

Abstract
To identify bacteria with potential for influencing gut health, 980 anaerobes were cultured from the swine intestinal tract and analyzed for butyrate production. Fifteen isolates in the order Clostridiales produced butyrate and had butyryl coenzyme A (CoA):acetate CoA transferase activity. Three of the isolates grew on mucin, suggesting an intimate association with host intestinal mucosa.
TEXT
Butyrate and butyrate-producing microbes have been associated with gastrointestinal health in humans (1–4) and various animal species, including swine (5–7). Despite the likely importance of butyrate-producing microbes to the swine intestinal microbiome, the nature and characteristics of these bacteria remain largely unknown. Consequently, this investigation was aimed at isolating and identifying bacteria producing butyrate in the swine intestine. These are important steps toward our long-range goal of characterizing, monitoring, and manipulating butyrate-producing bacteria in the swine intestinal tract to benefit host animal health and agricultural productivity.
Under anaerobic culture conditions, 980 bacteria were isolated as single colonies from the feces, intestinal contents, and intestinal mucosal scrapings of three sows and their 21- to 108-day-old piglets (8). Various culture media were used to increase the likelihood of selecting diverse taxa (Table 1). These animals were also used in parallel studies (12). Also evaluated for butyrate production were seven mucin-degrading bacterial strains previously isolated from the swine intestinal compartments, including mucosal scrapings (11). The mucin degraders were obtained by serial passage in enrichment broth cultures containing swine gastric mucus and grew in medium containing mucus or mucins as sole carbon sources. After subculture purification by single colony transfers, the isolates were cultured in broth media to maximum growth (optical density at 620 nm [OD620]), and culture supernatants were analyzed for butyrate by gas-liquid chromatography (13). Fifteen isolates that produced ≥5 mM butyrate were studied further (Table 1).
Table 1.
Characteristics of butyrate-producing bacteria isolated from the swine intestine
| Strain (origin)a | Isolation mediumb | Butyrate production |
Butyryl-CoA:acetate CoA transferase |
Closest related species (16S rRNA % identity; accession no.)e | ||
|---|---|---|---|---|---|---|
| Mediumb | Concn (mM)c | Sp actd | PCR assay | |||
| 27-5-10 (P Ce M) | HM | PYM1 | 9.0 ± 0.2 | 1.98 ± 0.02 | Pos | Flavonifractor plautii 265T (95; Y18187) |
| 35-6-1 (P Co M) | HM | PYGM1 | 6.8 ± 0.0 | 4.35 ± 0.10 | Neg | Peptoniphilus gorbachii Wal 10418T (97; DQ911241) |
| 68-3-10 (P Co C) | HM | M2GSC | 7.4 ± 0.0 | 0.07 ± 0.00 | Neg | Eubacterium nodatum VPI D6 A-5T (93; Z36274) |
| 984a (P F) | DRFp | PYGM2 | 4.7 ± 0.5 | 1.07 ± 0.02 | Neg | Megasphaera elsdenii LC-1T (99; NR_029207) |
| 813a (P F) | DRFp | PYGM2 | 4.7 ± 0.6 | 1.30 ± 0.05 | Neg | Megasphaera elsdenii LC-1T (99; NR_029207) |
| 702 (P F) | DRFp | DRF7-4X | 7.1 ± 0.2f | 2.50 ± 0.05 | Neg | Megasphaera elsdenii LC-1T (99; NR_029207) |
| 1161 (P F) | DRFla | DRFla | 11.0 ± 4.5f | 4.79 ± 0.41 | Neg | Megasphaera elsdenii LC-1T (99; NR_029207) |
| 1123 (P F) | DRFla | DRFla | 9.5 ± 0.5f | 1.92 ± 0.02 | Neg | Megasphaera elsdenii LC-1T (99; NR_029207) |
| 739a (P F) | DRFla | DRFla | 8.7 ± 1.9f | 4.18 ± 0.02 | Neg | Megasphaera elsdenii LC-1T (99; NR_029207) |
| 655a (P F) | DRFla | DRFla | 11.8 ± 3.7f | 1.93 ± 0.25 | Neg | Megasphaera elsdenii LC-1T (99; NR_029207) |
| 653a (P F) | DRFla | DRFla | 13.6 ± 2.6f | 3.91 ± 0.09 | Neg | Megasphaera elsdenii LC-1T (99; NR_029207) |
| 992a (P F) | DRF7 | DRF7-4X | 10.3 ± 0.3f | 4.32 ± 0.07 | Pos | Eubacterium hadrum VPI 82-52T (94; FR749934) |
| 494a (S F) | DRF7 | DRF7-2X | 9.0 ± 0.3f | 2.42 ± 0.09 | Pos | Eubacterium hadrum VPI 82-52T (96; FR749934) |
| 831b (P F) | DRF7 | DRF7-2X | 8.0 ± 0.1f | 7.77 ± 0.55 | Pos | Roseburia faecis M72/1T (97; AY305310) |
| 499 (S F) | DRF7 | DRF7-2X | 2.7 ± 0.1f | 3.37 ± 0.04 | Pos | Roseburia hominis A2-183T (95; AJ270482) |
P, postweaning pig, 3 to 15 weeks old; S, adult sow; Ce, cecum; Co, colon; F, feces; M, intestinal mucosa; C, intestinal compartment contents.
DRF, depleted rumen fluid medium (9). Media (1 liter) derived from DRF included DRFla (lactate and acetate, 2.5 g each), DRFp (Casamino Acids and Bacto tryptone, 2.5 g each), DRF7 (d-fructose, maltose, d-glucose, N-acetyl-d-glucosamine, d-ribose, cellobiose, l-arabinose, 0.25 g each). DRF7-2X and DRF7-4X contained, respectively, 2 and 4 times the concentrations of carbohydrates of DRF7. PYGM1 and PYGM2 have been described elsewhere (8). PYM1 is PYGM1 lacking glucose. M2GSC was modified from Miyazaki et al.'s formulation (10) to contain 300 ml depleted rumen fluid and 0.5 g l-cysteine/liter (29). HM medium has been described elsewhere (11).
Triplicate cultures were analyzed.
Expressed as μmoles/min/mg protein.
All are type strains of the species.
Strains also consuming acetate.
Nearly full-length 16S rRNA gene sequences were obtained after PCR amplification of genomic DNA preparations using the primers 8F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-ACGGCTACCTTGTTACGACTT-3′). Based on 16S rRNA gene sequence comparisons (Table 1; Fig. 1) and a >97% identity cutoff for species designation, eight isolates were strains of Megasphaera elsdenii, a commonly isolated swine intestinal anaerobe (14, 15). All M. elsdenii strains were isolated from the feces of piglets within 2 weeks after weaning. The other seven isolates were related to bacterial species in the genera Eubacterium, Anaerostipes, Peptoniphilus, Roseburia, and Flavonifractor (Table 1; Fig. 1). At least five isolates, 27-5-10, 68-3-10, 494a, 992a, and 499, appear to be new bacterial taxa (species or genera). Additional physiological, biochemical, and genetic characterizations, however, will be required to confirm or establish the taxonomic identities of the isolates, other than M. elsdenii strains.
Fig 1.
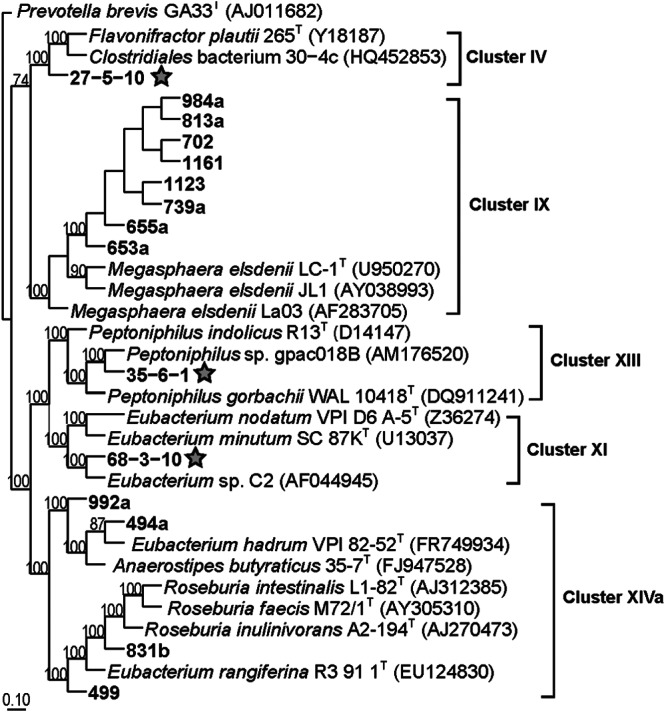
Maximum-likelihood phylogenetic tree of 16S rRNA gene sequences (1,231 bp) of swine butyrate producers (in bold), closely related cultured relatives, and select reference sequences, with superscript T denoting type strains. Clostridial clusters are labeled to the right of the tree. Stars indicates isolates that can degrade mucin. The tree is rooted with the Prevotella brevis GA33 16S rRNA sequence. Numbers by the nodes of the tree indicate the percentage bootstrap values of 1,000 resamplings and are noted only if the percentage was greater than 50%. The scale bar represents 10% sequence divergence.
Butyrate-producing bacteria in the human intestine are predominately members of clostridial clusters IV and XIVa (3, 16, 17). The last enzymatic step for butyrate formation in these anaerobes is commonly butyryl coenzyme A (CoA):acetate CoA transferase (But) (EC 2.8.3.8) rather than butyrate kinase (Buk) (EC 2.7.2.7) (3, 18). Cell-free lysates (prepared using a French press) of all 15 swine strains (Table 1) were negative in an assay for Buk activity (19) and lacked the Buk gene, as determined by PCR assay (18). In contrast, cell-free lysates of every strain were positive in an assay for But activity (20), with activities ranging from 0.07 to 7.77 μmol/min/mg protein (Table 1). Five strains (27-5-10, 494A, 499, 831b, and 992a) had But genes (Table 1), as determined in PCR assays using the previously described But primers (16) and the following amplification conditions: 5 min of denaturation at 95°C; 10 cycles of touchdown PCR of 95°C for 30 s, 53°C for 30 s (−0.4°C per cycle to 49°C), and 72°C for 30 s; 30 cycles of 95°C for 30 s, 49°C for 30 s, and 72°C for 30 s; and a final 10-min extension at 72°C. PCR amplicon sequences confirmed their identity as But genes, and the sequences were deposited in GenBank (accession numbers JX629256 to JX629270).
Notably, isolates 35-6-1 and 68-3-10 and the eight M. elsdenii strains did not have But genes detectable by degenerate PCR assay, although they all had But enzyme activity (Table 1). Previous studies using the same PCR primers for But genes (21, 22) would not have detected M. elsdenii or the two mucinolytic isolates. Our current findings combined with recent analyses of butyrate-producing bacteria from chicken ceca (23) suggest that But gene PCR assay with degenerate primers needs to be improved (broadened) to capture butyrate producer diversity in intestinal microbiomes of swine, chickens, and probably other animals. One approach could be arrays of PCRs using primer sets targeting specific But genes. The availability of bacterial cultures, such as isolates 35-6-1 and 68-3-10 and M. elsdenii strains, for physiological and genomic analyses will facilitate this effort for swine.
All M. elsdenii strains were isolated from piglets shortly after weaning, a finding that is consistent with a previous report of early colonization of piglets by this anaerobe (15). M. elsdenii is a well-known lactate fermenter (to butyrate and propionate), present in ruminant and nonruminant mammalian digestive tracts (24, 25), including swine intestinal mucosal surfaces (26). M. elsdenii colonizes the human intestinal tract (27, 28), although it has not been identified as a numerically dominant butyrate producer in analyses of (adult) human feces (3). To our knowledge, colonization of infants by M. elsdenii remains to be studied.
A high proportion (three of seven) of the previously isolated mucin-degrading bacteria (11) were found to produce butyrate and to be phylogenetically diverse (Table 1; Fig. 1). Another mucinolytic bacterium, strain CL-84T, was isolated from swine in the same study and has been designated a new bacterial species, Cloacibacillus porcorum CL-84T in the phylum Synergistetes (11). C. porcorum CL-84T produces butyrate when metabolizing serine. C. porcorum CL-84T and mucin-degrading isolates 27-5-10 and 35-6-1 were isolated from mucosal surfaces of swine intestinal tissues. In recent in vitro simulations of the human intestinal mucosal environment, mucin agar became specifically colonized by butyrate-producing bacteria of Clostridium cluster XIVa (9). Mucinolytic bacterial species producing butyrate and inhabiting intestinal mucosal surfaces could easily be overlooked in studies of butyrate producers in fecal samples. Both their niche and their microhabitat predict that these bacteria interact intimately with host intestinal tissues, thereby influencing gut health.
Nucleotide sequence accession numbers.
Newly determined 16S rRNA gene sequences have been deposited in GenBank under accession numbers JX629256 to JX629270.
ACKNOWLEDGMENTS
We gratefully acknowledge the excellent technical assistance of Deb Lebo, Sam Humphrey, Darrell Bayles, Loren Jones, and Stephanie Jones. We thank the NADC Animal Care and Use Committee for scrutiny and approval of all procedures involving animals in these studies.
Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Footnotes
Published ahead of print 12 April 2013
REFERENCES
- 1. Van Immerseel F, Ducatelle R, De Vos M, Boon N, Van De Wiele T, Verbeke K, Rutgeerts P, Sas B, Louis P, Flint HJ. 2010. Butyric acid-producing anaerobic bacteria as a novel probiotic treatment approach for inflammatory bowel disease. J. Med. Microbiol. 59:141–143 [DOI] [PubMed] [Google Scholar]
- 2. Roediger WEW. 1980. Role of anaerobic bacteria in the metabolic welfare of the colonic mucosa in man. Gut 21:793–798 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Louis P, Flint HJ. 2009. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 294:1–8 [DOI] [PubMed] [Google Scholar]
- 4. Hamer HM, Jonkers D, Venema K, Vanhoutvin S, Troost FJ, Brummer RJ. 2008. Review article: the role of butyrate on colonic function. Aliment. Pharmacol. Ther. 27:104–119 [DOI] [PubMed] [Google Scholar]
- 5. Torok VA, Hughes RJ, Mikkelsen LL, Perez-Maldonado R, Balding K, MacAlpine R, Percy NJ, Ophel-Keller K. 2011. Identification and characterization of potential performance-related gut microbiotas in broiler chickens across various feeding trials. Appl. Environ. Microbiol. 77:5868–5878 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Kien CL, Blauwiekel R, Bunn JY, Jetton TL, Frankel WL, Holst JJ. 2007. Cecal infusion of butyrate increases intestinal cell proliferation in piglets. J. Nutr. 137:916–922 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kien CL, Blauwiekel R. 2008. Cecal infusion of butyrate does not alter cecal concentration of butyrate in piglets fed inulin. J. Parenter. Enteral Nutr. 32:439–442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Levine UY, Bearson SM, Stanton TB. 2012. Mitsuokella jalaludinii inhibits growth of Salmonella enterica serovar Typhimurium. Vet. Microbiol. 159:115–122 [DOI] [PubMed] [Google Scholar]
- 9. Van den Abbeele P, Belzer C, Goossens M, Kleerebezem M, De Vos WM, Thas O, De Weirdt R, Kerckhof FM, Van de Wiele T. 13 December 2012, posting date Butyrate-producing Clostridium cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. [Epub ahead of print.] doi:10.1038/ismej.2012.158 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Miyazaki K, Martin JC, Marinsek-Logar R, Flint HJ. 1997. Degradation and utilization of xylans by the rumen anaerobe Prevotella bryantii (formerly P. ruminicola subsp. brevis) B(1)4. Anaerobe 3:373–381 [DOI] [PubMed] [Google Scholar]
- 11. Looft T, Levine UY, Stanton TB. 5 October 2012, posting date Cloacibacillus porcorum sp. nov., a mucin-degrading bacterium from the swine intestinal tract and emended description of the genus Cloacibacillus. Int. J. Syst. Evol. Microbiol. [Epub ahead of print.] doi: 10.1099/ijs.0.044719-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Allen HK, Looft T, Bayles DO, Humphrey S, Levine UY, Alt D, Stanton TB. 2011. Antibiotics in feed induce prophages in swine fecal microbiomes. mBio 2(6):e00260-11 doi: 10.1128/mBio.00260-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Salanitro JP, Muirhead PA. 1975. Quantitative method for the gas chromatographic analysis of short-chain monocarboxylic and dicarboxylic acids in fermentation media. Appl. Microbiol. 29:374–381 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Stanton TB, McDowall JS, Rasmussen MA. 2004. Diverse tetracycline resistance genotypes of Megasphaera elsdenii strains selectively cultured from swine feces. Appl. Environ. Microbiol. 70:3754–3757 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Stanton TB, Humphrey SB. 2011. Persistence of antibiotic resistance: evaluation of a probiotic approach using antibiotic-sensitive Megasphaera elsdenii strains to prevent colonization of swine by antibiotic-resistant strains. Appl. Environ. Microbiol. 77:7158–7166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Louis P, Flint HJ. 2007. Development of a semiquantitative degenerate real-time PCR-based assay for estimation of numbers of butyryl-coenzyme A (CoA) CoA transferase genes in complex bacterial samples. Appl. Environ. Microbiol. 73:2009–2012 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Collins MD, Lawson PA, Willems A, Cordoba JJ, Fernandez-Garayzabal J, Garcia P, Cai J, Hippe H, Farrow JA. 1994. The phylogeny of the genus Clostridium: proposal of five new genera and eleven new species combinations. Int. J. Syst. Bacteriol. 44:812–826 [DOI] [PubMed] [Google Scholar]
- 18. Louis P, Duncan SH, McCrae SI, Millar J, Jackson MS, Flint HJ. 2004. Restricted distribution of the butyrate kinase pathway among butyrate-producing bacteria from the human colon. J. Bacteriol. 186:2099–2106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Stanton TB. 1989. Glucose metabolism and NADH recycling by Treponema hyodysenteriae, the agent of swine dysentery. Appl. Environ. Microbiol. 55:2365–2371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Buckel W, Dorn U, Semmler R. 1981. Glutaconate CoA-transferase from Acidaminococcus fermentans. Eur. J. Biochem. 118:315–321 [DOI] [PubMed] [Google Scholar]
- 21. Metzler-Zebeli BU, Zijlstra RT, Mosenthin R, Ganzle MG. 2011. Dietary calcium phosphate content and oat beta-glucan influence gastrointestinal microbiota, butyrate-producing bacteria and butyrate fermentation in weaned pigs. FEMS Microbiol. Ecol. 75:402–413 [DOI] [PubMed] [Google Scholar]
- 22. Metzler-Zebeli BU, Hooda S, Pieper R, Zijlstra RT, van Kessel AG, Mosenthin R, Gänzle MG. 2010. Nonstarch polysaccharides modulate bacterial microbiota, pathways for butyrate production, and abundance of pathogenic Escherichia coli in the pig gastrointestinal tract. Appl. Environ. Microbiol. 76:3692–3701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Eeckhaut V, Van Immerseel F, Croubels S, De Baere S, Haesebrouck F, Ducatelle R, Louis P, Vandamme P. 2011. Butyrate production in phylogenetically diverse Firmicutes isolated from the chicken caecum. Microb. Biotechnol. 4:503–512 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Hespell RB, Akin DE, Dehority BA. 1997. Bacteria, fungi, and protozoa of the rumen, p 59–141 In Mackie RI, White BA, Isaacson RE. (ed), Gastrointestinal microbiology, vol 2 Gastrointestinal microbes and host interactions. Chapman & Hall, New York, NY [Google Scholar]
- 25. Stewart CS. 1997. Microorganisms in hindgut fermentors, p 142–186 In Mackie RI, White BA, Isaacson RE. (ed), Gastrointestinal microbiology, vol 2 Gastrointestinal microbes and host interactions. Chapman & Hall, New York, NY [Google Scholar]
- 26. Stanton TB, Humphrey SB. 2003. Isolation of tetracycline-resistant Megasphaera elsdenii strains with novel mosaic gene combinations of tet(O) and tet(W) from swine. Appl. Environ. Microbiol. 69:3874–3882 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Sugihara PT, Sutter VL, Attebery HR, Bricknell KS, Finegold SM. 1974. Isolation of Acidaminococcus fermentans and Megasphaera elsdenii from normal human feces. Appl. Microbiol. 27:274–275 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Sato T, Matsumoto K, Okumura T, Yokoi W, Naito E, Yoshida Y, Nomoto K, Ito M, Sawada H. 2008. Isolation of lactate-utilizing butyrate-producing bacteria from human feces and in vivo administration of Anaerostipes caccae strain L2 and galacto-oligosaccharides in a rat model. FEMS Microbiol. Ecol. 66:528–536 [DOI] [PubMed] [Google Scholar]
- 29. Allison MJ, Robinson IM, Bucklin JA, Booth GD. 1979. Comparison of bacterial populations of the pig cecum and colon based upon enumeration with specific energy sources. Appl. Environ. Microbiol. 37:1142–1151 [DOI] [PMC free article] [PubMed] [Google Scholar]