

Abstract
One of the remarkable features of networks is module that can provide useful insights into not only network organizations but also functional behaviors between their components. Comprehensive efforts have been devoted to investigating cohesive modules in the past decade. However, it is still not clear whether there are important structural characteristics of the nodes that do not belong to any cohesive module. In order to answer this question, we performed a large-scale analysis on 25 complex networks with different types and scales using our recently developed BTS (bintree seeking) algorithm, which is able to detect both cohesive and sparse modules in the network. Our results reveal that the sparse modules composed by the cohesively isolated nodes widely co-exist with the cohesive modules. Detailed analysis shows that both types of modules provide better characterization for the division of a network into functional units than merely cohesive modules, because the sparse modules possibly re-organize the nodes in the so-called cohesive modules, which lack obvious modular significance, into meaningful groups. Compared with cohesive modules, the sizes of sparse ones are generally smaller. Sparse modules are also found to have preferences in social and biological networks than others.
Introduction
Networks that can describe diverse complex systems are successful tools to understand unknown domains in nature [1]. One of the most interesting topics in the area of complex networks is the module structure and its detection. Modular structure detection has received a considerable amount of attention in various fields because of the significant feature that the nodes in the same module have similar attributes. In the literature, great efforts have been devoted to mining cohesive modules of networks in different fields, including social networks [2] such as collaboration networks, technological networks such as the WorldWide Web [3], [4], and biological networks such as protein-protein interaction (PPI) networks [5], metabolic networks [6], and neural networks [7]. A cohesive module indicates its intra-vertices are densely connected, which at the same time are sparsely connected with the vertices in other modules [3], [8], [9]. Detailed analysis on the meanings of the modules can thus be performed on the mining outputs. For example, in protein-protein interaction networks, modules may contain proteins having similar and specific functions within the cell [5]; in metabolic networks, they likely correspond to functional units such as metabolic pathways [10]; in social networks, individuals tend to form modules of similar hobbies, work environment, family, or friends [2].
Although community detection has been widely studied for a long time [11]–[13], early analysis mainly focuses on small networks, which can be accomplished by human power. In recent times, the size of real networks we can measure has grown considerably, reaching millions or even billions of nodes [9]. This calls for a new theoretical framework that uses computer science to deal with such big data for finding relationship among the nodes. Therefore, a large number of computer algorithm-based methods were proposed. One classical method [14] that aims at identification of edges lying between communities appeared in 2002. Another famous approach is the objective function known as modularity [3], [15], which can be used both to discover communities and to measure their strength. By assumption, high values of modularity indicate good partitions. So, several technologies, such as greedy techniques [15], simulated annealing [16], [17], extremal optimization [18], and spectral optimization [1], [19], have been employed to optimize the modularity. Communities in networks often overlap [10], [20], such that nodes can simultaneously belong to several groups. Clique percolation [10] and its derived CFinder [21] have been used for discovering overlapped nodes. In contrast to the above methods, which pay more attention to clustering nodes, link-based community detection [22] can discover both communities and overlapped nodes successfully.
In general, module detection is implemented based on the modular concept that connections of nodes within the same group are denser than connections with the rest of the network. The question is, are there any sparse modules in which the nodes are sparsely connected internally and densely connected with other sparse or cohesive modules possible in complex networks? Some studies on this issue have been reported in biological networks [23]–[25]. In 2010, using an error function, Pinkert et al proposed an alternative approach which does not consider any prior definitions of what actually constitutes a “module” to detect functional modules in PPI networks [23]. They applied their method to the PPI network derived from the Human Protein Reference Database (HPRD) and found some cohesive modules that proved to be functional modules. In addition, the authors also found some significant non-cohesive clusters, which are functionally related and can provide a better description of the PPI network when combined with the cohesive modules. This finding indicated that we need to extend our traditional concept about the functional unit in a complex network by paying more attention to the sparse modules. By overcoming the resolution limit and over-split phenomena of the alternative approach [24], [25], we proposed a BinTree Seeking (BTS) method based on the Edge Density of Module (EDM) and binary tree theory to mine both sparse and cohesive functional modules in biological networks [24]. Experimental results on three real PPI networks demonstrate that functional modules in PPI networks are not dominantly cohesive but can be sparse. Our studies also show that BTS can achieve the goal of mining both the cohesive and sparse modules simultaneously and automatically.
Based on the results obtained in PPI networks, the motivation of this paper is to study whether it is a general principle that sparse modules co-exist with cohesive modules in the same complex network regardless of its type. In order to answer this question, we firstly used BTS method to mine cohesive and sparse modules from 3 real networks with known modular structures. And then these mined cohesive and sparse modules were analyzed in detail by using known brokers in social networks, software classes in computer software networks and functional units, or metabolic pathways in biological networks. We further applied BTS method to 25 different networks including social, computer software, technological and biological networks. As a result, we detected sparse modules in all 25 networks. Although it seems from the results that it is a general rule for a network to have both cohesive and sparse modules, we find the preferences of sparse modules varied on different types of networks. We also illustrate the spatial organization of some of these sparse modules that are found in the real networks, which show that sparse and cohesive modules sometimes are spatially correlated.
Results
Comparison to Other Methods
Although our major aim is not to illustrate the performance of the BTS method, we compared BTS with three other methods on four networks with known modular structure, which are listed in Table 1. The first is a synthetic network that is generated using an algorithm similar to the one used for the SB Benchmark [26] and composed of 72 nodes and 448 edges. The synthetic network comprises three modules of 16, 32 and 24 nodes. Two of these modules are sparse modules and the third one form a cohesive module. The average degrees of the nodes in these communities are fixed to 16, 8, and 16, respectively (See Figure 1). Links are placed according the designed module structure. 12 out of 16 edges in module 1 (light yellow) are linked to module 2 (light green) and the other edges are connected to module 3 (red). Likewise, except for those edges that are connected to module 1, the rest of edges in module 2 are placed between module 2 and 3. For the cohesive module, most edges are connected to intra-module except links that are connected to modules 1 and 2 (The synthetic network data and code are shown in the supplement). The other three real networks are Davis’s southern woman [27], Scottish corpor. interlocks [28], and Jung networks [29] respectively. The Davis network is a well-known bipartite network, which describes the relationship of social collaborations between women in Natchez Mississippi. The other bipartite network is Scottish that supports corporate interlocks in Scotland between 1904 and 1905. The last network is a technological network, where nodes represent software classes and edges correspond to different types of dependencies among them (e.g. inheritance, parameters, variables etc.).
Table 1. Performance comparison of four methods on 4 networks.
| Network | Nodes | Edges | Number of Real Modules | BTSa | NLb | Infomoda | Pinkertb |
| NMI (Number of modules) | |||||||
| Synthesis | 72 | 448 | 3 | 0.646(6) | 0.423(3) | 0.533(4) | 0.275(3) |
| Davis | 32 | 89 | 4 | 0.666(2) | 0.818(4) | 0.669(2) | 0.665(4) |
| Scottish | 228 | 358 | 9 | 0.565(5) | 0.275(9) | 0.122(7) | 0.1536(9) |
| Jung | 398 | 943 | 38 | 0.588(35) | 0.591(38) | 0.537(6) | 0.451(38) |
The number of clusters in the network is determined automatically by the algorithms.
The number of clusters in the network is set according to the number of real modules beforehand.
Figure 1. Generated synthetic network of this study.

Different types of modules in these four networks were detected by BTS method. In addition, three methods that deal with the detection of block structures in networks were also applied to these networks. Among them are the mixture model method (NL method) [30] proposed by Newman et al, and the module detection method (Infomod method) [31] by optimizing the function of minimum description length principle. The third compared method is an alternative approach (Pinkert method) [23] that optimizes an error function. Modules detected by the four methods are first compared with known metadata, and then the compared outcomes are measured by normalized mutual information (NMI) [32]. High values of NMI indicate good partitions. The results are presented in Table 1. Note that the parameter of the number of modules is set to the actual number of modules in the NL and Pinkert methods.
From Table 1, BTS performed better than the other methods except on the Davis network, where Infomod performed slightly better and NL performed significantly better than all others due to the number of real modules passed in as input. Overall, the Pinkert method performed poorly.
The Meaning of Cohesive and Sparse Modules in Various Types of Networks
Although finding the coexistence of cohesive and sparse modules in complex networks is the aim of this study, another more significant task is to reveal their meaning in the complex system. To answer the question whether or not the discovered both sparse and cohesive modules are interesting, we applied BTS to social, computer software and biological networks, respectively, and then analyzed the output modules in detail.
Cohesive and sparse modules in social networks
A social network can represent a set of people or groups of people with some relationships between them [33]. These relationships may include friendships between individuals, business partners between companies and intermarriages between families. In contrast to the people in cohesive communities, brokers who trade over gaps in social structure [34], [35] must be credibly connected to actors, but they may not be connected to each other [36]. There are two significant features of brokers. One is that they bridge a gap in social structure, and the other is that they promote information, goods, opportunities, or knowledge flow across that gap [37]. In this section, we applied the BTS method to detect brokers in the well known Newcomb Fraternity network, which contains 17 students living together in a hostel. The development of the network was followed up for 15 weeks except for a holiday break of one week between weeks 9 and 10. In every week, the students were consulted on their friendship preference with the other 16 students [38]. In order to conveniently use BTS method to mine brokers in the Newcomb Fraternity, we transformed it into a network including nodes that represent students and edges that correspond to friendships in the following role: for a student in every week, five links are placed among the student and other five students who are ranked in the top five. Note that an edge that is placed between two students only means that the two students have a strong friendship, does not mean they are unacquainted. According to the BTS consequences (see supporting information at: www.csbio.sjtu.edu.cn/bioinf/SparseNetwork/for more details), in week 8, these students are divided into 5 groups, where groups 4 and 5 are sparse groups including nodes 9 and 17, 13 and 16 respectively. These results are largely consistent with the analysis by Taube [38] that nodes 9, 13 and 17 are coordinators who may mediate contradictions among students. Another example is the network generated by week 13 data. The modular results detected by BTS contain only 1 sparse group, and nodes 13 and 17, who are coordinators, are all in this cluster. Likewise, as a coordinator, node 17 appears in sparse modules in networks generated by weeks 14 and 9 data. We furthermore find that node 17 is observed in many sparse modules corresponding to the 15 week networks, meaning that it plays a significant role in these 17 students.
Cohesive and sparse modules in computer software networks
We further used BTS method to analyze a Jung network (in Table 1) in software systems in which the actual structure remains great unknown [29]. In the software network, nodes correspond to software classes and edges represent different types of dependencies among them (e.g., inheritance, parameters, variables etc.) [39]. As a result, we found 14 cohesive and 21 sparse modules respectively. For these modules, we analyzed some modules detected by BTS in detail below. Module 1 with 33 nodes is a cohesive group (See Table 2 and supplement information for detailed results), and 23 of them consist of Jung ‘graph’ class, and the other 10 nodes contain Jung ‘util’ class. As one could anticipate, the nodes in the module are densely connected internally and sparsely connected to the rest of the network. The second cohesive module we analyzed is module 20 that includes 18 nodes, where 13 nodes are divided into the ‘algorithms.scoring’ class and the rest of nodes are clustered into ‘algorithms.shortestpath’, ‘graph’, and ‘io’ classes (more details see Table 2). Three sparse modules are discussed next. Module 5 is a sparse cluster and its size is 20. The surprising result is that all the nodes in module 5 belong to Jung ‘visualization’ class. 9 and 6 nodes in the module 5 participate in ‘visualization.control’ and ‘visualization.renderers’ classes respectively. These results indicate that the nodes in this sparse module detected by BTS share similar attributes. The other two small sparse modules are module 14 and module 15 with 6 and 5 nodes respectively. 4 of 6 nodes in the module 14 contain ‘algorithms.layout3d’ class. In module 15, 4 nodes belong to ‘algorithms.importance’ class. As revealed by the network topology, this module has no intra-group links, but links with other modules, such as sparse modules 14 and 16.
Table 2. Analysis of the main modules in Jung network.
| Communitynumber | Type | Size | Description |
| 1 | cohesive | 33 | [jung.graph].*(23), .util.*(10). |
| 20 | cohesive | 18 | [jung.algorithms.scoring].*(13), .shortestpath.*(2); [jung.graph].Hypergraph(1); [jung.io].GraphReader(1), .graphml.GraphMLReader2(1). |
| 5 | sparse | 20 | [jung.visualization].*(2),.renderers.*(6),.control.*(9),.annotations.*(2),.transform.LensSupport(1). |
| 14 | sparse | 6 | [jung.algorithms].layout3d.*(4),.flows.EdmonskarpMaxFlow(1),.importance.AbstractRanker(1). |
| 15 | sparse | 5 | [jung.algorithms].importance.*(4),.shortestpath.ShortestPath(1). |
The detailed names of classes are omitted (refer to supporting information at www.csbio.sjtu.edu.cn/bioinf/SparseNetwork/for details).
Cohesive and sparse modules in biological networks
In PPI networks, functional subunits or protein complexes generally correspond to modular structures [40]. Recent literatures confirm that sparse clusters that contain few or no edges can form functional units, indicating functional units are not necessarily cohesive modules. In this study, we extend this viewpoint to gene co-expression networks, which are composed of nodes corresponding to genes and edges that represent significant co-expressed relationships between genes [41], [42]. Since genes on the same pathway or have related functions often exhibit similar expression patterns under diverse conditions in DNA microarray experiments [42], therefore, most of works on functional modules or units detection in gene co-expression network pay more attentions on cohesive modules [43]–[45], and rarely on sparse modules. In this section, we first employed BTS method to detect both cohesive and sparse modules in gene co-expression networks, and then discussed the meaning of these modules.
The genes that are used to construct gene co-expression networks are collected from Arabidopsis thaliana metabolic pathway data [46] (www.arabidopsis.org/). By removing repeated genes in the same pathway and pathways with less than 5 genes (in order to avoid many small clusters), we finally obtained 174 pathways that contain 1725 genes. For these 1725 Arabidopsis genes, Arabidopsis gene co-expressed data consisting of 20906 files from the ATTED-II database [47] (http.//atted.jp/) was used to identify their co-expressed relationships. Co-expression was measured using Pearson’s Correlation Coefficients (PCCs). If the PCC of any two genes is higher than 0.6, a link is placed between these two genes. At last, a gene co-expression network with 793 Arabidopsis genes and 10184 edges was constructed (see supporting information).
Using BTS, we detected 14 cohesive and 21 sparse modules from the network. To demonstrate their importance, we compared them with Arabidopsis metabolic pathway data based on the hypothesis that the genes belonging to the same pathway are highly co-expressed [48], [49]. In addition, the functions of these modules can also be measured by BiNGO [50], which is used to assess a set of genes with Gene Ontology (GO) annotations [51]. The cohesive group of Module 5 has 64 nodes, 22 of which participate in the adenosyl-L-methionine cycle. For the rest of genes in this group, 11 nodes and 12 nodes belong to the metabolic pathways of zeatin biosynthesis and galactose degradation respectively. From these results, we can clearly see that most of the genes in Module 5 actively participate in biosynthesis and degradation of adenosyl-L-methionine. Likewise, using BiNGO, we further find that most genes in this module are involved in acetyl-CoA biosynthesis with a low P-value (P-value = 5.0703E-12, Biological Process (BP)).
Besides cohesive groups, there are also some sparse modules found by BTS. For instance, Module 7 is a sparse module containing 8 nodes. 4 of these 8 nodes are related to the metabolic pathway of cutin biosynthesis, 2 nodes and 1 node belong to chorismate biosynthesis and zeatin biosynthesis respectively. These genes participate in biosynthesis and glucose catabolic process with a significant P-value of 1.8822E-9. Another sparse group, Module 1, has 29 genes. These genes participate in different metabolic pathways, but are significantly enriched in small molecule metabolic process (P-value = 4.7619E-10). These results demonstrate again that sparse modules not only form significant functional units or participate in metabolic pathways, but also can reveal important hidden relationships among nodes in the network.
Sparse Modules Co-exist with Cohesive Ones in Complex Networks
To better understand coexistence of cohesive and sparse modules in various complex networks, we further applied BTS approach to 25 networks with different scales and types (Table 3 shows the details). It is revealed by Table 4 (see supporting information for more details) that sparse modules are prevalent in all 25 networks rather than being isolated to specific networks. These results suggest that both cohesive and sparse modules characterize better functional units or modular structure of a complex network than cohesive modules alone. The nodes’ similar functions in a cohesive module are reflected by the direct links among them, while functions in sparse modules are exhibited by indirect linking and depending on other modules. Therefore, the relationship between different nodes should be evaluated by both types of modules.
Table 3. Descriptions of 25 networks studied in this paper.
| Network Name | Node | Edge | Ref | Description |
| Csphd (S) | 1384 | 1703 | [54] | PH.D. students to their advisors network |
| Erdos (S) | 492 | 1417 | [55] | Erdos collaboration network |
| Football (S) | 115 | 615 | [14] | Network of American football games between Division IA colleges |
| Lsle_of_Man (S) | 675 | 2007 | [54] | The British lsle of Man family of history |
| Jazz (S) | 198 | 2742 | [18] | Jazz musicians network |
| Science (S) | 1589 | 2742 | [56] | A coauthorship network of scientists |
| Collaboration (S) | 5242 | 14490 | [57] | Scientific collaboration network |
| Roget (S) | 1022 | 5075 | [58] | Roget’s thesaurus of English words and Phrases |
| Geom (S) | 7343 | 11898 | [54] | Collaboration network in computational geometry |
| Java (C) | 1538 | 7817 | [55] | Java dependency network |
| A00 (C) | 352 | 384 | [55] | A software project of classes and relationships |
| A96 (C) | 1096 | 1677 | [55] | Finite automaton network |
| C98 (C) | 112 | 168 | [54] | Theorethical graph network |
| Jung(C) | 398 | 943 | [29] | Jung 2.0.1 framework network |
| E-mail (T) | 1133 | 5451 | [59] | Network of E-mail interchanges |
| Odlis (T) | 2909 | 16380 | [60] | Online dictionary of library and information science network |
| SmallW (T) | 396 | 994 | [61] | Citation network produced by HisCite software |
| Polbook (T) | 105 | 441 | Network of books sold by online bookseller | |
| Power (T) | 4941 | 6594 | [62] | Power grid network |
| Usair (T) | 332 | 2126 | [54] | United States air line |
| Yeast PIN (B) | 2361 | 6646 | [63] | Protein interaction network in budding yeast |
| KPI (B) | 887 | 1844 | [64] | Protein kinase and phosphatase interaction network |
| DIP yeast (B) | 2147 | 4275 | [65] | Protein interaction network in yeas |
| BIND human (B) | 3724 | 8748 | [66] | Protein interaction network in human |
| Gene co-expression(B) | 793 | 10184 | Gene co-expression network in Arabidopsis | |
(S), (C), (T), and (B) indicate social network, computer software network, technological network and biological network, respectively.
Table 4. List of modules detected by BTS method in 25 complex networks.
| Network name | Cohesive modules | Sparse modules | Total modules |
| Csphd | 5 | 13 | 18 |
| Erdos | 8 | 16 | 24 |
| Football | 15 | 6 | 21 |
| Isle_of_Man | 1 | 9 | 10 |
| Jazz | 5 | 12 | 17 |
| Science | 7 | 17 | 24 |
| Collaboration | 6 | 19 | 25 |
| Roget | 16 | 10 | 26 |
| Geom | 4 | 16 | 20 |
| Java | 9 | 19 | 28 |
| A00 | 8 | 6 | 14 |
| A96 | 7 | 17 | 24 |
| C98 | 3 | 7 | 10 |
| Jung | 14 | 21 | 35 |
| 7 | 17 | 24 | |
| Odlis | 6 | 16 | 22 |
| SmallW | 3 | 2 | 5 |
| Polbook | 7 | 8 | 15 |
| Power | 5 | 19 | 24 |
| Usair | 5 | 9 | 14 |
| Yeast PIN | 5 | 9 | 14 |
| KPI | 8 | 21 | 29 |
| DIP yeast | 26 | 33 | 59 |
| BIND human | 26 | 39 | 65 |
| Gene co-expressed | 14 | 21 | 35 |
To rationalize the existence of sparse modules, we compared the outputs from BTS and those from the state-of-the-art cohesive-specific module detection approach of Newman-fast algorithm [15] because we do not know the modular structure of these networks. From ref. [52], we know that cohesive modules with less than 10 nodes possibly lack obvious modular significance. However, to provide a full network-based view of the modules, both cohesive modules detected by Newman-fast algorithm of larger and smaller than 10 nodes were considered. For our statistics, these modules are defined as large and small modules respectively. We then can analyze the relationship between the outputs from BTS and Newman method to reveal the inherent mechanism of the sparse module.
We first compared the results on the A00 network that has 352 nodes and 384 edges. Figure 2 shows the nodes distributions in different types of modules identified by the two methods. In Newman-fast, all the detected modules are cohesive modules; While BTS outputs, both cohesive and sparse modules. As shown in Figure 2, the total number of nodes in the small modules from the Newman-fast approach is 110 and the remaining 242 nodes are located in the large modules. At the same time, we found 138 and 214 nodes in the sparse and cohesive modules respectively from the BTS algorithm. By systematically comparing these outputs, we find that there is an intersection of 98 nodes between the small modules and the sparse modules, and an intersection of 202 nodes between the large modules from Newman-fast approach and the cohesive modules from BTS algorithm. It is also worth mentioning that although there are a large number of overlapped nodes between the small modules from Newman-fast method and the sparse modules from BTS, their node component organizations are quite different. The reason for the differences is that the small modules are subjective to the cohesive definitions in the Newman-fast method, while in the BTS, sparse modules are totally diverse from the cohesive definition [24].
Figure 2. Distributions of nodes in A00 network mined by BTS method and Newman-fast algorithm.

These findings indicate the following: (1) the core set of nodes in the cohesive modules can be well identified by both the cohesive-specific Newman-fast algorithm and the BTS method that can mine both dense and sparse modules; (2) although the modules with less than 10 nodes from the Newman-fast approach possibly lack obvious cohesive modular significance [52], these nodes can be potentially re-organized into sparse modules of important functional units, which should be investigated in a different way from the traditional cohesive-specific approach.
As a second example, let’s analyze modular structures of science network, which describes the cooperative relationship among scientists working on network theory and experiment. This network includes 1461 nodes and 2742 edges (128 isolated nodes are not considered). As a result, 24 modules and 275 modules were mined by BTS method and Newman-fast algorithm respectively. According to BTS, 1140 nodes are divided into sparse modules, and 321 nodes belong to cohesive modules. In the Newman-fast algorithm, 887 nodes belong to small cohesive modules, and the remaining 574 nodes belong to large cohesive modules. By comparing these results, we got a considerable intersection composed of 836 nodes between the 887 nodes of the small modules from Newman-fast algorithm and 1140 nodes in the sparse modules from BTS approach. In addition, 270 nodes are overlapped between the 574 nodes in the large modules from Newman-fast algorithm and 321 nodes in the cohesive modules from BTS.
At last, we compared the results on the Roget network, which has 1022 nodes and 5075 edges. 22 modules including 14 small modules and 8 large modules were mined by Newman-fast algorithm. These small modules contain 47 nodes and the other 975 nodes are in large modules. Using BTS method, we got 16 cohesive modules with 719 nodes and 10 sparse modules with 303 nodes. By comparison, we found that 707 nodes are identical between the cohesive modules from the BTS method and the large modules from the Newman-fast approach.
All these results show that sparse modules co-exist with cohesive ones in various networks. Furthermore, these results also imply that both sparse and cohesive modules can describe better functional groups of a complex network than cohesive modules alone. The reason is that sparse structure can reflect the functional relationship for those nodes in the cohesive modules, which lack obvious traditional modular significance.
Preferences of Sparse Structures
Which type of networks do sparse modules prefer? It is difficult to answer this question accurately, so we will study this problem in a straightforward way. The relative proportion of nodes in sparse and cohesive modules from BTS is showed in Figure 3, from which we can observe the difference among the different types of networks. In biological networks, high proportions of nodes in sparse modules that may correspond to functional units are found in Figure 3. This implies that biological networks have a higher tendency to possess sparse structure or weakly significant cohesive modules, which is consistent with previous findings [23]–[25].
Figure 3. The relative proportions of nodes in different networks from sparse and cohesive modules detected by BTS method.

Significant differences are found in the tested 9 social networks, where 4 of them may show obvious sparse structure (Csphd, Is, Sc, and Geom), and 3 of them have obvious cohesive feature (Fo, Jazz, and Roget). These results reveal that the modular structures in social networks vary very much according to the different relationships represented. For example, in the football network, individuals in a module may communicate with each other frequently or share more related attributes, but exchange rarely between diverse groups. Consequently, those, who share similar attributes, easily form social communities, but few people help information flow across communities. So, few brokers (nodes in sparse modules) are generated in this social network. While, in Csphd and Science networks, in which individuals are engaged in high technology fields, advanced information should be spread across different social communities, and promoting them development fleetly. Therefore, more brokers are needed to perform the task. Another potential reason for this phenomenon in social networks is that current networks are far from complete, and results on a partial network can simply reflect sub-organizations.
Although low proportion of nodes in sparse modules exists in computer software networks shown in Figure 3, it does not mean that sparse modular structures do not exist in them. For instance, we have found some meaningful sparse modules in the Jung and A00 networks as shown above. These results thus seem to imply that sparse modules appear ubiquitous in nature, but have a preference for some types of networks.
Sizes of Sparse Modules
We further study the size of sparse modules in various networks. Figure 4 compares the average sizes of the mined sparse and cohesive modules in 25 networks by BTS. Generally, sparse modules are smaller than cohesive modules in the same network, although 3 exceptions were observed (Csphd, Sc, and KPI). Here, we give a general explanation for this phenomenon because we do not have enough information on these networks. In social networks, people in sparse modules possibly play the role of brokers or mediator who help information flow across communities or mediate contradictions among people. Thus, these individuals may have low proportion of all people in a special industry. But in some social networks (for example Csphd and Science networks) in which people take part in high technology, the proportion can be larger since this industry needs more people to exchange or share advanced information, to promote their development. Compared to social networks, the ratio of sparse module size to cohesive module size is smaller in software and biological networks, which may be due to sparse modules forming functional units or software packages. However, the fact that the average size of sparse modules is smaller than cohesive modules is still not clear and waiting for further studies.
Figure 4. The average sizes of sparse and cohesive modules in various networks.

Possible Organizing Structures of Sparse Modules
We have shown above that sparse and cohesive modules can simultaneously exist in the same network, but the possible spatial structures of sparse modules are still not clear. Here we report two possible organizing structures for sparse modules in complex networks observed from the results on the 25 networks. Figure 5A.1 illustrates the bridge sparse module that links one or more other cohesive modules, and Figure 5A.2 shows a real example from the Science network. It is also worth pointing out that these bridge modules were also found in the biological networks as reported in refs. [23], [24]. Another common organization is different sparse modules interacting with each other as shown in Figure 5B.1, where Figure 5B.2 illustrates a real example from the Geom network.
Figure 5. Two possible organizations of sparse modules in the network.

Conclusion and discussion
Complex networks of people, proteins, webpages, or other elements with some pattern of contacts or interactions between them tend to exhibit modular features, which have been studied deeply by sociologists, mathematicians, and biologists. Based on the principle that elements in the same cohesive module have similar attributes, much effort has been devoted to mining cohesive modules and significant progress has been achieved. Recent studies reveal that networks comprise even more sophisticated modules than traditional cohesive modules [30], [39], [53]. For example, sparse modules in PPI networks have been verified to exist, and to have nodes with similar functions [23]–[25]. Using an extended BTS method that can successfully mine both cohesive and sparse clusters in various types of networks, we analyzed the meanings of cohesive and sparse modules detected from three types (social, computer software and biological) of networks. Furthermore, to better show the ubiquitousness of cohesive and sparse modules coexisting in complex networks, the modular structures of 25 different networks were also investigated. Our results suggest that sparse modules commonly exist with cohesive modules, indicating both types of modules should be analyzed simultaneously in order to reveal the functions of the whole network. We also observed some characteristics of the sparse modules and their possible spatial organizations.
Although meaningful results were obtained, great challenges remain. First, in order to further annotate the functions for the mined sparse modules, more information is needed when constructing the networks. This is particularly urgent in social networks since information for verifying the functions of the mined modules is lacking. Second, many functions for measuring cohesive modules have been proposed. But, few functions were developed to assess both cohesive and sparse modules that are simultaneously mined in the same network. Third, algorithms that are suitable for mining both cohesive and sparse modules should be further investigated especially when dealing with large real-world networks (e.g., more than 10000 nodes).
Datasets and Methods
Datasets
A total 25 complex networks were studied in this paper, which are summarized in Table 3 (See supplement for the websites of these 25 networks). These networks have following features: (1) different types divided into 4 categories of social, computer software, technological and biological networks, (2) different scales varied from 105 to 7,343 nodes, and from 168 to 16,380 edges, (3) different network topologies reflected by the edge densities. In general, a network can be represented as a graph where a node corresponds to an individual or object and an edge to a special relationship or interaction. For example, Csphd network was constructed to describe the relationship between Ph.D students and their advisors in theoretical computer science, which contains 1384 nodes and 1705 edges.
Methods
Unlike previous approaches that mainly extract functional units by identifying cohesive modules, we have recently developed an algorithm called BinTree Seeking (BTS) that can also find sparse modules in PPI networks [24]. Members in a sparse module are defined as sparsely connected internally and densely connected with other sparse or cohesive modules at the same time [24]. By using an adjacency matrix to represent a network, BTS detects modules by depicting edges and nodes simultaneously rather than nodes alone and its derivation procedure is based on matrix primary transpositions. When BTS finally converges, it will generate a binary tree, where each leaf represents a state of possible divisions composed of both cohesive and sparse modules. Then, we can use a kind of evaluation function to measure the qualities of the leaf states so that we can pick up the best formulation, e.g. the leaf corresponding to the lowest error function E [23]. The BTS method not only avoids the drawbacks (such as the resolution limit and over-split phenomena [25]) of previous methods but also has some significant merits. One of them is that the number of modules in a network can be automatically determined in this approach. Thus, both sparse and cohesive modules in 25 different networks were mined by BTS algorithm. For more details about the BTS method, the readers are referred to ref. [24], and the main ideas of BTS are given in the appendix. BTS software is available at: http://www.csbio.sjtu.edu.cn/bioinf/BTS/.
To effectively detect cohesive and sparse modules in various networks, three thresholds of (a
1, a
2, a
3) are introduced in BTS method, which play significant roles. a
1 is the lower limit of the link density of cohesive module, a
2 is the upper limit of link density of sparse module, and a
3 is the lower limit of edge density of bridge matrix required to confirm the existence of bridge matrix. In [24], we have discussed the choice of three thresholds in detail on PPI networks. In order to apply BTS method to various types of networks and detect modules effectively, in addition to the classical values of three thresholds ( ,
,  ,
,  , denoted as (0.7, 1.5, 1 ) in a simple form), we also provide here other five groups of three thresholds: (0.5, 1.3, 1), (0.5, 1.7, 1), (0.85, 1.3, 1), (0.8, 1.4, 1) and (0.5, 1.5, 1). Therefore, one can employ BTS method with these thresholds to mine cohesive and sparse modules, and then select a result corresponding to the smallest error function E value or a result that is consistent with known modular structure. Although one can select arbitrary one group of thresholds to detect modules, we suggest that researchers should first consider the classical values of three thresholds because it can generally yield better results. In this study, 11 of 25 networks, synthetic and gene co-expressed networks used the classical thresholds, and 5 networks employed the group of thresholds (0.5, 1.5, 1). The other 3, 2, and 1 networks adopted three thresholds of (0.85, 1.3, 1), (0.8, 1.4, 1), and (0.5, 1.3, 1) respectively (See supporting information for details). All 15 social networks generated by Newcomb Fraternity data used the thresholds of (0.5, 1.7, 1).
, denoted as (0.7, 1.5, 1 ) in a simple form), we also provide here other five groups of three thresholds: (0.5, 1.3, 1), (0.5, 1.7, 1), (0.85, 1.3, 1), (0.8, 1.4, 1) and (0.5, 1.5, 1). Therefore, one can employ BTS method with these thresholds to mine cohesive and sparse modules, and then select a result corresponding to the smallest error function E value or a result that is consistent with known modular structure. Although one can select arbitrary one group of thresholds to detect modules, we suggest that researchers should first consider the classical values of three thresholds because it can generally yield better results. In this study, 11 of 25 networks, synthetic and gene co-expressed networks used the classical thresholds, and 5 networks employed the group of thresholds (0.5, 1.5, 1). The other 3, 2, and 1 networks adopted three thresholds of (0.85, 1.3, 1), (0.8, 1.4, 1), and (0.5, 1.3, 1) respectively (See supporting information for details). All 15 social networks generated by Newcomb Fraternity data used the thresholds of (0.5, 1.7, 1).
As an improvement over the original BTS method, extended BTS method can also expediently detect modular structure from bipartite networks, such as Divas and Scottish networks. Likewise, detecting bipartite structure also faces the problem of selecting three thresholds. Differing from the problem of three thresholds mentioned above, only one parameter 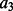 is changed to get better results, and the other two parameters are fixed (
is changed to get better results, and the other two parameters are fixed ( ,
,  ). Figure 6 displays the relationship between
). Figure 6 displays the relationship between  and E values [23]. From Figure 6, we can see that different
and E values [23]. From Figure 6, we can see that different 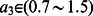 lead to different E values with low fluctuation. So, several values of
lead to different E values with low fluctuation. So, several values of 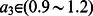 were selected with an interval of 0.05.
were selected with an interval of 0.05.
Figure 6. The relationship between a3 and E value.

Supporting Information
Brief description of BTS algorithm.
(DOC)
Acknowledgments
We thank Dr. Richard Jang for reading through the manuscript, and the anonymous reviewers for suggestions and comments which helped improving the quality of this paper.
Funding Statement
This work was supported by the National Natural Science Foundation of China (No. 91130033, 61175024), Shanghai Science and Technology Commission (No. 11JC1404800), A Foundation for the Author of National Excellent Doctoral Dissertation of PR China (No. 201048), and Program for New Century Excellent Talents in University (NCET-11-0330). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Newman MEJ (2006) Modularity and community structure in networks. Proceedings of the National Academy of Sciences 103: 8577–8582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Girvan M, Newman MEJ (2002) Community structure in social and biological networks. Proceedings of the National Academy of Sciences 99: 7821–7826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Newman MEJ, Girvan M (2004) Finding and evaluating community structure in networks. Physical Review E 69: 026113. [DOI] [PubMed] [Google Scholar]
- 4. Flake GW, Lawrence S, Giles CL, Coetzee FM (2002) Self-organization and identification of web communities. Computer 35: 66–70. [Google Scholar]
- 5. Barabasi AL, Oltvai ZN (2004) Network biology: Understanding the cell’s functional organization. Nature Reviews Genetics 5: 101–U115. [DOI] [PubMed] [Google Scholar]
- 6. Ravasz E, Somera AL, Mongru DA, Oltvai ZN, Barabási AL (2002) Hierarchical organization of modularity in metabolic networks. Science 297: 1551–1555. [DOI] [PubMed] [Google Scholar]
- 7. Park YS, Chon TS, Kwak IS, Lek S (2004) Hierarchical community classification and assessment of aquatic ecosystems using artificial neural networks. Science of the Total Environment 327: 105–122. [DOI] [PubMed] [Google Scholar]
- 8. Ye Z, Hu S, Yu J (2008) Adaptive clustering algorithm for community detection in complex networks. Physical Review E 78: 046115. [DOI] [PubMed] [Google Scholar]
- 9. Fortunato S (2010) Community detection in graphs. Physics Reports 486: 75–174. [Google Scholar]
- 10. Palla G, Derényi I, Farkas I, Vicsek T (2005) Uncovering the overlapping community structure of complex networks in nature and society. Nature 435: 814–818. [DOI] [PubMed] [Google Scholar]
- 11.Coleman JS (1964) Introduction to mathematical sociology. London Free Press Glencoe.
- 12. Moody J, White DR (2003) Structural cohesion and embeddedness: A hierarchical concept of social groups. American Sociological Review 68: 103–127. [Google Scholar]
- 13. Weiss RS, Jacobson E (1955) A method for the analysis of the structure of complex organizations. American Sociological Review 20: 661–668. [Google Scholar]
- 14. Girvan M, Newman MEJ (2002) Community structure in social and biological networks. Proceedings of the National Academy of Sciences 99: 7821–7826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Newman MEJ (2004) Fast algorithm for detecting community structure in networks. Physical Review E 69: 066133. [DOI] [PubMed] [Google Scholar]
- 16. Guimera R, Sales-Pardo M, Amaral LAN (2004) Modularity from fluctuations in random graphs and complex networks. Physical Review E 70: 025101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Guimera R, Amaral LAN (2005) Functional cartography of complex metabolic networks. Nature 433: 895–900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Duch J, Arenas A (2005) Community detection in complex networks using extremal optimization. Physical Review E 72: 027104. [DOI] [PubMed] [Google Scholar]
- 19. Richardson T, Mucha PJ, Porter MA (2009) Spectral tripartitioning of networks. Physical Review E 80: 036111. [DOI] [PubMed] [Google Scholar]
- 20. Palla G, Barabasi AL, Vicsek T (2007) Quantifying social group evolution. Nature 446: 664–667. [DOI] [PubMed] [Google Scholar]
- 21. Adamcsek B, Palla G, Farkas IJ, Derényi I, Vicsek T (2006) CFinder: locating cliques and overlapping modules in biological networks. Bioinformatics 22: 1021–1023. [DOI] [PubMed] [Google Scholar]
- 22. Ahn YY, Bagrow JP, Lehmann S (2010) Link communities reveal multiscale complexity in networks. Nature 466: 761–764. [DOI] [PubMed] [Google Scholar]
- 23. Pinkert S, Schultz J, Reichardt J (2010) Protein interaction networks–more than mere modules. Plos Computational Biology 6: e1000659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Jiao QJ, Zhang YK, Li LN, Shen HB (2011) Bintree seeking: a novel approach to mine both bi-sparse and cohesive modules in protein interaction networks. Plos One 6: e27646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Zhang XF, Dai DQ, Ou-Yang L, Wu MY (2012) Exploring Overlapping Functional Units with Various Structure in Protein Interaction Networks. Plos One 7: e43092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Šubelj L, Bajec M (2011) Generalized network community detection. In Proceedings of the ECML PKDD Workshop on Finding Patterns of Human Behaviors in Network and Mobility Data: 66–84.
- 27.Davis A, Gardner BB, Gardner MR (1941) Deep south: University of Chicago Press Chicago.
- 28.Scott J, Hughes M, Mackenzie J (1980) The anatomy of Scottish capital: Scottish companies and Scottish capital, 1900–1979: Croom Helm London.
- 29. Šubelj L, Bajec M (2011) Community structure of complex software systems: Analysis and applications. Physica A: Statistical Mechanics and its Applications 390: 2968–2975. [Google Scholar]
- 30. Newman M, Leicht E (2007) Mixture models and exploratory analysis in networks. Proceedings of the National Academy of Sciences 104: 9564–9569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Rosvall M, Bergstrom CT (2007) An information-theoretic framework for resolving community structure in complex networks. Proceedings of the National Academy of Sciences 104: 7327–7331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Danon L, Díaz-Guilera A, Duch J, Arenas A (2005) Comparing community structure identification. Journal of Statistical Mechanics: Theory and Experiment 2005: P09008. [Google Scholar]
- 33. Newman MEJ (2003) The structure and function of complex networks. Siam Review 45: 167–256. [Google Scholar]
- 34.Burt RS (2005) Brokerage and closure: An introduction to social capital. New York: Oxford University Press.
- 35.Small ML (2009) Unanticipated gains: Origins of network inequality in everyday life. New York: Oxford University Press.
- 36. Stovel K, Golub B, Milgrom EMM (2011) Stabilizing brokerage. Proceedings of the National Academy of Sciences 108: 21326–21332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Stovel K, Shaw L (2012) Brokerage. Annual Review of Sociology 38: 139–158. [Google Scholar]
- 38. Täube VG (2004) Measuring the social capital of brokerage roles. Connections 26: 29–52. [Google Scholar]
- 39. Šubelj L, Bajec M (2012) Ubiquitousness of link-density and link-pattern communities in real-world networks. The European Physical Journal B-Condensed Matter and Complex Systems 85: 1–11. [Google Scholar]
- 40. Clauset A, Newman MEJ, Moore C (2004) Finding community structure in very large networks. Physical Review E 70: 066111. [DOI] [PubMed] [Google Scholar]
- 41. Lee HK, Hsu AK, Sajdak J, Qin J, Pavlidis P (2004) Coexpression analysis of human genes across many microarray data sets. Genome Research 14: 1085–1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Stuart JM, Segal E, Koller D, Kim SK (2003) A gene-coexpression network for global discovery of conserved genetic modules. Science 302: 249–255. [DOI] [PubMed] [Google Scholar]
- 43. Fukushima A, Nishizawa T, Hayakumo M, Hikosaka S, Saito K, et al. (2012) Exploring Tomato Gene Functions Based on Coexpression Modules Using Graph Clustering and Differential Coexpression Approaches. Plant Physiology 158: 1487–1502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Ciriello G, Cerami E, Sander C, Schultz N (2012) Mutual exclusivity analysis identifies oncogenic network modules. Genome Research 22: 398–406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Spangler JB, Ficklin SP, Luo F, Freeling M, Feltus FA (2012) Conserved Non-Coding Regulatory Signatures in Arabidopsis Co-Expressed Gene Modules. Plos One 7: e45041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Swarbreck D, Wilks C, Lamesch P, Berardini TZ, Garcia-Hernandez M, et al. (2008) The Arabidopsis Information Resource (TAIR): gene structure and function annotation. Nucleic Acids Research 36: D1009–D1014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Obayashi T, Nishida K, Kasahara K, Kinoshita K (2011) ATTED-II updates: condition-specific gene coexpression to extend coexpression analyses and applications to a broad range of flowering plants. Plant and Cell Physiology 52: 213–219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Wei H, Persson S, Mehta T, Srinivasasainagendra V, Chen L, et al. (2006) Transcriptional coordination of the metabolic network in Arabidopsis. Plant Physiology 142: 762–774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Srinivasasainagendra V, Page GP, Mehta T, Coulibaly I, Loraine AE (2008) CressExpress: a tool for large-scale mining of expression data from Arabidopsis. Plant Physiology 147: 1004–1016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Maere S, Heymans K, Kuiper M (2005) BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 21: 3448–3449. [DOI] [PubMed] [Google Scholar]
- 51. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, et al. (2000) Gene Ontology: tool for the unification of biology. Nature Genetics 25: 25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Leskovec J, Lang KJ, Dasgupta A, Mahoney MW (2008) Statistical properties of community structure in large social and information networks. In Proceedings of the 17th International Conference on World Wide Web (WWW): 695–704.
- 53. Airoldi EM, Blei DM, Fienberg SE, Xing EP (2008) Mixed membership stochastic blockmodels. The Journal of Machine Learning Research 9: 1981–2014. [PMC free article] [PubMed] [Google Scholar]
- 54.De Nooy W, Mrvar A, Batagelj V (2011) Exploratory social network analysis with Pajek. New York: Cambridge University Press.
- 55.Pajek datasets. Available: http://vlado.fmf.uni-lj.si/pub/networks/pajek/data/gphs.htm. Accessed 2013 May 8.
- 56. Newman MEJ (2001) The structure of scientific collaboration networks. Proceedings of the National Academy of Sciences 98: 404–409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Leskovec J, Kleinberg J, Faloutsos C (2007) Graph evolution: Densification and shrinking diameters. ACM Transactions on Knowledge Discovery from Data (TKDD) 1: 2. [Google Scholar]
- 58.Knuth DE (1993) The Stanford GraphBase: a platform for combinatorial computing: AcM Press.
- 59. Guimera R, Danon L, Diaz-Guilera A, Giralt F, Arenas A (2003) Self-similar community structure in a network of human interactions. Physical Review E 68: 065103. [DOI] [PubMed] [Google Scholar]
- 60.Reitz JM (2010) Online dictionary for library and information science. Westport, CT: Libraries Unlimited.
- 61.Garfield E (2001) From computational linguistics to algorithmic historiography. paper presented at the Symposium in Honor of Casimir Borkowski at the University of Pittsburgh School of Information Sciences.
- 62. Watts DJ, Strogatz SH (1998) Collective dynamics of small world networks. Nature 393: 440–442. [DOI] [PubMed] [Google Scholar]
- 63. Bu D, Zhao Y, Cai L, Xue H, Zhu X, et al. (2003) Topological structure analysis of the protein–protein interaction network in budding yeast. Nucleic Acids Research 31: 2443–2450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Breitkreutz A, Choi H, Sharom JR, Boucher L, Neduva V, et al. (2010) A global protein kinase and phosphatase interaction network in yeast. Science Signalling 328: 1043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Salwinski L, Miller CS, Smith AJ, Pettit FK, Bowie JU, et al. (2004) The database of interacting proteins: 2004 update. Nucleic Acids Research 32: D449–D451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Brown KR, Jurisica I (2005) Online predicted human interaction database. Bioinformatics 21: 2076–2082. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Brief description of BTS algorithm.
(DOC)