

Abstract
Four experiments were conducted with rats in order to determine whether being placed on a platform in one corner of a rectangular swimming pool results in latent spatial learning. Rats in Experiments 1–3 received four trials a day of being placed on the platform. During a subsequent test trial, in which they were released into the pool without the platform, the rats exhibited a preference for swimming in the correct corners of the pool (those with the same geometric properties as the corner containing the platform during training), than the two remaining, incorrect corners. This effect was seen when the interval between the final placement trial and the test trial was as much as 24 hr (Experiment 2) and after varying numbers of sessions of placement training (Experiment 3). Experiment 4 revealed that when the test took place in a kite-shaped arena, after placement training in a rectangle, a stronger preference was shown for the corner that was geometrically equivalent to the correct rather than the incorrect corners in the rectangle. The placement treatment is said to result in latent spatial learning based on the development of S-S associations.
Keywords: latent learning, geometry, passive placement
Numerous experiments have shown that animals are able to identify the location of a submerged platform in a swimming pool by reference to cues that are some distance from this goal (e.g., Morris, 1981). According to an associative analysis (e.g., White, 2008), such spatial learning may result from the formation of either stimulus-response (S-R) or stimulus-stimulus (S-S) associations. The former would be effective by allowing the animal to make specific responses to specific stimuli on its journey to the goal. The latter would enable animals to learn that the goal is situated in a particular location. An early debate in the study of spatial learning concerned the extent to which each of these associations is responsible for successful spatial behavior. Hull (1943), for example, argued that spatial learning is based entirely on S-R associations, whereas Tolman (1948) argued that S-S associations play an important role in allowing animals to reach a desired goal. More recently, it has been assumed that both kinds of association are involved in spatial learning, and an important focus of research has been to identify the different neural systems that are responsible for the formation of S-S and S-R associations (e.g., Packard & McGaugh, 1996; White, 2008). It should be acknowledged, however, that at least as far as the swimming pool is concerned there is relatively little evidence that shows convincingly that spatial behavior is governed by one kind of association rather than the other.
In the original version of this task, rats were required to escape from a circular pool of water by swimming to a submerged platform, the position of which could be identified by reference to landmarks outside the pool. Morris (1981) trained rats in one experiment to find the platform by repeatedly releasing them from the same position. The rats were subsequently able to find the platform rapidly, even if they were released from a novel location. One explanation for this outcome is that the initial training allowed animals to identify the position of the platform relative to cues outside the pool. Morris suggested the position of the platform was identified with reference to a map-like representation of the relative positions of these cues (e.g., O'Keefe & Nadel, 1978). As an alternative, its position may have been identified with reference to selected landmarks through S-S associations. We shall return to this distinction in Experiment 4. Another possibility is that the training trials resulted in the development of a number of S-R associations. The rat on being released into the pool from a novel location would then swim until it came across a familiar stimulus, which would elicit a response that would lead it either to the goal or to another familiar stimulus which, in turn, would elicit a response that would help it on its way to the goal. Although Morris viewed this interpretation skeptically, it nonetheless remains possible that the original training resulted in S-R associations that guided rats to the goal on the test trial. In order to demonstrate that animals use S-S associations to find a hidden goal, it is necessary for them to be able to develop in a context where it is not possible for S-R associations to develop at the same time. If such training can then be shown to facilitate future attempts to find the goal, then it would suggest that S-S associations do indeed play a role in spatial learning in a swimming pool.
One method for achieving the foregoing objective would be to place the animal on the platform from where it can view the cues surrounding the goal, before releasing it into the pool for a test trial. The placement treatment would provide the opportunity for the formation of an S-S association between the view provided by the surrounding cues and the goal. At the same time, it is hard to see how such treatment would permit the growth of any useful S-R associations, as the animal remains in the same place throughout this training. Accordingly, if the exposure treatment should result in latent learning, and facilitate finding the platform on the test trial, then this outcome is likely to be a consequence of the formation of S-S rather than S-R associations. A number of experiments have adopted this methodology and their results are conflicting.
An example of successful latent learning is reported by Sutherland and Linggard (1982) who placed one group of rats on a platform in a Morris pool for 44 placement trials with each trial lasting 10 s. Ten min after the final placement trial, rats were released into the pool and required to find the platform that was in the same place as for the placement trials. The time taken to reach the platform by this experimental group was significantly less than for a control group for which the platform was in a different location to that used for the placement trials. A similar result was observed by Keith and McVety (1988) except that rats received a single placement trial of 2 min before being released into the pool (see Chew, Sutherland & Whishaw, 1989, for a critique of this experiment). Finally, Jacobs, Zaborowski, and Whishaw (1989a) placed rats on a platform for 15 placement trials, each lasting 30 s, before they were released from the side of the pool. In keeping with the previous studies, the mean escape latency for a group in which the platform during the test trial was in the same position as during the exposure trials was significantly shorter than for a group for which the platform was situated in a new location for the test trials. On the basis of these results Jacobs et al. (1989a) concluded that placement on a platform in a Morris pool can result in latent spatial learning. Jacobs, Zaborowski, and Whishaw (1989b), however, subsequently retracted this conclusion in a brief report that summarizes seven replications of the original study, all of which failed to reveal a significant effect of the placement treatment. At best, therefore, it appears that spatial learning as a consequence of being placed on a platform in a Morris pool is unreliable, and it would therefore be imprudent to draw any theoretical conclusions from the results just considered. In passing we can note that unpublished experiments in our laboratory, similar to those just described, have also revealed conflicting outcomes. On some occasions the placement treatment has resulted in evidence of spatial learning, and on other occasions it has not.
A potential problem with the foregoing studies is that animals may have paid relatively little attention to the landmarks during the placement treatment. The landmarks were a considerable distance from the platform and may thus have been of low salience. If the landmarks were paid little attention, then it is hardly surprising that latent spatial learning was elusive. In an attempt to investigate further whether being placed on a platform in a swimming pool can result in latent spatial learning, the present experiments were based on the design just described but took place in a rather different environment. Instead of using a circular pool with landmarks provided by room cues, the experiments were conducted in a rectangular pool surrounded by curtains so that the position of the platform could be defined only by geometric cues provided by the shape of the pool. It was hoped that positioning the platform near one corner of the pool would result in the walls creating the corner being of considerable salience, and thus enhance the likelihood of the placement treatment being successful. There is ample evidence that animals find a hidden goal by reference to the shape of a rectangular arena when they are consistently released some distance from the goal (e.g., Cheng, 1986; Pearce, Good, Jones, & McGregor, 2004). The initial purpose of the present experiments was to determine if a similar effect can be found when the subject is placed at the goal for all of its training.
Experiment 1 provided a successful demonstration of latent spatial learning, and one objective of the remaining experiments was to confirm the reliability and generality of this effect. Experiment 2 examined whether it was affected by the delay between the final placement trial and the test trial. Experiment 3 examined the effect of manipulating the number of placement trials. A further purpose of Experiment 3 was to investigate the way in which the information acquired during the placement treatment is used to find the platform. Finally, Experiment 4 was conducted in order to gain an insight into the information that is acquired during the placement trial. Thus, the initial purpose of the experiments was to confirm that stimulus-stimulus associations are influential in spatial learning. A further purpose was to gain an understanding of the conditions that promote their formation, the information they contain, and how they influence behavior.
Experiment 1
A single group of rats was placed on a platform situated in one corner of a rectangular pool for a number of trials before a test trial in which they were released into the pool for 60 sec without the platform. During the test trial the amount of time spent in the two correct corners of the rectangle—those with the same geometric properties as the corner used for the placement training—and in the remaining two, incorrect, corners was recorded. If the placement treatment is successful in promoting spatial learning, then during the test the group should spend more time in the correct than the incorrect corners. In this experiment, and all subsequent ones, it was important to ensure that rats did not leave the platform during their placement training, and thus gain experience of swimming in the rectangular pool. If they gained such experience, then it might encourage the development of S-R associations that may then be responsible for a successful outcome to the test trial. Thus, in order to ensure rats remained on the platform for the placement trials, the experiment commenced with three sessions of training in a circular pool. Rats were placed on the platform in this pool, and if they fell off, they were immediately guided back to the platform. We have found that once a rat has become adept at standing on the platform, it is extremely reluctant to leave it.
Method
Subjects
Ten, experimentally naïve, male, hooded Lister Rats (Rattus norvegicus), obtained from Harlan Olac (Bicester, Oxon, England), and weighing between 250g and 300g at the start of the experiment were used. Rats were housed in white plastic cages with secured metal grid lids and maintained on a 12-hr/12-hr light/dark cycle with lights on at 0700. Subjects were housed in pairs and had continuous access to food and water in their home cages.
Apparatus
A white, circular pool measuring 2 m in diameter and 0.6 m deep was used. The pool was mounted on a platform 0.6 m from the floor in the middle of the room (4 m × 4 m × 2.3 m). The pool was filled with water to a depth of 27 cm and was maintained at a temperature of 25 °C (± 2 °C). To make the water opaque, 0.5 L of white opacifer E308 (Roehm and Haas, U.K., Ltd., Dewsbury) was used. The water was changed daily.
A white circular ceiling, measuring 2 m in diameter, was suspended 1.75 m above the floor of the pool. In the center of the ceiling was a hole measuring 30 cm in diameter in which a video camera with a wide-angled lens was situated. The lens of the camera was 25 cm above the hole and was connected to a video monitor and computer equipment in an adjacent room. During tests, the rats' movements were analyzed using Watermaze software (Morris & Spooner, 1990). The pool was illuminated by eight, 45-W lights that were located in the circular ceiling above the pool. The lights were 22.5 cm in diameter and were equidistant from each other in a 1.6 m diameter circle whose center was coincident with the center of the circular ceiling. A platform measuring 10 cm in diameter and mounted on a column was used during all training trials. The surface of the platform had a series of concentric ridges. For all trials, the base of the column rested on the bottom of the pool, and the platform surface was 2 cm below the surface of the water. A white curtain was drawn around the pool during all training and test trials. The curtain, which was attached to the edge of the circular ceiling, was 1.5 m high and fell 25 cm below the edge of the pool.
The training room was additionally lit by two 1.53-m strip lights connected end to end on each of the East and West walls. These lights ran parallel with the floor and were situated 75 cm above the floor. There was a door (1.75 m × 2 m) in the center of the South wall. Throughout the experiment rats were trained in a rectangular-shaped pool constructed from two gray, long Perspex boards (1.8 m long, 0.59 m high, and 2 mm thick) and two gray, short Perspex boards (0.9 m long, 0.59 m high, and 2 mm thick). Each board was placed vertically in the pool and suspended by bars that extended over the edge of the pool.
Procedure
Rats completed one session of four training trials each day. For each session they were carried into a room adjacent to the test room in groups of five in a light-tight box. They remained in this box between trials. For each trial, the rat was carried from the box to the pool and placed on the platform. The rat was allowed to stay on the platform for 30 s, undisturbed, before being removed, dried, and returned to the holding box. The intertrial interval was approximately 5 min.
All rats initially underwent three sessions of pretraining in the circular pool. For these sessions the platform was placed in a quadrant of the pool (NE, NW, SW, or SE), with each quadrant used once in a given session. The platform was randomly positioned either 25 cm or 50 cm from the edge of the pool, each for two trials per session. Following pretraining the rats received 12 sessions of training in the rectangle. For half of the rats, the platform was placed in a corner where the short wall was to the right of the long wall, and for the remainder it was in a corner where the short wall was to the left of the long wall.
In the rectangle, the platform was positioned 25 cm from a corner on an imaginary line that bisected the corner. Between each trial, the rectangle was randomly rotated 90°, 180°, or 270° clockwise. Four possible orientations were used (North, South, East, or West). The orientation of the arena across trials varied randomly with the only stipulation being that each orientation was used once for any given session.
The first three trials of Session 12 were conducted in the same manner as for the previous sessions. The fourth trial, which took place approximately 5 min after the final placement trial, consisted of a test trial in the rectangle, whose orientation was different to the previous three training trials, in the absence of the platform. Rats were released from the center of the pool and allowed to swim for 60 s.
For the purpose of analyzing the results from the test trial, circular search zones in each corner were used. Each zone had a diameter of 30 cm with its center positioned 25 cm from a corner on a line that bisected the corner. The percentage of time spent in the zones in the correct corners (the two geometrically equivalent corners where the platform had been located during training) and in the incorrect corners (the remaining two corners) of the rectangular pool were analyzed. A Type-1 error rate of .05 was adopted for all reported statistical comparisons.
Results and Discussion
The percentage of time spent in the correct and incorrect zones in the rectangle for the test trial was 19.61% (SEM = 2.59%) and 8.41% (SEM = 1.63%) respectively. The animals expressed a clear preference for searching in the correct rather than the incorrect corners of the pool. In support of this observation, a paired t test, based on individual times spent in the correct and incorrect search zones, revealed that significantly more time was spent in the former than the latter, t(9) = 3.39.
The results demonstrate that being placed on a platform in one corner of a rectangular pool is sufficient to encourage rats when released into the pool to spend more time in this corner, and the diagonally opposite corner, than in the remaining, incorrect corners. One interpretation of this finding is that the placement treatment resulted in the formation of an S-S association between cues provided by the shape of the pool and cues provided by the platform, and this association was then responsible for guiding the rats to the correct corner during the test trial.
We shall consider in Experiment 3 how an S-S association might influence behavior in this way. First, however, we describe an experiment that was intended to test further the possibility that the placement training resulted in the formation of an S-S association. A general property of associations, both S-S and S-R, is that once their strength is close to asymptote their influence is relatively long-lasting. For example, if rats receive appetitive conditioning in which a tone signals food, then the tone will normally elicit a conditioned response at least 24 hours after the final conditioning trial. In the present experiment, the effects of the placement training were examined approximately 5 min after the final placement trial, and it is not clear that a similar outcome of the experiment would have been found if this interval was increased to 24 hours. Indeed, although the results reported by Jacobs et al. (1989a) have been retracted (Jacobs et al., 1989b), it is worthy of note that they concluded the effects of their placement training were quickly forgotten (within 2 hr). If the same is true for the methodology we have adopted, then it might be misleading to attribute the effects of our placement treatment to the formation of an S-S association. With this concern in mind, two groups in Experiment 2 received the same placement training that was administered in Experiment 1. One group then received the test Trial 5 min after the final placement trial, and a second group was tested after an interval of 24 hr.
Experiment 2
Method
Subjects and apparatus
Twenty-three animals were maintained in the same manner, and were from the same stock as in Experiment 1. Eleven of the animals had previously been used in a standard object recognition experiment but had no experience in a swimming pool. The remaining rats were experimentally naïve. The apparatus was identical to that used in Experiment 1.
Procedure
The procedure was identical to Experiment 1 with the following exceptions. Rats were randomly assigned to two groups, a short delay group (n = 12) and a long delay group (n = 11). For the short delay group, the test trial on Session 12 was conducted approximately 5 min after the third trial. For the long delay group, following the three training trials in the rectangle on Session 12, there was a delay of 24 hours before the rats were tested. In both instances, the test trial was conducted and analyzed in the same manner as for Experiment 1.
Results and Discussion
Figure 1 shows the percentage of time spent searching in the correct and incorrect zones for the test trial for the short (left-hand panel) and long (right-hand panel) delay groups. Irrespective of the delay, both groups spent a higher proportion of time in the correct than the incorrect zone. A 2 × 2 (Group × Zone) ANOVA was conducted and revealed a significant main effect of zone, F(1, 21) = 7.04. The main effect of group and the Group × Zone interaction was not significant, Fs (1, 21) < 1.06. It is clear that with 12 sessions of placement training, rats develop a relatively enduring representation of where the goal is located. One way of characterizing this representation is in the form of an S-S association.
Figure 1. The mean (+SEM) percentage of time spent in the correct and incorrect zones during the test trial for the short and long delay groups of Experiment 2.

Experiment 3
Experiments 1 and 2 have shown that four placement trials a day for 12 days is sufficient to result in rats subsequently expressing a significant preference for the correct over the incorrect corners of the rectangular pool. One purpose of the next experiment was to assess whether a similar preference will be expressed after fewer training trials. Thus, three groups were trained identically to the group in Experiment 1, except that 2, 5, and 9 sessions of training were given before the test trial. If the results from the first experiment can be replicated with at least some of these groups, then it would serve to confirm the reliability and generality of the results revealed by Experiment 1.
A further purpose of the experiment was to evaluate two different explanations for the way in which the S-S associations formed during the placement trials influence the outcome of the test trial. According to one explanation, on being placed in the pool the rats might swim at random until, by chance, they enter a corner. If they should enter a correct corner, then they might recognize it as the one where they received their placement training and spend time in it searching for the platform. On the other hand, if they should enter an incorrect corner, then it will not be familiar, and they may waste no time there searching for the platform. According to an alternative explanation, after being released into the pool, the rats might inspect the boundary and identify one of the corners as being equivalent to the one where the placement training was conducted. They might then head toward this corner and search for the platform when they reach it. Thus, both accounts predict that rats will spend more time in the correct than the incorrect corners during the test trial, but they differ in the manner in which the rat is assumed to select a correct corner. One account asserts the corner will be chosen by chance, and the other asserts it will be chosen deliberately. In order to evaluate these accounts, a record was taken of which corner a rat entered first after being released. The first account predicts that rats will fail to show a preference of heading for a correct than an incorrect corner when they are released, whereas the second account predicts that rats will head more often for a correct than an incorrect corner.
Method
Subjects and apparatus
Forty-eight experimentally naïve animals were maintained, housed, and were from the same stock as in Experiment 1. The apparatus was identical to that used in Experiment 1.
Procedure
The procedure was identical to Experiment 1 with the following exceptions. Rats were randomly assigned, in equal numbers (n = 16) to three groups: Group 2, Group 5, and Group 9. The number indicates the session in which testing occurred. Also, the start of training was staggered for each group to ensure testing occurred on the same day for all animals. The test was conducted immediately following the third trial of the final session for each group. Apart from recording the percentage of time spent in the search zones during the test trial, a record was taken of whether, after being released, a rat entered first one of the correct corners of the pool. A rat was deemed to have entered any of the four corners if its snout crossed a notional circular line with a radius of 40 cm and with its center at the point where the walls creating the corner met.
Results and Discussion
Figure 2 shows the percentage of time spent in the correct and incorrect zones for the three groups. All groups showed a preference for the correct over the incorrect zone, although this effect was more pronounced in Group 9 than in the other two groups. A 3 × 2 (Group × Zone) ANOVA revealed a significant main effect of zone, F(1, 45) = 21.29. The main effect of group and the Group × Zone interaction was not significant, Fs (2, 45) <1.22. Paired t tests using the results from each of the three groups separately revealed that significantly more time was spent in the correct than the incorrect corner by Group 2, t(15) = 2.82, Group 5, t(15) = 2.15, and Group 9, t(15) = 3.19.
Figure 2. The mean (+SEM) percentage of time spent in the correct and incorrect zones during the test trial for the three groups of Experiment 3.
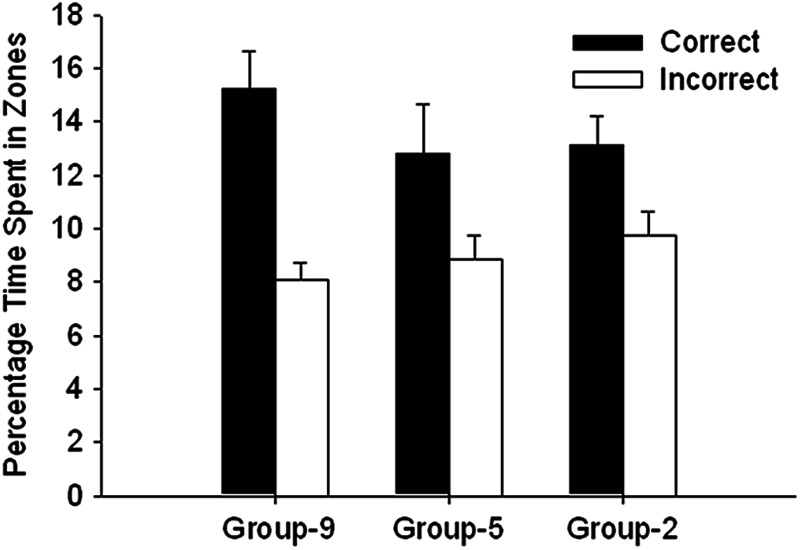
Upon being released into the pool during the test trial, 12 out of 16 rats in Group 2 and in Group 9, and 11 out of 16 rats in Group 5 entered a correct corner before entering an incorrect corner. A binomial test on the choices combined across the three groups revealed that the number of rats that chose the correct corner first (35) was significantly above chance, z = 3.18, p = .002.
The significantly greater proportion of time that was spent in the correct than the incorrect search zones by each of the three groups during the test trial confirms that the effect of the placement training observed in Experiments 1 and 2 is reliable and robust. Moreover, it appears that as few as seven placement trials are sufficient in order for a preference for the correct corner to develop.
The experiment also revealed that after being released from the center of the pool, rats were significantly more likely to enter first a correct rather than an incorrect corner. A rat was deemed to have made this choice when its snout entered a notional circle with its center coincident with the join between the walls creating the corner, and with a radius of 40 cm. Thus, the decision by a rat to enter a correct corner was made while its current view of the corner must have been quite different to that experienced as it received its placement training. The obvious question is then raised as to how is an animal able to identify the correct corner when it experiences a novel view of that corner. We shall return to this issue in the General Discussion.
Experiment 4
The results from the previous experiments strongly suggest that an association between cues provided by the shape of the environment, and those provided by the submerged platform, is formed as a consequence of the placement training. The purpose of the final experiment is to understand a little about the nature of information concerning the shape of the environment that is contained within this association. We have already noted Morris (1981) argued that animals trained in a swimming pool form a map-like representation of their environment, and then identify the location of the goal with reference to this representation (O'Keefe & Nadel, 1978). A related view has been expressed by Cheng (1986) in order to explain how rats are able to use the shape of a rectangular arena in order to find food hidden in one corner. He argued that the rats constructed a representation of the overall shape of the environment, and that the position of the goal was pasted onto this representation (see also Gallistel, 1990; Cheng & Gallistel, 2005). One way of characterizing these accounts is to say that rats construct a global representation of their environment in which the geometric relations among the various objects play a prominent role.
Rather than form a global representation during their placement training, rats may rely on less complex information to identify where the platform is located. For example, being placed on a platform in one corner of a rectangular arena might result in a representation of just this corner entering into an association with the goal. Alternatively, a particular end of a wall of a certain length could serve as a simple cue for indicating where the platform is located (Pearce et al., 2004; Pearce, 2009).
As a step toward choosing among these possibilities, Experiment 4 was based on a study reported by Pearce et al. (2004). During the first stage of the experiment, rats received placement training in a rectangular pool in the same manner as for the previous experiments. They then received a single test trial in a kite-shape pool. The kite was constructed from the same walls as the rectangle and contained two right-angled corners. One of these right-angled corners will be referred to as the correct corner as it is equivalent geometrically to the corners in the rectangle where the platform was located. The other right-angled corner in the kite is referred to as the incorrect corner because of its equivalence to the incorrect corners in the rectangle. If the placement training in the rectangle should result in animals forming a global geometric representation of this environment, then on being placed in the kite they should effectively be lost as the representation of the rectangle will fail to match the shape of the kite. According to this account, therefore, subjects should fail to show a preference for the correct over the incorrect corner during the test in the kite. In contrast, if the initial training should result in the position of the platform being identified with reference to a particular corner, then there is no reason why rats should not show a preference for the equivalent right-angled corner in the kite. The outcome of the test trial should therefore indicate whether placement training in the rectangle results in a reliance on local or global cues to indicate where the platform is located.
The previous experiment revealed that as few as seven placement trials was sufficient for this treatment to be effective. We were not sure how many trials to give in the present experiment, however, because the difference between the training and test environments might weaken responding during the test trials through generalization decrement, and result in poor performance if insufficient training had been given. Accordingly, the experiment contained three groups that received 4, 8, and 16 sessions of placement training, with the final trial of the last session being a test in the kite.
Method
Subjects and apparatus
Forty-eight experimentally naïve animals were maintained and from the same stock as in Experiment 1. The four boards that were used to form the rectangle were also used to create the kite for the test trial. All other details were the same as for Experiment 1.
Procedure
The animals were randomly assigned to three groups in equal number (n = 16): Group 4, Group 8, and Group 16 (the number indicates the number of training sessions the group received in the rectangle). The groups were trained in the same manner as in the previous experiments. Three animals were removed from the behavioral analysis because they left the platform and gained experience of swimming in the rectangle during the placement trials. Thus, final group sizes were 16, 15, and 14 for Group 4, Group 8, and Group16, respectively.
The first three trials of the final session of training for each group were conducted in the rectangle in the same manner as previous trials. The fourth trial of these sessions, which commenced 5 min after the final placement trial, was a test trial in a kite. Rats were gently released into the center of the pool and allowed to swim for 60 s.
To analyze the test trial in the kite, 30-cm diameter search zones were identified in each of the four corners of the kite. The percentage of time spent in the zone in the correct corner (the corner in the kite that was geometrically identical to the corner housing the platform in the rectangle), the incorrect corner (the opposite corner), the apex corner, and the obtuse corner was recorded. In addition to the analysis of time spent in the corners, upon being released into the pool for the test trial, a record of which corner the animal visited first was taken.
Results and Discussion
The left-hand panel of Figure 3 shows the percentage of time spent in the correct and incorrect zones for the three groups during the test in the kite. All groups showed a marked preference for the correct than the incorrect zone. A 3 × 2 (Group × Zone) ANOVA was conducted and revealed a significant effect of zone, F(1, 42) = 14.37. The main effect of group and the Group × Zone interaction were not significant, Fs (2, 42) < 2.57. Paired t tests for the results of each of the three groups separately, revealed that significantly more time was spent in the correct than the incorrect search zone by Group 8 t(14) = 3.22, and Group 16, t(13) = 2.42, but not Group 4, t(15) = 1.37.
Figure 3. The mean (+SEM) percentage of time spent during the test trial in the kite in Experiment 4 in the correct and incorrect search zones by each of the three groups (left-hand panel), in each of the four corners by the three groups combined (center panel) and the number rats for the three groups combined that entered first each of the four corners (right-hand panel).

For the sake of convenience, the results from the three groups have been combined for the purposes of presenting and analyzing the percentage of time spent in each of the four corners and the choice data. The center panel of Figure 3 therefore shows the mean time spent by the three groups in each of the four corners of the pool. The striking feature of this figure is that more time was spent in the correct corner than any of the other corners. A one-way ANOVA revealed that the effect of zone was significant, F(3, 132) = 10.45. Subsequent comparisons then revealed that significantly more time was spent in the correct corner than in any of the other three corners individually, ts (44) > 2.97, while the remaining comparisons between pairs of corners were not significant, ts (44) < 1.40.
The right-hand panel of Figure 3 shows for the three groups combined, the number of rats that entered first each of the four corners of the kite. There was a strong tendency to head directly toward the correct corner rather than any other corner. In support of this observation, a χ2 test revealed a significant difference among the number of rats entering the different corners, χ2(3) = 10.95. In addition, 30 of the 45 rats entered the correct corner before the incorrect corner. This preference for the correct corner was significant with the binomial test, z = 2.09, p = .04. The number of rats in each group that entered the correct corner before the incorrect corner was as follows: Group 4, 9 out of 16; Group 8, 11 out of 15; Group 16, 10 out of 14.
In keeping with the results from the three previous experiments, the placement training in the rectangle was again found to be successful, both in terms of the time spent in the correct corner of the kite and the number of rats that headed directly to this corner. This pattern of results would not be expected if the placement training resulted in the position of the platform being based on a global representation of the overall shape of the rectangular pool. If this had been the case, then it is hard to see how the difference between the shapes of the arenas would permit the preference for the correct over the incorrect corner to transfer from the rectangle to the kite. Instead, it appears that the position of the platform was identified by simpler means.
Pearce et al. (2004) conducted a similar experiment to the present one, except that rats were required to swim to the platform on every trial in the rectangle. Testing in the kite revealed that subjects preferred to swim directly toward the correct rather than the incorrect right-angled corner, which again suggests their ability to find the platform in the training stage did not depend upon the use of a global representation of the geometric attributes of the rectangle. A plausible alternative explanation for the outcome of the experiment is that rats during the training stage identified the position of the platform by referring to a fragment of the rectangle. Although an obvious candidate for this fragment is the corner in which the platform was situated, further results by Pearce et al. (2004) point to a different conclusion. During the test stage, subjects showed a stronger tendency to head toward both the correct corner and the apex of the kite than the incorrect corner. Such a preference for the apex is hard to explain if animals were searching for a right-angled corner with particular properties. Instead, as a result of their initial training, they may have learned to search for the platform at a particular end of a long wall that, depending on the wall that was selected, would then lead them to either the correct corner or the apex in the kite.
It does not appear that the rats in the present study adopted a similar strategy. On being released into the pool substantially fewer rats headed directly toward the apex of the pool than the correct corner, and substantially more time was spent in the correct corner than the apex. Both of these findings suggest that the initial training resulted in animals favoring the correct corner over the apex, which would not be expected if they were searching for the platform at a particular end of a long wall. Instead, the results suggest that during the test trial rats were searching for a corner with the same geometric properties as the correct corner in the rectangle.
General Discussion
The four experiments demonstrate for the first time that being placed on a platform in one corner of a rectangular swimming pool results in a subsequent preference for this, and the diagonally opposite corner. Such a preference can be observed after as few as 7 placement trials and can be observed even when testing is delayed for 24 hours after the final placement trial. Taken together, the results are consistent with the claim that the placement training results in the growth of an S-S association.
The above experiments are not the first to demonstrate that S-S associations influence spatial behavior in an environment with a distinctive shape. Horne and Pearce (2009; see also Rhodes, Creighton, Killcross, Good, & Honey, 2009) have provided a rather different demonstration of the influence of these associations to that revealed here. Rats were trained to find a platform in one corner of a kite. The walls creating the corner with the platform were black, while those creating the opposite corner were white. The significance of the black and white walls was then reversed by requiring the rats to find a platform in the white corner, but not the black corner, of a square with two adjacent white walls and two adjacent black walls. A subsequent test trial in an all white kite then revealed a preference for the previously incorrect over the previously correct corner. In essence, Horne and Pearce (2009) explained this result by suggesting that S-S associations formed during the first stage between the black walls and the correct corner. During the test trial, the sight of the correct corner would retrieve a memory of the black walls, the value of which would have been reversed in the second stage of the experiment. As a result, the rats would then be encouraged to swim away from the correct corner and express a preference for the incorrect corner.
In the foregoing experiment, it is possible that, apart from the between-cue associations that have just been identified, the remaining associations were all S-R in nature. Indeed, in all experiments where animals have been required to escape from a pool by swimming to a submerged platform, it is possible that behavior was a consequence of S-R associations guiding them in the correct direction. The present experiments might therefore be unique in being the first to establish that S-S associations play a role in helping an animal reach a goal in a Morris pool. The important question is then raised: How does an association between two stimuli result in the animal making a response that leads it to its goal in such an environment? We noted in the introduction to Experiment 3 that S-S associations might be effective by enabling animals to recognize a correct corner once they had reached it by chance. Such an account is unable to explain the results of Experiments 3 and 4, where there was a significant tendency for animals to select a correct rather than an incorrect corner when they were at least 40 cm from it.
One explanation for the emergence of this preference to enter a correct corner over an incorrect corner is that animals possessed a representation of the overall shape of the arena and used this to determine where to head after being released. Although the results of Experiment 3 are consistent with this explanation, for reasons already spelled out, the results from Experiment 4 do not appear to be amenable to this sort of explanation. As an alternative it is possible that rats were able to differentiate between correct and incorrect corners on the basis of template matching (Cartwright & Collett, 1983; Collett & Collett, 2002). The animal is assumed to take a snapshot, as it were, of the view at the goal. When released into the pool, it then compares its current view with the remembered view and moves in such a way as to minimize the difference between the two. Stürzl, Cheung, Cheng, and Zeil (2008) have shown how template matching can result in successful search for a platform in rectangular pool. On reaching the goal the subject is assumed to take a panoramic 360o image, which is then compared with similar panoramic views as it moves through the environment. Cheung, Stürzl, Zeil, and Cheng (2008) go on to demonstrate formally how this account can predict the successful transfer from a rectangle to a kite that was reported by Pearce et al. (2004). The account by Stürzl et al. (2008) assumes that the animal makes its own way to the goal before taking the all important snapshot, but there is no reason why a similar snapshot should not be taken when the animal is placed on the platform. The snapshot could then be used in the manner described by Stürzl et al. (2008) to guide the animal when it is released for the first time into either a rectangular or kite-shaped test arena. A clear implication of the account proposed by Stürzl et al. is that animals should be able to navigate to a goal without the benefit of S-R associations. The present experiments, by making it likely that S-R associations were not involved in the successful outcome of the test trials, thus provide novel support for an important assumption of this type of theory.
A distinctive feature of the proposals of Stürzl et al. (2008) is that animals take a panoramic snapshot of the entire environment when they reach the goal. In the template-matching account of navigation proposed by Cartwright and Collett (1983), the snapshot is concentrated on the cues situated close to the goal. It remains to be seen whether a formal model based on this account is able to explain our results.
Shenikhovich, Chavarriaga, Strösslin, Arleo, and Gerstner (2009) have also proposed that navigation in a rectangular environment is based on snapshots. However, in contrast to Stürzl et al. (2008), these snapshots are assumed to guide behavior by entering into associations with responses that lead to reward. It would seem to follow from this essentially S-R analysis that placement training by itself will not be sufficient to enable a rat to plot a course to a hidden goal when it is required to swim in the pool for the first time. In view of the present results, it may be necessary for the model proposed by Shenikhovich et al. (2009) to be modified.
The discussion thus far has considered the nature of the information contained within the first element of an S-S association. But what about the second element? One possibility is that this element represents the stimulation provided by the platform when the rat is placed on it. Another possibility is that the second element represents the reinforcement provided by standing on the platform when surrounded by water. White (2008) referred to associations involving this second type of information as Stimulus-Reinforcement associations (S-Rft). He has further argued that such S-Rft assocations arise when rats are placed in a particular location in a maze and given food. Although it is quite possible that S-Rft associations also form when rats are placed on a platform in a swimming pool, there is no evidence in the present experiments to indicate this was the case. As a result, it is not possible to comment with any certainty about the nature of the information contained within the second element of the S-S associations that resulted from our placement training.
The four experiments have shown that allowing a rat to view a rectangular pool from a submerged platform in one corner is sufficient for it to discriminate subsequently between the correct and incorrect corners of the pool. The success of this placement treatment contrasts with the mixed outcomes from investigations of latent spatial learning in a Morris pool when the platform can be found by reference to cues outside the pool (e.g., Jacobs et al., 1989a, b). It is hard to offer with any degree of confidence an explanation for why the results from our experiments have been more reliable than those from the other studies. It is possible, however, that the cues that indicated where the platform is situated are easier to discern when the placement training takes place in a rectangular pool, rather than in a circular pool with distal landmarks.
Having established that S-S associations develop during spatial learning in a rectangular environment, it might be argued that all demonstrations of spatial learning in this environment, even when the animal must make its own way to the goal, are due to the influence of S-S associations. The experiments thus raise the challenge of ascertaining whether S-R associations are acquired when an animal must find a hidden goal in an environment with a distinctive shape. As a step toward addressing this challenge we can return to the results of Experiment 4, and the study by Pearce et al. (2004). Pearce et al., it will be recalled, concluded that after being required to swim to a platform in a rectangle, rats developed the strategy of swimming to the appropriate end of one of the long walls. As there was no evidence of this strategy being adopted in the test trial of Experiment 4, it is tempting to speculate that the strategy was based on an S-R habit that resulted from being required to swim to the platform during the training trials. Further research is needed before the merits of this speculation can be determined.
Acknowledgments
The reported research was supported by a grant from the Wellcome Trust.
References
- Cartwright B. A., & Collett T. S. (1983). Landmark learning in bees: Experiments and models. Journal of Comparative Physiology A, 151, 521–543. doi:10.1007/BF00605469 [Google Scholar]
- Cheng K., & Gallistel C. R. (2005). Shape parameters explain data from spatial transformations: Comment on Pearce et al. (2004) and Tommasi & Polli (2004). Journal of Experimental Psychology: Animal Behavior Processes, 31, 254–259. doi:10.1037/0097-7403.31.2.254 [DOI] [PubMed] [Google Scholar]
- Cheng K. (1986). A purely geometric module in the rat's spatial representation. Cognition, 23, 149–178. doi:10.1016/0010-0277(86)90041-7 [DOI] [PubMed] [Google Scholar]
- Cheung A., Stürzl W. S., Zeil J., & Cheng K. (2008). The information content of panoramic images II: View-based navigation in nonrectangular experimental arenas. Journal of Experimental Psychology: Animal Behavior Processes, 34, 15-30. [DOI] [PubMed] [Google Scholar]
- Chew G. L., Sutherland R. J., & Whishaw I. Q. (1989). Latent learning does not produce instantaneous transfer of place navigation: A rejoinder to Keith and McVety. Psychobiology, 17, 207–209. [Google Scholar]
- Collett T. S., & Collett M. (2002). Memory use in insect visual navigation. Nature Reviews Neuroscience, 3, 542–552. doi:10.1038/nrn872 [DOI] [PubMed] [Google Scholar]
- Gallistel C. R. (1990). The Organisation of Learning. Cambridge, MA: MIT Press. [Google Scholar]
- Horne M. R., & Pearce J. M. (2009). Between-cue associations influence searching for a hidden goal in an environment with a distinctive shape. Journal of Experimental Psychology: Animal Behavior Processes, 35, 99–107. doi:10.1037/0097-7403.35.1.99 [DOI] [PubMed] [Google Scholar]
- Hull C. L. (1943). Principles of behavior. New York, NY: Appleton-Century-Crofts. [Google Scholar]
- Jacobs W. J., Zaborowski J. A., & Whishaw I. Q. (1989a). Rats repeatedly placed on a hidden platform learn but quickly forget its location. Journal of Experimental Psychology: Animal Behavior Processes, 15, 36–42. doi:10.1037/0097-7403.15.1.36 [Google Scholar]
- Jacobs W. J., Zaborowski J. A., & Whishaw I. Q. (1989b). Failure to find latent spatial learning in the Morris water task: Retraction of Jacobs, Zaborowski, and Whishaw (1989). Journal of Experimental Psychology: Animal Behavior Processes, 15, 286. doi:10.1037/0097-7403.15.3.286 [Google Scholar]
- Keith J. R., & McVety K. M. (1988). Latent place learning in a novel environment and the influence of prior training in rats. Psychobiology, 16, 146–151. [Google Scholar]
- Morris R. G. M. (1981). Spatial localization does not require the presence of local cue. Learning and Motivation, 12, 239–260. doi:10.1016/0023-9690(81)90020-5 [Google Scholar]
- Morris R. G. M., & Spooner R. I. W. (1990). Watermaze software (computer software). Edinburgh, UK: Watermaze Software. [Google Scholar]
- O'Keefe J., & Nadel L. (1978). The hippocampus as a cognitive map. Oxford: Clarendon Press. [Google Scholar]
- Packard M. G., & McGaugh J. L. (1996). Inactivation of hippocampus or caudate nucleus with lidocaine differentially affects expression of place and response learning. Neurobiology of learning and memory, 65, 65–72. doi:10.1006/nlme.1996.0007 [DOI] [PubMed] [Google Scholar]
- Pearce J. M., Good M. A., Jones P. M., & McGregor A. (2004). Transfer of spatial behavior between different environments: Implications for theories of spatial learning and for the role of the hippocampus in spatial learning. Journal of Experimental Psychology: Animal Behavior Processes, 30, 135–147. doi:10.1037/0097-7403.30.2.135 [DOI] [PubMed] [Google Scholar]
- Pearce J. M. (2009). An associative analysis of spatial learning. Quarterly Journal of Experimental Psychology, 62, 1665–1684. doi:10.1080/17470210902805589 [DOI] [PubMed] [Google Scholar]
- Rhodes S. E., Creighton G., Killcross A. S., Good. M., & Honey R. C. (2009). Integration of geometric with luminance information in the rat: Evidence from within-compound associations. Journal of Experimental Psychology: Animal Behavior Processes, 35, 92–98. doi:10.1037/0097-7403.35.1.92 [DOI] [PubMed] [Google Scholar]
- Sheynikhovich D., R C., Strosslin T., Arleo A., & Gerstner W. (2009). Is there a geometric module of spatial orientation? Insights from a rodent model of navigation. Psychological Review, 116, 540–546. doi:10.1037/a0016170 [DOI] [PubMed] [Google Scholar]
- Stürzl W. S., Cheung A., Cheng K., & Zeil J. (2008). The information content of panoramic images I: The rotational errors and the similarity of views in rectangular experimental arenas. Journal of Experimental Psychology: Animal Behavior Processes, 34, 1–14. doi:10.1037/0097-7403.34.1.1 [DOI] [PubMed] [Google Scholar]
- Sutherland R. J., & Linggard R. C. (1982). Being there: A novel demonstration of latent spatial learning in the rat. Behavioral and Neural Biology, 36, 103–107. doi:10.1016/S0163-1047(82)90101-7 [DOI] [PubMed] [Google Scholar]
- Tolman E. C. (1948). Cognitive maps in rats and men. Psychological Review, 55, 189–208. doi:10.1037/h0061626 [DOI] [PubMed] [Google Scholar]
- White N. M. (2008). Multiple memory systems in the brain: Cooperation and competition. In Eichenbaum H. B. (Ed.), Memory systems (pp. 9–46). Oxford: Elsevier. [Google Scholar]