

Abstract
There are increasing reports of cognitive and psychological declines related to occupational stress in subjects without psychiatric premorbidity or major life trauma. The underlying neurobiology is unknown, and many question the notion that the described disabilities represent a medical condition. Using PET we recently found that persons suffering from chronic occupational stress had limbic reductions in the 5-HT1A receptor binding potential. Here we examine whether chronic work-related stress is also associated with changes in brain structure. We performed MRI-based voxel-based morphometry and structural volumetry in stressed subjects and unstressed controls focusing on gray (GM) and white matter (WM) volumes, and the volumes of hippocampus, caudate, and putamen – structures known to be susceptible to neurotoxic changes. Stressed subjects exhibited significant reductions in the GM volumes of the anterior cingulate cortex and the dorsolateral prefrontal cortex. Furthermore, their caudate and putamen volumes were reduced, and the volumes correlated inversely to the degree of perceived stress. Our results add to previous data on chronic psychosocial stress, and indicate a morphological involvement of the frontostriatal circuits. The present findings of morphological changes in these regions confirm our previous conclusion that symptoms from occupational stress merit careful investigations and targeted treatment.
Introduction
Western societies are facing increasing reports of stress-related sickness among otherwise healthy and high-performing persons who report that they have not experienced any major negative life events or particular stress in early life [1], [2], [3], [4], [5]. These persons describe having stereotyped symptoms, including memory and concentration problems, sleeplessness, diffuse aches, profound fatigue, irritability, anxiety, and a feeling of being emotionally drained, which they often attribute to occupational stress. It has also been found that such stressed individuals can experience an acute phase with symptoms of hypertension, chest pain, dizziness and serious cognitive disabilities [6]. Even though many individuals recover from the acute symptoms, the cognitive and emotional dysfunction as well as the increased sensitivity to stress often last for months, or years, forcing the affected individuals to work part-time, change jobs or retire early. Many professionals are still not accepting the described disabilities as a medical condition, and when accepted, they tend to be misdiagnosed as depression. Only a minor portion of the affected individuals are helped by treatment with serotonin reuptake inhibitors or other antidepressants [7]. The major pathways of the physiological response to stress involve autonomic nervous system as well as hypothalamic–pituitary–adrenal (HPA) axis [8]. Although subjects suffering from symptoms attributed to chronic occupational stress are believed to have an altered reactivity of the HPA system, it is unknown whether this is a consistent finding [9], and both normal [10], [11], reduced [12], [13], [14], [15], and elevated [16], [17], [18] cortisol levels after awakening have been reported. Furthermore, in contrast to patients with major depression [19], subjects with chronic occupational stress symptoms show reduced cortisol and ACTH responses to the Corticotropin Releasing Hormone after dexametasone pretreatment [5], [20]. Emotional reactions to chronic stress and major depression, thus, seem to represent at least partly separate constructs, even if some symptoms (such as anxiety and attention and memory deficits) may overlap.
In a recent PET study, we found that patients suffering from chronic work-related psychosocial stress had significant reductions in the 5-HT1A receptor binding in three limbic structures: the hippocampus, the anterior cingulate cortex (ACC), and the anterior insular cortex [21]. A functional disconnection was also found between the amygdala, the ACC, and the medial prefrontal cortex (mPFC), despite the absence of psychiatric co-morbidity and major negative life events [21]. The locations of changes, in several aspects, corresponded to the locations of structural changes detected through MRI in persons suffering from other stress-related conditions, such as stress in early life, repeated stressful negative life events, and post-traumatic stress disorder (PTSD), [22], [23], [24], [25], [26], [27], [28], [29].
This similarity raises the question of whether certain changes in structural volumes also occur among persons with symptoms related to chronic work-related stress. This is of interest for a couple reasons. Firstly, since the pathophysiology of symptoms attributed to occupational stress is highly debated, there is a need for clarification on whether this condition is associated with any cerebral changes. Secondly, if structural changes similar to those described in other stress-related conditions are also linked to occupational stress, one may hypothesize that chronic psychosocial stress affects our brains in a rather stereotyped manner, regardless of the underlying cause, and that cerebral changes are not limited to exposure to extreme and life threatening situations, but can also be related to accumulated everyday stress.
We therefore carried out comparative MRI studies of cerebral gray matter (GM) and white matter (WM) volumes between patients with chronic occupation-related stress and healthy controls. The study also included an analysis of the structural volumes of the hippocampus, caudate, and putamen. The two latter regions, to the best of our knowledge, have not been previously investigated in this population. They were, however, of interest because they have been shown to process stress stimuli [30], [31], [32], [33] Furthermore, magnetic resonance imaging (MRI) of a large sample of adults with no history of psychopathology revealed that people who have experienced significant early life stress have volumetric reductions in the caudate nucleus in addition to the anterior cingulate cortex [24]. Also, a recent study of effects of life traumas during youth, reveal reductions in the caudate and putamen volumes [34], further emphasizing that volume changes in these structures may be related to stress. On the basis of these previous data, and assuming that a repetitive stress-induced activation could lead to neuronal and dendritic damage in the structures involved, it was hypothesized that the caudate and putamen volumes may be smaller in persons suffering from prolonged occupational stress than in controls. Given previous findings on the effects of stress on the brain [5], [6], [21], [35], a further hypothesis was that our stressed subjects would have reductions in the GM and WM volumes in the ACC and mPFC, and in the grey matter volumes of the hippocampus, amygdala and the insular cortex. These hypotheses were tested in a magnetic resonance (MR) study, which combined two different analyses – voxel based morphometry (VBM) and structural volumetry. VBM was carried out in two ways: 1) exploratively, with the entire brain as search space, to investigate whether there were any regional GM and WM changes in the brain on pixel by pixel basis. 2) It was also used with the assumption that GM and WM would be changed in regions known to be consistently involved in the regulation of stress (such as the hippocampus, mesial prefrontal cortex, insular cortex and the amygdala), and therefore, restricting the search space to a mask covering these areas (see methods). Structural volumetry was carried out in addition to the VBM analyses because some subcortical structures, basal ganglia in particular, have a poor white and gray matter demarcation, and the volumes of these structures are better defined with manual delineation.
Methods
Subjects
Thirty right-handed [36], non-smoking patients (23 women and 7 men, age 41.3±6.6, range 36–55 years, education 13.5±2 years), who had been diagnosed as having had a ‘reaction to severe stress and an adjustment disorder’ according to the International Classification of Diseases (ICD-10, F43), were recruited from the Stress Research Institute at Stockholm University. In order to compose a study group with a homogenous etiology and to reduce variability, the selection was limited to subjects who attributed their illness to prolonged work-related stress, after working 60 to 70 hours per week continuously over several years prior to the onset of symptoms. Inclusion criteria consisted of a characteristic symptom course of sleeplessness, diffuse aches, palpitations and fatigue, a subsequent onset of irritability, anxiety, memory and concentration problems, feeling of depersonalization, and reduced work capacity (confirmed by the employers) [5], [6]. All of the subjects attributed their symptoms to chronic stress and had no other known etiology for their distress.
Subjects were also required to have had a symptom duration of at least one year, to have been on sick leave (≥50%) for stress-related symptoms for a minimum of 6 months before entering the study, and to have an average stress-burnout score of ≥3.0 on the Maslach Stress-Burnout Inventory – General Survey (MBI-GS), [37]. This 7-point rating scale, ranging from 0 (never) to 6 (daily), consists of three subscales: exhaustion (five items), cynicism (five items), and lack of professional efficacy (six items). When rating perceived stress, subjects were asked to take into consideration the last six months, and not only the actual time-point. The average scores for Scandinavian populations are around 2 for MBI-GS, [1], [38].
Subjects were excluded if they had previous history of psychosis, personality disorder, major or bipolar depression, alcohol or substance abuse, chronic fatigue, chronic pain, fibromyalgia, or neurological or endocrine disease. Subjects, who had experienced prominent stress factors in their private life or a major traumatic event at any time in their life, including sexual abuse, were also excluded. No daily medication was allowed during the two months prior to the study, except contraceptives. Review of past history of pharmacological treatment revealed no drugs known to affect brain structure (for example, psychopharmaca).
Sixty-eight healthy, right-handed, non-smoking volunteers (53 females and 15 males, age 37.5±7.2 years, range 27–51 years, education 13.0±2.35 years) with no history of chronic stress or heredity for neuropsychiatric disorders were used as the control group. The patient and control groups therefore had similar gender distributions, and both groups had a female dominance to accommodate with the female-dominated epidemiology of the condition studied [1]. The study was approved by the Ethics Committee at the Karolinska Institute, and written informed consent was received from each participant.
Before the interview, participants completed questionnaires in order to evaluate their stress symptoms and assess their previous life events. In addition, the occurrence of major life events among the subjects was assessed through a clinical psychiatric interview based on the non-work-related items of the Holmes and Rahe Scale [39]. The participants were asked to answer yes or no to whether they had experienced any non-work-related stressful life events (e.g., death of a relative or spouse, recent divorce, forced family relocation). Subjects were excluded if they answered positively to having experienced such an event in their lives. Patients also received a medical screening, (physical examination, test of thyroid and liver function). The possible presence of psychiatric disorders or personality disturbances were assessed according to the Diagnostic and Statistical Manual of the American Psychiatric Association, 4th Edition (DSM–IV), including the Structured Questionnaire for DSM-IV® Axis I and II (Structured Clinical Interview for DSM-IV® (SCID-I, and II) (American Psychiatric Publishing Inc, Arlington, 1997), along with a test for depression using the Montgomery-Asberg Depression scale [40].
Magnetic Resonance Imaging
Data acquisition
All magnetic resonance imaging data was acquired on a whole-body 1.5-Tesla MRI medical scanner (General Electric, Milwaukee, Wisconsin) equipped with an 8-channel phased array coil. The MRI protocol included the following scans: 1) 3D-weighted T1 SPGR images with 1 mm isotropic voxel size according to a previously described protocol [41]; and 2) 2D T2-weighted fast spin echo (FSE) images in the axial plane (effective TE = 56 ms, TR = 2500 ms, FOV = 24 cm, 23 slices of 3 mm thickness). The 2D images were not used in the present analysis.
Voxel-Based Morphometry (VBM)
We used a version of the VBM in the SPM5 package (www.fil.ion.ucl.ac.uk/spm) with the Gaser toolbox (http://dbm.neuro.uni-jena.de/vbm/) and Matlab 7.3 (Math Works, Natick, MA). The VBM pre-processing included five steps:
Check for scanner artifacts and gross anatomical abnormalities for each subject.
Set of image origin at the Anterior Commissure AC.
Using the Hidden Markov Random Field (HMRF) option in the segmentation of the VBM5 toolbox to minimize the noise level of the segmentation.
Using the Diffeomorphic Anatomical Registration Through Lie Algebra toolbox (DARTEL, Wellcome Department of Imaging Neuroscience, University College London, UK; http://www.fil.ion.ucl.ac.uk/spm) for a high-dimensional normalization protocol. We followed the standard version of John Ashburner's chapter including the MNI space transformation [42].
To restore the original volume information within each voxel, voxel values in the segmented images were modulated (multiplied) by the Jacobian determinants derived from the spatial normalization step. The analyses of modulated data allowed direct comparisons of regional differences in the amount of each tissue type.
After pre-processing, visually checking for homogeneity across the sample, smoothed (8 mm), modulated, and normalized images were obtained and used for the statistical analyses. These modulated volume images of the respective tissue types are hereafter referred to as GM, WM, and cerebrospinal fluid (CSF).
Structural volumetry
In regions with poor white and gray matter demarcation, as in basal ganglia, volumetry is regarded as more reliable than VBM analysis [43], and manual volumetry was, therefore, used in addition to VBM. Homologous VOIs were delineated manually for the hippocampus, caudate, and putamen in all the subjects.
All the volumes were delineated on original, un-reformatted T1 images using MRIcro software (www.sph.sc.edu/comd/rorden/mricro.html) by two investigators who were uninformed about the identity of the subjects. Values presented in Results and Table 1b were generated by investigator 1, who analyzed all the data (investigator 2 analyzed structural volumes from 15 randomized subjects in each study group). Both raters analyzed 10 images twice to establish the intra-rater reliability. VOIs were outlined according to previously described protocols [43], [44], [45]. In summary.
Table 1. Demographics.
| Patients (n = 30) | Controls (n = 68) | P and F values | |
| Age (years) | 41.3±6.6 | 37.5±7.2 | p = 0.378 F = 0.783 |
| Education (years) | 13.5±1.8 | 13.0±2.4 | p = 0.423 F = 0.648 |
| MBI- GS (score) | 4.2±1.1 | 2.2±0.5 | p<0.0001 F = 100.6 |
| MADRAS (score) | 10.6±6.3 | 7.3±3.9 | p = 0.12 F = 2.58 |
Age and education are expressed in years; MBI-GS is a questionnaire to score work-related stress symptoms. MADRAS = Montgomery Asberg Depression Scale.
The hippocampus: the hippocampus was traced according to the protocol of Watson et al. [46]. The posterior boundary of the hippocampus was defined as the first image in which the crus of the fornix became visible. The superior boundary consisted of the alveus; the lateral boundary was the inferior corn of the lateral ventricle; the ambient cistern was the medial boundary, whereas the WM of the parahippocampal gyrus represented the inferior boundary. The hippocampus VOI included the tail of the hippocampus.
The caudate nucleus: the caudate nuclei were traced separately, in accordance with Raz et al. [47]. The lateral ventricle was used as the medial boundary; the subcallosal fasciculus served as the anterior boundary; the stria terminalis was the posterior boundary (identified by a change in signal intensity relative to the caudate nucleus); and the anterior limb of the internal capsule served as the lateral boundary. The region of interest included the head, the body of the caudate, and the tail (excluding the portion that turned anterior). The nucleus accumbens was excluded as well.
The putamen: the delineation of the putamen started in the first slice where it became visible laterally to the caudate. The superior boundary was defined by the corona radiata, the internal capsule was the medial border, and the lateral border was the external capsule. The posterior limit was the point at which the putamen was no longer visible in the corona radiata. The lower limit was just above the amygdala, excluding nucleus accumbens, when the operator could see the most inferior aspect of the third ventricle and the chiasmatic cistern was no longer visible.
The hippocampus was delineated on coronal images, and the other structures on horizontal sections. All the volumes were then viewed and corrected on coronal, horizontal, as well as sagital images.
Statistical analysis
Group differences in age and education, as well as differences in the total intracranial volume (TIV), calculated as the total volume of GM + WM + CSF, and the total tissue volume (TV), calculated as the total volume of GM + WM, were tested with unpaired Students t-test (p<0.05). Group comparisons of the total GM and WM volumes were tested using analysis of covariance (ANCOVA, p<0.05), taking into consideration individual differences in TIV, which was used as the covariate in addition to age and sex (the latter was not strictly necessary as the groups were matched for sex). Group comparisons of relative structural volumes (VOI/TIV) were carried out with unpaired Students t-tests using the mean individual relative values of the two homologous VOIs for each type of structure as input values (p<0.016), after Bonferroni correction for the three separate regions). The aforementioned analyses were carried out with PASW Statistics 18 (SPSS Inc., Chicago, IL).
Group differences in GM and WM volumes within the regions that were expected to show changes in stressed subjects (the ACC and mPFC, the hippocampus, amygdala and the insular cortex) were tested with VBM by restricting the search space to a mask encompassing the amygdalae (both sides), hippocampi, the ACC, the medial and superior frontal gyrus, and the insular cortex (both sides). This mask was derived using the WFU Pick Atlas (maldjian@wfubmc.edu), by adding the respective regional areas as defined by the atlas into a confluent, large mask (Fig. 1). In addition, we employed explorative analysis using also the entire brain as the search space to investigate possible differences between the two subject groups in other regions. Significant clusters were defined with flexible factorial design in SPM5 (voxel threshold p<0.001, with FDR correction at p<0.05) using age, sex, and the total brain volume (TIV) as covariates of no interest. We also implemented non-stationary cluster extent correction in the Gaser toolbox to correct for the non-uniform smoothness in the VBM images.
Figure 1. Illustration of the limbic mask, constructed from the WFU-pick atlas (please see methods), superimposed on the mean grey matter images from the entire study population.
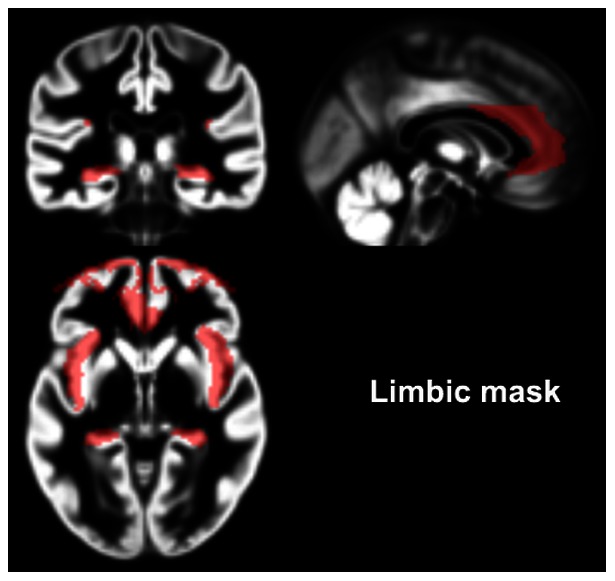
The co-ordinates are reported in Montreal Neurological Institute (MNI) space.
We hypothesized that changes in regional GM and WM, and in the structural volumes would be related to the degree of perceived stress. To test this, correlation analyses were carried out between MBI-GS scores and the individual GM or WM volumes extracted from the clusters showing significant group differences in GM and WM volumes as well as the relative volumes (the VOI/TIV ratios) of structures showing a significant difference between patients and controls. For the correlation analyses between VOI/TIV and MBI-GS, we used the mean of the two homologous regions (provided that a possible difference from controls was bilateral) in order to reduce the number of comparisons. The significance value for these linear regressions was set at p<0.05, because each of the regions included in this calculation was assumed to be affected by stress.
Inter- and intra-rater reliability was tested with linear correlation analysis, Pearson's coefficient, p<0.05.
Results
Clinical data
The groups did not differ in age or education. (Table 1).
No significant difference was detected between the two subject groups in respect to depression, as assessed by MADRAS scores (p = 0.12), although the mean value was higher among patients (Table 1). However, the MBI-GS scores, indicating perceived work-related stress levels, were significantly higher among the patients (4.2±1.1 vs. 2.2±0.5; p<0.0001; F = 100.6, df = 1); among controls, the scores were below 3.0 and in the range reported in other studies [1], [38], (Table 1).
No gross anatomical abnormalities were found, as judged by an experienced neuroradiologist, and none of the subjects had to be excluded due to either movement artifacts during scanning or segmentation errors.
We did not detect any group difference regarding total GM volume, WM volume, total TV, or the TIV (Table 2).
Table 2. Structural volumes.
| Volumes (cm3) | Patients | Controls | P and F values |
| L caudate volume | 3.8±0.5 | 4.6±0.6 | p<0.0001 F = 26.8 |
| R caudate volume | 3.8±0.5 | 4.5±0.6 | p<0.001 F = 11.7 |
| L putamen volume | 3.8±0.5 | 4.6±0.7 | p<0.0001 F = 23.4 |
| R putamen volume | 3.8±0.4 | 4.7±0.6 | p<0.0001 F = 25.8 |
| L hippocampus volume | 3.0±0.4 | 3.0±0.4 | p = 0.312 F = 1.054 |
| R hippocampus volume | 3.0±0.4 | 3,0±0.5 | p = 0.493 F = 0.483 |
| GM volume | 690.3±57.9 | 699.2±66.4 | p = 0.208 F = 1.608 |
| WM volume | 458.9±50.4 | 457.5±48.4 | p = 0.88 F = 1.148 |
| CSF volume | 338.4±100.9 | 457.5±48.4 | p = 0.86 F = 0.033 |
| TIV volume | 1487.6±163.3 | 1490.5±151.3 | p = 0.57 F = 0.331 |
TIV = total intracranial volume.
P-values for structural volumes were based on calculations of ratios between the respective structural volume and the TIV.
Structural volumetry
Patients showed a significantly lower structural volume relative to the TIV (VOI/TIV) bilaterally in the caudate and putamen (Table 2). The results remained when adding age and MADRAS as covariate (p<0.001 for both structures, post hoc analysis with ANCOVA). No significant group difference was detected in the volume of the hippocampus. The inter-rater correlation was 0.8 for the measurements of the hippocampus, and 0.9 for the caudate and putamen; the corresponding intra-rater values were 0.8, 1.0, respectively.
Voxel-based morphometry
Significant reductions in the GM volumes were detected in three clusters among the patients. They were located in the ACC (a region covered by the a priori hypothesis), and, in addition, in the left and right middle frontal gyrus (regions constituting portions of the dorsolateral prefrontal cortex – dlPFC), (Table 3, Fig. 2). Contrary to the hypothesis, we found no changes in the hippocampus, the amygdala, or the anterior insular cortex, (Table 3). No regional increases in GM were found in among patients. No group differences were detected in regional WM.
Table 3. Significant group difference in GM volume.
| Controls > Patients | Patients > Controls | |||
| Z-level | Size (cm3) | Coordinates | ||
| Left middle frontal gyrus | 4.3 | 1.9 | 25–11 59 | |
| Anterior cingulate cortex (BA 32) # | 3.4 | 0.5 | −39 6 46 | NONE |
| Right middle frontal gyrus | 5.0 | 2.1 | −2 31 13 | |
# Cluster showing a significant group difference when using a limbic mask comprising the anterior cingulate cortex, the mPFC, the insular cortex, the hippocampus and the amygdala, using peak threshold at p = 0.001, FDR corrected at p<0.05. The other clusters were calculated with same level of significance, but using the entire brain as search space (no a priori hypothesis).
No group differences were observed in white matter volumes and no significant clusters were detected when reversing the contrast (using the contrast: stressed patients – controls).
Figure 2. Reductions in GM volumes in stressed subjects.
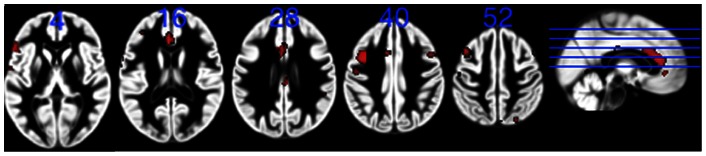
Significant reductions in the GM (red) in stressed subjects compared with controls. Clusters calculated using peak threshold at p = 0.001, FDR corrected at p<0.05. All the clusters are superimposed on the GM template from the entire study group. The numbers indicate z-levels in MNI co-ordinates. R = right side.
Post hoc group comparison when adding MADRAS as covariate of no interest did not alter the results.
Correlation analyses
No correlations were detected between stress scores (MBI-GS) and the individual GM volumes extracted from the clusters showing significant differences between controls and patients (the ACC, the left and right middle frontal gyrus). There was, however, a statistically significant, negative correlation between the relative structural volume of the caudate and putamen (mean of the right and left values for the respective structure) and the MBI-GS scores (r = −0.47 and p = 0.0001, r = −0.45 p = 0.0001, respectively), Fig. 3A. When estimating the regression lines separately for stressed subjects and controls, significant inverse correlations remained amongst the controls for both structures (r = −0.57, and r = −0.45; p<0.0001 for both), Fig. 3B, whereas amongst the stressed group there was only a tendency for an inverse correlation in the caudate (r = −0.36, p = 0.054), and no correlation with the relative volume of the putamen (Fig. 3C).
Figure 3. Correlation analysis between stress scores (MBI-GS) and the relative putamen volume (mean of the right and left putamen/TIV) – upper panel, and the relative volume of the caudate (mean of the right and left caudate/TIV) – lower panel.

A = All subjects; B = Control subjets only; C = Stressed subjects only.
Discussion
The present study tests the hypothesis that perceived prolonged occupational stress is associated with regional morphological changes in the brain. Such changes have been found in patients with PTSD (thus, in relation to life threatening trauma), in persons having experienced early life traumas, and in those reporting negative major life events [23], [27]. However, to the best of our knowledge, the possibility of such changes has not been previously discussed in relation to subjects undergoing occupational stress who have not faced extreme life conditions or psychosocial trauma and who have no history of psychiatric disorders.
The principal findings were the reductions in the GM volumes of the ACC and the dlPFC, and reductions in the volumes of the caudate and putamen. The basal ganglia volumes were also inversely correlated with the degree of perceived stress. All the structures in which we detected changes have been described to be sensitive to stress stimuli [21], [23], [48], [49] and are known to be involved in stress physiology [31], [33], [50], [51]. Decreased GM volume and density in the prefrontal cortex and the ACC in particular has been detected in several studies of persons suffering from PTSD [26], [27], [52], [53], [54]. Of particular interest is the data of Kasai et al., showing atrophy of the ACC, hippocampus and insular cortex in combat veterans with PTSD but not in their identical twins, suggesting that the detected abnormality was acquired [26]. Furthermore, measurements of cortical thickness in a separate group of patients with PTSD have shown dynamic, stress-related changes in the dlPFC which were characterized by an increase in cortical thickness that gradually normalized over time during recovery [55].
Data from studies on basal ganglia in corresponding populations are less abundant. There are reports about an affection of the caudate and putamen in relation to early life traumas [56] and PTSD, [57], but in general, these structures have received relatively minor interest in the context of stress. The present finding of reduced structural volumes in the caudate and putamen, therefore, deserves particular consideration. This finding is in accordance with the reported activations of the basal ganglia occurring during acute stress [31], [58], [59]. It is also compatible with the notion that both the putamen and caudate receive powerful glutamatergic input from the prefrontal cortex [60] and are susceptible to excitotoxicity [61], [62]. The present findings are also congruent with the well-known phenomenon of stress-related freezing [63] and raises the question of whether and how motor performance may be affected in persons suffering from chronic psychosocial stress.
The regions showing changes in our stressed patients are part of a network believed to mediate the integration of cognitive, affective, and autonomic responses [64].
It is tenable that an affection of these structures could lead to poor attention, and working memory deficits, symptoms which individuals with chronic occupational stress have described experiencing [5], [6], [65]. The preset data is in line with results from a fMRI study by Sandström et al., showing a decreased activation of the dlPFC among 10 patients suffering from occupational stress [6], and with a fMRI study by Qin et al., which detected a reduced activation of the dlPFC in healthy subjects who performed a working memory task under acute stress [51].
Contrary to our hypothesis, but in accordance with the presently available reports from MRI studies of subjects with occupational stress [5], [21], no changes were detected in the hippocampus or amygdala. Among studies of other chronic stress conditions, relatively few have examined amygdala volumes, and the results are variable. Among persons with PTSD, smaller volumes have been reported in some studies [66], [67], while others have detected normal values [68], [69], [70]. Similar inconsistency appears with regard to the hippocampus [5], [69], [71].
One possible explanation for the mixed findings could be that the small size of these structures, the amygdala in particular, may lead to greater variability in volume measures, which hampers the probability of detecting significant group differences, especially when the investigated study groups are undersized. Given the size of the present population and the use of 1.5 Tesla scanner, we did not try to specifically assess the structural volume of the amygdala in the present study. Another potential explanation may be associated with time of the stress exposure in relation to age as such exposure may have effects on these structures under sensitive developmental periods [72], [73].
The fact that the presently detected volume reductions were confined to regions believed to be involved in the processing of stress stimuli [23] makes it highly unlikely that they were random findings. The threshold used in the SPM analysis was in accordance with other VBM studies of psychosocial stress in humans [27], [74], [75] and the detected differences in the structural volumes of the caudate and putamen were highly significant.
At present, we can only speculate about the underpinnings of the observed changes. Because the study was cross-sectional, it is difficult to state whether the detected reductions represent the neurotoxic effect of stress, effect of other factors, such as nitric oxide, or are associated with a pre-existing condition that could have rendered the brain more vulnerable to the development of pathological stress responses. Due to our strict selection criteria, it is, however, possible to exclude potential confounding factors such as major life traumas, psychiatric premorbidity, depression, chronic pain, and pharmacological treatment. Considering the congruence with data from animal experiments as well as with the longitudinal data from patients with PTSD, we find it probable that the present findings reflect effects of chronic psychosocial stress, in this case, occupational stress. The observed inverse correlation between the stress scores and the relative volumes of the caudate and putamen (Fig. 3) could be taken as an argument for this view. It should be notified, however, that this correlation is dominated by the data from controls (r = −0.57, p = 0,0001, and r = −0.45, p = 0.0001, whereas the corresponding regression line in stressed subjects only showed a non-significant trend for the caudate (r = −0.36 p = 0.054), and no trend for the putamen (r = −0.08; p = 0.69). The reason for that is not evident, one possibility could be that the control group was much larger, and that the spread of volumes was larger among the controls. Worth mentioning is that in our recent follow up investigation of two new cohorts of controls and subjects with occupational stress, shows very similar results (Osika&Savic, manuscript in preparation).
The molecular underpinnings of the morphological changes after stress are just beginning to be studied in detail, please see Leuner and Shors 2012, and Conrad 2008, for a comprehensive review [76], [77]. The major mediators of stress-related neuronal modulation involving dendritic retraction and in some cases neurotoxicity are glucocorticoids and glutamate [23]. They have own neuronal effects, and are also reported to interact [78], [79]. Circulating glucocorticoids interact with various neurotransmitters, [80] and chronic stress in tree shrews is found to reduce the number of dopamine transporter (DAT) binding sites (Bmax) in the caudate nucleus and the putamen [81]. Adrenal ectomy is reported to increase the proliferation of hippocampal neurons whereas excess in glucocorticoid is reported to decrease it sharply, as do psychosocial stressors [82]. Experiments with betamethasone infusion in fetal sheep show glucocorticoid-related loss of synaptic density in the frontal neocortex, caudate, putamen, and hippocampus [83], [84]. While these alterations were initially viewed as a neurodegenerative event, it is worth mentioning that more recent studies suggest that stress induced dendritic alterations are reversible if animals are given time to recover from chronic stress [77].
The second stress-associated factor, which could have contributed to the observed changes, is glutamate. Data from animal experiments show that stress causes an enhanced release of glutamate, and that a stress-related elevation of extracellular glutamate levels induces retraction in the spines in stress-targeted regions, such as the mPFC, ACC, and the basal ganglia [79], [85], [86], [87], [88]. Medial prefrontal cortex is reported to be sensitive to repetitive stress [89]. See also the review by Leuner and Shors [77]. Via glutamatergic excitation the mPFC and ACC send inhibitory GABA-ergic impulses to the amygdala, which is the primary cerebral relay for the processing of psychosocial stress stimuli [90]. One possible scenario is that the absence of prefrontal inhibition of the amygdala, due to the stress-mediated neurotoxic damage of the mPFC (due to high glutamate, cortisol or the combination of both [79], [91], may cause amygdala hyperactivity [92], providing a context for a vicious circle with increased excitation and excitotoxic changes along the networks linked to the amygdala and mPFC. These networks primarily include the dlPFC, the basal ganglia, and the limbic brain (the hippocampus and insular cortex). Whereas the ACC, the dlPFC, and the basal ganglia showed distinct changes, neither the hippocampus nor the insular cortex was seemingly affected according to our results, even though these regions were included in the specific search space defined by our limbic mask. This was unexpected, especially when considering that both structures had significant reductions in 5-HT1A receptor binding potential in our previous study of a similar population [21]. The underlying reasons are not clear, and a larger population of subjects needs to be investigated before this lack of significance can be taken as an argument for specificity of the previously detected 5-HT1A receptor changes.
Methodological limitations and strengths
Several methodological limitations deserve comment. Firstly, the sample was rather small even though the sensitivity was enhanced by the use of a homogenous study group. Secondly, causality cannot be determined in the present study, and longitudinal studies would be needed to provide a better understanding of the temporal relationships between morphological brain alterations and periods of psychosocial stress. The issue of comorbidity with depression is always a problem in a study group like our – as some symptoms are overlapping and the condition of profound fatigue and inability to lead a normal life is perceived as extremely taxing by some subjects. The stressed subjects were not deemed as depressed by the experienced psychiatrist. We also re-run the comparisons between groups using MADRAs scores as covariate in addition to age and gender when comparing the mean relative caudate and putamen values between the groups. The results remained (p = 0.0061 for caudate and p = 0.0002 for putamen). Also in the VBM analysis, when controlling for depression by entering depression scores as covariate the results remained unchanged. Finally, neuropsychological data was not collected systematically, as we whished to investigate weather there are any anatomical changes in the brain that could be related to occupational stress, before designing a study to test possible behavioural correlates. Such study is on going.
The advantage of the present study is that it combines VBM analysis and investigations of structural volumes. In regions with poor white and gray matter demarcation, as in the basal ganglia, volumetry is regarded as more reliable than VBM [43], and the two methods should, therefore, be used in tandem. Consequently, it is not surprising that structural volumetry showed reductions in the basal ganglia while the corresponding GM and VM volumes did not differ from controls.
Conclusions
By investigating the GM and WM volumes and structural volumes of subjects who are experiencing chronic psychosocial stress, but do not have a history of a particular psychosocial trauma, the present study expands upon the data from animal experiments and reports from PTSD patients, and focuses on the involvement of the frontostriatal circuits in chronic stress. The finding of GM atrophy in regions known to be associated with chronic psychosocial stress confirms our previous conclusion that subjects reporting stereotyped symptoms from occupational stress have a medical condition requiring careful investigations and a targeted treatment. Larger series and consecutive investigations before and after treatments, as well as parallel mapping of cognitive and motor functions are highly encouraged in the future, and might have important implications for the understanding of this increasingly common condition.
Acknowledgments
We are extremely grateful to Dr. Walter Osika for his careful patient recruitment. Dr. Savic had full access to all of the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Funding Statement
The study was supported by the Swedish Council for Working Life and Social Research (FAS), Swedish Research Council, AFA Insurance, Stockholm City Council, as well as the Centre for Gender Related Medicine, Karolinska Institute. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Ahola K, Honkonen T, Isometsa E, Kalimo R, Nykyri E, et al. (2006) Burnout in the general population. Results from the Finnish Health 2000 Study. Soc Psychiatry Psychiatr Epidemiol 41: 11–17. [DOI] [PubMed] [Google Scholar]
- 2. Copertaro A, Barbaresi M, Tarsitani L, Battisti F, Baldassari M, et al. (2007) [Fast stress evaluation in nurses]. Giornale italiano di medicina del lavoro ed ergonomia 29: 350–352. [PubMed] [Google Scholar]
- 3. Copertaro A, Bracci M, Amati M, Mocchegiani E, Barbaresi M, et al. (2010) [Biological risk and health care workers: analysis of the effects of work chronobiology on the immune system]. Med Lav 101: 427–436. [PubMed] [Google Scholar]
- 4. Fernandez Torres B, Roldan Perez LM, Guerra Velez A, Roldan Rodriguez T, Gutierrez Guillen A, et al. (2006) [Prevalence of burnout among anesthesiologists at Hospital Universitario Virgen Macarena de Sevilla]. Rev Esp Anestesiol Reanim 53: 359–362. [PubMed] [Google Scholar]
- 5. Rydmark I, Wahlberg K, Ghatan PH, Modell S, Nygren A, et al. (2006) Neuroendocrine, cognitive and structural imaging characteristics of women on longterm sickleave with job stress-induced depression. Biological psychiatry 60: 867–873. [DOI] [PubMed] [Google Scholar]
- 6. Sandstrom A, Rhodin IN, Lundberg M, Olsson T, Nyberg L (2005) Impaired cognitive performance in patients with chronic burnout syndrome. Biol Psychol 69: 271–279. [DOI] [PubMed] [Google Scholar]
- 7. Asberg M, Grape T, Krakau I, Nygren A, Rohde M, et al. (2010) [Stress as the cause of mental illness]. Lakartidningen 107: 1307–1310. [PubMed] [Google Scholar]
- 8. Ulrich-Lai YM, Herman JP (2009) Neural regulation of endocrine and autonomic stress responses. Nat Rev Neneurosci 6: 397–409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Fries E, Dettenborn L, Kirschbaum C (2009) The cortisol awakening response (CAR): facts and future directions. Int J Psychophysiol 72: 67–73. [DOI] [PubMed] [Google Scholar]
- 10. Langelaan S, Bakker AB, Schaufeli WB, van Rhenen W, van Doornen LJ (2007) Is burnout related to allostatic load? Int J Behav Med 14: 213–221. [DOI] [PubMed] [Google Scholar]
- 11. Mommersteeg PM, Heijnen CJ, Verbraak MJ, van Doornen LJ (2006) Clinical burnout is not reflected in the cortisol awakening response, the day-curve or the response to a low-dose dexamethasone suppression test. Psychoneuroendocrinology 31: 216–225. [DOI] [PubMed] [Google Scholar]
- 12. Moch SL, Panz VR, Joffe BI, Havlik I, Moch JD (2003) Longitudinal changes in pituitary-adrenal hormones in South African women with burnout. Endocrine 21: 267–272. [DOI] [PubMed] [Google Scholar]
- 13. Mommersteeg PM, Keijsers GP, Heijnen CJ, Verbraak MJ, van Doornen LJ (2006) Cortisol deviations in people with burnout before and after psychotherapy: a pilot study. Health Psychol 25: 243–248. [DOI] [PubMed] [Google Scholar]
- 14. Pruessner JC, Hellhammer DH, Kirschbaum C (1999) Burnout, perceived stress, and cortisol responses to awakening. Psychosomatic medicine 61: 197–204. [DOI] [PubMed] [Google Scholar]
- 15. Chida Y, Steptoe A (2009) Cortisol awakening response and psychosocial factors: a systematic review and meta-analysis. Biol Psychol 80: 265–278. [DOI] [PubMed] [Google Scholar]
- 16. Grossi G, Perski A, Ekstedt M, Johansson T, Lindstrom M, et al. (2005) The morning salivary cortisol response in burnout. J Psychosom Res 59: 103–111. [DOI] [PubMed] [Google Scholar]
- 17. Grossi G, Perski A, Evengard B, Blomkvist V, Orth-Gomer K (2003) Physiological correlates of burnout among women. J Psychosom Res 55: 309–316. [DOI] [PubMed] [Google Scholar]
- 18. Melamed S, Ugarten U, Shirom A, Kahana L, Lerman Y, et al. (1999) Chronic burnout, somatic arousal and elevated salivary cortisol levels. J Psychosom Res 46: 591–598. [DOI] [PubMed] [Google Scholar]
- 19. Friess E, Schmid D, Modell S, Brunner H, Lauer CJ, et al. (2008) Dex/CRH-test response and sleep in depressed patients and healthy controls with and without vulnerability for affective disorders. J Psychiatr Res 42: 1154–1162. [DOI] [PubMed] [Google Scholar]
- 20. Wahlberg K, Ghatan PH, Modell S, Nygren A, Ingvar M, et al. (2009) Suppressed neuroendocrine stress response in depressed women on job-stress-related long-term sick leave: a stable marker potentially suggestive of preexisting vulnerability. Biol Psychiatry 65: 742–747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Jovanovic H, Perski A, Berglund H, Savic I (2011) Chronic stress is linked to 5-HT(1A) receptor changes and functional disintegration of the limbic networks. NeuroImage 55: 1178–1188. [DOI] [PubMed] [Google Scholar]
- 22.Bremner JD (2007) Neuroimaging in posttraumatic stress disorder and other stress-related disorders. Neuroimaging Clin N Am 17: 523–538, ix. [DOI] [PMC free article] [PubMed]
- 23. Bremner JD, Elzinga B, Schmahl C, Vermetten E (2008) Structural and functional plasticity of the human brain in posttraumatic stress disorder. Prog Brain Res 167: 171–186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Cohen RA, Grieve S, Hoth KF, Paul RH, Sweet L, et al. (2006) Early life stress and morphometry of the adult anterior cingulate cortex and caudate nuclei. Biological psychiatry 59: 975–982. [DOI] [PubMed] [Google Scholar]
- 25. Geuze E, Westenberg HG, Heinecke A, de Kloet CS, Goebel R, et al. (2008) Thinner prefrontal cortex in veterans with posttraumatic stress disorder. NeuroImage 41: 675–681. [DOI] [PubMed] [Google Scholar]
- 26. Kasai K, Yamasue H, Gilbertson MW, Shenton ME, Rauch SL, et al. (2008) Evidence for acquired pregenual anterior cingulate gray matter loss from a twin study of combat-related posttraumatic stress disorder. Biological psychiatry 63: 550–556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Papagni SA, Benetti S, Arulanantham S, McCrory E, McGuire P, et al. (2011) Effects of stressful life events on human brain structure: a longitudinal voxel-based morphometry study. Stress 14: 227–232. [DOI] [PubMed] [Google Scholar]
- 28. Pavlisa G, Papa J, Pavic L, Pavlisa G (2006) Bilateral MR volumetry of the amygdala in chronic PTSD patients. Coll Antropol 30: 565–568. [PubMed] [Google Scholar]
- 29. van Harmelen AL, van Tol MJ, van der Wee NJ, Veltman DJ, Aleman A, et al. (2010) Reduced medial prefrontal cortex volume in adults reporting childhood emotional maltreatment. Biological psychiatry 68: 832–838. [DOI] [PubMed] [Google Scholar]
- 30.Chen L, Lui S, Wu QZ, Zhang W, Zhou D, et al..(2011) Impact of acute stress on human brain microstructure: An MR diffusion study of earthquake survivors. Hum Brain Mapp. [DOI] [PMC free article] [PubMed]
- 31. Goldstein JM, Jerram M, Abbs B, Whitfield-Gabrieli S, Makris N (2010) Sex differences in stress response circuitry activation dependent on female hormonal cycle. J Neurosci 30: 431–438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. McEwen BS (1998) Stress, adaptation, and disease. Allostasis and allostatic load. Annals of the New York Academy of Sciences 840: 33–44. [DOI] [PubMed] [Google Scholar]
- 33. Wang J, Rao H, Wetmore GS, Furlan PM, Korczykowski M, et al. (2005) Perfusion functional MRI reveals cerebral blood flow pattern under psychological stress. Proceedings of the National Academy of Sciences of the United States of America 102: 17804–17809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Baker LM, Williams LM, Korgaonkar MS, Cohen RA, Heaps JM, et al.. (2012) Impact of early vs. late childhood early life stress on brain morphometrics. Brain Imaging Behav. [DOI] [PMC free article] [PubMed]
- 35. Flugge G (1995) Dynamics of central nervous 5-HT1A-receptors under psychosocial stress. J Neurosci 15: 7132–7140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9: 97–113. [DOI] [PubMed] [Google Scholar]
- 37. Schaufeli WB, Van Dierendonck D (1995) A cautionary note about the cross-national and clinical validity of cut-off points for the Maslach Burnout Inventory. Psychological reports 76: 1083–1090. [DOI] [PubMed] [Google Scholar]
- 38. Stenlund T, Ahlgren C, Lindahl B, Burell G, Knutsson A, et al. (2007) Patients with burnout in relation to gender and a general population. Scand J Public Health 35: 516–523. [DOI] [PubMed] [Google Scholar]
- 39. Holmes TH, Rahe RH (1967) The Social Readjustment Rating Scale. J Psychosom Res 11: 213–218. [DOI] [PubMed] [Google Scholar]
- 40.Montgomery S, Asberg M, Traskman L, Montgomery D (1978) Cross cultural studies on the use of CPRS in English and Swedish depressed patients. Acta Psychiatr Scand Suppl: 33–37. [DOI] [PubMed]
- 41. Ciumas C, Wahlin TB, Espino C, Savic I (2010) The dopamine system in idiopathic generalized epilepsies: identification of syndrome-related changes. Neuroimage 51: 606–615. [DOI] [PubMed] [Google Scholar]
- 42. Ashburner J (2007) A fast diffeomorphic image registration algorithm. NeuroImage 38: 95–113. [DOI] [PubMed] [Google Scholar]
- 43. Ciumas C, Savic I (2006) Structural changes in patients with primary generalized tonic and clonic seizures. Neurology 67: 683–686. [DOI] [PubMed] [Google Scholar]
- 44. Helms G, Ciumas C, Kyaga S, Savic I (2006) Increased thalamus levels of glutamate and glutamine (Glx) in patients with idiopathic generalised epilepsy. J Neurol Neurosurg Psychiatry 77: 489–494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Savic I, Lindstrom P (2008) PET and MRI show differences in cerebral asymmetry and functional connectivity between homo- and heterosexual subjects. Proc Natl Acad Sci U S A 105: 9403–9408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Watson C, Andermann F, Gloor P, Jones-Gotman M, Peters T, et al. (1992) Anatomic basis of amygdaloid and hippocampal volume measurement by magnetic resonance imaging. Neurology 42: 1743–1750. [DOI] [PubMed] [Google Scholar]
- 47. Raz N, Lindenberger U, Rodrigue KM, Kennedy KM, Head D, et al. (2005) Regional brain changes in aging healthy adults: general trends, individual differences and modifiers. Cereb Cortex 15: 1676–1689. [DOI] [PubMed] [Google Scholar]
- 48. McEwen BS (2007) Physiology and neurobiology of stress and adaptation: central role of the brain. Physiol Rev 87: 873–904. [DOI] [PubMed] [Google Scholar]
- 49. Sinha R, Lacadie C, Skudlarski P, Wexler BE (2004) Neural circuits underlying emotional distress in humans. Annals of the New York Academy of Sciences 1032: 254–257. [DOI] [PubMed] [Google Scholar]
- 50. Dedovic K, Rexroth M, Wolff E, Duchesne A, Scherling C, et al. (2009) Neural correlates of processing stressful information: an event-related fMRI study. Brain research 1293: 49–60. [DOI] [PubMed] [Google Scholar]
- 51. Qin S, Hermans EJ, van Marle HJ, Luo J, Fernandez G (2009) Acute psychological stress reduces working memory-related activity in the dorsolateral prefrontal cortex. Biological psychiatry 66: 25–32. [DOI] [PubMed] [Google Scholar]
- 52. Nardo D, Hogberg G, Looi JC, Larsson S, Hallstrom T, et al. (2010) Gray matter density in limbic and paralimbic cortices is associated with trauma load and EMDR outcome in PTSD patients. J Psychiatr Res 44: 477–485. [DOI] [PubMed] [Google Scholar]
- 53. Thomaes K, Dorrepaal E, Draijer N, de Ruiter MB, van Balkom AJ, et al. (2010) Reduced anterior cingulate and orbitofrontal volumes in child abuse-related complex PTSD. J Clin Psychiatry 71: 1636–1644. [DOI] [PubMed] [Google Scholar]
- 54. Yamasue H, Kasai K, Iwanami A, Ohtani T, Yamada H, et al. (2003) Voxel-based analysis of MRI reveals anterior cingulate gray-matter volume reduction in posttraumatic stress disorder due to terrorism. Proceedings of the National Academy of Sciences of the United States of America 100: 9039–9043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Lyoo IK, Kim JE, Yoon SJ, Hwang J, Bae S, et al. (2011) The neurobiological role of the dorsolateral prefrontal cortex in recovery from trauma. Longitudinal brain imaging study among survivors of the South Korean subway disaster. Archives of general psychiatry 68: 701–713. [DOI] [PubMed] [Google Scholar]
- 56. Sheu YS, Polcari A, Anderson CM, Teicher MH (2010) Harsh corporal punishment is associated with increased T2 relaxation time in dopamine-rich regions. NeuroImage 53: 412–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Filipovic BR, Djurovic B, Marinkovic S, Stijak L, Aksic M, et al. (2011) Volume changes of corpus striatum, thalamus, hippocampus and lateral ventricles in posttraumatic stress disorder (PTSD) patients suffering from headaches and without therapy. Cent Eur Neurosurg 72: 133–137. [DOI] [PubMed] [Google Scholar]
- 58. Liston C, McEwen BS, Casey BJ (2009) Psychosocial stress reversibly disrupts prefrontal processing and attentional control. Proceedings of the National Academy of Sciences of the United States of America 106: 912–917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Wang J, Korczykowski M, Rao H, Fan Y, Pluta J, et al. (2007) Gender difference in neural response to psychological stress. Soc Cogn Affect Neurosci 2: 227–239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. McEwen BS (2000b) Protective and damaging effects of stress mediators: central role of the brain. Prog Brain Res 122: 25–34. [DOI] [PubMed] [Google Scholar]
- 61. Bernal F, Saura J, Ojuel J, Mahy N (2000) Differential vulnerability of hippocampus, basal ganglia, and prefrontal cortex to long-term NMDA excitotoxicity. Exp Neurol 161: 686–695. [DOI] [PubMed] [Google Scholar]
- 62. Chen Q, Harris C, Brown CS, Howe A, Surmeier DJ, et al. (1995) Glutamate-mediated excitotoxic death of cultured striatal neurons is mediated by non-NMDA receptors. Exp Neurol 136: 212–224. [DOI] [PubMed] [Google Scholar]
- 63. Siemiatkowski M, Rokicki D, Czlonkowska AI, Sienkiewicz-Jarosz H, Bidzinski A, et al. (2000) Locomotor activity and a conditioned fear response: correlation with cortical and subcortical binding of the dopamine D1 receptor antagonist. Neuroreport 11: 3953–3956. [PubMed] [Google Scholar]
- 64. Critchley HD (2009) Psychophysiology of neural, cognitive and affective integration: fMRI and autonomic indicants. Int J Psychophysiol 73: 88–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Jovanovic H, Cerin A, Karlsson P, Lundberg J, Halldin C, et al. (2006) A PET study of 5-HT1A receptors at different phases of the menstrual cycle in women with premenstrual dysphoria. Psychiatry Res 148: 185–193. [DOI] [PubMed] [Google Scholar]
- 66. Matsuoka Y, Yamawaki S, Inagaki M, Akechi T, Uchitomi Y (2003) A volumetric study of amygdala in cancer survivors with intrusive recollections. Biological psychiatry 54: 736–743. [DOI] [PubMed] [Google Scholar]
- 67. Rogers MA, Yamasue H, Abe O, Yamada H, Ohtani T, et al. (2009) Smaller amygdala volume and reduced anterior cingulate gray matter density associated with history of post-traumatic stress disorder. Psychiatry Res 174: 210–216. [DOI] [PubMed] [Google Scholar]
- 68. Bonne O, Brandes D, Gilboa A, Gomori JM, Shenton ME, et al. (2001) Longitudinal MRI study of hippocampal volume in trauma survivors with PTSD. Am J Psychiatry 158: 1248–1251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Gilbertson MW, Shenton ME, Ciszewski A, Kasai K, Lasko NB, et al. (2002) Smaller hippocampal volume predicts pathologic vulnerability to psychological trauma. Nat Neurosci 5: 1242–1247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Wignall EL, Dickson JM, Vaughan P, Farrow TF, Wilkinson ID, et al. (2004) Smaller hippocampal volume in patients with recent-onset posttraumatic stress disorder. Biological psychiatry 56: 832–836. [DOI] [PubMed] [Google Scholar]
- 71. Villarreal G, Hamilton DA, Petropoulos H, Driscoll I, Rowland LM, et al. (2002) Reduced hippocampal volume and total white matter volume in posttraumatic stress disorder. Biological psychiatry 52: 119–125. [DOI] [PubMed] [Google Scholar]
- 72. Andersen SL, Teicher MH (2008) Stress, sensitive periods and maturational events in adolescent depression. Trends Neurosci 31: 183–191. [DOI] [PubMed] [Google Scholar]
- 73. Andersen SL, Tomada A, Vincow ES, Valente E, Polcari A, et al. (2008) Preliminary evidence for sensitive periods in the effect of childhood sexual abuse on regional brain development. J Neuropsychiatry Clin Neurosci 20: 292–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Chen S, Xia W, Li L, Liu J, He Z, et al. (2006) Gray matter density reduction in the insula in fire survivors with posttraumatic stress disorder: a voxel-based morphometric study. Psychiatry Res 146: 65–72. [DOI] [PubMed] [Google Scholar]
- 75. Zhang J, Tan Q, Yin H, Zhang X, Huan Y, et al. (2011) Decreased gray matter volume in the left hippocampus and bilateral calcarine cortex in coal mine flood disaster survivors with recent onset PTSD. Psychiatry Res 192: 84–90. [DOI] [PubMed] [Google Scholar]
- 76. Conrad CD (2008) Chronic stress-induced hippocampal vulnerability: the glucocorticoid vulnerability hypothesis. Rev Neurosci 19: 395–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Leuner B, Shors TJ (2012) Stress, anxiety, and dendritic spines: What are the connections? Neuroscience. [DOI] [PubMed]
- 78. Brown ES, Zaidel L, Allen G, McColl R, Vazquez M, et al. (2010) Effects of lamotrigine on hippocampal activation in corticosteroid-treated patients. Journal of affective disorders 126: 415–419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Magarinos AM, McEwen BS (1995) Stress-induced atrophy of apical dendrites of hippocampal CA3c neurons: involvement of glucocorticoid secretion and excitatory amino acid receptors. Neuroscience 69: 89–98. [DOI] [PubMed] [Google Scholar]
- 80. McEwen BS (2000a) The neurobiology of stress: from serendipity to clinical relevance. Brain research 886: 172–189. [DOI] [PubMed] [Google Scholar]
- 81. Isovich E, Mijnster MJ, Flugge G, Fuchs E (2000) Chronic psychosocial stress reduces the density of dopamine transporters. Eur J Neurosci 12: 1071–1078. [DOI] [PubMed] [Google Scholar]
- 82. Wong EY, Herbert J (2004) The corticoid environment: a determining factor for neural progenitors' survival in the adult hippocampus. Eur J Neurosci 20: 2491–2498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83. Colberg C, Antonow-Schlorke I, Muller T, Schubert H, Witte OW, et al. (2004) Recovery of glucocorticoid-related loss of synaptic density in the fetal sheep brain at 0.75 of gestation. Neurosci Lett 364: 130–134. [DOI] [PubMed] [Google Scholar]
- 84. Conrad CD, LeDoux JE, Magarinos AM, McEwen BS (1999) Repeated restraint stress facilitates fear conditioning independently of causing hippocampal CA3 dendritic atrophy. Behav Neurosci 113: 902–913. [DOI] [PubMed] [Google Scholar]
- 85. Hunter RG, Bellani R, Bloss E, Costa A, McCarthy K, et al. (2009) Regulation of kainate receptor subunit mRNA by stress and corticosteroids in the rat hippocampus. PloS one 4: e4328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86. Iijima M, Shimazaki T, Ito A, Chaki S (2007) Effects of metabotropic glutamate 2/3 receptor antagonists in the stress-induced hyperthermia test in singly housed mice. Psychopharmacology 190: 233–239. [DOI] [PubMed] [Google Scholar]
- 87. Lowy MT, Gault L, Yamamoto BK (1993) Adrenalectomy attenuates stress-induced elevations in extracellular glutamate concentrations in the hippocampus. Journal of neurochemistry 61: 1957–1960. [DOI] [PubMed] [Google Scholar]
- 88. McEwen BS, Conrad CD, Kuroda Y, Frankfurt M, Magarinos AM, et al. (1997) Prevention of stress-induced morphological and cognitive consequences. Eur Neuropsychopharmacol 7 Suppl 3S323–328. [DOI] [PubMed] [Google Scholar]
- 89. Brown SM, Henning S, Wellman CL (2005) Mild, short-term stress alters dendritic morphology in rat medial prefrontal cortex. Cereb Cortex 15: 1714–1722. [DOI] [PubMed] [Google Scholar]
- 90. Herman JP, Ostrander MM, Mueller NK, Figueiredo H (2005) Limbic system mechanisms of stress regulation: hypothalamo-pituitary-adrenocortical axis. Progress in neuro-psychopharmacology & biological psychiatry 29: 1201–1213. [DOI] [PubMed] [Google Scholar]
- 91. Martin KP, Wellman CL (2011) NMDA receptor blockade alters stress-induced dendritic remodeling in medial prefrontal cortex. Cereb Cortex 21: 2366–2373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Roozendaal B, McReynolds JR, McGaugh JL (2004) The basolateral amygdala interacts with the medial prefrontal cortex in regulating glucocorticoid effects on working memory impairment. J Neurosci 24: 1385–1392. [DOI] [PMC free article] [PubMed] [Google Scholar]