

Abstract
Pericytes are small cells that are apposed to brain and meningeal microvasculature and control capillary contraction, thereby regulating local cerebral perfusion. Pericytes respond to exogenously applied glutamate in vitro and express metabotropic glutamate receptors. However, it is unclear if pericytes have the capacity to release glutamate. We therefore determined whether pericytes express vesicular glutamate transporters (VGLUTs), which are considered to be unambiguous markers of cells that use glutamate as an intercellular signaling molecule. Leptomeningeal and brain microvasculature-associated pericytes of the adult rat, as defined by the presence of NG2 proteoglycan, expressed both VGLUT2- and VGLUT3-immunoreactivity, but did not express VGLUT1. Consistent with the hypothesis that pericytes release glutamate, VGLUT2- and VGLUT3-immunoreactivities appeared to be localized to secretory vesicles. These results suggest that glutamate is released from pericytes of the leptomeninges and brain microvasculature, and demonstrate for the first time the co-localization of VGLUT2 and VGLUT3.
Keywords: Glutamate, Lysosome, Pericyte, Vesicle, VGLUT2, VGLUT3
Pericytes reside within the basal lamina of microvasculature, adhering to arterioles, venules, and capillaries. Within the microvasculature of brain parenchyma and the leptomeninges, where these cells are referred to as central nervous system (CNS) pericytes, they perform a variety of functions, including the regulation of local cerebral blood flow and macrophage activity. The phagocytotic immune function of CNS pericytes is subserved by numerous lysosomes located within the cytoplasm, leading to a granular appearance of pericytes [5]. CNS pericytes are also replete with plasmalemmal and cytoplasmic vesicles [6,10], but the function of these organelles is unknown.
CNS pericytes have ovular to oblong cell bodies spaced along brain parenchymal blood vessels at intervals of ~50 µm [27]. Protruding from the cell body are processes that extend along and wrap around microvasculature. A recent report [18] demonstrated that the processes of CNS pericytes, which express both actin and myosin filaments [3,12], contract and relax to bidirectionally control capillary diameter. Consistent with this observation are data showing that electrical stimulation of the cortex causes arteriolar dilation not only at the site of stimulation but also in upstream arterioles that branch to supply the cortical area that was stimulated [14]. Moreover, electrical stimulation of a CNS pericyte in cerebellar slice cultures elicits contractile responses from upstream and downstream CNS pericytes along the same microvessel [18]. Taken together, these data suggest the presence of bidirectional communication between spatially segregated CNS pericytes, and thus some form of “paracrine” intercellular signaling between these cells.
A growing body of evidence indicates that CNS pericytes respond to signaling molecules, including neurotransmitters. Upon exogenous application, certain signaling molecules elicit CNS pericyte-induced capillary constriction or dilation [18]. Among these is glutamate, which reverses norepinephrine-induced CNS pericyte constriction of capillaries. It has also been demonstrated that adult rat CNS pericytes express the metabotropic glutamate receptors mGluR1α, mGluR2/3, mGluR4a, and mGluR7 [11]. This observation bolsters the notion that CNS pericytes respond to paracrine cues from other cells, including glutamate-containing neurons and glia. It is not known, however, if CNS pericytes express the molecular machinery necessary to store and release glutamate for intercellular signaling. We therefore determined if CNS pericytes express any of the three known vesicular glutamate transporters (VGLUTs), the proteins required for vesicular storage and exocytotic release of glutamate.
Adult male Sprague–Dawley rats (Harlan, Indianapolis, IN) were group housed on a 12 h light–dark cycle with lights on at 0700, and provided with food and water ad libitum. All studies were performed in accordance with the National Institutes of Health Guide for Care and Use of Laboratory Animals and under the oversight of the Vanderbilt University Animal Care and Use Committee.
Adult male rats were perfused with room temperature 0.1 M phosphate buffer, pH 7.3, followed by ice-cold 4% paraformaldehyde in phosphate buffer. Brains were postfixed in 4% paraformaldehyde, cryoprotected, and stored in a solution of phosphate buffer with 30% sucrose and 30% ethylene glycol at −20 °C. Frozen coronal sections were subsequently cut at 40 µm through the forebrain on a sliding microtome. Using our previously described dual immunofluorescence method [4], we determined if the pericyte marker NG2 proteoglycan [13] was co-expressed with one of the three known VGLUTs. Primary antibodies included rabbit anti-NG2 chondroiton sulfate proteoglycan (1:250; Chemicon, Temecula, CA), rabbit anti-VGLUT1 (1:1000; Mab Technologies, Stone Mountain, GA), mouse anti-VGLUT1 (1:100; Antibodies, Inc., Davis, CA), guinea pig anti-VGLUT2 (1:1000; Chemicon), rat anti-VGLUT2 (1:50; Mab Technologies), guinea pig anti-VGLUT3 (1:10,000; Chemicon), and rabbit anti-von Willebrand factor (1:500; Chemicon). Secondary antibodies included AlexaFluor 488-conjugated donkey anti-rabbit IgG (1:400; Invitrogen, Carlsbad, CA), cyanine-2 (Cy2)-conjugated donkey anti-rat (1:400; Jackson ImmunoResearch Laboratories, West Grove, PA), cyanine-3 (Cy3)-conjugated donkey anti-rabbit and donkey anti-guinea pig IgGs (1:1000; Jackson ImmunoResearch). Images of immunoreactive cells were acquired using an LSM Meta confocal laser scanning microscopy system (Carl Zeiss) equipped with a 63 × 1.4 NA objective and using a 2.5 digital zoom factor.
In order to determine if VGLUTs expressed in CNS pericytes are localized to functional vesicles, we examined the co-localization of the transporters with the anterograde tract tracer biotinylated dextran amine (BDA), which when applied extracellularly is taken up by cells through vesicles that dock with plasma membrane and release contents [1]. Similarly, we determined if VGLUTs in CNS pericytes are present in functional lysosomes by determining co-localization of the transporters with the retrograde tracer FluoroGold (FG), which upon uptake by cells is accumulated into lysosomes [16]. This approach provided for an unambiguous identification of lysosomes and secretory vesicles based on their function. Briefly, rats were deeply anesthetized with isoflurane and a 20 µm (outside tip diameter) glass micropipette was used to iontophoretically deposit a 3% solution of FG (Fluorochrome, Inc., Englewood, CO) in 0.1 M cacodylic acid into the nucleus reuniens of the thalamus, or a 15% solution of the anterograde tracer BDA (10,000 MW; Invitrogen, Carlsbad, CA) in 0.1 M phosphate buffer into the prefrontal cortex. FG and BDA were iontophoresed using pulsed positive 1.5 µA current (7 s on/off) for 10 and 15 min, respectively. Animals were allowed to recover for 12 days before being sacrificed. The presence of BDA was revealed by incubation in Cy3-conjugated streptavidin (1:1000; Jackson ImmunoResearch), while FG was detected using rabbit-anti-FG (1:4000; Chemicon) and subsequent Alexa 488-conjugated donkey anti-rabbit IgG (1:400; Invitrogen).
NG2-proteoglycan-immunoreactive (-ir) pericytes were observed in the leptomeninges (Fig. 1A) as well as associated with brain microvasculature (data not shown). These NG2-positive pericytes had a mean (±S.E.M.) cross-sectional area of 64.23 ± 1.77 µm2. NG2-positive pericytes were uniformly immunoreactive for VGLUT2 (Fig. 1D–F) and for VGLUT3 (Fig. 1G–I). However, we did not observe any pericytes that displayed VGLUT1-ir (Fig. 1A–C).
Fig. 1.
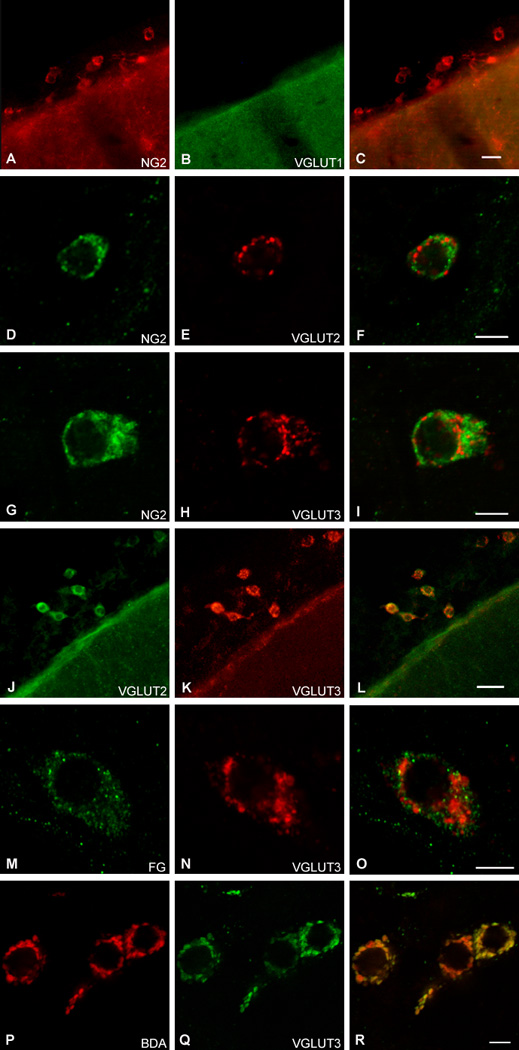
Leptomeningeal CNS pericytes co-express VGLUT2 and VGLUT3 in vesicles. VGLUT1 is not expressed in CNS pericytes (revealed by NG2 proteoglycan (NG2)-ir) (A–C). However, both VGLUT2 (D–F) and VGLUT3 (H–J) are expressed in CNS pericytes, where they appear to be co-localized (J–K). VGLUTs were not seen in lysosomes as revealed using FluoroGold (FG) as a functional marker of these organelles (M–O). Instead,VGLUT3 was localized to secretory vesicles as revealed by the co-localization of the transporter with biotinylated dextran amine (BDA), which is taken up into vesicles from the extracellular environment during exocytosis (P–R). Antibodies: (A–C) mouse anti-VGLUT1; (D–F) guinea pig anti-VGLUT2; (J–L) rat anti-VGLUT2. Scale bars: (C, L) 20 µm; (F, I, O, R) 5 µm. *Blood vessel.
In addition, we observed that single CNS pericyte expressed both VGLUT2- and VGLUT3-ir (Fig. 1J–L). VGLUT2- and VGLUT3-ir were prominent along the cytoplasmic surface of the cell membrane, and could be seen in both the soma and processes. In addition, using VGLUT2 or VGLUT3 antibodies, we observed the orthogonal first order branching of processes that is typical of CNS pericytes [25]. It should be noted that two different VGLUT2 antibodies were used, but these antibodies did not differ in their ability to reveal VGLUT2 protein in CNS pericytes. Two different VGLUT1 antibodies were also used, neither of which were able to detect the presence of VGLUT1 protein in CNS pericytes.
In order to verify the specificity of the VGLUT2 and VGLUT3 antibodies, we performed a control in which the primary antibodies were omitted (Suppl. Fig. 1D–F); no staining was seen. In addition, to examine possible cross-reactivity, we incubated sections in a solution containing the VGLUT3 primary antibody, but not the VGLUT2 antibody, with both secondary antibodies present; only VGLUT3-ir CNS pericytes were observed (Suppl. Fig. 1A–C). Thus, no nonspecific immunofluorescence was observed with either the VGLUT2 or VGLUT3 primary antibodies.
To distinguish CNS pericytes from endothelial cells, double immunofluorescence was performed using guinea pig anti-VGLUT3 and a rabbit antibody directed against the epithelial cell marker von Willebrand Factor (vWF). The vWF antibody revealed capillaries as well as larger vessels. VGLUT-ir CNS pericytes did not co-localize with endothelial cells (Suppl. Fig. 1G–I).
Double staining for VGLUT3-ir and FG revealed that the transporter was not expressed in lysosomes, which accumulate FG (Fig. 1N–O). In contrast, BDA, which is accumulated by vesicles, was co-localized with VGLUT3 (Fig. 1P–R), consistent with a functional vesicular localization of the VGLUTs 2 and 3.
We observed that CNS pericytes express VGLUT2 and VGLUT3 but not vGLUT1. VGLUT2- and VGLUT3-ir profiles mainly appeared as aggregates of spherical bodies approximately 0.5–1 µm in diameter along the cell membrane. This pattern of localization is consistent with ultrastructural studies of pericyte vesicles [9,10] that noted that vesicles appeared aggregated and continuous in their association with the inner surface of the cell membrane.
We also demonstrated that BDA and VGLUT3 were co-localized in CNS pericytes, but that FG and the transporter were not co-localized. Because BDA when applied extracellularly to cultured cells is taken up by synaptic vesicles as they fuse with the plasma membrane to release their contents, it has been used as an indicator of synaptic vesicle release and re-incorporation [1]. Thus, our observation of BDA in CNS pericytes and the co-localization of BDA and VGLUT3 suggest that release and recycling of glutamate may be occurring in these cells. This suggestion is buttressed by the finding that pericytes of human foreskin venules express synaptobrevin, a protein required for vesicular docking and release [6].
Although there are indirect data that suggest that VGLUT3-containing vesicles release glutamate from cortical pyramidal cell dendrites [7,26], direct evidence supporting VGLUT3-mediated exocytotic glutamate release from neurons is lacking [22]. The relative dearth of information concerning VGLUT3 function makes the generalization of the functional significance of CNS pericyte expression of VGLUT3 difficult. However, our observation that BDA is incorporated into VGLUT3-containing vesicles of CNS pericytes suggests that these vesicles are undergoing exocytosis of, presumably, glutamate. In contrast to VGLUT3, far more is known about the functional role of VGLUT2. Takamori and co-workers [23,24] demonstrated that heterologous expression of VGLUT2 is sufficient to confer a glutamatergic phenotype in neurons and endocrine cells. This suggests that CNS pericytes release glutamate, at least through VGLUT2-mediated exocytosis.
Our demonstration that VGLUT2 and VGLUT3 are co-expressed in CNS pericytes differs from previous studies of the adult CNS, where glutamatergic neurons typically express just one VGLUT isoform [8,21]. Although a relatively small number of neurons in the adult brain co-express VGLUT1 and VGLUT2 [15,17,19], we are not aware of any studies of any organ, including the brain, in which co-expression of VGLUT2 and VGLUT3 has been reported. Accordingly, CNS pericytes, which can readily be cultured, appear to be ideal candidates for the study of the functional differences between VGLUT2 and VGLUT3 isoforms in a natively expressing cell type.
Why do CNS pericytes express both VGLUT2 andVGLUT3? The subcellular localization of VGLUT1 and VGLUT2 in neurons is restricted to presynaptic terminals, whereas VGLUT3 is localized to neuronal cell bodies and dendrites as well as presynaptic terminals [21]. Thus, because CNS pericytes have distinct vesicle populations defined by VGLUT isoform expression, these vesicles may be functioning in different subcellular compartments. Although the staining pattern for both VGLUT2 and VGLUT3 appear to be very similar when examined separately, we cannot rule out the existence of two distinct subpopulations of vesicles defined by VGLUT2- or VGLUT3-ir under confocal microscopic examination; future electron microsopic studies of immunogold labeling will be required to conclusively resolve this issue.
To our knowledge, this is the first report demonstrating that CNS pericytes contain the cellular machinery necessary for exocytotic release of glutamate. A logical extension of our observations is that CNS pericytes may communicate with one another through a glutamatergic mechanism. It is also possible that glutamate-dependent pericyte-neuron, pericyte-endothelial cell, or pericyte-astrocyte communication also exists, considering the close apposition of CNS pericytes and neurons, astroctyes, and endothelial cells, all of which express glutamate receptors [2,20]. The dual expression of VGLUT2 and VGLUT3 in CNS pericytes may allow new avenues of research into the functional differences between VGLUT2 and VGLUT3 isoforms. Moreover, these findings clearly add to the growing list of complex and highly varied functions of CNS pericytes, and provide insight into the roles of these cells in health and disease.
Supplementary Material
{kind=link}
Acknowledgments
This work was supported in part by the National Parkinson Foundation Center of Excellence at Vanderbilt University and PO1 NS 44282. We would like to extend our deep gratitude to Jennifer L. Madison for confocal microscopy aid and Michael Bubser for BDA-injected material.
Footnotes
Appendix A. Supplementary data
Supplementary data associated with this article can be found, in the online version, at doi:10.1016/j.neulet.2008.01.082.
References
- 1.Bonzelius F, Zimmermann H. Recycled synaptic vesicles contain vesicle but not plasma membrane marker, newly synthesized achetylcholine, and a sample of extracellular medium. J. Neurochem. 1990;55:1266–1273. doi: 10.1111/j.1471-4159.1990.tb03134.x. [DOI] [PubMed] [Google Scholar]
- 2.Cornell-Bell AH, Finkbeiner SM, Cooper MS, Smith SJ. Glutamate induces calcium waves in cultured astrocytes: long-range glial signaling. Science. 1990;247:470–473. doi: 10.1126/science.1967852. [DOI] [PubMed] [Google Scholar]
- 3.Ehler E, Karlhuber G, Bauer HC, Draeger A. Heterogeneity of smooth muscle-associated proteins in mammalian brain microvasculature. Cell Tissue Res. 1995;279:393–403. doi: 10.1007/BF00318497. [DOI] [PubMed] [Google Scholar]
- 4.Fadel J, Deutch AY. Anatomical substrates of orexin-dopamine interactions: lateral hypothalamic projections to the ventral tegmental area. Neuroscience. 2002;111:379–387. doi: 10.1016/s0306-4522(02)00017-9. [DOI] [PubMed] [Google Scholar]
- 5.Farrell CR, Stewart PA, Farrell CL, Del Maestro RF. Pericytes in human cerebral Mmcrovasculature. Anat. Rec. 1987;218:466–469. doi: 10.1002/ar.1092180416. [DOI] [PubMed] [Google Scholar]
- 6.Feng D, Flaumenhaft R, Bandeira-Melo C, Weller PF, Dvorak AM. Ultrastructural localization of vesicle-associated membrane protein(s) to specialized membrane structures in human pericytes, vascular smooth muscle cells, endothelial cells, neutrophils, and eosinophils. J. Histochem. Cytochem. 2001;49:293–304. doi: 10.1177/002215540104900303. [DOI] [PubMed] [Google Scholar]
- 7.Fremeau RT, Jr, Burman J, Qureshi T, Tran CH, Proctor J, Johnson J, Zhang H, Sulzer D, Copenhagen DR, Storm-Mathisen J, Reimer RJ, Chaudhry FA, Edwards RH. The identification of vesicular glutamate transporter 3 suggests novel modes of signaling by glutamate. PNAS. 2002;99:14488–14493. doi: 10.1073/pnas.222546799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fremeau RT, Voglmaier S, Seal RP, Edwards RH. VGLUTs define subsets of excitatory neurons and suggest novel roles for glutamate. Trends Neurosci. 2004;27:98–103. doi: 10.1016/j.tins.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 9.Frokjaer-Jensen J. The plasmalemmal vesicular system in striated muscle capillaries and in pericytes. Tissue Cell. 1984;16:31–42. doi: 10.1016/0040-8166(84)90016-8. [DOI] [PubMed] [Google Scholar]
- 10.Frokjaer-Jensen J, Wagner RC, Andrews SB, Hagman P, Reese TS. Three-dimensional organization of the plasmalemmal vesicular system in directly frozen capillaries of the rete mirabile in the swim bladder of the Eel. Cell Tissue Res. 1988;254:17–24. doi: 10.1007/BF00220012. [DOI] [PubMed] [Google Scholar]
- 11.Gillard SE, Tzaferis J, Tsui HT, Kingston AE. Expression of metabotropic glutamate receptors in rat meningeal and brain microvasculature and choroid plexus. J. Comp. Neurol. 2003;461:317–332. doi: 10.1002/cne.10671. [DOI] [PubMed] [Google Scholar]
- 12.Herman IM, D’Amore PA. Microvascular pericytes contain muscle and nonmuscle actins. J. Cell Biol. 1985;101:43–52. doi: 10.1083/jcb.101.1.43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hughes S, Chan-Ling T. Characterization of smooth muscle cell and pericyte differentiation in the rat retina in vivo. Invest. Opthalmol. Vis. Sci. 2004;45:2795–2806. doi: 10.1167/iovs.03-1312. [DOI] [PubMed] [Google Scholar]
- 14.Iadecola C, Yang G, Ebner TJ, Chen G. Local and propagated vascular responses evoked by focal synaptic activity in cerebellar cortex. J Neurophysiol. 1997;78:651–659. doi: 10.1152/jn.1997.78.2.651. [DOI] [PubMed] [Google Scholar]
- 15.Li JL, Xiong KH, Dong YL, Fujiyama F, Kaneko T, Mizuno N. Vesicular glutamate transporters, VGluT1 and VGluT2, in the trigeminal ganglion neurons of the rat, with special reference to coexpression. J. Comp. Neurol. 2003;463:212–220. doi: 10.1002/cne.10755. [DOI] [PubMed] [Google Scholar]
- 16.Naumann T, Hartig W, Frotscher M. Retrograde Tracing with Fluoro-Gold: different methods of tracer detection at the ultrastructural level and neurodegenerative changes of back-filled neurons in long-term studies. J. Neurosci. Methods. 2000;103:11–21. doi: 10.1016/s0165-0270(00)00292-2. [DOI] [PubMed] [Google Scholar]
- 17.Oliveira AL, Hydling F, Olsson E, Shi T, Edwards RH, Fujiyama F, Kaneko T, Hökfelt T, Cullheim S, Meister B. Cellular localization of three vesicular glutamate transporter mRNAs and proteins in rat spinal cord and dorsal root ganglia. Synapse. 2003;50:117–129. doi: 10.1002/syn.10249. [DOI] [PubMed] [Google Scholar]
- 18.Peppiatt CM, Howarth C, Mobbs P, Attwell D. Bidirectional control of CNS capillary diameter by pericytes. Nature. 2006;443:700–704. doi: 10.1038/nature05193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Sakata-Haga H, Kanemoto M, Maruyama D, Hoshi K, Moqi K, Narita M, Okado N, Ikeda Y, Nogami H, Fukui Y, Kojima I, Takeda J, Hisano S. Differential localization and colocalization of two neuron-types of sodium-dependent inorganic phosphate cotransporters in rat forebrain. Brain Res. 2001;902:143–155. doi: 10.1016/s0006-8993(01)02290-9. [DOI] [PubMed] [Google Scholar]
- 20.Sharp CD, Hines I, Houghton J, Warren A, Jackson TH, 4th, Jawahar A, Nanda A, Elrod JW, Long A, Chi A, Minagar A, Alexander JS. Glutamate causes a loss in human cerebral endothelial barrier integrity through activation of NMDA receptor. Am. J. Physiol. Heart Circ. Physiol. 2003;285:H2592–H2598. doi: 10.1152/ajpheart.00520.2003. [DOI] [PubMed] [Google Scholar]
- 21.Takamori S. VGLUTs: ‘Exciting’ times for glutamatergic research? Neurosci. Res. 2006;55:343–351. doi: 10.1016/j.neures.2006.04.016. [DOI] [PubMed] [Google Scholar]
- 22.Takamori S, Malherbe P, Broger C, Jahn R. Molecular cloning and functional characterization of human vesicular glutamate transporter 3. EMBO Rep. 2002;3:798–803. doi: 10.1093/embo-reports/kvf159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Takamori S, Rhee JS, Rosenmund C, Jahn R. Identification of a vesicular glutamate transporter that defines a glutamatergic phenotype in neuron. Nature. 2000;407:189–194. doi: 10.1038/35025070. [DOI] [PubMed] [Google Scholar]
- 24.Takamori S, Rhee JS, Rosenmund C, Jahn R. Identification of differentiation-associated brain-specific phosphate transporter as a second vesicular glutamate transporter (VGLUT2) J Neurosci. 2001;21:RC182. doi: 10.1523/JNEUROSCI.21-22-j0002.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Thomas WE. Brain macrophages: on the role of pericytes and perivascular cells. Brain Res. Brain Res. Rev. 1999;31:42–57. doi: 10.1016/s0165-0173(99)00024-7. [DOI] [PubMed] [Google Scholar]
- 26.Zilberter Y. Dendritic release of glutamate suppresses synaptic inhibition of pyramidal neurons in rat neocortex. J. Physiol. 2000;528:489–496. doi: 10.1111/j.1469-7793.2000.00489.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zimmerman K. Der peinere bau der blutcapillaren. A Anat. Entwicklungsgesch. 1923;68:29–109. Cited in: K.K. Hirschi, P.A. D’Amore, Pericytes in the microvasculature, Cardiovasc. Res. 32(1996) 687–698. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.