods
Ethics statement
This study was performed in strict accordance with the recommendations in the Guide for Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Institutional Animal Care and Use of the Michigan State University (Application #: 03/11-040-00).
Mosquito rearing and cell culture maintenance
All mosquito strains used in these experiments, including the wild-type APM strain and the transfected line MTB of Ae. polynesiensis, were maintained on sugar solution at 27°C and 85% humidity with a 12-hr light/dark cycle according to standard rearing procedures. After transinfection of wAlbB into AMPT, MTB females were out-crossed with AMPT males for six consecutive generations [14]. APM and MTB were reared in a regular condition for at least two generations before used for experiments with the goal to recover and re-colonize gut bacteria [7]. Female mosquitoes, 3–5 d after eclosion, were fed on the blood of anesthetized white mice to initiate egg development. The Ae. albopictus cell line C6/36 was grown in minimal essential medium (MEM) with 10% heat-inactivated FBS, 1% L-glutamine, and 1% non-essential amino acids at 32°C and 5% CO2.
DENV-2 infections
The New Guinea C strain of DENV-2 was propagated in C6/36 cells according to standard conditions [27]: In brief, 0.5-ml aliquots of virus stock were used to infect 75-cm2 flasks of C6/36 cells, at 80% confluence, with a multiplicity of infection (MOI) of 3.5 virus particles/cell. Infected cells were incubated for 7 days. Then, cells were harvested with a cell scraper and lysed by repeated freezing and thawing in dry ice and a 37°C water bath. Virus isolated from these cells was combined with the supernatant [28], resulting in virus suspension with a titer of 2×107 PFU/ml. For infection through intrathoracic injection, 69 nl of the above virus suspension was used for injection into each female. For infection through oral feeding, the resulting virus suspension was mixed 1∶1 with commercial human blood. A flask with uninfected C6/36 cells was maintained under similar conditions and used to create the noninfectious blood meal that served as our control. The blood meal was maintained at 37°C for 30 min prior to use for feeding 7-day-old mosquitoes [28].
Mosquito dissections
Mosquitoes were dissected to collect the midguts in RNALater at 4, 7 and 10 days post infection (dpi) and thorax at 14 dpi, with three individual mosquitoes in a single replicate. At least five replicate biological assays were performed. Total RNA was extracted using the RNeasy kit (QIAGEN). To measure the virus titers in mosquito bodies, at 14 days after a blood meal, mosquitoes were briefly washed in 70% ethanol, then rinsed in sterile distilled water. The midgut and thorax were dissected in sterile PBS and transferred separately to microcentrifuge tubes containing 150 µl of MEM, and then homogenized with a Kontes pellet pestle motor in a sterile environment. To measure the density of Wolbachia in mosquito tissues, the midguts, salivary glands, fat bodies, and ovaries of 7-day-old non-blood-fed females were dissected and transferred separately to microcentrifuge tubes containing 50 µl of STE buffer for extraction of total genomic DNA.
Real-time qPCR assays
To measure the number of viral genome copies, total virus RNA was extracted using the RNeasy kit (QIAGEN) and reverse-transcribed using Superscript III (Invitrogen, Carlsbad, California, USA) with random hexamers. qRT-PCR was conducted using primers targeting the dengue NS5 gene and the host RPS6 [29]. The dengue genome copy number was normalized using the RPS6 results. Two recombinant plasmids containing the targeted fragments were diluted from 101 to 108 copies/reaction and used to generate separate standard curves for NS5 and RPS6. Real-time quantitation was performed using the QuantiTect SYBR Green PCR Kit (Qiagen) and ABI Detection System ABI Prism 7000 (Applied Biosystems, Foster City, California, USA). Three independent biological replicates were assayed, and all PCR reactions were performed in triplicate. To determine the number of copies of the Wolbachia genome, real-time PCR was carried out as previously described [22], [30].
Plaque assays for DENV-2 virus titration
Virus titers in the tissue homogenates were measured as previously reported [28]: The virus-containing homogenates were serially diluted and inoculated into C6/36 cells in 24-well plates. After incubation for 5 days at 32°C and 5% CO2, the plates were assayed for plaque formation by peroxidase immunostaining, using mouse hyperimmune ascitic fluid (MHIAF, specific for DENV-2) and a goat anti-mouse HRP conjugate as the primary and secondary antibodies, respectively.
Indirect immunofluorescence assay (IFA)
At 7 dpi, the viral antigen in the midguts of mosquitoes was detected by using an indirect IFA. Mosquito midguts were dissected in PBS and fixed in 4% paraformaldehyde for 5 h, and then incubated for 1 h at room temperature in a PBS-BSA-Triton solution (1× PBS, 1% Bovine Serum Albumin, and 0.1% Triton X-100), a mouse anti-dengue complex monoclonal antibody (obtained from the Centers for Disease Control, Atlanta, GA), and a fluorescein-conjugated affinity-purified secondary antibody (Millipore) were used in all midguts assays. Specimens were examined with a Zeiss (Germany) fluorescence microscope.
Transmission assay
Mosquitoes that had been infected with DENV-2 as described above were maintained for 14 days for forced salivation assays. The assays were conducted as previously reported [31], [32]: In brief, mosquitoes were deprived of food for 24 h prior to forced salivation. The legs and wings of each mosquito were cut away, and the proboscis was inserted into 25 µl of feeding solution (50% FBS/164 mM NaCl/100 mM NaHCO3/0.2 mM ATP/≈50 µg sucrose/phenol red, pH 7.0) [32] in a 0.2-ml PCR tube. After 90 min, the mosquitoes were removed, and the feeding solution from each mosquito was sterilized by Millex-GV filter for plaque assays.
Results
A reduced viral infection in the whole bodies of MTB mosquitoes
The potential use of the MTB strain to control Ae. polynesiensis in a dengue endemic area provides a rationale to test if the MTB strain is resistant to DENV as compared to the wild type of Ae. polynesiensis. To test this, we first compared DENV-2 genome copies in the whole body between the MTB strain and the wild type APM strain at 14 days post infection (dpi). Mosquitoes were infected through either oral feeding with DENV-2 infected blood or intrathoracic injection with cell medium containing DENV-2. As a result, we observed that the median number of viral genomes in the whole bodies of MTB mosquitoes was 1.4×104 times lower than that of APM mosquitoes when mosquitoes were infected through oral feeding (Mann-Whitney U test, P<0.05) (Fig. 1). Similarly, mosquito infection through intrathoracic injection led to a significantly lower viral infection in whole bodies of MTB mosquitoes as compared to APM mosquitoes (Mann-Whitney U test, P<0.001). There was a 28.3-fold reduction in the median number of viral genome copies in MTB mosquitoes comparing to APM mosquitoes (Fig. 1).
Figure 1. Inhibition of dengue infection in the whole bodies of MTB mosquitoes.

At 14 dpi (days post infection) through a blood meal or intrathoracic injection, the whole bodies of MTB and APM mosquitoes were collected, and the number of genome copies of the DENV genome was determined by qRT-PCR using primers for the NS5 gene; the results were normalized to the Ae. polynesiensis ribosomal protein S6 (RPS6). Lines indicate the median of the ten biological replicates. Significance was determined using a Mann-Whitney U test.
An inhibited viral replication in the midguts of MTB mosquitoes
In order to determine how viral inhibition occurs in a tissue-specific manner in MTB mosquitoes, we examined whether or not viral replication in the midguts of MTB mosquitoes was suppressed. Both MTB and APM mosquitoes were fed with the DENV-2 infected blood and viral genome copy numbers in the midguts were measured at three different time points: 4, 7 and 10 dpi. The DENV-2 copy numbers were significantly lower at all of the three time points in the midguts of MTB mosquitoes than in APM mosquitoes (Mann-Whitney U test, P<0.01) (Fig. 2A). The viral infection in mosquito midguts at 7 dpi was further assayed by IFA to visualize the intensity and distribution of DENV-2 using an antibody against the viral envelop protein. Consistently, 76.2% (16/21) of APM midguts showed strong positive signals while only 10% (2/20) of MTB midguts had weak positive signals (Fisher's Exact Test, P<0.001) (Fig. 2B; Table 1). This indicates that viral replication is strongly inhibited in the midguts of MTB mosquitoes.
Figure 2. Inhibition of dengue infection in the midgut of MTB mosquitoes.

(A) At 4, 7 and 10 dpi with a blood meal containing DENV-2, mosquito midguts were collected, and the number of genome copies of the DENV-2 genome was determined by qRT-PCR using primers for the NS5 gene; the results were normalized to the Ae. polynesiensis RPS6. Lines indicate the median of the ten biological replicates. Significance was determined using a Mann-Whitney U test. (B) At 7 dpi through a blood meal, mosquito midguts were collected and fixed, and the viral antigen was detected using a mouse anti-Dengue complex monoclonal antibody by indirect fluorescence assay (IFA). One representative midgut from each category is shown. APM mosquitoes fed with dengue-infected or -uninfected blood meal was used as positive (POS.) and negative (NEG.) control, respectively. +, dengue-positive; −, dengue-negative.
Table 1. wAlbB confers resistance to DENV-2 in both midguts and heads of MTB mosquitoes.
| % of infected samples | P value | ||
| Tissues | APM | MTB | |
| Midguts* | 76.2 (16/21) | 10.0 (2/20) | <0.001 |
| Heads# | 100 (30/30) | 26.7 (8/30) | <0.001 |
At 7 dpi through an infectious blood meal, the midguts of MTB and APM mosquitoes were collected and fixed, and the viral antigen was detected using a mouse anti-Dengue complex monoclonal antibody by indirect fluorescence assay (IFA).
At 14 dpi, the heads of both mosquitoes were collected to extract the total RNA. DENV-2 was diagnosed by RT-PCR using primers for the NS5 gene. Amplification of Ae. polynesiensis RPS6 was used to verify the quality of the RNA samples. Significance was shown in Fisher's exact test.
An inhibited viral dissemination to the heads of MTB mosquitoes
Following replication in the midgut, DENV-2 migrates to the mosquito head where it reaches a peak infection 14 dpi [25]. To test whether viral dissemination was affected in MTB mosquitoes, we assayed the viral infection at this time point by RT-PCR and compared the head infection rate between MTB and APM mosquitoes. As shown in Table 1, 100% (30/30) of APM heads were positive whereas viruses were only detected in 26.7% (8/30) of MTB heads (Fisher's Exact Test, P<0.001). This indicates the possibility that viral dissemination to mosquito heads is significantly reduced in MTB mosquitoes as compared with APM mosquitoes.
Inhibition of viral transmission potential of MTB mosquitoes
To determine whether viral transmission potential is reduced in MTB mosquitoes, we measured infectious viral particles in the saliva released during feeding from the proboscises of mosquitoes at 14 dpi by plaque assay. Mosquitoes were infected through either oral feeding with DENV-2 infected blood or by intrathoracic injection with cell medium containing DENV-2. When mosquitoes were infected through a bloodmeal, 21.4% (6/28) of the APM saliva samples were positive for the envelope protein of DENV-2, whereas DENV-2 was detected in 3.6% (1/28) of the MTB saliva samples (Chi-Square test, P<0.05) (Fig. 3). In the intrathoracic injection experiment, 100% (8/8) of saliva samples were positive for both mosquito strains, although the viral infection level was significantly lower in MTB than APM mosquitoes (Mann-Whitney U test, P<0.05) (Fig. 3). These results indicate MTB mosquitoes have a lower viral transmission potential than APM mosquitoes.
Figure 3. In vitro assay of DENV-2 transmission by MTB and APM mosquitoes.

At 14 days after a blood meal containing DENV-2 or infection through intrathoracic injection, the wings and legs were removed, and the proboscis of each mosquito was inserted into a feeding solution for 90 min. Solution from each mosquito was analyzed for infectious DENV-2 by plaque assays. Lines indicate the log of the median values. Twenty eight and eight biological replicates were used in oral feedings and intrathoracic injection assays, respectively.
A large increase in Wolbachia density in the somatic tissues of MTB mosquitoes
We previously found that Wolbachia can induce resistance to dengue infection in a Wolbachia-density dependent and a tissue-specific manner [18]. To test whether the above viral inhibition is caused by an increased Wolbachia density in the somatic tissues of MTB mosquitoes, we compared the Wolbachia density between MTB and APM mosquitoes using qPCR. We found that the Wolbachia density is significantly higher in the salivary glands (5,738-fold), fat bodies (68-fold), and midguts (269-fold) of MTB mosquitoes as compared to APM mosquitoes, while no differences were observed in the ovaries (1-fold) of APM and MTB mosquitoes (Fig. 4). These results suggest that the high density of Wolbachia in somatic tissues of MTB mosquitoes may contribute directly to their low susceptibility for DENV.
Figure 4. Resistance of MTB mosquitoes to DENV-2 is associated with a high density of Wolbachia in mosquito somatic tissues.
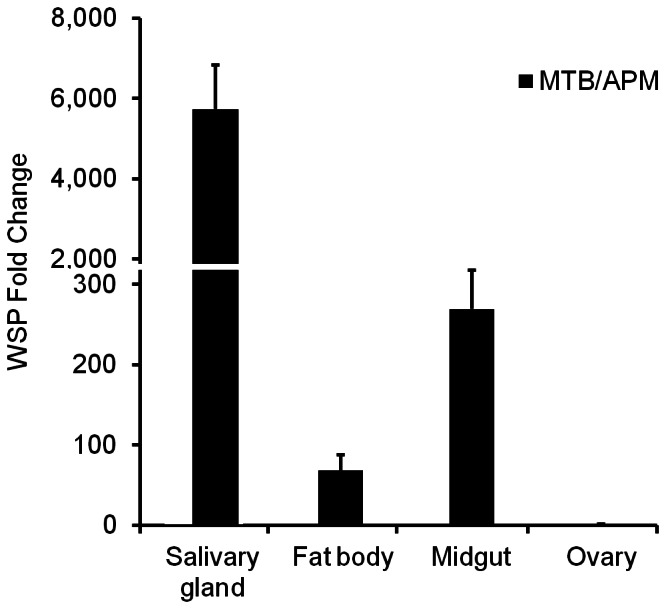
The fold change in genome copy of the Wolbachia surface protein (WSP) gene in MTB mosquitoes is compared to APM mosquitoes. The copy number of the Wolbachia wsp was normalized by Ae. polynesiensis RPS6. In all the assays, the midguts, salivary glands, fat bodies, and ovaries of 7-day-old non-blood-fed females were dissected and used for extraction of total genomic DNA. Error bars are standard errors of the mean of twelve biological replicates.
Discussion
The Wolbachia strain wAlbB is able to induce resistance to DENV in both mosquito vectors and cell lines [6], [18]. After the native wPolA was replaced with wAlbB in Ae. polynesiensis, we observed that wAlbB inhibits dengue viral replication in mosquito midguts and dissemination to mosquito heads. This artificial infection also reduces viral transmission potential through mosquito bites and suppresses the viral infection level in the whole bodies of mosquitoes. Our results further show that the density of wAlbB is 269-fold and 5,738-fold higher in the midguts and salivary glands, respectively, of MTB mosquitoes than the native wPolA infection in APM mosquitoes. This supports the previous finding that the strength of Wolbachia-mediated viral inhibition depends on the Wolbachia density [18].
Although it naturally carries a Wolbachia infection, Ae. polynesiensis is a compatible vector for both DENV and Wuchereria bancrofti in South Pacific [9], [12]. This is similar to Ae. albopictus and Culex quinquefasciatus in that the presence of the native Wolbachia infection in mosquitoes does not inhibit their abilities as vectors to transmit human pathogens. Evidence supports that Wolbachia-mediated viral inhibition occurs in a tissue-specific manner with the effect dependent on the Wolbachia density in the local tissue [18]. In a host cell with a very low level of Wolbachia infection, coexistence of Wolbachia and DENV can be observed in the cytoplasm [18]. Before a mosquito can transmit DENV to humans, viruses ingested by a mosquito in a blood-meal must sequentially migrate through and replicate in the midgut and salivary glands. We previously found that the Wolbachia density in both the midgut and salivary gland of Ae. albopictus was too low to induce a resistance to DENV [6], [18]. A similar situation may be true for Ae. polynesiensis. Consistent with this predication, the Wolbachia density in the midgut and salivary gland of the wild type APM strain is hundreds of times lower than that of the resistant MTB strain.
It is interesting to note that the above difference in the Wolbachia density between APM and MTB mosquitoes occurs only in the somatic tissues, and not the reproductive tissues (ovaries). This distribution pattern could be due to the strain of Wolbachia or the history of Wolbachia-host association. Previous studies showed a Wolbachia strain-specific distribution in insect hosts [33]. The native Wolbachia in APM mosquitoes may not be able to develop an infection at as high a level as wAlbB in somatic tissues. Alternatively, the native infection may have initially been at high levels, similar to wAlbB in the somatic tissues of MTB mosquitoes, but was gradually reduced to the current level in APM mosquitoes as Wolbachia and host co-adapted during evolution. Previous studies show a long-term attenuation of wMelPop in a non-native host, Drosophila simulans [34]. If wAlbB behaves similarly, it may compromise the effectiveness of this bacterium in pest control.
Our results indicate that Wolbachia in midguts can have a significant impact in the development of mosquito resistance to DENV. DENV has to pass the midgut infection barrier with a peroral infection. When infected by an intrathoracic injection, viruses bypass the midgut infection barrier and directly disseminate through the hemolymph. We observed a 1.4×104 –fold reduction in the viral infection in the whole bodies of MTB mosquitoes as compared to those of APM mosquitoes with a peroral infection by DENV-2, while just a 28.3-fold reduction was observed for the same comparison with the viral intrathoracic injection. In addition, only 3.6% of MTB saliva was dengue-positive with the viral peroral infection, as compared to 100% of MTB saliva positive with the viral intrathoracic injection. The above results suggest that Wolbachia-induced viral interference in midguts may contribute to the majority of the virus-blocking effect in MTB mosquitoes.
The fact that Wolbachia-mediated resistance has a broad spectrum against a variety of pathogens in Ae. aegypti indicates that general killing mechanisms should be triggered by Wolbachia [7], [8], [16], [26]. Similarly, wAlbB is also found to suppress filarial worm loads in MTB mosquitoes as conducted by a parallel study in a recent publication [14]. Thus, wAlbB is the second example showing that Wolbachia can interact with mosquito hosts to inhibit both DENV and worm infection. We previously found that Wolbachia can induce ROS production, which activates the Toll pathway to control DENV in Ae. aegypti [26]. When the levels of H2O2 were compared before blood meal, there was a significantly higher level of ROS in MTB mosquitoes than in APM mosquitoes [14]. This suggests that a high density of wAlbB induces more oxidative stress in MTB mosquitoes than the native wPolA does in APM mosquitoes, consistent with our prediction that a high ROS production can contribute to the antiviral resistance [26].
Until now, wAlbB has been observed inducing resistance to DENV in the Ae. albopictus Aa23 cell line [18], Ae. aegypti [6], and Ae. polynesiensis mosquitoes. Among them, wAlbB is a native infection in Aa23 cells, but an artificial infection in Ae. aegypti and Ae. polynesiensis. It is important to note here that the native wAlbB infection can completely clear DENV in Aa23 cells [18]. This suggests that wAlbB has the ability to block DENV in a host cell when the cellular physiological environment is optimized for wAlbB to grow, such that it reaches a high infection level. Until now, both wMel and wMelPop were reported to induce complete blockage of DENV transmission in Ae. aegypti [4], [7], while wAlbB induces a strong resistance to DENV with some viruses leaking from its inhibition in Ae. aegypti [6]. Considering that experimental methods, viral strains and mosquito strains are different in these studies, a direct comparison of viral interference and fitness between wAlbB and wMel/wMelPop in the same host genetic background is needed to ascertain which Wolbachia strain is better suited for use in dengue control. From the disease control standpoint, an ideal strain of Wolbachia should be able to block pathogens in mosquitoes effectively enough to interrupt the disease transmission, and also be benign enough to the mosquito hosts that it can be persistently maintained in populations. It would be interesting to know if a perfect viral blockage is essential for disease control, and what extent of the host fitness cost associated with Wolbachia is acceptable.
Previous studies show Wolbachia-host interactions are determined by the Wolbachia strain, the host genotype, and the environment [35], [36], [37], [38]. It is still unclear why some native Wolbachia are unable to induce pathogen interference in their original hosts, although it appears related to their density. Although the artificial infections all reported inducing a resistance to pathogens [4], [6], [7], [8], this does not mean that any strain of Wolbachia, when introducing into a new mosquito host, will induce pathogen interference. We also cannot exclude the possibility that certain Wolbachia-host associations may even facilitate pathogen infection in an insect vector [39]. Those effects are determined by how a mosquito physiological system is perturbed or modified by its interaction with each specific strain of Wolbachia. The outcome becomes even more of a challenge to predict when considering that Wolbachia-host interactions may evolve over time. Despite these gaps in our knowledge, our results show replacement of native Wolbachia with a novel infection is a potentially practical way to develop pathogen resistant mosquitoes for modification of those vector populations.
Both the ability of wAlbB to induce resistance to filarial worms and DENV and the bi-directional nature of CI between wAlbB and wPolA in Ae. polynesiensis make it possible to develop MTB line as a tool to block disease transmission through population replacement. This result can be achieved by large-scale releases of MTB females. Alternatively, a strategy can be designed to start with population suppression by inundative releases of MTB males, followed by release of MTB females to initiate population replacement. Technically, the latter will be more acceptable to the public because males do not feed on human blood or transmit disease. When the population is then suppressed to a low density, the number of females that need to be released will be reduced. This would make population replacement happen parallel to a decrease in annoyance by mosquito biting. However, its deployment would require the development of a system with a high efficacy for mass rearing and sex separation, which is an ongoing effort in the field.
Acknowledgments
We thank S. L. Dobson for providing us with the APM and MTB strains of Ae. polynesiensis, G. Dimopoulos for supplying the DENV-2 and the C6/36 cell line, M. Weill for providing the pQuantAlb plasmid, and the Arbovirus Diseases Branch at the CDC for providing the anti-dengue antibodies (mouse hyperimmune ascitic fluid). We are also grateful to S. M. Thiem and Melissa Huang for editing this manuscript.
Funding Statement
This work was supported by a grant (R01 AI-080597) from the National Institutes of Health/National Institute for Allergy and Infectious Disease, and the Bill and Melinda Gates Foundation (#44190). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hilgenboecker K, Hammerstein P, Schlattmann P, Telschow A, Werren JH (2008) How many species are infected with Wolbachia?–A statistical analysis of current data. FEMS Microbiol Lett 281: 215–220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Werren JH, Baldo L, Clark ME (2008) Wolbachia: master manipulators of invertebrate biology. Nat Rev Microbiol 6: 741–751. [DOI] [PubMed] [Google Scholar]
- 3. Xi Z, Khoo CC, Dobson SL (2005) Wolbachia establishment and invasion in an Aedes aegypti laboratory population. Science 310: 326–328. [DOI] [PubMed] [Google Scholar]
- 4. Walker T, Johnson PH, Moreira LA, Iturbe-Ormaetxe I, Frentiu FD, et al. (2011) The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 476: 450–453. [DOI] [PubMed] [Google Scholar]
- 5. Hoffmann AA, Montgomery BL, Popovici J, Iturbe-Ormaetxe I, Johnson PH, et al. (2011) Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 476: 454–457. [DOI] [PubMed] [Google Scholar]
- 6. Bian G, Xu Y, Lu P, Xie Y, Xi Z (2010) The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. PLoS Pathog 6: e1000833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu G, Pyke AT, et al. (2009) A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 139: 1268–1278. [DOI] [PubMed] [Google Scholar]
- 8. Kambris Z, Cook PE, Phuc HK, Sinkins SP (2009) Immune activation by life-shortening Wolbachia and reduced filarial competence in mosquitoes. Science 326: 134–136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Rosen L, Rozeboom LE, Sweet BH, Sabin AB (1954) The transmission of dengue by Aedes polynesiensis Marks. Am J Trop Med Hyg 3: 878–882. [DOI] [PubMed] [Google Scholar]
- 10. Gratz NG (2004) Critical review of the vector status of Aedes albopictus. Med Vet Entomol 18: 215–227. [DOI] [PubMed] [Google Scholar]
- 11. Lambrechts L, Scott TW, Gubler DJ (2010) Consequences of the expanding global distribution of Aedes albopictus for dengue virus transmission. PLoS Negl Trop Dis 4: e646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Brelsfoard CL, Sechan Y, Dobson SL (2008) Interspecific hybridization yields strategy for South Pacific filariasis vector elimination. PLoS Negl Trop Dis 2: e129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Armbruster P, Damsky WE Jr, Giordano R, Birungi J, Munstermann LE, et al. (2003) Infection of New- and Old-World Aedes albopictus (Diptera: Culicidae) by the intracellular parasite Wolbachia: implications for host mitochondrial DNA evolution. Journal of medical entomology 40: 356–360. [DOI] [PubMed] [Google Scholar]
- 14. Andrews ES, Crain PR, Fu Y, Howe DK, Dobson SL (2012) Reactive oxygen species production and Brugia pahangi survivorship in Aedes polynesiensis with artificial Wolbachia infection types. PLoS Pathog 8: e1003075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Xi Z, Khoo CC, Dobson SL (2006) Interspecific transfer of Wolbachia into the mosquito disease vector Aedes albopictus. Proc Biol Sci 273: 1317–1322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Rances E, Ye YH, Woolfit M, McGraw EA, O'Neill SL (2012) The relative importance of innate immune priming in Wolbachia-mediated dengue interference. PLoS Pathog 8: e1002548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Glaser RL, Meola MA (2010) The native Wolbachia endosymbionts of Drosophila melanogaster and Culex quinquefasciatus increase host resistance to West Nile virus infection. PLoS One 5: e11977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Lu P, Bian G, Pan X, Xi Z (2012) Wolbachia induces density-dependent inhibition to dengue virus in mosquito cells. PLoS Negl Trop Dis 6: e1754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Mousson L, Zouache K, Arias-Goeta C, Raquin V, Mavingui P, et al. (2012) The Native Wolbachia Symbionts Limit Transmission of Dengue Virus in Aedes albopictus. PLoS Negl Trop Dis 6: e1989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Blagrove MS, Arias-Goeta C, Failloux AB, Sinkins SP (2012) Wolbachia strain wMel induces cytoplasmic incompatibility and blocks dengue transmission in Aedes albopictus. Proc Natl Acad Sci U S A 109: 255–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Beerntsen BT, James AA, Christensen BM (2000) Genetics of mosquito vector competence. Microbiol Mol Biol Rev 64: 115–137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Xi Z, Ramirez JL, Dimopoulos G (2008) The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog 4: e1000098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Sanchez-Vargas I, Scott JC, Poole-Smith BK, Franz AW, Barbosa-Solomieu V, et al. (2009) Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito's RNA interference pathway. PLoS Pathog 5: e1000299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Souza-Neto JA, Sim S, Dimopoulos G (2009) An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc Natl Acad Sci U S A 106: 17841–17846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Salazar MI, Richardson JH, Sanchez-Vargas I, Olson KE, Beaty BJ (2007) Dengue virus type 2: replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC Microbiol 7: 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Pan X, Zhou G, Wu J, Bian G, Lu P, et al. (2012) Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc Natl Acad Sci U S A 109: E23–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Troyer JM, Hanley KA, Whitehead SS, Strickman D, Karron RA, et al. (2001) A live attenuated recombinant dengue-4 virus vaccine candidate with restricted capacity for dissemination in mosquitoes and lack of transmission from vaccinees to mosquitoes. Am J Trop Med Hyg 65: 414–419. [DOI] [PubMed] [Google Scholar]
- 28. Das S, Garver L, Ramirez JR, Xi Z, Dimopoulos G (2007) Protocol for Dengue Infections in Mosquitoes (A. aegypti) and Infection Phenotype Determination. J Vis Exp 5: e220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Molina-Cruz A, Gupta L, Richardson J, Bennett K, Black Wt, et al. (2005) Effect of mosquito midgut trypsin activity on dengue-2 virus infection and dissemination in Aedes aegypti. Am J Trop Med Hyg 72: 631–637. [PubMed] [Google Scholar]
- 30. Tortosa P, Courtiol A, Moutailler S, Failloux A, Weill M (2008) Chikungunya-Wolbachia interplay in Aedes albopictus. Insect Molecular Biology 17: 677–684. [DOI] [PubMed] [Google Scholar]
- 31.Higgs S, Olson KE, Kamrud KI, Powers AM, Beaty BJ (1997) Viral expression systems and viral infections in insects. In: Crampton JM, Beard, C B., Louis, C., editor. The molecular biology of biology of insect disease vectors, a method manual. London: Chapman & Hall. pp. 459–483.
- 32. Franz AW, Sanchez-Vargas I, Adelman ZN, Blair CD, Beaty BJ, et al. (2006) Engineering RNA interference-based resistance to dengue virus type 2 in genetically modified Aedes aegypti. Proc Natl Acad Sci U S A 103: 4198–4203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Veneti Z, Clark ME, Karr TL, Savakis C, Bourtzis K (2004) Heads or tails: host-parasite interactions in the Drosophila-Wolbachia system. Appl Environ Microbiol 70: 5366–5372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Carrington LB, Hoffmann AA, Weeks AR (2010) Monitoring long-term evolutionary changes following Wolbachia introduction into a novel host: the Wolbachia popcorn infection in Drosophila simulans. Proc Biol Sci 277: 2059–2068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. McGraw EA, Merritt DJ, Droller JN, O'Neill SL (2002) Wolbachia density and virulence attenuation after transfer into a novel host. Proc Natl Acad Sci U S A 99: 2918–2923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Boyle L, O'Neill SL, Robertson HM, Karr TL (1993) Interspecific and intraspecific horizontal transfer of Wolbachia in Drosophila. Science 260: 1796–1799. [DOI] [PubMed] [Google Scholar]
- 37. Werren JH (1997) Biology of Wolbachia. Annu Rev Entomol 42: 587–609. [DOI] [PubMed] [Google Scholar]
- 38. Feder ME, KARR TL, YANG W, HOEKSTRA JM, JAMES AC (1999) Interaction of Drosophila and its endosymbiont Wolbachia: natural heat shock and the overcoming of sexual incompatibility. Am Zool 39: 363–373. [Google Scholar]
- 39. Graham RI, Grzywacz D, Mushobozi WL, Wilson K (2012) Wolbachia in a major African crop pest increases susceptibility to viral disease rather than protects. Ecol Lett 15: 993–1000. [DOI] [PubMed] [Google Scholar]