

Abstract
Disruption of finely coordinated neuropeptide signals in the hypothalamus can result in altered food intake and body weight. We identified neuron-derived neurotrophic factor (NENF) as a novel secreted protein through a large-scale screen aimed at identifying novel secreted hypothalamic proteins that regulate food intake. We observed robust Nenf expression in hypothalamic nuclei known to regulate food intake, and its expression was altered under the diet-induced obese (DIO) condition relative to the fed state. Hypothalamic Nenf mRNA was regulated by brain-derived neurotrophic factor (BDNF) signaling, itself an important regulator of appetite. Delivery of purified recombinant BDNF into the lateral cerebral ventricle decreased hypothalamic Nenf expression, while pharmacological inhibition of trkB signaling increased Nenf mRNA expression. Furthermore, recombinant NENF administered via an intracerebroventricular cannula decreased food intake and body weight and increased hypothalamic Pomc and Mc4r mRNA expression. Importantly, the appetite-suppressing effect of NENF was abrogated in obese mice fed a high-fat diet, demonstrating a diet-dependent modulation of NENF function. We propose the existence of a regulatory circuit involving BDNF, NENF, and melanocortin signaling. Our study validates the power of using an integrated experimental and bioinformatic approach to identify novel CNS-derived proteins with appetite-modulating function and reveals NENF as an important central modulator of food intake.
Keywords: brain-derived neurotrophic factor, melanocortin, whole-body energy balance, neural circuitry, agouti-related protein
energy homeostasis involves finely tuned coordination of signals arising from both peripheral sources and the central nervous system (CNS) to establish a well-balanced homeostatic feedback loop. Hypothalamic neuropeptides and other secreted proteins play a critical role in modulating body weight and food intake. Lesions of the lateral nuclei of the hypothalamus result in anorexia, and while lesions of both the ventromedial (VMH) or paraventricular nuclei (PVN) result in excessive food intake and body weight gain (3, 22). Further, disruption of hypothalamic neuropeptide signals (e.g., brain-derived neurotrophic factor, BDNF; Agouti-related protein, AGRP) in loss-of-function mouse models can lead to metabolic diseases and obesity (48, 63).
Neurotrophic factors act as neuromodulators to maintain growth, survival, and differentiation of neurons (24, 49). Genetic evidence implicates the BDNF signaling pathway in regulating feeding and body weight. A missense mutation in the BDNF receptor, tyrosine kinase receptor B (TRKB), has been identified in severely obese children (19, 59). Additionally, postnatal Bdnf deletion and haploinsufficiency both produce hyperphagia and obesity in mice (17, 46). Further, mice lacking the melanocortin-4 receptor (Mc4r) exhibit excessive weight gain, and Bdnf has been shown to be a downstream effector by which Mc4r signaling modulates energy balance (6, 58). Thus, the Bdnf/trkB signaling pathway contributes toward maintaining energy homeostasis in the CNS, and its disruption results in increased susceptibility to obesity.
To discover novel regulators of feeding and body weight, we conducted a large-scale experimental and bioinformatic screen for hypothalamic factors that may signal downstream of Bdnf/trkB to regulate food intake and body weight. Several characteristics prompted us to select one candidate protein from this screen for in-depth characterization, neuron-derived neurotrophic factor (Nenf or neudesin). Mouse Nenf contains 171 amino acids, is 91% similar to human NENF, is primarily expressed in neurons, but is absent from glia (31). In addition, it is expressed in white adipose tissue (28). Nenf is a secreted heme-binding protein with neurotrophic and neuroprotective activities, and possesses the ability to suppress adipocyte differentiation (28–31). However, a role for NENF in energy balance has not yet been explored.
We show here that BDNF signaling inversely regulates expression of NENF in the CNS and that melanocortin signaling also regulates Nenf mRNA expression in response to food intake. Since increased levels of hypothalamic BDNF suppress food intake and body weight and alter gene expression (13, 55), our findings suggest that NENF may function as a downstream effector of the Bdnf/trkB pathway to influence melanocortin signaling and appetite and that this relationship is disrupted in diet-induced obese (DIO) mice. Indeed, central administration of recombinant NENF significantly increased Pomc and Mc4r mRNA expression. This phenomenon was not observed in DIO mice, suggesting that overfeeding results in potential resistance to NENF signaling. These findings demonstrate the power of the integrated approach used here to identify novel hypothalamic regulators of ingestive behavior.
METHODS
Animals and Housing
C57BL/6J male mice (Jackson Laboratories, Bar Harbor, ME) were used for all studies, and all mice were fed their respective diets from weaning. Wild-type mice were fed a standard rodent chow diet (2018; Harland-Teklad, Indianapolis, IN). Diet-induced obese (DIO) mice were fed a high-fat diet (60% kcal derived from fat content, D12492i; Research Diets, New Brunswick, NJ). TrkBF616A knockin mice (14) were backcrossed to C57BL/6J, and these mice were also fed a standard rodent chow (2018; Harland-Teklad). All mice were individually housed in clear Plexiglas cages, with a solid bottom that contained alpha dry or corn cob bedding. Before the start of a feeding experiment, mice were adapted to wire-mesh floors for 3 days to allow for collection of food spillage throughout the study. The housing environment was regulated to maintain a constant temperature (72°F), humidity (45% ± 5), and a 12:12-h light-dark cycle. All animals had access to water ad libitum, and their respective diets were restricted only during indicated fasting times. All animal experiments were approved by the Animal Care and Use Committee of Johns Hopkins University School of Medicine and were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Experiment 1. Screening Strategy Identified Novel Hypothalamic Secreted Proteins That May Regulate Neural Pathways Associated With Food Intake
In silico analysis.
Publicly available in situ hybridization (ISH) data from adult male C57BL/6J mice were obtained the Allen Brain Atlas (2a) and cross-referenced against in situ hybridization data generated in our own laboratory in the course of a large-scale microarray and ISH-based screen aimed at identifying genes that regulate mouse hypothalamic development (9, 50). Genes that appear to show stronger expression in hypothalamus relative to other brain regions were then cross-checked against the SAGE Genie database (http://cgap.nci.nih.gov/SAGE) to confirm enrichment in hypothalamus relative to both other neuronal and nonneuronal tissues. Sequences of genes that showed prominent and selective expression in adult hypothalamus were then screened for the presence of a signal peptide (http://uswest.ensembl.org/index.html). The chromosomal locations of putative secreted proteins were then checked against the human obesity gene map (41) to ascertain whether they mapped to genomic regions that contain a QTL implicated in the regulation of body composition or food intake. Potential function of the candidate protein was checked against the PubMed database (http://www.ncbi.nlm.nih.gov) to ensure that no previous study implicated the candidate proteins in appetite control.
The following five criteria were applied to further narrow candidates for functional characterization: 1) enriched for expression in hypothalamic nuclei known to regulate body composition, hunger, satiety, or the endocrine axis; 2) maps to the genomic region containing a QTL implicated in the regulation of body composition or food intake; 3) contains a signal peptide sequence in the NH2-terminus for protein secretion, as our focus is on identification of novel secreted neural proteins that regulate food intake; 4) shows altered gene expression under fed, fasted, and DIO conditions; and 5) no previous study implicates the candidate protein in appetite or weight control.
Experiment 2. Changes in Physiological State of Energy Balance Regulate Nenf Gene Expression
Fed control mice (8 wk of age; n = 4) were given ad libitum access to water and standard rodent chow. Fasted mice (8 wk of age; n = 4) were raised on a diet similar to fed controls, but given access only to water for 24 h prior to euthanasia. DIO mice (8 wk of age; n = 4) were provided a high-fat diet. The fed and DIO mice received no food for 2 h to prevent random consumption of a meal prior to euthanasia but were allowed access to water.
In situ hybridization.
To generate an Nenf riboprobe for in situ hybridization (ISH), plasmid encoding the Nenf cDNA was purified using a DNA miniprep kit, according to manufacturer's protocol (K2100–10; Invitrogen, Carlsbad, CA) and used as a template for in vitro transcription. The Nenf riboprobe was synthesized with T3 RNA polymerase (Roche, Indianapolis, IN) for 2 h at 37°C and purified by LiCl precipitation. Brains were collected from ad libitum fed mice, with food removed 2 h before tissue collection. Freshly isolated brain tissues were immediately frozen on dry ice in OCT compound, and then stored at [minus]80 °C. Frozen tissue was cut into sections (25-μm thickness) using a cryostat (Microm, model HM550; Thermo Scientific, Waltham, MA) and mounted onto Superfrost Plus slides (Fisher Scientific, Pittsburgh, PA). ISH was performed as previously described (10, 12).
Experiments 2–6: Quantitative Real-Time PCR
As previously described (12), the anterior commissure and the oculomotor nerve were used as neuroanatomical markers to dissect hypothalami in PBS solution. RNA was extracted and quantified using real-time PCR analysis, as previously described (12, 13). Briefly, RNeasy Midi kits (Qiagen, Valencia, CA) were used to extract RNA. Superscript II reverse transcriptase (Invitrogen) and random primers were used to create cDNA from 1 μg of RNA. SYBR Green PCR master mix (Applied Biosystems, Life Technologies, Carlsbad, CA) was used in quantitative real-time PCR analysis, which generated a threshold cycle (Ct) value of each sample. The ΔCt value was generated by normalizing the data to 18S rRNA, or ΔΔCt generated by normalizing the data to the control treatment. The primers used for quantitative PCR (qPCR) are listed in Table 1.
Table 1.
qPCR primers used to measure hypothalamic gene expression
| Gene | Sense Primer | Antisense Primer |
|---|---|---|
| Nenf | 5′ GAAGGGAGTGGT GTTCGA 3′ | 5′ GTGTCGTGAGTGAGGTCTGC 3′ |
| Pomc | 5′ ATGCCGAGATTCTGCTACAG 3′ | 5′ TGCTGCTGTTCCTGGGGC 3′ |
| Agrp | 5′ ATGCTGACTGCAATGTTGCTG 3′ | 5′ CAGACTTAGACCTGGGA ACTCT 3′ |
| Bdnf | 5′ TCATACTTCGGTTGCATGAAGG 3′ | 5′ AGACCTCT CGAACCTGCCC 3′ |
| Mc4r | 5′ CGGACGGAGGATGCTATGAG 3′ | 5′ CGCCACGATCACTAGAATGTT 3′ |
| 18S rRna | 5′ GCAATTATTCCCCATGAACG 3′ | 5′ GGCC TCACTAAACCATCCAA 3′ |
Experiments 3–5. Recombinant Protein Production
A full-length open reading frame (ORF) encoding human NENF was obtained (Invitrogen, CA), and the region of the ORF lacking the NENF signal peptide sequence was amplified by PCR. Gateway cloning was used to insert NENF cDNA (lacking the signal peptide) into the pDEST17 vector encoding an N-terminal GST followed by a 3C2 protease cleavage site. Optimal bacterial culture conditions for the expression of NENF-GST fusion protein were determined. Bacteria were harvested, lysed, and the soluble recombinant GST fusion protein was purified according to standard protocols. The purified protein was verified via immunoblotting for GST (Invitrogen, according to manufacturer's protocol). The N-terminal GST tag was removed by 3C2 protease, and the purity (>95%) of the resulting untagged NENF was verified by silver staining. Protein concentration was determined by reference to a BSA standard curve and was measured using a Nanodrop 1000 spectrophotometer (Thermo Scientific). Concentration of purified NENF was determined by fitting the value to the known standard using the equation y = mx + b. Full-length recombinant NENF or BDNF with a COOH-terminal FLAG epitope tag (DYKDDDDK) was expressed in mammalian GripTite HEK 293 cells (Invitrogen). Serum-free Opti-MEM media (Invitrogen) containing secreted NENF or BDNF was purified as previously described (40). Purified proteins were dialyzed against 20 mM HEPES buffer (pH 8.0) containing 135 mM NaCl in a 10-kDa cut-off Slide-A-Lyzer dialysis cassette (Pierce, Rockford, IL). After protein concentration was determined using Coomassie Plus protein assay reagent (Thermo Scientific), samples were aliquoted and stored at −80°C.
Experiment 3: Nenf Levels Are Modulated by the BDNF Signaling Pathway
TrkB inhibition.
Bdnf is expressed in the hypothalamus of the adult mouse brain (23), primarily in the VMH with low expression levels in the dorsomedial hypothalamus, in the arcuate nucleus (ARC), and at very low levels in the lateral hypothalamus (51, 55, 58). Therefore, a knockin allele containing a point mutation (F616A) in the trkB gene was used to pharmacologically and temporally inhibit trkB signaling in mice via a highly permeable and selective small-molecule inhibitor, 1NMPP1 (14). The treated group received 1NMPP1 (80 μM) for 14 days via drinking water starting at 2 mo of age. 1NMPP1 was subsequently removed to allow for reactivation of trkB signaling after chemical inhibition. ISH was conducted 4 mo after treatment, as previously described, to measure Nenf mRNA levels in the ARC of the hypothalamus; semiquantitative differences were analyzed using densitometry with ImageJ software (http://rsbweb.nih.gov/ij/), which was normalized to the background. Mice were fasted 2 h prior to collection of brain samples.
Stereotaxic cannulation.
Unilateral cannulas for infusion of proteins were implanted into the lateral cerebral ventricle of C57BL/6J male mice (8 wk of age; n = 10), as described previously (2). One week after surgery, the correct placement of cannula was verified, as previously described, with the following minor modifications: intracerebroventricular injection of 1 nmol of neuropeptide Y (NPY; American Peptide , Sunnyvale, CA), during the light cycle, was used to elicit a ∼100% increase in food intake relative to vehicle (saline), during a 1-h food intake test relative to saline injection (2). We used the dose of the purified BDNF (130 ng) or HEPES buffer (25 mM HEPES pH 8.0 and 135 mM NaCl) as a vehicle, since the protein was suspended in this buffer, food intake and body weight were measured for 24 h after intracerebroventricular injection. The intracerebroventricular injection was administered just prior to the start of the dark cycle. The spillage of food was accounted for and subtracted from total food intake.
Experiment 4: NENF Administration Inhibited Food Intake and Decreased Body Weight
C57BL/6J male mice (8 wk of age) were cannulated, and placement of the cannula was verified as described previously (2). Two weeks after recovering from surgery, baseline food intake was measured at the start of the dark cycle at 1, 2, 4, and 24 h. Saline (0.9%) was used as a vehicle for the recombinant NENF purified from bacterial cells, and HEPES buffer was used as a vehicle control for the recombinant NENF purified in mammalian cells, since the proteins were suspended in these buffers after purification. Central administration of saline or 0.1, 1, 10, and 100 nmol of NENF (made in bacteria) (n = 10), or HEPES or 12 nmol (200 ng) of NENF-FLAG (made in mammalian cells) (n = 11), was given to the mice via intracerebroventricular cannulation. These doses were selected as an initial dose-response curve since other neuropeptides known to modulate food intake have been administered via intracerebroventricular cannulation within this range of doses (39, 61). For studies involving an intracerebroventricular NENF injection, NENF was injected just before onset of the dark cycle, and food intake measurements began after completion of the injections, which approximately coincided with the start of the dark cycle. Before the NENF injection, mice were not food restricted, although mice do tend to decrease food intake naturally during the light cycle. Food intake was measured after subtracting spillage from the total food consumed. Body weights were taken at the time of injection and 24 h after injection.
Kaolin test.
C57BL/6J male mice (4 mo of age, n = 12) were cannulated, and placement of the cannula was verified as previously described (2). An ad libitum amount of kaolin pellets (Research Diets, New Brunswick, NJ), along with ad libitum access to chow, was provided for 1 wk to acclimatize the mice to the novelty of the kaolin. Food intake measurements were recorded at the start of the dark cycle. After acclimatization, an intracerebroventricular injection of vehicle (HEPES buffer) was administered just prior to the start of the dark cycle, and baseline food intake measurements were recorded for both the kaolin and chow diets, with spillage of each diet subtracted, at 2 and 24 h. An intracerebroventricular injection of the recombinant NENF protein (12 nmol) produced in mammalian cells was delivered intracerebroventricularly, and food intake measurements for both the kaolin and chow diets were recorded at 2 and 24 h, with spillage of each deducted.
Experiment 5: NENF Administration Increased Pomc and Mc4r Gene Expression, and High-Fat Diet Abrogated the Appetite-Suppressing Effect of NENF
Mice were cannulated (C57BL/6J; n = 10) to compare the central effects of NENF on food intake in animals consuming a chow vs. high-fat diet. We compared the intakes of mice that were ∼1 yr old and that consumed only a chow diet with the intakes of body weight-matched DIO mice (6 mo old). Weight matching provided a control for potential physiological changes arising from the phenotype induced by body weight. All mice were fed their respective diets since weaning. At the start of the dark cycle, baseline food intake was measured 2 wk after surgery. Recombinant mammalian NENF was injected at 12 nmol, and food intake was measured at 1, 2, 4, and 24 h after injection of the vehicle at the start of the dark cycle, subtracting spillage from the total amount of food consumed. One week after the food intake test, a second injection of the mammalian recombinant NENF was administered intracerebroventricularly, 6 h before the dark cycle. Mice were not allowed access to food after this injection, and hypothalami were collected 3 h later.
Experiment 6: Inhibition of Melanocortin Signaling Increased Food Intake, Which Subsequently Increased Nenf Gene Expression
C57BL/6J male mice were cannulated (n = 14), and placement of the cannula was verified as above. AGRP (61) and SHU-9119 (20) were obtained from Phoenix Pharmaceuticals (St. Joseph, MO). One nanomole of AGRP, an inverse agonist for MC4R, 1 nmol SHU-9119, an antagonist for MC4R, or vehicle (saline) was injected intracerebroventricularly in a 2-μl volume, 8 h before start of the dark cycle. Mice were allowed to consume food for the first 2 h, then food was removed for 2 h. Hypothalami were collected 4 h after initial intracerebroventricular injection, and gene expression was measured using quantitative real-time PCR, as previously described.
Statistical analysis.
All comparisons were made using a one-way ANOVA. A two-way ANOVA for main effects between treatment groups was also performed where appropriate. A Student's t-test was utilized to analyze the semiquantitative values generated from the ISH data and qPCR. All statistical analyses were conducted using Statistica (v.8.0; Tulsa, OK). Values reported are means ± SE, and for all statistical tests, P < 0.05 was considered significant.
RESULTS
Screening Strategy Identified Novel Hypothalamic Secreted Proteins That May Regulate Neural Pathways Associated With Food Intake
We recently conducted a microarray-based screen to identify genes that are dynamically and/or selectively expressed in the developing and adult mouse hypothalamus from embryonic (E) 10 to postnatal day (P) 42, and expression patterns for 1,200 genes were verified by ISH (50). This data set was used as a starting point to identify secreted hypothalamic proteins that may play novel roles in regulating neural pathways involved in control of food intake and metabolism. Further data were obtained by analyzing the publicly available gene expression data generated by the Allen Brain Atlas consortium (38). In total, we identified 250 candidate genes that were prominently and selectively expressed in adult hypothalamic nuclei known to regulate food intake, body weight, and expression of relevant neuropeptide hormones. One candidate, NENF, met all five criteria for candidates of interest and was explored in detail.
Changes in Physiological State of Energy Balance Regulate Nenf Gene Expression
ISH indicated that Nenf mRNA is expressed primarily in the PVN of the hypothalamus, with lower levels of expression in the hypothalamic ARC (Figs. 1 and 2E). We examined whether metabolic state affects expression levels of Nenf in the hypothalamus. Indeed, Nenf, Bdnf, and Pomc mRNA expression in the hypothalamus was suppressed in the DIO, but not the fasted state (Fig. 1, B–D; Nenf F1,8 = 5.63, P = 0.04; Bdnf F1,8 = 10.64, P = 0.013; and Pomc F1,8 = 17.68, P = 0.005). As expected, fasting upregulated mRNA for the orexigenic hypothalamic neuropeptide, Agrp (Fig. 1E: Agrp F1,8=15.49, P = 0.005). Interestingly, the changes in expression patterns of Nenf in response to diet-induced obesity closely resembled changes seen for Bdnf. Since BDNF signaling has been associated with the development of both obesity and anorexia (1, 4, 15, 18, 21, 26, 36, 37, 42–45, 47, 51–53), we determined whether BDNF signaling modulates Nenf mRNA expression.
Fig. 1.

Nenf and neuropeptide gene expression in the hypothalamus. A: in situ hybridization (ISH) for Nenf in the paraventricular nucleus (PVN) and the arcuate nucleus (ARC) of the hypothalamus. B–E: Neuropeptide and neurotrophin gene expression during fed, fasted, and diet-induced obese (DIO) food intake states (n = 4 per group). Nenf (B), Pomc (C), and Bdnf (D) were decreased in both the DIO state relative to the fed state. E: Agrp was increased in the fasted state relative to the fed state. Data are expressed as means ± SE; *P < 0.05.
Fig. 2.

Changes in BDNF signaling alter Nenf gene expression. A–C: BDNF delivered intracerebroventricularly to mice decreased food intake (A), body weight (B), and hypothalamic Nenf mRNA levels (C) after a single injection relative to vehicle (BDNF: n = 5; vehicle: n = 6). D and E: TrkB signaling was inhibited in adult mice for 14 days. The control group received no 1NMPP1 (n = 5 or 6/group). TrkBF616A mice administered 1NMPP1 to inhibit trkB signaling demonstrated increased Nenf gene expression in the ARC at 4 mo of age by both quantitative PCR (D) and ISH (E). Data are expressed as means ± SE; *P < 0.05.
Nenf mRNA Levels Are Modulated by the BDNF Signaling Pathway
To determine whether Nenf mRNA levels are regulated by the BDNF/trkB signaling pathway, we administered recombinant BDNF protein through intracerebroventricular injection just before onset of the dark cycle. Importantly, central administration of BDNF does not cause a conditioned taste aversion (55). An acute increase in BDNF levels in the brain resulted in robust decrease in food intake just 1 h later, and body weight decreased the day after injection, relative to vehicle (Fig. 2, A and B; food intake: 0–1 h F1,9 = 9.07, P = 0.016; 1–2 h F1,9=10.81, P = 0.01; 2–4 h F1,9 = 6.77, P = 0.03; and body weight: F1,9 = 12.33, P = 0.007). No difference in food intake between groups was observed at 24 h postinjection (data not shown). However, Nenf mRNA expression, detected by quantitative real-time PCR, was reduced in the hypothalamus 3 h after BDNF injection (Fig. 2C, F1,9 = 6.40, P = 0.03).
We next sought to test whether transient inhibition of BDNF activity affects hypothalamic Nenf mRNA levels. Mice homozygous for a targeted mutation of trkB, which alters the highly conserved phenylalanine at position 616 within the catalytic kinase domain to alanine (F616A), enabled us to examine the effects of selective and reversible inhibition of trkB signaling on Nenf mRNA levels by using the synthetic compound 1NMPP1 (8, 14). 1NMPP1 is a highly membrane-permeable compound that can be conveniently delivered in drinking water and readily crosses both the placental and blood-brain barriers to reversibly regulate the activity of neuronally expressed mutant trkB (56). Therefore, 1NMPP1, or vehicle solution without 1NMPP1, was administered via drinking water to 2-mo-old mice for 14 days. Hypothalamic Nenf mRNA levels were then characterized in both treatment groups at 4 mo of age. Following inhibition of trkB signaling at 2 mo of age, mice at 4 mo of age displayed a robust increase in ARC Nenf mRNA levels via ISH (Fig. 2, D and E, F1,11 = 29.43, P = 0.0002).
NENF Administration Inhibited Food Intake and Decreased Body Weight
To directly determine whether NENF could affect food intake and body weight, we bacterially produced recombinant NENF or two purified control proteins (CNTF and GST) in mice weighing, on average, 25 g. In addition, recombinant NENF produced in mammalian cells was injected as an additional control for bacterially produced NENF. A single dose of recombinant NENF protein, with the GST tag removed, was injected. Central administration of 0.1, 1, 10, and 100 nmol doses of NENF significantly decreased food intake (Fig. 3, A–E, food intake: 0.1 nmol: 0–1 h, F1,20 = 2.92, P = 0.09; 1–2 h, F1,20 = 7.00, P = 0.013; 2–4 h, F1,20 = 12.56, P = 0.001; 4–22 h, F1,20 = 13.36, P = 0.001; 1 nmol: 0–1 h, F1,20 = 32.67, P = 0.00001; 1–2 h, F1,20 = 26.32, P = 0.00005; 2–4 h, F1,20 = 15.49, P = 0.0008; 4–22 h, F1,20 = 63.53, P = 0.0000001; 10 nmol: 0–1 h, F1,20 = 0.01, P = 0.98; 1–2 h, F1,20 = 11.22, P = 0.002; 2–4 h, F1,20 = 9.22, P = 0.005; 4–22 h, F1,20 = 19.00, P = 0.0001; 100 nmol: 0–1 h, F1,20 = 8.98, P = 0.006; 1–2 h, F1,20 = 16.98, P = 0.0004; 2–4 h, F1,20 = 9.39, P = 0.005; and 4–22 h, F1,20 = 18.48, P = 0.0002) and body weight (Fig. 3F, body weight: 0.1 nmol: F1,20 = 8.75, P = 0.007; 1 nmol: F1,20 = 5.87, P = 0.02; 10 nmol: F1,20 = 9.56, P = 0.004; 100 nmol: F1,20 = 19.50, P = 0.0001). The 100-nmol dose was most efficacious, with decreased food intake at 0–1 h (Fig. 3A); the 1, 10, and 100-nmol doses decreased food intake at 1–2 h, and all doses decreased food intake at 2–4 and 4–22 h (Fig. 3, B–D, respectively). Cumulative food intake over 24 h was significantly reduced for all doses of recombinant NENF (Fig. 3E, cumulative food intake: vehicle: 11.02 kcal ± 0.45, 0.1 nmol: 5.77 kcal ± 1.13, F1,20 = 27.11, P = 0.00002; 1 nmol: 9.86 kcal ± 0.55, F1,20 = 8.77, P = 0.02; 10 nmol: 6.34 kcal ± 1.27, F1,20 = 18.27, P = 0.0002; 100 nmol: 6.05 kcal ± 1.12, F1,20 = 27.91, P = 0.00001). Differences in food intake were not significant between groups at 48 h (vehicle: 13.43 kcal ± 0.6, NENF: 9.67 kcal ± 0.5). In contrast, central administration of 30 nmol GST elicited no changes in food intake or body weight (data not shown). As expected, our positive control, CNTF, decreased food intake following intracerebroventricular injection (data not shown). To ensure that the appetite-suppressing effect of recombinant NENF was not due to bacterial contaminants found in the purified protein, we confirmed our results with recombinant protein produced in a serum-free mammalian expression system. Indeed, 12 nmol (200 ng) of recombinant NENF produced in mammalian HEK 293 cells also decreased food intake in mice when given by intracerebroventricular injection (Fig. 3G, 2–4 h: F1,11 = 6.74, P = 0.02; 4–22 h: F1,11 = 5.79, P = 0.03). In addition, recombinant NENF produced in mammalian cells was able to reduce body weight 24 h after one intracerebroventricular injection (Fig. 3H, F1,11 = 12.74, P = 0.003).
Fig. 3.

Intracerebroventricular recombinant NENF injection decreased food intake and body weight. A–D: 0.1-, 1-, 10-, and 100-nmol doses of bacterially derived NENF were injected into mice (n = 10 per dose), and food intake was measured at 0–1, 1–2, 2–4, and 4–22 h after injection and compared with mice injected with vehicle (saline). Cumulative food intake (E) and body weight (F) were decreased 24 h after a single injection of NENF for all doses. G and H: NENF protein purified from HEK 293 cells was injected as a control for the bacterially expressed purified protein. Protein derived from mammalian cells (12-nmol dose) decreased food intake (G) and body weight (H) relative to vehicle (HEPES buffer)-injected mice, after 24 h, in a similar manner as bacterially expressed NENF. Data are expressed as means ± SE; *P < 0.05.
Fig. 4.

Intracerebroventricular NENF injection in age- and body weight-matched control or DIO mice. A: recombinant NENF injection decreased body weight in control mice [body weight-matched (BWM) were used for control] consuming chow, but not in DIO mice. B and C: BWM mice had decreased food intake after 1 h and 2 h following intracerebroventricular NENF injection relative to vehicle injection (BWM; n = 4–6/group), but DIO mice (n = 8) had no change in food intake relative to saline-injected mice. D–F: neuropeptide gene expression levels were altered after NENF injection. D: Nenf mRNA levels were increased following injection in both BWM and DIO mice. E: relative mRNA levels of Pomc. F: Mc4r gene expression increased in BWM mice, but not in DIO mice, 3 h after NENF injection. *P < 0.05 vs. vehicle. #P = 0.054 vs. vehicle.
Central BDNF administration does not cause a conditioned taste aversion (55); we tested whether NENF administration produced a kaolin response to determine whether the animals had decreased food intake that was attributable to visceral illness and, consequently, an aversion to food. Kaolin and chow food intake were measured at 2 and 24 h after intracerebroventricular injection of either vehicle or the mammalian recombinant NENF. No significant difference in kaolin intake was observed between the two groups at the 2- and 24-h time points (kaolin 2 h: vehicle = 0.02 g ± 0.01, NENF = 0.01 g ± 0.01; 24 h: vehicle = 0. 2 g ± 0.1, NENF = 0.13 g ± 0.06), but chow intake was decreased in the group injected with NENF (chow 2 h: vehicle = 0.36 g ± 0.06, NENF = 0.14 g ± 0.08; chow 24 h: vehicle = 3.89 g ± 0.26, NENF = 1.64 g ± 0.6).
Central NENF Administration Increased Pomc and Mc4r mRNA Expression, and High-Fat Diet Abrogated the Appetite-Suppressing Effect of NENF
In DIO mice, the expression of Pomc is profoundly suppressed as a direct result of obesity-induced physiological changes (25), which alters central melanocortin secretion and signal activation in response to satiety signals, such as leptin (16). To investigate whether NENF might decrease food intake by modulating melanocortin mRNA levels, we injected recombinant NENF intracerebroventricularly in DIO mice (51.5 ± 2.5 g) and body weight-matched (BWM) mice that had consumed only chow (BWM; 46.6 ± 1.6 g). Because DIO mice, consuming a high-fat diet then switched to a chow diet, exhibit restored secretion of melanocortin neuropeptides (16), the BWM group is an appropriate control for the DIO group to investigate the role of melanocortin effects downstream of NENF. Given that NENF is downstream of BDNF, whose expression is modulated by melanocortin signaling, we hypothesized that NENF administration might be much less effective at reducing food intake and body weight in DIO mice. Indeed, we failed to observe any changes in food intake or body weight relative to vehicle-injected DIO mice when the mammalian recombinant NENF was injected (Fig. 4, A–C), with an overall main effect of group (BWM vs. DIO: F1,26 = 5.09, P = 0.02), and time (F1,26 = 6.01, P = 0.02), but no overall interaction between group and time (F1,26 = 1.40, P = 0.27). Cumulative food intake (day 1) was not different between the DIO mice treated with NENF or vehicle (DIO vehicle: 13.73 ± 3.37 kcal; DIO NENF = 15.04 ± 5.98 kcal, F1,11 = 0.01, P = 0.90). However, similar to Fig. 4, B and C, the BWM mice injected with NENF intracerebroventricularly showed suppressed cumulative food intake (day 1), relative to the vehicle group (BWM vehicle: 19.60 ± 0.86 kcal; BWM NENF = 15.29 ± 1.19 kcal, F1,9 = 12.74, P = 0.03). There was no overall difference in the calories consumed between the DIO vehicle and BWM vehicle groups (BWM vehicle: 19.60 ± 0.86 kcal; DIO vehicle = 13.73 ± 3.37 kcal, F1,11 = 1.11, P = 0.31).
Next, we used qPCR to analyze changes in gene expression induced by central administration of recombinant NENF in the hypothalamus of BWM and DIO mice (Fig. 4, D–F). Mice receiving a single intracerebroventricular injection of mammalian recombinant NENF exhibited increased hypothalamic Nenf, Pomc, and Mc4r gene expression in BWM mice. Interestingly, recombinant NENF injected intracerebroventricular in DIO mice increased hypothalamic Nenf mRNA levels but did not alter Pomc or Mc4r mRNA levels (Fig. 4, D–F, respectively, BWM, Nenf: F1,10 = 5.06, P = 0.054; Pomc: F1,10 = 5.36, P = 0.04; Mc4r: F1,10 = 14.16, P = 0.004; and DIO, Nenf: F1,16 = 6.03, P = 0.03; Pomc: F1,16 = 0.11, P = 0.73; Mc4r: F1,16 = 1.06, P = 0.32).
Inhibition of Melanocortin Signaling Increased Food Intake, Which Subsequently Increased Nenf Gene Expression
To determine whether and where NENF may function with the BDNF and melanocortin signaling pathways, we used an experimental paradigm based on studies of Boghossian et al. (11), who demonstrated that AGRP delivery to the amygdala increases Bdnf mRNA levels after allowing animals 2 h of food access following injection. We hypothesized that central delivery of AGRP or SHU-9119 would induce increased food intake, and this, in turn, would induce hypothalamic Nenf gene expression as a signal to reduce food intake, likely in response to satiety factors.
We injected mice with either 0.1 nmol AGRP (n = 6), 1 nmol SHU-9119 (n = 6), or vehicle (saline) control (n = 6). Mice injected with AGRP or SHU-9119 had increased food intake relative to control mice, verifying the accuracy of cannulations and the efficacy of drugs injected (Fig. 5A: control vs. AGRP: F1,12 = 7.00, P = 0.03; control vs. SHU-9119: F1,12 = 6.66, P = 0.03). As expected (11), Bdnf mRNA levels were increased in the hypothalamus after intracerebroventricular injection of either AGRP or SHU-9119 (Fig. 5B: control vs. AGRP: F1,12 = 6.39, P = 0.03; control vs. SHU-9119: F1,12 = 10.94, P = 0.01). Importantly, Nenf mRNA levels were increased as well (Fig. 5C: control vs. AGRP: F1,12 = 8.85, P = 0.02; and control vs. SHU-9119: F1,12 = 6.00, P = 0.03).
Fig. 5.
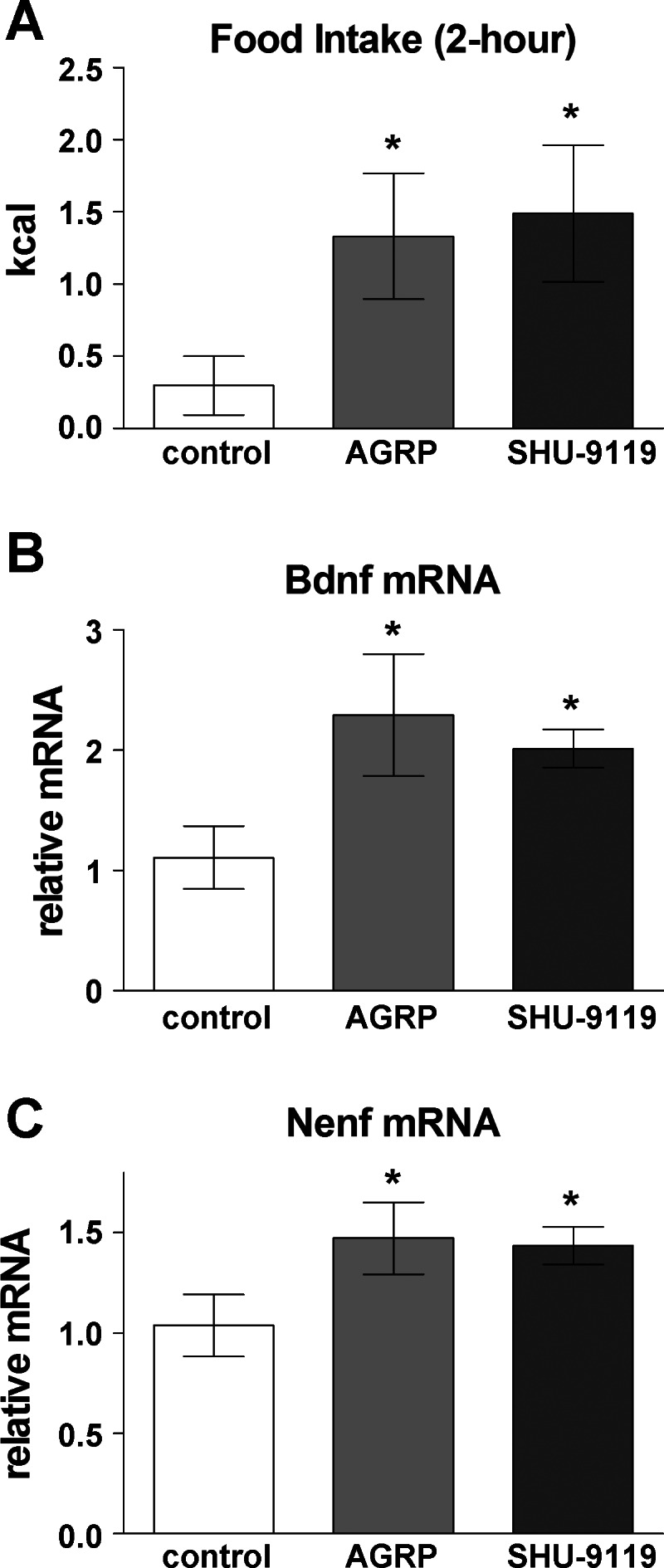
Effect of agouti-related peptide (AGRP) and SHU-9119 on food intake and hypothalamic gene expression. Mice received AGRP (0.1 nmol), SHU-9119 (1 nmol) or saline (control) via intracerebroventricular injection. A: after injection, food was provided and intake was measured from hours 0 to 2; then no food was provided from hours 2 to 4. B and C: hypothalamic Nenf and Bdnf gene expression was increased 4 h after initial injection, following access to food and increased food intake. Values are shown as means ± SE; *P < 0.05 vs. saline.
DISCUSSION
Hypothalamic anorexigenic and orexigenic neurotrophic factors and neuropeptides change expression patterns in response to dietary signals to modulate overall energy balance. In response to a long- or short-term fast, expression of orexigenic neuropeptides increases to drive food intake until satiety is reached. In contrast, expression of anorexigenic neuropeptides increases to suppress food intake and utilizes energy resources stored in adipose tissues, maintaining the body's equilibrium for energy balance. The proportion of orexigenic to anorexigenic expression levels alternates in response to central and peripheral signals known to modulate energy balance (63). We propose that hypothalamic NENF interacts with BDNF and melanocortin signaling to modulate the production of either positive or negative energy balance, which may contribute toward long-term whole-body energy balance.
The mature form of BDNF regulates food intake and body weight primarily by activating trkB receptor signaling. Genetic loss-of-function of Bdnf, either during development or in postnatal animals, results in increased food intake and obesity (17, 27, 46, 51, 58). A similar phenotype is seen following chemical-genetic mediated inhibition of trkB during development (Byerly MS, unpublished data), suggesting that rewiring central neural circuitry regulating feeding during the prenatal or postnatal period can result in permanent changes in the adult. Previous studies have reported that high-fat/high-sugar diets decrease hypothalamic expression of both Bdnf and trkB (34, 35, 57, 60), and we have confirmed that Bdnf mRNA levels were also suppressed in DIO mice. In a state of positive energy balance, short-term changes (14 days) in trkB inhibition resulted in upregulation of Nenf mRNA levels up to 4 mo later. In a state of intracerebroventricularly induced negative energy balance, created by central administration of BDNF to reduce food intake, rapid decreases in hypothalamic Nenf mRNA levels were observed. However, changes in Bdnf, Nenf, and Pomc mRNA levels were not observed after a 24-h fast, suggesting that the duration of the fast may not have been the optimal duration to observe gene expression changes (e.g., the fast was too long or not long enough).
Nenf is expressed in nuclei of the hypothalamus known to be involved in energy regulation, and changes in diet (DIO mice) altered Nenf gene expression in the same direction as Bdnf gene expression; thus, we predicted that NENF would inhibit food intake. Indeed, central delivery of recombinant NENF decreased food intake and body weight, suggesting that it is a novel anorexigenic neurotrophic factor. The dose of NENF delivered intracerebroventricularly altered the duration of time until decreased food intake was observed, with the highest dose (100 nmol) suppressing food intake by 1 h and the lowest dose (0.1 nmol) by 4 h, relative to vehicle-injected mice. However, the lower limit of physiological detection for NENF did not fall into the range of doses delivered, and remains to be determined.
Food intake is controlled by hypothalamic melanocortins acting on the Mc4r, with Agrp acting as an orexigenic inverse agonist of α-melanocyte-stimulating-hormone (α-msh), a peptide product of proopiomelanocortin (Pomc), which acts as an anorexigenic receptor agonist. We have shown that NENF is also a catabolic neurotrophic factor expressed in the PVN and ARC nuclei, with expression only observed in the ARC after long-term trkB inhibition. Nenf mRNA is modulated by melanocortin signals, a pathway also known to modulate Bdnf expression (6, 58). However, under conditions in which hypothalamic expression of Pomc is profoundly suppressed, such as is seen in DIO mice, central administration of NENF failed to elicit changes in food intake, and did not increase Pomc and Mc4r mRNA levels. The lack of effect in the DIO mice does not appear to be secondary to body weight, since chow-fed mice that had reached comparable body weight still responded. DIO mice have been shown to have only one-third of the activation of the Mc4r relative to control, resulting in decreased central melanocortin activation (16). Therefore, these data support the hypothesis that NENF acts as an anorexigenic neurotrophic factor that contributes to energy balance, and DIO mice may be resistant to NENF signals as a result of altered central melanocortin signaling.
The DIO diet is high in fat and dense in energy, which may result in it being a more palatable diet given the increased fat content relative to a standard chow diet. Therefore, the high-fat diet fed to DIO mice may have altered reward cues associated with food intake, relative to BWM mice fed a standard chow diet. Environmental factors, such as energy density of the diet, palatability/taste, or large quantities of available food, provide reward cues to reinforce and drive food intake behavior (7, 33, 62). These reward cues have been shown to weaken feedback signals, such as leptin, which serve to decrease food intake and modulate whole-body energy balance (7, 62). Similar quantities of food were provided to DIO and BWM mice daily. Therefore, the quantities of available food most likely did not influence the reward cues between the two groups. However, the energy density of the diet, as well as palatability and/or taste differences between the diets, may have influenced the amount consumed between the two groups (DIO vs. BWM), overriding the signals triggered by centrally administered recombinant NENF. Thus, it is possible that the DIO mice have alterations in reward-driven food intake behavior (62). It remains to be determined whether NENF is less effective in altering energy balance when reward-driven food intake is reinforced by components of the high-fat diet fed to the DIO mice, relative to the standard chow fed to the BWM mice.
The percent reduction of food intake for the mammalian recombinant NENF treatment group, relative to saline, resembled the percent reduction observed for the 1-nmol dose of the bacterially produced recombinant NENF at 2–4 and 4–22 h. The dose injected for the mammalian recombinant NENF was 12 nmol, suggesting that the mammalian recombinant protein was 10-fold less effective than the bacterially produced recombinant NENF protein. This difference between the mammalian and bacterially derived NENF may be attributable to residual contaminants, such as LPS or muramyl dipeptide (MDP), present in bacterially produced recombinant protein, even after LPS removal by a commercially available kit. LPS and MDP have been shown to reduce food intake via intraperitoneal injection (32) and intracerebroventricular injection (54). In addition, the recombinant NENF produced in mammalian cells is an ideal control for the recombinant NENF produced in bacteria, because the mammalian expression system allows for native protein modifications that would not be found in bacteria, such as glycosylation. Our findings suggest that the purified mammalian recombinant NENF is, indeed, effective in regulating food intake when produced in a system that does not contain LPS and possesses endogenous protein modifications found in the mouse. Given that we used the mammalian protein in DIO and BWM mice to show alterations in hypothalamic neuropeptide gene expression, we confirm that NENF is an anorexigenic neurotrophic factor capable of inhibiting food intake and regulating expression of genes.
Perspectives and Significance
Many genes have been identified to be associated with obesity (41); however, the function of proteins produced by these genes has not yet been characterized in regard to food intake or whole-body energy balance. An integrated approach, combining experimental and bioinformatic techniques, led to the identification of candidate hypothalamic neuropeptides with novel appetite-modulating function. One such candidate, NENF, when centrally administered to mice, decreased food intake and body weight, thus validating the general approach used to identify functionally relevant secretory proteins that regulate neuronal pathways controlling ingestive behavior. Alterations to BDNF/trkB or melanocortin signaling led to a compensatory change in hypothalamic Nenf mRNA levels, suggesting that Nenf gene expression is responsive to both trkB and melanocortin signaling to regulate food intake and energy balance. Our results indicate that DIO mice may be resistant to NENF signaling, since DIO mice have weakened central melanocortin activation (16) and do not show upregulated melanocortin signaling following intracerebroventricular administration of recombinant NENF. We propose that this regulatory circuit contributes to the maintenance of normal energy homeostasis and is likely to be disrupted in obesity. We demonstrated that hypothalamic signaling pathways in the mouse respond to the human NENF protein. Physiological processes activated by NENF signaling may be evolutionarily conserved since the human and mouse proteins have 91% similarity (31), which suggests that NENF may be a novel candidate biomarker for disease susceptibility or development of novel drug therapeutics in humans.
GRANTS
This work was supported by a W.M. Keck Distinguished Young Scholar Award in Medical Research to S.B. and by a National Institute of Diabetes and Digestive and Kidney Diseases training fellowship to M.S.B. (T32DK007751). G.W.W. is supported by NIH grant DK084171.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
Author contributions: M.S.B., S.A., T.H.M., G.W.W., and S.B. conception and design of research; M.S.B., R.D.S., N.N.S., P.S.M., K.K., and G.W.W. performed experiments; M.S.B. analyzed data; M.S.B. and S.B. interpreted results of experiments; M.S.B. prepared figures; M.S.B. drafted manuscript; M.S.B., N.N.S., T.H.M., G.W.W., and S.B. edited and revised manuscript; M.S.B., S.A., T.H.M., G.W.W., and S.B. approved final version of manuscript.
ACKNOWLEDGMENTS
We thank Dr. David Ginty for kindly providing the trkB knock-in mice (Department of Neuroscience, Johns Hopkins University School of Medicine).
REFERENCES
- 1. Adan RA, Hillebrand JJ, De Rijke C, Nijenhuis W, Vink T, Garner KM, Kas MJ. Melanocortin system and eating disorders. Ann NY Acad Sci 994: 267–274, 2003 [DOI] [PubMed] [Google Scholar]
- 2. Aja S, Landree LE, Kleman AM, Medghalchi SM, Vadlamudi A, McFadden JM, Aplasca A, Hyun J, Plummer E, Daniels K, Kemm M, Townsend CA, Thupari JN, Kuhajda FP, Moran TH, Ronnett GV. Pharmacological stimulation of brain carnitine palmitoyl-transferase-1 decreases food intake and body weight. Am J Physiol Regul Integr Comp Physiol 294: R352–R361, 2008 [DOI] [PubMed] [Google Scholar]
- 2a. Allen Institute for Brain Science Allen Mouse Brain Atlas. Seattle, WA: Allen Institute for Brain Science; http://mouse.brain-map.org, 2009. [Google Scholar]
- 3. Anand BK, Brobeck JR. Hypothalamic control of food intake in rats and cats. Yale J Biol Med 24: 124–140, 1951 [PMC free article] [PubMed] [Google Scholar]
- 4. Argyropoulos G, Rankinen T, Neufeld DR, Rice T, Province MA, Leon AS, Skinner JS, Wilmore JH, Rao DC, Bouchard C. A polymorphism in the human agouti-related protein is associated with late-onset obesity. J Clin Endocrinol Metab 87: 4198–4202, 2002 [DOI] [PubMed] [Google Scholar]
- 6. Bariohay B, Roux J, Tardivel C, Trouslard J, Jean A, Lebrun B. BDNF/TrkB signaling is a downstream effector of the brainstem melanocortin system in food intake control. Endocrinology 150: 2646–2653, 2009 [DOI] [PubMed] [Google Scholar]
- 7. Berthoud HR. Metabolic and hedonic drives in the neural control of appetite: who is the boss? Curr Opin Neurobiol 21: 888–896, 2011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Bishop AC, Ubersax JA, Petsch DT, Matheos DP, Gray NS, Blethrow J, Shimizu E, Tsien JZ, Schultz PG, Rose MD, Wood JL, Morgan DO, Shokat KM. A chemical switch for inhibitor-sensitive alleles of any protein kinase. Nature 407: 395–401, 2000 [DOI] [PubMed] [Google Scholar]
- 9. Blackshaw S, Scholpp S, Placzek M, Ingraham H, Simerly R, Shimogori T. Molecular pathways controlling development of thalamus and hypothalamus: from neural specification to circuit formation. J Neurosci 30: 14925–14930, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Blackshaw S, Snyder SH. Developmental expression pattern of phototransduction components in mammalian pineal implies a light-sensing function. J Neurosci 17: 8074–8082, 1997 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Boghossian S, Park M, York DA. Melanocortin activity in the amygdala controls appetite for dietary fat. Am J Physiol Regul Integr Comp Physiol 298: R385–R393, 2010 [DOI] [PubMed] [Google Scholar]
- 12. Byerly MS, Al Salayta M, Swanson RD, Kwon K, Peterson JM, Wei Z, Aja S, Moran TH, Blackshaw S, Wong GW. Estrogen-related receptor beta deletion modulates whole-body energy balance via estrogen-related receptor gamma and attenuates neuropeptide Y gene expression. Eur J Neurosci 37: 1033–1047, 2013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Byerly MS, Simon J, Lebihan-Duval E, Duclos MJ, Cogburn LA, Porter TE. Effects of BDNF, T3, and corticosterone on expression of the hypothalamic obesity gene network in vivo and in vitro. Am J Physiol Regul Integr Comp Physiol 296: R1180–R1189, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Chen X, Ye H, Kuruvilla R, Ramanan N, Scangos KW, Zhang C, Johnson NM, England PM, Shokat KM, Ginty DD. A chemical-genetic approach to studying neurotrophin signaling. Neuron 46: 13–21, 2005 [DOI] [PubMed] [Google Scholar]
- 15. de Rijke CE, Jackson PJ, Garner KM, van Rozen RJ, Douglas NR, Kas MJ, Millhauser GL, Adan RA. Functional analysis of the Ala67Thr polymorphism in agouti related protein associated with anorexia nervosa and leanness. Biochem Pharmacol 70: 308–316, 2005 [DOI] [PubMed] [Google Scholar]
- 16. Enriori PJ, Evans AE, Sinnayah P, Jobst EE, Tonelli-Lemos L, Billes SK, Glavas MM, Grayson BE, Perello M, Nillni EA, Grove KL, Cowley MA. Diet-induced obesity causes severe but reversible leptin resistance in arcuate melanocortin neurons. Cell Metab 5: 181–194, 2007 [DOI] [PubMed] [Google Scholar]
- 17. Fox EA, Byerly MS. A mechanism underlying mature-onset obesity: evidence from the hyperphagic phenotype of brain-derived neurotrophic factor mutants. Am J Physiol Regul Integr Comp Physiol 286: R994–R1004, 2004 [DOI] [PubMed] [Google Scholar]
- 18. Friedel S, Horro FF, Wermter AK, Geller F, Dempfle A, Reichwald K, Smidt J, Bronner G, Konrad K, Herpertz-Dahlmann B, Warnke A, Hemminger U, Linder M, Kiefl H, Goldschmidt HP, Siegfried W, Remschmidt H, Hinney A, Hebebrand J. Mutation screen of the brain derived neurotrophic factor gene (BDNF): identification of several genetic variants and association studies in patients with obesity, eating disorders, and attention-deficit/hyperactivity disorder. Am J Med Genet 132: 96–99, 2005 [DOI] [PubMed] [Google Scholar]
- 19. Gray J, Yeo G, Hung C, Keogh J, Clayton P, Banerjee K, McAulay A, O'Rahilly S, Farooqi IS. Functional characterization of human NTRK2 mutations identified in patients with severe early-onset obesity. Int J Obes (Lond) 31: 359–364, 2007 [DOI] [PubMed] [Google Scholar]
- 20. Grill HJ, Ginsberg AB, Seeley RJ, Kaplan JM. Brainstem application of melanocortin receptor ligands produces long-lasting effects on feeding and body weight. J Neurosci 18: 10128–10135, 1998 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Hashimoto K, Koizumi H, Nakazato M, Shimizu E, Iyo M. Role of brain-derived neurotrophic factor in eating disorders: recent findings and its pathophysiological implications. Prog Neuropsychopharmacol Biol Psychiatry 29: 499–504, 2005 [DOI] [PubMed] [Google Scholar]
- 22. Hetherington A, Ranson S. Hypothalamic lesions and adiposity in the rat. Anatom Record 78: 149–182, 1940 [Google Scholar]
- 23. Hofer M, Pagliusi SR, Hohn A, Leibrock J, Barde YA. Regional distribution of brain-derived neurotrophic factor mRNA in the adult mouse brain. EMBO J 9: 2459–2464, 1990 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Huang EJ, Reichardt LF. Trk receptors: roles in neuronal signal transduction. Annu Rev Biochem 72: 609–642, 2003 [DOI] [PubMed] [Google Scholar]
- 25. Huang XF, Han M, South T, Storlien L. Altered levels of POMC, AgRP and MC4-R mRNA expression in the hypothalamus and other parts of the limbic system of mice prone or resistant to chronic high-energy diet-induced obesity. Brain Res 992: 9–19, 2003 [DOI] [PubMed] [Google Scholar]
- 26. Kalnina I, Kapa I, Pirags V, Ignatovica V, Schioth HB, Klovins J. Association between a rare SNP in the second intron of human Agouti related protein gene and increased BMI. BMC Med Genet 10: 63, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Kernie SG, Liebl DJ, Parada LF. BDNF regulates eating behavior and locomotor activity in mice. EMBO J 19: 1290–1300, 2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Kimura I, Konishi M, Asaki T, Furukawa N, Ukai K, Mori M, Hirasawa A, Tsujimoto G, Ohta M, Itoh N, Fujimoto M. Neudesin, an extracellular heme-binding protein, suppresses adipogenesis in 3T3–L1 cells via the MAPK cascade. Biochem Biophys Res Commun 381: 75–80, 2009 [DOI] [PubMed] [Google Scholar]
- 29. Kimura I, Konishi M, Miyake A, Fujimoto M, Itoh N. Neudesin, a secreted factor, promotes neural cell proliferation and neuronal differentiation in mouse neural precursor cells. J Neurosci Res 83: 1415–1424, 2006 [DOI] [PubMed] [Google Scholar]
- 30. Kimura I, Nakayama Y, Yamauchi H, Konishi M, Miyake A, Mori M, Ohta M, Itoh N, Fujimoto M. Neurotrophic activity of neudesin, a novel extracellular heme-binding protein, is dependent on the binding of heme to its cytochrome b5-like heme/steroid-binding domain. J Biol Chem 283: 4323–4331, 2008 [DOI] [PubMed] [Google Scholar]
- 31. Kimura I, Yoshioka M, Konishi M, Miyake A, Itoh N. Neudesin, a novel secreted protein with a unique primary structure and neurotrophic activity. J Neurosci Res 79: 287–294, 2005 [DOI] [PubMed] [Google Scholar]
- 32. Langhans W, Balkowski G, Savoldelli D. Differential feeding responses to bacterial lipopolysaccharide and muramyl dipeptide. Am J Physiol Regul Integr Comp Physiol 261: R659–R664, 1991 [DOI] [PubMed] [Google Scholar]
- 33. Lenard NR, Berthoud HR. Central and peripheral regulation of food intake and physical activity: pathways and genes. Obesity (Silver Spring) 16 Suppl 3: S11–S22, 2008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Molteni R, Barnard RJ, Ying Z, Roberts CK, Gomez-Pinilla F. A high-fat, refined sugar diet reduces hippocampal brain-derived neurotrophic factor, neuronal plasticity, and learning. Neuroscience 112: 803–814, 2002 [DOI] [PubMed] [Google Scholar]
- 35. Molteni R, Wu A, Vaynman S, Ying Z, Barnard RJ, Gomez-Pinilla F. Exercise reverses the harmful effects of consumption of a high-fat diet on synaptic and behavioral plasticity associated to the action of brain-derived neurotrophic factor. Neuroscience 123: 429–440, 2004 [DOI] [PubMed] [Google Scholar]
- 36. Monteleone P, Maj M. Genetic susceptibility to eating disorders: associated polymorphisms and pharmacogenetic suggestions. Pharmacogenomics 9: 1487–1520, 2008 [DOI] [PubMed] [Google Scholar]
- 37. Nakazato M, Hashimoto K, Shimizu E, Kumakiri C, Koizumi H, Okamura N, Mitsumori M, Komatsu N, Iyo M. Decreased levels of serum brain-derived neurotrophic factor in female patients with eating disorders. Biol Psychiatry 54: 485–490, 2003 [DOI] [PubMed] [Google Scholar]
- 38. Ng L, Bernard A, Lau C, Overly CC, Dong HW, Kuan C, Pathak S, Sunkin SM, Dang C, Bohland JW, Bokil H, Mitra PP, Puelles L, Hohmann J, Anderson DJ, Lein ES, Jones AR, Hawrylycz M. An anatomic gene expression atlas of the adult mouse brain. Nat Neurosci 12: 356–362, 2009 [DOI] [PubMed] [Google Scholar]
- 39. Olszewski PK, Bomberg EM, Grace MK, Levine AS. Alpha-melanocyte stimulating hormone and ghrelin: central interaction in feeding control. Peptides 28: 2084–2089, 2007 [DOI] [PubMed] [Google Scholar]
- 40. Peterson JM, Wei Z, Wong GW. C1q/TNF-related protein-3 (CTRP3), a novel adipokine that regulates hepatic glucose output. J Biol Chem 285: 39691–39701, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Rankinen T, Zuberi A, Chagnon YC, Weisnagel SJ, Argyropoulos G, Walts B, Perusse L, Bouchard C. The human obesity gene map: the 2005 update. Obesity (Silver Spring) 14: 529–644, 2006 [DOI] [PubMed] [Google Scholar]
- 42. Ribases M, Gratacos M, Armengol L, de Cid R, Badia A, Jimenez L, Solano R, Vallejo J, Fernandez F, Estivill X. Met66 in the brain-derived neurotrophic factor (BDNF) precursor is associated with anorexia nervosa restrictive type. Mol Psychiatry 8: 745–751, 2003 [DOI] [PubMed] [Google Scholar]
- 43. Ribases M, Gratacos M, Badia A, Jimenez L, Solano R, Vallejo J, Fernandez-Aranda F, Estivill X. Contribution of NTRK2 to the genetic susceptibility to anorexia nervosa, harm avoidance and minimum body mass index. Mol Psychiatry 10: 851–860, 2005 [DOI] [PubMed] [Google Scholar]
- 44. Ribases M, Gratacos M, Fernandez-Aranda F, Bellodi L, Boni C, Anderluh M, Cavallini MC, Cellini E, Di Bella D, Erzegovesi S, Foulon C, Gabrovsek M, Gorwood P, Hebebrand J, Hinney A, Holliday J, Hu X, Karwautz A, Kipman A, Komel R, Nacmias B, Remschmidt H, Ricca V, Sorbi S, Wagner G, Treasure J, Collier DA, Estivill X. Association of BDNF with anorexia, bulimia and age of onset of weight loss in six European populations. Hum Mol Genet 13: 1205–1212, 2004 [DOI] [PubMed] [Google Scholar]
- 45. Ribases M, Gratacos M, Fernandez-Aranda F, Bellodi L, Boni C, Anderluh M, Cristina Cavallini M, Cellini E, Di Bella D, Erzegovesi S, Foulon C, Gabrovsek M, Gorwood P, Hebebrand J, Hinney A, Holliday J, Hu X, Karwautz A, Kipman A, Komel R, Nacmias B, Remschmidt H, Ricca V, Sorbi S, Tomori M, Wagner G, Treasure J, Collier DA, Estivill X. Association of BDNF with restricting anorexia nervosa and minimum body mass index: a family-based association study of eight European populations. Eur J Hum Genet 13: 428–434, 2005 [DOI] [PubMed] [Google Scholar]
- 46. Rios M, Fan G, Fekete C, Kelly J, Bates B, Kuehn R, Lechan RM, Jaenisch R. Conditional deletion of brain-derived neurotrophic factor in the postnatal brain leads to obesity and hyperactivity. Mol Endocrinol 15: 1748–1757, 2001 [DOI] [PubMed] [Google Scholar]
- 47. Saito S, Watanabe K, Hashimoto E, Saito T. Low serum BDNF and food intake regulation: A possible new explanation of the pathophysiology of eating disorders. Prog Neuropsychopharmacol Biol Psychiatry 33: 312–316, 2009 [DOI] [PubMed] [Google Scholar]
- 48. Schwartz MW, Woods SC, Porte D, Jr, Seeley RJ, Baskin DG. Central nervous system control of food intake. Nature 404: 661–671, 2000 [DOI] [PubMed] [Google Scholar]
- 49. Segal RA. Selectivity in neurotrophin signaling: theme and variations. Annu Rev Neurosci 26: 299–330, 2003 [DOI] [PubMed] [Google Scholar]
- 50. Shimogori T, Lee DA, Miranda-Angulo A, Yang Y, Wang H, Jiang L, Yoshida AC, Kataoka A, Mashiko H, Avetisyan M, Qi L, Qian J, Blackshaw S. A genomic atlas of mouse hypothalamic development. Nat Neurosci 13: 767–775, 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Unger TJ, Calderon GA, Bradley LC, Sena-Esteves M, Rios M. Selective deletion of Bdnf in the ventromedial and dorsomedial hypothalamus of adult mice results in hyperphagic behavior and obesity. J Neurosci 27: 14265–14274, 2007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. van Rossum CT, Pijl H, Adan RA, Hoebee B, Seidell JC. Polymorphisms in the NPY and AGRP genes and body fatness in Dutch adults. Int J Obes (Lond) 30: 1522–1528, 2006 [DOI] [PubMed] [Google Scholar]
- 53. Vink T, Hinney A, van Elburg AA, van Goozen SH, Sandkuijl LA, Sinke RJ, Herpertz-Dahlmann BM, Hebebrand J, Remschmidt H, van Engeland H, Adan RA. Association between an agouti-related protein gene polymorphism and anorexia nervosa. Mol Psychiatry 6: 325–328, 2001 [DOI] [PubMed] [Google Scholar]
- 54. von Meyenburg C, Langhans W, Hrupka BJ. Evidence for a role of the 5-HT2C receptor in central lipopolysaccharide-, interleukin-1 beta-, and leptin-induced anorexia. Pharmacol Biochem Behav 74: 1025–1031, 2003 [DOI] [PubMed] [Google Scholar]
- 55. Wang C, Bomberg E, Levine A, Billington C, Kotz CM. Brain-derived neurotrophic factor in the ventromedial nucleus of the hypothalamus reduces energy intake. Am J Physiol Regul Integr Comp Physiol 293: R1037–R1045, 2007 [DOI] [PubMed] [Google Scholar]
- 56. Wang H, Shimizu E, Tang YP, Cho M, Kyin M, Zuo W, Robinson DA, Alaimo PJ, Zhang C, Morimoto H, Zhuo M, Feng R, Shokat KM, Tsien JZ. Inducible protein knockout reveals temporal requirement of CaMKII reactivation for memory consolidation in the brain. Proc Natl Acad Sci USA 100: 4287–4292, 2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Wu A, Ying Z, Gomez-Pinilla F. The interplay between oxidative stress and brain-derived neurotrophic factor modulates the outcome of a saturated fat diet on synaptic plasticity and cognition. Eur J Neurosci 19: 1699–1707, 2004 [DOI] [PubMed] [Google Scholar]
- 58. Xu B, Goulding EH, Zang K, Cepoi D, Cone RD, Jones KR, Tecott LH, Reichardt LF. Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nat Neurosci 6: 736–742, 2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Yeo GS, Connie Hung CC, Rochford J, Keogh J, Gray J, Sivaramakrishnan S, O'Rahilly S, Farooqi IS. A de novo mutation affecting human TrkB associated with severe obesity and developmental delay. Nat Neurosci 7: 1187–1189, 2004 [DOI] [PubMed] [Google Scholar]
- 60. Zeeni N, Chaumontet C, Moyse E, Fromentin G, Tardivel C, Tome D, Jean A, Darcel N. A positive change in energy balance modulates TrkB expression in the hypothalamus and nodose ganglia of rats. Brain Res 1289: 49–55, 2009 [DOI] [PubMed] [Google Scholar]
- 61. Zheng H, Corkern MM, Crousillac SM, Patterson LM, Phifer CB, Berthoud HR. Neurochemical phenotype of hypothalamic neurons showing Fos expression 23 h after intracranial AgRP. Am J Physiol Regul Integr Comp Physiol 282: R1773–R1781, 2002 [DOI] [PubMed] [Google Scholar]
- 62. Zheng H, Lenard NR, Shin AC, Berthoud HR. Appetite control and energy balance regulation in the modern world: reward-driven brain overrides repletion signals. Int J Obes (Lond) 33 Suppl 2: S8–S13, 2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Zigman JM, Elmquist JK. Minireview: From anorexia to obesity—the yin and yang of body weight control. Endocrinology 144: 3749–3756, 2003 [DOI] [PubMed] [Google Scholar]