

Abstract
Eicosanoids (prostaglandins, leukotrienes and lipoxins) are a family of signaling lipids derived from arachidonic acid that have important roles in physiological and pathological processes. Over the past years, it has been established that successful eicosanoid production is not merely determined by arachidonic acid and eicosanoid-forming enzymes availability, but requires sequential interactions between specific biosynthetic proteins acting in cascade and may involve very unique spatial interactions. Direct assessment of specific subcellular locales of eicosanoid synthesis has been elusive, as those lipid mediators are newly formed, not stored and often rapidly released upon cell stimulation. In this chapter, we discuss the EicosaCell protocol for intracellular detection of eicosanoid-synthesizing compartments by means of a strategy to covalently cross-link and immobilize the lipid mediators at their sites of synthesis followed by immunofluorescent-based localization of the targeted eicosanoid.
Keywords: Eicosanoids, prostaglandin, leukotriene, biosynthesis, compartmentalization, carbodiimide, EDAC (1-ethyl-3-(3-dimethylaminopropyl) carbodiimide), lipid droplets, phagosomes, perinuclear
1. Introduction
Eicosanoids – including leukotrienes and prostaglandins – are a family of signaling lipids derived from the enzymatic oxygenation of arachidonic acid (AA) that control key processes involving cell–cell communication, including cell activation, proliferation, apoptosis, metabolism and migration (1, 2). Thus, eicosanoids have important roles in physiological and pathological conditions such as tissue homeostasis, host defense, inflammation and cancer. In view of the magnitude of eicosanoid actions, great efforts have been aimed at understanding the biochemical, cellular and molecular aspects of their biosynthetic pathway.
In all cells, the highly regulated generation of eicosanoids is dependent on activation of specific phospholipases and specific eicosanoid-synthesizing enzymes and involves small molecules (e.g. Ca2+) and activation-dependent localization of enzymes at specific compartments within cells (3–8). Intracellular compartmentalization of eicosanoid synthesis within leukocytes has emerged as a key feature that regulates the amount and may also regulate the eicosanoid produced. Such intracellular sites of eicosanoid formation in any cell have been inferred based on the permanent or temporary localization of specific eicosanoid-forming enzymes under proper cell activation, since the direct observation of sites of eicosanoid synthesis has been hard to define as those lipid mediators are newly formed, non-storable and often rapidly released upon cell stimulation. It was recently established that successful eicosanoid production is not merely determined by AA and eicosanoid-forming enzymes availability, but requires sequential interactions between specific biosynthetic proteins acting in cascade, and may involve very unique spatial interactions. Therefore, just by detecting eicosanoid-forming enzymes within discrete subcellular structures, one cannot assure that those sites are indeed accountable for the efficient and enhanced eicosanoid synthesis observed during inflammatory responses. The immunolocalization of eicosanoid-forming proteins does not necessarily ascertain that the localized protein is functional and activated to synthesize a specific eicosanoid lipid at an intracellular site. We previously developed a method to capture and localize the eicosanoid, prostaglandin E2 (PG E2), released extracellularly by a nematode parasite (9). By means of a strategy to covalently cross-link, capture and localize newly formed eicosanoids at their sites of synthesis, we developed a more direct approach to detect the intracellular sites of arachidonic acid (AA)-derived lipid mediator formation in leukocytes and other cell types.
To develop our new strategy for in situ immunolocalization of newly formed eicosanoids to ascertain the intracellular compartmentalization of their synthesis – the EicosaCell assay – modifications of a prior technique was used (9). The EicosaCell rational relies on the specific features of the heterobifunctional cross-linker 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (C8H17N3-HCl; EDAC) used. EDAC immobilizes newly synthesized eicosanoids by cross-linking the eicosanoid carboxyl groups to the amines of adjacent proteins localized at eicosanoid-synthesizing compartment. Such EDAC-mediated reaction forms a bond without any spacer length between the two molecules, favoring an accurate positioning of the newly synthesized eicosanoid within the cell. In addition, while other cross-linkers formed bonds that often generate foreign molecules, EDAC-driven eicosanoid-bond is homologous to native eicosanoid that allows immunoassays like EicosaCell. Besides the precise positioned coupling of an immunodetectable eicosanoid at its sites of formation, EDAC enables: (I) the ending of cell stimulation step; (II) cell fixation; (III) cell permeabilization, allowing the penetration of both anti-eicosanoid and the detecting fluorochrome-conjugated antibodies into cells; and, importantly, (IV) the relative preservation of lipid domains, such as membranes and droplets, which dissipate with air drying or commonly used alcohol fixation.
2. Materials
2.1. Conventional EicosaCell
EDAC (1-ethyl-3-(3-dimethylaminopropyl) carbodiimide hydrochloride) is diluted in Hanks-buffered salt solution without calcium chloride and magnesium chloride (HBSS−/−). Refer to Note 1 for EDAC solution handling. EDAC final concentration with cells varies according to cell type and protocol used (see next subheadings). The working solution should have twice concentration of the final concentration with cells. For instance, specifically regarding purified human eosinophils stimulated as a cell suspension, EDAC final concentration with eosinophils should be 0.1% in HBSS−/−, therefore the EDAC working solution should be diluted to 0.2%. Alternatively, with adherent macrophages stimulated in 6 wells plate, EDAC final concentration should be 0.5% in HBSS−/−, therefore the EDAC working solution should be diluted to 1.0%.
Primary antibody to the eicosanoid of interest.
Fluorescent-labeled secondary antibodies.
Glass microscope slides and coverslips.
Anti-fading mounting medium for fluorescence.
2.2. Double-Labeling Purposes
DAPI (4′, 6′-Diamidino-2-phenylindole dihydrochloride) stock solution is prepared by dissolving 1 mg/mL of powder in distilled water. Aliquots should be stored at −20° protected from light.
Monoclonal antibody against lysosome-associated membrane protein (LAMP) 1.
BODIPY® 493/503 (4,4-difluoro-1,3,5,7,8-pentamethyl-4-bora-3a,4a-diaza-s-indacene) (Molecular Probes; cat no. D-3922, molecular weight: 262). To prepare BODIPY stock solution, BODIPY should be dissolved in DMSO (1 mM), aliquoted in small Eppendorf tubes (~10 μL per tube) and stored at −20°C protected from light. BODIPY working solution should be diluted fresh 1000× in HBSS−/− and kept from light.
Monoclonal or polyclonal antibody to adipose differentiation-related protein (ADRP).
3. Methods
3.1. EicosaCell with Cells in Suspension
EicosaCell can be easily performed with a varied of cell types in suspension, such as purified human blood leukocytes, cell lineages, as well as, peritoneal, pleural or bronchoalveolar animal cells. After in vivo or in vitro stimulation of these cell populations, incubation with EDAC should instantaneously guarantee the immobilization of eicosanoids at their synthesizing spot within the cell, just before cytospin slides are prepared to allow microscopic analysis. As schematically illustrated in Fig. 10.1a, after preparing a cell suspension, EDAC working solution should be added to cell suspension and incubated for a period of time to ensure cell fixation, immobilization of eicosanoid and cell permeabilization.
Fig. 10.1.

Schematic illustration of EicosaAssay method. EicosaCell preparations, which undergo EDAC-dependent capturing and fixation of newly formed-eicosanoids at their sites of synthesis, are analyzed by phase-contrast and fluorescence microscopy and can employ cytospun cells (a), adherent cells or cells embedded in a gel matrix (b).
Prepare a cell suspension of 2 × 106/mL. Gently and immediately add an equal volume of EDAC solution, prepared as described in Section 2.1, Step 1 (refer to Notes 1 and 2 for details), to the cell suspension.
Incubate the cell suspension with EDAC for 30 min to 1 h at 37°C.
Cytospin the cells onto slides using 100 μL of the cell suspension at 23 g for 5 min.
Wash twice in HBSS−/−.
Labeling of newly formed eicosanoids can be done with a variety of already tested antibodies, as already published elsewhere (10–13). Incubate cells with the primary antibody to the eicosanoid of interest for 1 h at room temperature. The non-immune serum from the animal where the secondary antibody was produced may be added to the primary antibody so as to decrease unspecific labeling.
Wash 2–3 times in HBSS−/−.
Incubate with the fluorescent-labeled secondary antibody for 1 h at room temperature.
At the end of the staining procedure, cytospun cells should be always extensively washed with HBSS−/−, at least 3 times for 5 min each.
Slides should be mounted using an aqueous mounting medium, preferentially with anti-fading.
Analysis is performed on phase contrast to observe cell morphology and fluorescence microscope or confocal scanning laser microscope to identify the eicosanoid labeling. For example, analyses and image acquisition can be obtained using an Olympus BX-FLA fluorescence microscope equipped with a Plan Apo 100× 1.4 Ph3 objective and CoolSNAP-Pro CF digital camera in conjunction with Image Pro Plus® software (Media Cybernetics) (see Note 3 for details).
The specificity of the eicosanoid immunolabeling using EicosaCell system should be always ascertained by including some mandatory control conditions as detailed in Note 4.
As shown in Fig. 10.2, EicosaCell system was successfully employed on macrophages recovered from pleural cavities of BCG-infected or control mice (11). Briefly, cells obtained 24 h after infection with BCG and controls were recovered from the pleural cavity with 500 μL of HBSS−/− and immediately mixed with 500 μL of EDAC (1% in HBSS−/−). After 30 min incubation at 37°C with EDAC, pleural leukocytes were then washed with HBSS−/−, cytospun onto glass slides and incubated with mouse anti-PGE2 in 0.1% normal goat serum and guinea pig polyclonal anti-mouse ADRP (see Section 2.2) in 0.1% normal donkey serum simultaneously for 1 h at room temperature. Isotyping matching antibodies (murine IgG1) were used as controls (Fig. 10.2). Cells were washed twice and incubated with secondary antibodies, goat anti-mouse conjugated with AlexaFluor-488 (1/1000, Molecular Probes) and CY3-conjugated donkey anti-guinea pig (1/1000). Slides were washed (three times, 10 min each) and mounted with aqueous mounting medium. Cells were analyzed by both phase-contrast and fluorescence microscopy. As a control for PGE2 specificity of detection, one group of BCG-infected animals was treated with indomethacin (4 mg/kg), 4 h before sacrificing animals for cell recovery (not shown).
Fig. 10.2.

EicosaCell for PGE2 immunolocalization within BCG-infected cytospun macrophages. In upper panels, macrophages from BCG-infected animals were labeled for ADRP-associated lipid bodies (red staining) and for newly formed PGE2 (green staining). Merged image showed co-localization of PGE2 in ADRP-associated lipid bodies (yellow staining). In bottom panels, IgG1 irrelevant isotype (MOPC) was used as control for PGE2 labeling. Briefly, pleural macrophages obtained 24 h after infection with BCG were recovered from the thoracic cavity with 500 μL of HBSS, immediately mixed with 500 μL of EDAC (1% in HBSS) and incubated for 30 min at 37°C. Cells were then washed with HBSS, cytospun onto glass slides and incubated with mouse anti-PGE2 (1/100) or MOPC 21 in 0.1% normal goat serum and guinea pig polyclonal anti-mouse ADRP (1/1000) in 0.1% normal donkey serum simultaneously for 1 h at RT. Cells were washed twice and incubated with secondary Abs, goat anti-mouse conjugated with AlexaFluor-488 (1/1000) and CY3-conjugated donkey anti-guinea pig (1/1000). The slides were washed (three times, 10 min each) and mounted with aqueous mounting medium.
3.2. EicosaCell with Adherent Cells
To study the intracellular compartmentalization of eicosanoid synthesis by EicosaCell in adherent cells, extra care should be taken to ensure the conservation of cell adherence and morphology during EDAC step. EicosaCell have succeeded to immunolocalize PGE2 within at least three distinct cell types: plated murine macrophages (D’Avila et al., unpublished) and two lineages of intestinal cells, CACO-2 (a human colon adenocarcinoma cell line) (Fig. 10.3) (14) and IEC-6 (a rat epithelial cell line (15)).
Fig. 10.3.

EicosaCell for PGE2 immunolocalization within adherent CACO-2 cells. The largest panel shows fluorescent microscopy of CACO-2 cells labeled for newly formed PGE2 (red staining). Bottom three images panel showed immunofluorescent PGE2 (red staining), BODIPY-associated lipid bodies (green staining) and a merged image showing co-localization of PGE2 in lipid bodies (yellow staining). Insert panel showed lack of PGE2 immunolabeling within lipid-body-enriched CACO-2 cells, which were treated with indomethacin (4 mg/kg) 1 h before EDAC. Briefly, CACO-2 cells were fixed and permeabilized during 1 h at 37°C with EDAC (0.5% in HBSS−/−). Then, cells were washed with HBSS and blocked with 2% donkey serum for 15 min before incubation with anti-PGE2 monoclonal antibody (Cayman Chemicals) for 45 min. Cells were washed with HBSS and incubated with fluorescent secondary antibody Cy3-conjugated affiniPure F(ab′) fragment donkey anti-mouse and BODIPY 493/503 (Molecular Probes, CA) for 45 min.
While adherent on glass coverslips, cells can be incubated for 30 min to 1 h at 37°C with EDAC at 0.5% in HBSS−/− to cross-link the lipid mediator of interest to carboxyl groups to amines in adjacent proteins (Fig. 10.3) without affecting cell morphology (refer to Note 5). Alternatively, cells grown in Lab-Tek chambers can be used.
Gently wash cells 2–3 times in HBSS−/−.
Incubate cells with the primary antibody to the eicosanoid of interest for 1 h at room temperature. The non-immune serum from the animal where the secondary antibody was produced may be added to the primary antibody so as to decrease unspecific labeling.
Gently wash 2–3 times in HBSS−/−.
Incubate with the fluorescent-labeled secondary antibody for 1 h at room temperature.
At the end of the staining procedure, cells should be gently washed with HBSS−/−, at least 3 times for 5 min each.
Cell-containing coverslips should be carefully glued to the slide and mounted using an aqueous mounting medium, preferentially with anti-fading.
Analysis is performed on phase contrast to observe cell morphology and fluorescence microscope or confocal scanning laser microscope to identify the eicosanoid labeling. For example, analyses and image acquisition can be obtained using an Olympus BX-FLA fluorescence microscope equipped with a Plan Apo 100 × 1.4 Ph3 objective (Olympus) and CoolSNAP-Pro CF digital camera in conjunction with Image Pro Plus® software (Media Cybernetics) (see Note 3 for details).
As for cytospun cells, the specificity of the eicosanoid immunolabeling using EicosaCell system should be ascertained by including mandatory controls listed in Note 4.
3.3. EicosaCell with Cells Embedded in a Gel Matrix
In contrast to analyzing cytospun cells which do not preserve in situ morphology, cells embedded in an agarose matrix, that are kept in a hydrated system with a substrate where they can crawl, display tissue-like cell morphology exhibiting polarization and other characteristics of activated leukocyte, for instance. Therefore, by immunolocalizing eicosanoids at its formation sites within agarose-embedded cells (as schematically illustrated in Fig. 10.1b), generated products may be microscopically localized at cell structures assembled during stimulation and preserved in cells that are not cytospun into slides (10, 16).
To prepare the agarose matrix, 2.5% agarose (24 C gelling point) (Promega) in sterile distilled H2O, is melted at 70°C; and while liquid at 37°C, 9 volumes of agarose are mixed with 1 volume of 10× concentrated RPMI 1640 medium.
One volume of this medium-supplemented agarose is mixed with one volume of RPMI 1640 medium containing 2% fatty acid free-albumin at 37°C and with three volumes of the studied cell, exemplified here as human eosinophils (Fig. 10.1b), which should be at 15 × 106 cells/mL in RPMI 1640 medium containing 1% fatty acid free-human albumin.
Stimuli are then added in 0.1 volumes to agarose/eosinophil mixtures. As schematically illustrated in Fig. 10.1b, immediately thereafter, 20 μL samples are gently spread onto microscope slides and covered with perfusion chamber (CoverWell™).
Each slide is overlaid with RPMI 1640 medium containing 1% albumin and an identical concentration of the stimulus present in the agarose/eosinophil mixture.
Slides can be incubated (37°C, humidified 5% CO2) for varying periods of time.
. Overlying medium should be removed and replaced with RPMI 1640, 1% albumin medium, that may contain or not 0.1 μM calcium ionophore (A23187) and incubated for extra 15 min (37°C; 5% CO2).
Stimulations are stopped by removing chambers and adding EDAC. Fixation and permeabilization of cells with proper immobilization of newly formed eicosanoids at its intracellular sites of synthesis are achieved by immersing the slides containing stimulated cells in 0.5% EDAC (in HBSS−/−) for 30 min.
-
After three washes (5 min each) with HBSS−/−, the fluorochrome-labeled anti-eicosanoid, for instance Alexa488-labeled rat anti-cysteinyl leukotriene (LT) detection mAb (Sigma) (Alexa™488 protein labeling using a kit from Molecular Probes) should be added (400 μL of 10 μg/mL) for 1 h.
Slides need to be extensively washed with HBSS−/−, at least 3 times for 5 min each.
Aqueous mounting medium should be applied to each slide before coverslip attachment.
Slides can be viewed by both phase-contrast and fluorescence microscopy as detailed above – Section 3.1 (Step 11).
Mandatory control conditions, as listed in Note 4, should be always included as for cytospun and adherent cells to ascertain the specificity of the eicosanoid immunolabeling using EicosaCell system.
3.4. Double-Labeling Procedures to Identify Eicosanoid-Synthesizing Intracellular Sites
3.4.1. Nuclear Localization
To better visualize perinuclear eicosanoid synthesis by EicosaCell, a double labeling with DAPI is advised.
After EDAC and antibody incubation steps, EicosaCell slides preparations should be extensively washed in HBSS−/− and then incubated with DAPI (DAPI working solution 100 ng/mL or 300 nM, see Section 2.2, Step 1) for 5 min before aqueous mounting medium application.
The morphology of the cells’ nuclei is observed using a fluorescence microscope at excitation wavelength 350 nm.
3.4.2. Phagosomal Localization
As performed by Balestrieri and coworkers (17), phagosome involvement in eicosanoid synthesis can be ascertained by co-localizing the phagosomal protein marker LAMP-1 in EicosaCell preparations.
After incubation at 37°C with EDAC, cells should be washed with HBSS−/−, cytospun onto glass slides and incubated with the anti-eicosanoid of interest and the primary antibody against LAMP-1 (2.5 μg/mL) in blocking buffer (5% normal donkey serum) for 2 h at room temperature.
Negative control cells are instead incubated for 2 h with appropriate IgG.
After 2 h, the cells are washed extensively with HBSS−/− and incubated for 1 h at room temperature with fluorescent-labeled secondary antibody to detect the primary antibody against the eicosanoid of interest and with fluorescent-labeled secondary antibody to detect the primary antibody against LAMP-1(1:200).
The cells should be washed five times with HBSS−/− and then mounted in aqueous mounting medium.
3.4.3. Lipid Body Localization
To investigate lipid body role in eicosanoid synthesis by Eicosa-Cell assay, two double-labeling strategies can be employed: BODIPY or anti-ADRP immunostaining (for further information on lipid body labeling refer to Chapter 9). Both approaches can be used for adherent, suspension or agarose-embedded cells.
3.4.3.1. BODIPY® 493/503 Lipid Body-Labeling
To employ BODIPY® 493/503 strategy, incubate Eicosa-Cell preparations (coverslips or slides) with 1 μm BODIPY (working solution) simultaneously to secondary antibody incubation for 45–60 min at 37°C (water bath).
To remove free BODIPY after incubation, EicosaCell preparations should be washed at least twice in HBSS−/− before aqueous mounting medium application and coverslip attachment to slides.
3.4.3.2. ADRP Lipid Body-Labeling
Alternatively, to visualize lipid bodies, anti-ADRP immunolabeling may be performed as detailed in Chapter 9.
Add anti-ADRP antibodies together with the anti-eicosanoid antibody of interest for 1 h at room temperature. (a) For human cells incubate with mouse anti-human ADRP at dilution of 1:20 (2.5 μg/mL, final concentration); (b) For mouse, rat, human or bovine cells, incubate with guinea pig anti-human ADRP polyclonal antibody at dilution of 1:300 (final dilution).
Wash three times in HBSS−/− or PBS.
Incubate with fluorescent-labeled secondary antibody for 1 h room temperature (together with the secondary antibody for the EicosaCell labeling.
Wash three times in HBSS−/− or PBS.
Mount in mounting medium for fluorescence microscopy. Common problems and non-obvious features found in immunofluorescent-detection of eicosanoids in EDAC preparations by EicosaCell with their possible explanations and potential solutions are described in Note 6.
5. Applications
Over the past decade, intracellular compartmentalization of eicosanoid-synthetic machinery has emerged as a key component of the regulation of eicosanoid synthesis (reviewed in (4–6, 19). However, the direct evaluation of specific subcellular locales of eicosanoid synthesis has been elusive, as those lipid mediators are newly formed, not stored and often rapidly released upon cell stimulation. Thus, in the majority of studies, intracellular sites of eicosanoid synthesis have been inferred based on the identification of eicosanoid-forming enzymes localization.
The EicosaCell technique described herein enables to directly pinpoint the intracellular locales of eicosanoid synthesis by detecting the newly formed lipids and has been successfully able to confirm the dynamic aspect involved in the localization of eicosanoid synthesis, providing direct evidence of compartmentalization within perinuclear envelope (10, 14, 15, 20), phagosomes (17) or lipid bodies in accord to cell type and stimulatory condition (10–12, 15, 21); (Figs. 10.4 and 10.5). So far, the EicosaCell assay has been used to identify the production of leukotriene C4 (LTC4) (10, 13, 17, 22), leukotriene B4 (LTB4) (12, 23), prostaglandin E2 (PGE2) (11, 14, 15) and prostaglandin D2 (PGD2) (unpublished observations) in different cell types and under different stimulatory conditions. Moreover, it could in principle be adapted to intracellular detection of other lipid mediators as long as specific antibodies are available.
Fig. 10.4.
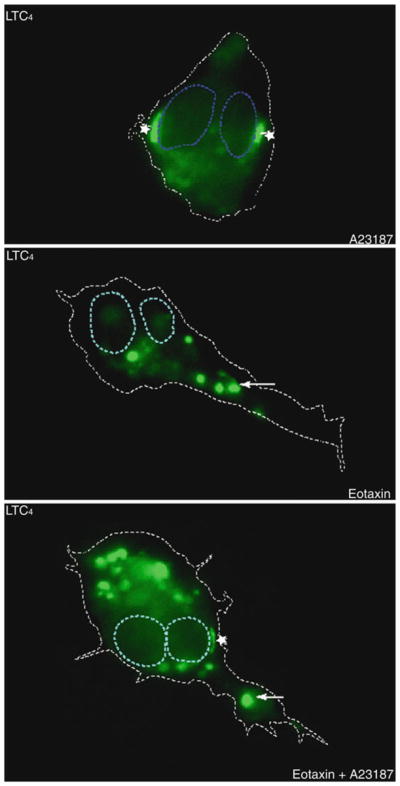
EicosaCell for LTC4 immunolabeling within gel-immersed human eosinophils. Fluorescent microscopy of agarose-embedded eosinophils, fixed with EDAC (0.5% in HBSS−/−) and stained with Alexa488-labeled anti-cysteinyl LT mAb. To facilitate intracellular localization of anti-LTC4 immunoreactive sites (green staining) within representative eosinophils, blue and white dotted lines were drawn to delineate, respectively, the nuclear and cellular perimeters. As indicated, A23187-, eotaxin- or eotaxin plus A23187-stimulated eosinophils display fluorescent LTC4 immunostaining. While, eosinophils stimulated with A23187 (0.1 μM) for 15 min exhibited exclusively perinuclear (stars) immunoreactive LTC4, eotaxin (100 ng/mL)-stimulated eosinophils showed punctate cytoplasmic lipid body-comprised LTC4 (arrows). Differently, eosinophils stimulated with eotaxin for 1 h and activated for extra 15 min with A23187 exhibit perinuclear (stars) and punctate cytoplasmic (arrows) immunoreactive LTC4.
Fig. 10.5.
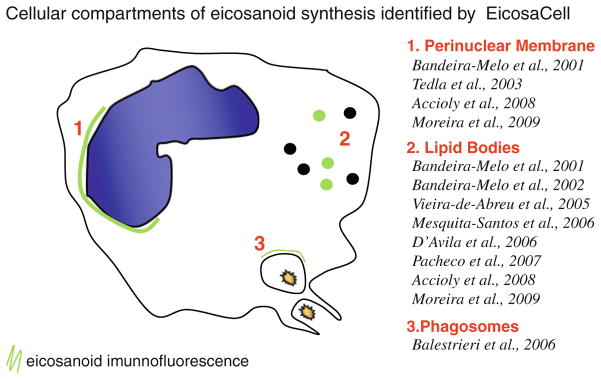
Schematic summary of EiocosaCell-derived reports identifying three distinct intracellular compartments of eicosanoid synthesis: the nuclear envelope, cytoplasmic lipid bodies and zymozan-driven phagosomes.
Of note, the EicosaCell Assay has high sensitivity, enabling the detection of low levels of intracellular generated eicosanoids even when extracellular released eicosanoids could not be detected by conventional eicosanoid enzyme immune assay (10, 15). Indeed, it has been shown that besides paracrine/autocrine activities, eicosanoids may display intracrine functions (22, 24). For instance, by employing EicosaCell technique, it has been uncovered that a lipid-body-derived LTC4 have intracellular functions in controlling cytokine release from eosinophils (25). Therefore, by identifying compartmentalized levels of eicosanoids, besides providing new insights of regulation of eicosanoid biosynthesis, EicosaCell assay may contribute to identification of likely intracellular functions of newly synthesized eicosanoids.
Acknowledgments
The work of authors is supported by PRONEX-MCT, Conselho Nacional de Desenvolvimento Cientifico e Tecnológico (CNPq, Brazil), PAPES-FIOCRUZ, Fundação de Amparo à Pesquisa do Rio de Janeiro (FAPERJ, Brazil) and NIH grants (AI022571, AI020241, AI051645). Authors are indebted with Dr. Heloisa D’Avila and Dr. Patricia Pacheco for the contributions to figures used in the manuscript.
Footnotes
EDAC working solution should be prepared fresh, kept protected from light and discarded after each experiment.
Incubation of cells with EDAC can be carried out on either cell in suspension or with the cells already cytospun onto slides by dropping EDAC on top of the cells. Even though the latter method is less costly, some differences in preservation of cell morphology, cell permeabilization and eicosanoid detection may occur and should be analyzed with care.
Analysis of EicosaCell preparations should be performed as soon as slides are mounted, inasmuch as immunofluorescent labeling is usually not stable for a long period bleaching after a certain time. Even though freezing may preserve fluorescence overnight, EDAC-treated cells may display altered cell appearance after freezing–thawing cycle.
The specificity of the eicosanoid immunolabeling using EicosaCell system should be always ascertained by including some mandatory control conditions: (i) non-stimulated EDAC-treated cells labeled with the proper anti-eicosanoid antibody; (ii) the incubation (1 h min before EDAC) with the eicosanoid synthesis inhibitors, such as cPLA2-a inhibitor (e.g. pyrrolidine-2; 1 μM), COX inhibitor (e.g. indomethacin; 1 μg/mL), FLAP inhibitor (e.g. MK886; 50 μg/animal or 10 μM for in vitro incubations) or 5-LO inhibitor (e.g. zileuton; 50 μg/animal or 10 μM for in vitro incubations) to avoid the eicosanoid synthesis and (iii) the use of an irrelevant antibody control. Optionally, other suitable controls to check specificity and performance of Eicosa-Cell are (i) to use, instead of EDAC, paraformaldehyde, which will not immobilize the newly synthesized eicosanoid within cells; (ii) to, in parallel, carry out the EicosaCell in a different cell type that lacks the ability to synthesize the targeted eicosanoid (for instance, to use neutrophils to check specificity of LTC4 immunodetection by EicosaCell) or (iii) to analyze mixed populations of responsive plus unresponsive cells to a specific stimulus, so you can reassure that the targeted eicosanoid is specifically detected only within stimulated cells.
While adherent CACO 2 cells can be incubated with EDAC for 1 h, IEC-6 cells can be incubated for at most 30 min (at same concentration; 0.5% in HBSS−/−) to retain reasonable cell morphology and PGE2 immunodetection at synthesizing compartments (refer to the original articles (14, 15) for details of blocking and staining conditions with anti-PGE2 monoclonal antibody (Cayman Chemicals) and proper secondary antibodies.
Common problems and non-obvious features found in immunofluorescent-detection of eicosanoids in EDAC preparations by EicosaCell:
Lack of eicosanoid detection: When few or no eicosanoid specific immunostaining is observed (but expected), the problem usually lies in the improper fixation (e.g. EDAC-driven cross-linking) of targeted eicosanoid at its sites of synthesis. Thus, the newly formed eicosanoid would be washed-out from the EicosaCell preparation turning detection impossible. Resolution of this problem is normally achieved by adjusting (slight increase) concentration and/or time of incubation of EDAC. Alternatively, the lack of immunodetection of newly formed eicosanoids can be due to inefficient stimulation; a positive control with a known agonist should be always included in experiments.
Eicosanoid detection within non-stimulated cells: Eicosanoids are lipid mediators non-storable in the cell and newly formed upon stimulation, therefore non-stimulated cells should not show any immunostaining for the targeted eicosanoid. Thus, non-stimulated cells should always be included as an important negative control. However, cell activation during procedures including cell incubation at 37°C or cell fixation/permeabilization with EDAC can lead to spontaneous, stimulus-independent eicosanoid synthesis. Throughout cell preparation, care is needed to ensure that cells are not mechanically, chemically or immunologically stimulated. Unexpected eicosanoid detection within Eicosa-Cell preparations can also result from non-specific detection (discussed below).
Non-specific detection: Fluorescent detection antibodies may non-specifically bind to other lipids found within cells or bind to other cellular structures. The crosslinking properties of EDAC may favor the tendency for cells to be sticky; therefore antibodies could interact through low-affinity non-antigen binding site. To investigate non-specific binding in EicosaCell preparations, a proper control using host/isotype-matched irrelevant antibodies, must be always included. An additional mandatory control that needs to be always included in the experimental design to rule out non-specific immunostaining is the condition with a synthesis inhibitor of the targeted eicosanoid. Synthesis inhibitor-treated controls should show no immune-labeling confirming specific detection of targeted eicosanoid. If non-specific staining is too high (>10% positive), there are several possible remedies. The detecting antibody may be diluted further, or a different one from a different host may be tried. Also, it is possible to try an adsorbing reagent that effectively blocks out non-specific sites, such as a normal serum (same host of the detecting antibody). Non-specific fluorescence can also be detected when the solution of detecting antibody contains a high degree of aggregated antibody; therefore, it is important to centrifuge the detecting antibody before adding to cell preparations.
Poor preservation of cell morphology: During EDAC incubation step of EicosaCell assays, cell appearance may change from unimportant to severe modification of typical cell morphology. This undesirable effect of EDAC on cells can be avoided by adjusting both EDAC concentration and incubation time.
Losing cell adherence with EDAC: Similar to unwanted EDAC effect on cell morphology, the ability of cells to stay adhered to coverslips or other substrates can be affected by EDAC incubation. Again, previous careful setting of EDAC incubation step is obligatory and should be adjusted for each cell type.
Lost of cell integrity: Eicosanoid localization within cells by EicosaAssay may be tricky sometimes since some cell types are destroyed during EDAC-driven cross-linking/permeabilization step. For instance, even though lipid bodies of human neutrophils and basophils are sites of 5-LO localization (10, 18), EicosaCell assays with agarose-embedded neutrophils and basophils were not feasible since these cells did not endure to EDAC-driven fixation/permeabilization process, which precedes eicosanoid immunodetection by EicosaCell, indicating that the combination of gel matrix with EDAC step may be useful to study only a small group of tough cells, like eosinophils. Compartmentalization studies of eicosanoid synthesis within more fragile cells like neutrophils and basophils, however, can be carried out with EicosaCell system in non-gel solutions.
References
- 1.Yaqoob P. Fatty acids as gatekeepers of immune cell regulation. Trends Immunol. 2003;24:639–645. doi: 10.1016/j.it.2003.10.002. [DOI] [PubMed] [Google Scholar]
- 2.Wymann MP, Schneiter R. Lipid signaling in disease. Nat Rev Mol Cell Biol. 2008;9:162–176. doi: 10.1038/nrm2335. [DOI] [PubMed] [Google Scholar]
- 3.Smith WL, DeWitt DL, Garavito RM. Cyclooxygenases: structural, cellular, and molecular biology. Annu Rev Biochem. 2000;69:145–182. doi: 10.1146/annurev.biochem.69.1.145. [DOI] [PubMed] [Google Scholar]
- 4.Bozza PT, Magalhaes K, Weller PF. Leukocyte lipid bodies – biogenesis and functions in inflammation. Biochim Biophys Acta. 2009;1791:540–51. doi: 10.1016/j.bbalip.2009.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Mandal AK, Skoch J, Bacskai BJ, Hyman BT, Christmas P, Miller D, Yamin TT, Xu S, Wisniewski D, Evans JF, Soberman RJ. The membrane organization of leukotriene synthesis. Proc Natl Acad Sci USA. 2004;101:6587–6592. doi: 10.1073/pnas.0308523101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Peters-Golden M, Brock TG. Intracellular compartmentalization of leukotriene synthesis: unexpected nuclear secrets. FEBS Lett. 2001;487:323–326. doi: 10.1016/s0014-5793(00)02374-7. [DOI] [PubMed] [Google Scholar]
- 7.Diaz BL, Arm JP. Phospholipase A(2). Prostaglandins leukotrienes essent. Fatty Acids. 2003;69:87–97. doi: 10.1016/s0952-3278(03)00069-3. [DOI] [PubMed] [Google Scholar]
- 8.Bandeira-Melo C, Weller PF. Eosinophils and cysteinyl leukotrienes. Prostaglandins Leukot Essent Fatty Acids. 2003;69:135–143. doi: 10.1016/s0952-3278(03)00074-7. [DOI] [PubMed] [Google Scholar]
- 9.Liu LX, Buhlmann JE, Weller PF. Release of prostaglandin E2 by microfilariae of Wuchereria bancrofti and Brugia malayi. Am J Trop Med Hyg. 1992;46:520–523. doi: 10.4269/ajtmh.1992.46.520. [DOI] [PubMed] [Google Scholar]
- 10.Bandeira-Melo C, Phoofolo M, Weller PF. Extranuclear lipid bodies, elicited by CCR3-mediated signaling pathways, are the sites of chemokine-enhanced leukotriene C4 production in eosinophils and basophils. J Biol Chem. 2001;276:22779–22787. doi: 10.1074/jbc.M101436200. [DOI] [PubMed] [Google Scholar]
- 11.D’Avila H, Melo RCN, Parreira GG, Werneck-Barroso E, Castro-Faria-Neto HC, Bozza PT. Mycobacterium bovis bacillus Calmette-Guerin induces TLR2-mediated formation of lipid bodies: intracellular domains for eicosanoid synthesis in vivo. J Immunol. 2006;176:3087–3097. doi: 10.4049/jimmunol.176.5.3087. [DOI] [PubMed] [Google Scholar]
- 12.Pacheco P, Vieira-de-Abreu A, Gomes RN, Barbosa-Lima G, Wermelinger LB, Maya-Monteiro CM, Silva AR, Bozza MT, Castro-Faria-Neto HC, Bandeira-Melo C, Bozza PT. Monocyte chemoattractant protein-1/CC chemokine ligand 2 controls microtubule-driven biogenesis and leukotriene B4-synthesizing function of macrophage lipid bodies elicited by innate immune response. J Immunol. 2007;179:8500–8508. doi: 10.4049/jimmunol.179.12.8500. [DOI] [PubMed] [Google Scholar]
- 13.Vieira-de-Abreu A, Assis EF, Gomes GS, Castro-Faria-Neto HC, Weller PF, Bandeira-Melo C, Bozza PT. Allergic challenge-elicited lipid bodies compartmentalize in vivo leukotriene C4 synthesis within eosinophils. Am J Respir Cell Mol. 2005;33:254–261. doi: 10.1165/rcmb.2005-0145OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Accioly MT, Pacheco P, Maya-Monteiro CM, Carrossini N, Robbs BK, Oliveira SS, Kaufmann C, Morgado-Diaz JA, Bozza PT, Viola JP. Lipid bodies are reservoirs of cyclooxygenase-2 and sites of prostaglandin-E2 synthesis in colon cancer cells. Cancer Res. 2008;68:1732–1740. doi: 10.1158/0008-5472.CAN-07-1999. [DOI] [PubMed] [Google Scholar]
- 15.Moreira LS, Piva B, Gentile LB, Mesquita-Santos FP, D’Avila H, Maya-Monteiro CM, Bozza PT, Bandeira-Melo C, Diaz BL. Cytosolic phospholipase A2-driven PGE2 synthesis within unsaturated fatty acids-induced lipid bodies of epithelial cells. Biochim Biophys Acta. 2009;1791:156–165. doi: 10.1016/j.bbalip.2009.01.003. [DOI] [PubMed] [Google Scholar]
- 16.Bandeira-Melo C, Sugiyama K, Woods LJ, Phoofolo M, Center DM, Cruikshank WW, Weller PF. IL-16 promotes leukotriene C(4) and IL-4 release from human eosinophils via CD4- and autocrine CCR3-chemokine-mediated signaling. J Immunol. 2002;168:4756–4763. doi: 10.4049/jimmunol.168.9.4756. [DOI] [PubMed] [Google Scholar]
- 17.Balestrieri B, Hsu VW, Gilbert H, Leslie CC, Han WK, Bonventre JV, Arm JP. Group V secretory phospholipase A2 translocates to the phagosome after zymosan stimulation of mouse peritoneal macrophages and regulates phagocytosis. J Biol Chem. 2006;281:6691–6698. doi: 10.1074/jbc.M508314200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pacheco P, Bozza FA, Gomes RN, Bozza M, Weller PF, Castro-Faria-Neto HC, Bozza PT. Lipopolysaccharide-induced leukocyte lipid body formation in vivo: innate immunity elicited intracellular loci involved in eicosanoid metabolism. J Immunol. 2002;169:6498–6506. doi: 10.4049/jimmunol.169.11.6498. [DOI] [PubMed] [Google Scholar]
- 19.Bandeira-Melo C, Bozza PT, Weller PF. The cellular biology of eosinophil eicosanoid formation and function. J Allergy Clin Immunol. 2002;109:393–400. doi: 10.1067/mai.2002.121529. [DOI] [PubMed] [Google Scholar]
- 20.Tedla N, Bandeira-Melo C, Tassinari P, Sloane DE, Samplaski M, Cosman D, Borges L, Weller PF, Arm JP. Activation of human eosinophils through leukocyte immunoglobulin-like receptor 7. Proc Natl Acad Sci USA. 2003;100:1174–1179. doi: 10.1073/pnas.0337567100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Mesquita-Santos FP, Vieira-de-Abreu A, Calheiros AS, Figueiredo IH, Castro-Faria-Neto HC, Weller PF, Bozza PT, Diaz BL, Bandeira-Melo C. Cutting edge: prostaglandin D2 enhances leukotriene C4 synthesis by eosinophils during allergic inflammation: synergistic in vivo role of endogenous eotaxin. J Immunol. 2006;176:1326–1330. doi: 10.4049/jimmunol.176.3.1326. [DOI] [PubMed] [Google Scholar]
- 22.Devchand PR, Keller H, Peters JM, Vazquez M, Gonzalez FJ, Wahli W. The PPARalpha-leukotriene B4 pathway to inflammation control. Nature. 1996;384:39–43. doi: 10.1038/384039a0. [DOI] [PubMed] [Google Scholar]
- 23.Silva AR, Pacheco P, Vieira-De-Abreu A, Dalegria B, Bandeira-Melo C, Castro-Faria-Neto HC, Bozza PT. Lipid bodies in oxidized LDL-induced foam cells are leukotriene-synthesizing organelles: a MCP-1/CCL2 regulated phenomenon. Biochim Biophys Acta. 2009;1791:1066–75. doi: 10.1016/j.bbalip.2009.06.004. [DOI] [PubMed] [Google Scholar]
- 24.Kliewer SA, Lenhard JM, Willson TM, Patel I, Morris DC, Lehmann JM. A prostaglandin J2 metabolite binds peroxisome proliferator-activated receptor gamma and promotes adipocyte differentiation. Cell. 1995;83:813–819. doi: 10.1016/0092-8674(95)90194-9. [DOI] [PubMed] [Google Scholar]
- 25.Bandeira-Melo C, Woods LJ, Phoofolo M, Weller PF. Intracrine cysteinyl leukotriene receptor-mediated signaling of eosinophil vesicular transport-mediated interleukin-4 secretion. J Exp Med. 2002;196:841–850. doi: 10.1084/jem.20020516. [DOI] [PMC free article] [PubMed] [Google Scholar]