

Abstract
Recent evidence suggests that affect acts as modulator of cognitive processes and in particular that induced mood has an effect on the way semantic memory is used on-line. We used event-related potentials (ERPs) to examine affective modulation of semantic information processing under three different moods: neutral, positive and negative. Fifteen subjects read 324 pairs of sentences, after mood induction procedure with 30 pictures of neutral, 30 pictures of positive and 30 pictures of neutral valence: 108 sentences were read in each mood induction condition. Sentences ended with three word types: expected words, within-category violations, and between-category violations. N400 amplitude was measured to the three word types under each mood induction condition. Under neutral mood, a congruency (more negative N400 amplitude for unexpected relative to expected endings) and a category effect (more negative N400 amplitude for between- than to within-category violations) were observed. Also, results showed differences in N400 amplitude for both within- and between-category violations as a function of mood: while positive mood tended to facilitate the integration of unexpected but related items, negative mood made their integration as difficult as unexpected and unrelated items. These findings suggest the differential impact of mood on access to long-term semantic memory during sentence comprehension.
Keywords: mood, language, semantic memory, ERP, N400
INTRODUCTION
In spite of the fact that the way mood modulates cognition has intrigued researchers for decades, there is still a dearth of studies on brain correlates of these processes. Several studies have shown that affective stimuli are processed differently from neutral stimuli, both in visual and auditory modalities (e.g. Bradley and Lang, 2000; Cuthbert et al., 2000; Schupp et al., 2003; Paulmann and Kotz, 2008). In addition, a number of studies have shown that affect acts as a modulator of cognitive processes. However, the neurobiological underpinnings of emotion–cognition interactions remain unclear.
Studies in normal populations suggest that mood has an impact on several cognitive processes, including language (Bolte et al., 2003), memory (Gray, 2001; Hurlemann et al., 2005; Storbeck and Clore, 2005; Bauml and Kuhbandner, 2007), attention (Wang et al., 2006) and executive functions (Dreisbach and Goschke, 2004; Simon-Thomas and Knight, 2005; Dreisbach, 2006; Larson et al., 2006; Wiswede et al., 2009). For example, positive mood states are associated with increased flexibility of thinking and creative problem solving (e.g. Isen and Daubman, 1984; Isen et al., 1987, 1992; Fiedler, 2001; Bolte et al., 2003; Dreisbach and Goschke, 2004; Dreisbach, 2006). These findings suggest that positive mood is associated with a heuristic, global or holistic (‘top-down’) processing style (Bohner et al., 1994; Park and Banaji, 2000; Bolte et al., 2003; Fredrickson and Branigan, 2005; Vissers et al., 2010).
Even though the effects of negative mood are less consistent (Mitchell and Phillips, 2007) and more difficult to predict (Ashby et al., 1999), some studies indicate that it is associated with narrowed, local, detailed-oriented (‘bottom-up’) information processing (Fiedler, 1988, 2001). Together, the existing studies investigating the effects of mood on cognitive processes suggest that: (i) even mild affective states can produce changes in cognitive processes; (ii) emotion and cognition processes are not independent of each other; (iii) positive and negative mood produce qualitative (but not quantitative) changes in information processing (see also Ashby et al., 1999; Vissers et al., 2010; Chwilla et al., 2011).
In spite of the fact that the effects of mood on language processing are not well understood, similar effects of mood on linguistic processing have been reported. Behavioral studies suggest that, under positive mood, subjects’ performance in linguistic tasks includes increased verbal fluency (Phillips et al., 2002), increased production of unusual associations (Isen et al., 1985), enhanced access to remote associates in semantic memory (Bolte et al., 2003; Dreisbach and Goschke, 2004; Dreisbach, 2006; Rowe, et al., 2007), and facilitated implicit judgments of semantic coherence (Bolte et al., 2003). In contrast, negative mood is associated with restricted spread of activation in the semantic network (e.g. Bolte et al., 2003).
According to spreading-activation theories of semantic processing (e.g. Collins and Loftus, 1975), semantic networks are structured along semantic similarity dimension: closely related concepts share more semantic features than concepts distantly related; exemplars of the same semantic category share more attributes and are more interconnected, whereas exemplars of a different or loosely related category are less interconnected. The concept of ‘category’ is based on the similarity of perceptual and functional attributes, and category membership may contain a graded structure, depending on whether and how many attributes are shared by items of that category (e.g. Rosch and Mervis, 1975; Dry and Storms, 2010). Our knowledge about the world is organized in hierarchical taxonomies, so that a ‘superordinate’ level refers to the most inclusive categories (e.g. ‘animals’); at a ‘basic’ level, category members share a significant number of features in common, have similar shapes and similar motor programs (e.g. ‘dog’); finally, less inclusive concepts constitute the ‘subordinate’ level (e.g. ‘golden retriever’) (Rosch and Mervis, 1975; Rosch et al., 1976). More recent studies (Federmeier and Kutas, 1999a, b; Federmeier et al., 2001) bring additional support for the taxonomic organization of semantic memory.
Behavioral findings discussed above suggest that mood can influence the access to knowledge in semantic long-term memory, thus supporting the notion that language and emotion interact. Different theoretical accounts have been proposed to explain the effects of mood on language processing. For example, the network model proposed by Bower (Bower, 1981; Forgas and Bower, 1987) suggests that the current mood experienced by a person will enter into interactions with current cognitive events and influence such cognitive processes as free associations, elaborations and inferences: mood congruent events will be remembered better than incongruent ones. The differential effects of positive and negative mood may also be related to different storage of emotional information in lexical networks: positive mood and related positive information may be associated with a larger or better interconnected network of associations, when compared to negative mood and related negative material (Isen et al., 1978). Accordingly, positive affect may result in changes of cognitive organization, including increased perception of relatedness of cognitive material that may lead to retrieval of a broader range of associates in word association tasks (Isen et al., 1985): for example, in a word association task, Isen et al. (1985) found that people in whom positive mood was induced responded to neutral words (although not to negative words) with a broader range of first associates, when compared with controls under neutral mood. A somewhat different theoretical perspective is taken by a theory of semantic cohesiveness developed by Maratos et al. (2000) who suggested that better recall for emotional words is related to greater semantic cohesiveness of their local semantic networks, and by the theory of emotional salience developed by McNeely et al. (2004) who proposed that the differences observed in retrieval of neutral vs emotional information from semantic memory are related to the emotional salience of the stimulus, such that emotional (in particular, negative) information will be associated with increased perceived saliency. We note that while both the Maratos et al. (2000) and McNeely et al. (2004) proposals are important for the understanding of emotional linguistic material processing, they do not address the effects of mood on neutral content language materials.
Event-related brain potentials (ERPs) methodology is particularly well suited to explore neural underpinnings of mood and language interaction by offering superior millisecond-range temporal resolution that allows tracking neural events as they happen in real time (e.g. Niznikiewicz et al., 1997, 2010). However, the ERP studies focusing on this topic are few. Effects of mood on linguistic tasks were observed for the N400 (a component associated with processing meaning—Federmeier et al., 2001; Chwilla et al., 2011; Federmeier and Kutas, 2011), as well as for the P600 (associated with monitoring and reanalysis processes—Vissers et al., 2010). The N400 is regarded as an index of the difficulty of integrating the meaning of a word into its sentence context (Holcomb, 1993) or the difficulty with which a word can be accessed from the lexicon or semantic memory (Deacon et al., 2000; Kutas and Federmeier, 2000): it is least negative if an item fits well into the previous context (or is more easily accessed from the lexicon) and most negative if the word is not a good fit (or is less easily accessed from the lexicon). Please note that these two accounts of functional significance of the N400 are not mutually exclusive but rather that the N400 is influenced both by the context and by semantic relationships, i.e. the structure of semantic memory, as persuasively argued by Federmeier and Kutas (1999a,b; see also Kutas and Federmeier, 2011). We would like to note that we do not believe that the N400 reflects the final computation of meaning but rather that it reflects a dynamic interplay between contextually influenced, currently available semantic information and contents of semantic long-term memory (Kutas and Federmeier, 2011). Yet another perspective of the N400 has been offered by the inhibition hypothesis of the N400 where the emphasis is placed on the inhibitory processes (Debruille, 2007) and their role in arriving at meaning representations. However, given our focus on the interaction between mood and the structure of semantic memory, this hypothesis is less applicable to the current results.
In addition, it has been recently demonstrated that N400 is sensitive to the impact of mood on linguistic processes (positive mood in Federmeier et al., 2001, and positive and negative mood in Chwilla et al., 2011). Thus, it can be successfully used to probe processes operating within semantic memory and their interactions with context (Federmeier and Kutas, 1999a,b; Khateb et al., 2010), as well as the impact of positive (Federmeier et al., 2001; Chwilla et al., 2011) and negative mood on these processes (Chwilla et al., 2011).
In the studies of Federmeier and colleagues, participants (males and females) either read two-sentence paragraphs presented by themselves (Federmeier, 1999 a,b), or after induction of neutral and positive moods (Federmeier et al., 2001) via pictures selected from the International Affective Pictures System database (IAPS—Lang et al., 2008). The final (target) words in the second sentence were: (i) expected words; (ii) words contextually unexpected but sharing several features with an expected target; and (iii) words contextually unexpected and not sharing many features with the expected target. Two sets of findings emerged. First, it was demonstrated that the N400 amplitude was modulated as a function of semantic distance: it was least negative to contextually expected sentence completions (expected words: EW), followed by contextually unexpected but sharing a category membership targets (within-category violations: WCVs), with the most negative N400 to unexpected words from a different but neighboring category (between-category violation: BCV). These findings suggested that language processing is affected both by the immediate language context used to generate a set of predictions and that these predictions influence word choices in a way that reflects context-independent structure of semantic memory. Second, the N400 effect was modulated by mood: under induced positive mood, decreased N400 amplitude was observed to BCV, with no difference between BCV and WCV, and with both types of sentence endings being more negative than EW. Federmeier et al. (2001) took these findings as evidence that mood can influence the dynamic relationship between prediction processes (i.e. the activation of semantic features of upcoming words prior to their occurrence) and the structure of long-term memory. Of note, a clear demonstration of the mood effect was observed in female but not in male participants.
More recently, Chwilla et al. (2011) explored the relationship between mood and semantic processing in women. Participants were presented with high- (i.e. valid) and low-cloze (i.e. semantically anomalous) sentences, in which the critical word was always in mid-sentence position. Positive and negative mood states were induced via film fragments. A significant mood by cloze interaction revealed that the N400 cloze effect was reduced in the sad relative to happy mood condition. Also, a positive correlation was observed between mood ratings and N400 amplitude: the happier the mood, the larger the N400 effect (i.e. difference between N400 amplitude in the high-cloze condition and N400 amplitude in the low-cloze condition). These findings supported the hypothesis that different mood states lead to qualitative differences in information processing style (happy and global; sad and local).
Predictions for the present study
In the current study, we extended the Federmeier et al. (1999a,b, 2001) design by including both the manipulations of a category membership, and of a positive and a negative mood induction in order to investigate a role of positive and negative mood on semantic processing of categories in the same group of subjects. We also extended the Chwilla et al. (2011) findings by including the neutral mood in addition to the positive and negative moods examined in that study. Finally, the present study differed from both the Federmeier et al. (2001) and the Chwilla et al. (2011) studies in the population studied (males only), details of paradigm design and task requirements. We examined the interaction between (i) mood, (ii) contextual congruity and (iii) semantic memory organization.
We hypothesized that the structure of semantic memory would interact with the sentence context resulting in least negative N400 to EW, followed by WCV and BCV. Importantly, we predicted that this effect would interact with mood. Under the neutral mood, we expected the N400 effects as described above for the Federmeier et al. (1999a,b). For example, the sentence ‘The Jones’ made so much money that they no longer wanted to live in their small house so they moved to the outskirts of the town to a very fashionable subdivision. There, they bought a huge…’ would create an expectation for a final word ‘mansion’. The N400 recorded to this final word would be reduced relative to the word ‘apartment’ that is unexpected but shares some semantic features with ‘mansion’, and would be most negative to ‘tepee’ that shares few features with ‘mansion’: both ‘mansion’ and ‘apartment’ are types of dwellings that are meant to be non-portable and permanent, while ‘tepee’ is a fragile and portable shelter. Thus, even though all three words belong to a super-ordinate category of human dwellings, the preceding context makes ‘mansion’ the best choice. This hypothesis is consistent with a predictive process borne out of available context and interacting with the extent of feature overlap between the target and its close and distant associates resulting in N400 EW < N400 WCV and N400 WCV < N400 BCV.
Considering the broadening effects of positive mood demonstrated in lexical access (e.g. Isen et al., 1985) and semantic memory tasks (e.g. Bolte et al., 2003; Dreisbach, 2006; Rowe et al., 2007) and its facilitatory effect on semantic processing (Chwilla et al., 2011), we hypothesized that positive mood would facilitate word choices such that a broader set of semantic features would be interpreted as closely overlapping with the best fit target. This would be reflected in similar N400 amplitude to both EW and WCV.
Given behavioral studies suggesting a restrictive effect of negative mood on lexical access and indicating its association with a narrowed/local information processing style (e.g. Fiedler, 1988; Bolte et al., 2003; Chwilla et al., 2011), we speculated that the negative mood would restrict prediction mechanisms and thus increase the sensitivity of the language comprehension system to contextual information allowing for a distinction between contextually expected (EW) and unexpected (WCV and BCV) words, but not between the two types of violations. This would be translated into similar N400 amplitude for both WCV and BCV. Two sets of predictions were possible for EW: (i) the restrictive effects of mood will not apply to EW and the N400 to EW will be unchanged and (ii) the effects of mood will also affect EW and the N400 to EW will be further reduced.
METHODS
Participants
Fifteen subjects (all males) (mean age = 45.07; s.d. = 8.62; age range 26–54) participated in the ERP experiment (none participated in stimuli norming procedures). Inclusion criteria for participation in the study were: (i) age between 18 and 50 years; (ii) right handedness (Edinburgh Inventory, Oldfield, 1971); (iii) no history of neurological illness; (iv) no history of alcohol or drug abuse; (v) no history of psychiatric disorder in oneself or in first-degree relatives; (vi) no current medication for medical disorders that would have effects on electroencephalogram (EEG) morphology or consequences at the level or neurological and/or cognitive functioning; (vii) verbal intelligence quotient (IQ) above 75; (viii) no alcohol use in the 24 h before testing; (ix) an ability and desire to cooperate with the experimental procedures, as demonstrated by signed informed consent, following Harvard Medical School and Veterans Affairs Boston Healthcare System guidelines.
Participants were recruited from internet advertisements to serve as controls in a larger study on the neurophysiological correlates of language processes in schizophrenia spectrum disorders. All subjects were screened using the Structured Clinical Interview for DSM-IV for Axis I (SCID-I—First et al., 2002) and Axis II (SCID-II—First et al., 1995) disorders. In addition, they underwent neurocognitive assessment (Wechsler Intelligence Adult Scales-III—Wechsler, 1997) (Table 1).
Table 1.
Socio-demographic and cognitive characterization of participants (Mean ± s.d.)
| Participants (n = 15) | |
|---|---|
| Age | 45.07 (8.62) |
| Years of education | 14.70 (1.79) |
| Parent’s social–economic statusa | 2.40 (1.12) |
| Subject’s social–economic statusa | 2.13 (1.13) |
| Verbal IQb | 103.93 (16.53) |
| Full-Scale IQb | 103.73 (13.50) |
| Vocabularyb, c | 10.50 (2.96) |
| Verbal Comprehension Indexb | 100.13 (12.74) |
aHollingshead Two-Factor Index of Social Position (Hollingshead, 1965).
bWAIS-III: Weschler Adult Intelligence Scale – III (Weschler, 1997).
cScaled score.
Materials
Three hundred and twenty-four pairs of sentences, distributed across three main lists, were used as stimuli. Additionally, 30 positive, 30 negative and 30 neutral pictures selected from the IAPS data set were used for the emotional elicitation procedure1 (Table 2), following previous studies where IAPS pictures proved effective in eliciting mild changes in affect (e.g. Federmeier et al., 2001; Simon-Thomas and Knight, 2005).
Table 2.
Mean (s.d.) scores for the pictures’ valence and arousal, in each mood condition
| Mood condition | Valence Mean (s.d.) | Arousal Mean (s.d.) |
|---|---|---|
| Neutral | 5.05 (0.19) | 2.65 (0.36) |
| Positive | 7.85 (0.25) | 4.48 (0.38) |
| Negative | 2.74 (0.35) | 4.78 (0.28) |
Note: IAPS picture ratings as published by Lang et al. (2008).
Following the procedure of Federmeier and Kutas (1999a,b), the sentences ended with three possible types of target words: (i) EW (items with the highest cloze probability); (ii) WCV (items that were unexpected given the semantic context and that were derived from the same semantic category as the expected items); (iii) BCV (items that were unexpected given the semantic context and that were derived from a different, but related semantic category to the expected word) (Appendix 1).
The target words were taken from 15 categories including: plants, animals, body parts, foods, beverages, types of dwellings, land formations, vehicles, tools and household objects, office supplies, furniture, garments and personal articles, leisure and hobby-related items (e.g. sports), jobs and music instruments.
The first sentence of the pair established the context and the second sentence completed it with an expected or an unexpected ending given the previous context. The second sentence was always appropriately completed by each of the three final word types (concrete nouns only). The sentence did not contain lexical associates to any ending type to avoid lexical associative priming.
We followed Federmeier and Kutas (1999a,b) in stimuli development. The expected words were obtained in a separate study where cloze probabilities for the final words of each of the sentence contexts (324) presented in the experiment were calculated. Twenty-six volunteers (mean age = 24.31, s.d. = 6.93, age range 21–57 years; 21 females and 5 males) were asked to complete each sentence context with the first word that came to mind and that made its appropriate completion. Cloze probability was calculated as the proportion of individuals who chose a particular word to complete a given sentence context. Mean cloze probability for the expected words was 0.75.
In addition, a different group of twenty individuals (mean age = 26.20, s.d. = 12.17, age range 17–61 years; 16 females and 4 males) was asked to rate the plausibility of all sentence endings within their contexts. Sentences were divided into three lists of 108 sentences each (so that no sentence context or critical word was repeated within each list), corresponding to the three experimental blocks. Each block was presented to a different subset of subjects. They were asked to rate ‘How well do these sentences make sense together?’ on a percent scale. Mean plausibility was calculated as the average of the ratings for all items of each of the three conditions (EW, WCV and BCV). Mean rated plausibility for EW was 86.44%, for WCV was 24.22% and for BCV was 12.54%. A repeated-measures analysis of variance (ANOVA) showed a significant effect of ending type [F(2,40) = 123.12, P < 0.001], with EW rated as significantly more plausible than WCV (P < 0.001) and BCV (P < 0.001), and WCV rated as more plausible than BCV (P < 0.001).
Sentences were distributed across three blocks [block 1 (1–108); block 2 (109–216); block 3 (217–324)]. Each sentence context could end with a plausible (EW) or implausible ending (WCV; BCV). Each target word appeared three times, once in each type of ending—EW, WCV or BCV,2 and once per set. The three target word types were matched for length (mean number of letters = 5.76; s.d. = 1.73), frequency (M = 50.21; s.d. = 79.37), imageability (M = 585.93; s.d. = 39.67), concreteness (M = 582.63; s.d. = 44.09) and age of acquisition (M = 282.55; s.d. = 81.86), taken from the MRC Psycholinguistic Database—http://www.psy.uwa.edu.au/mrcdatabase/uwa_mrc.htm (Coltheart, 1981; Wilson, 1998).
The sentences were rotated across blocks and lists, so that no sentence context or critical word was repeated for a particular participant within a given block (Figure 1). Across participants, each target word appeared in one of the three possible conditions (EW, WCV and BCV). In order to balance the number of plausible and implausible sentences, 36 plausible filler sentences were added to each list (Federmeier and Kutas, 1999a,b). In the end, all the lists had the same number of expected and unexpected sentence endings (72 sentences of each type).
Fig. 1.

Illustration of the counterbalancing procedure: each subject saw a different set of sentences across three moods; at the same time, across subjects, each sentence set was seen in all three moods. Both moods and sentence sets were counterbalanced across subjects. Colors show different orders for the presentation of blocks in each mood. Each block included 36 sentences of each critical condition (expected words, within-category violations and between-category violations).
Each participant viewed a different list in each mood. Sentence contexts and items were used only once per list and each list consisted of 36 of each target type (Figure 1). All sentences and pictures were presented via Superlab software 4.0.6.b (Cedrus Corporation, 2008). Pictures were not semantically related to sentence contexts and sentence endings.
Procedure
Participants were tested in three experimental sessions (one for each mood induction: neutral, positive and negative). At least a week separated the sessions. The experiment was conducted in a soundproof, electrically shielded chamber. Participants were seated in a comfortable chair 40 inches in front of a monitor.
Before the experiment, they were asked to complete the ‘Profile of Mood States’ (McNair et al., 1971/1981), a questionnaire with 65 adjectives rated on a 5-point scale (from 0 = not at all to 4 = extremely) that covers different mood states indexed by seven subscales: Depression, Tension, Anger, Vigor, Fatigue, Confusion and Friendliness. In order to have a more comprehensive description of participants’ current mood states, the following positive items were added: ‘happy’, ‘clear headed’, ‘relaxed’, ‘content’, ‘satisfied’, ‘hopeful’. This checklist has been used by several studies aiming at investigating mood effects (e.g. Steele et al., 1999; Canli et al., 2004; Demirtas-Tatlidede et al., 2010), and was demonstrated to have good test–retest reliability.
The same questionnaire was administered at the end of an ERP experimental session. Thus, each subject responded to the questionnaire in each affect induction session twice: before and after mood induction (Figure 2).
Fig. 2.

Schematic illustration of the experimental design.
Before sentences’ presentation, participants saw 30 pictures from the IAPS database (positive, negative or neutral). Each picture was presented for 1000 ms; a fixation cross appearing in the centre of the screen for 2000 ms preceded each picture; the inter-stimulus interval was 2000 ms. The order of mood induction was counterbalanced across participants. They were not informed about the true purpose of the pictures but, as in the Federmeier et al. (2001) study, they were instructed to pay attention to the stimuli and told that later they would be asked to judge if the pictures were appropriate for use in another experiment. In order to familiarize participants with the instructions and task, a short practice block preceded the testing session.
To keep mood constant across the experiment, a picture from the set of selected IAPS pictures for a specific mood induction was presented briefly (350 ms) before each pair of sentences. It was followed by a fixation cross, presented for 1500 ms (Figure 3). The first sentence of the pair appeared fully on the screen. Participants read each sentence at their own pace and pushed a button to view the second sentence. The second sentence was presented one word at a time, horizontally, each word for about 600 ms. Final words were presented in the centre of the screen for 1000 ms. Participants were asked not to blink or move their eyes during the second sentence, to minimize EEG artifacts. The final word was followed by a question mark (for 3000 ms), prompting the participants to make a button press response, after which the next sentence appeared automatically. Subjects were asked to decide if the two sentences together made sense or not. The order of ‘yes’ and ‘no’ response buttons was counterbalanced across subjects. A short break was given after every 20 pairs of sentences.
Fig. 3.
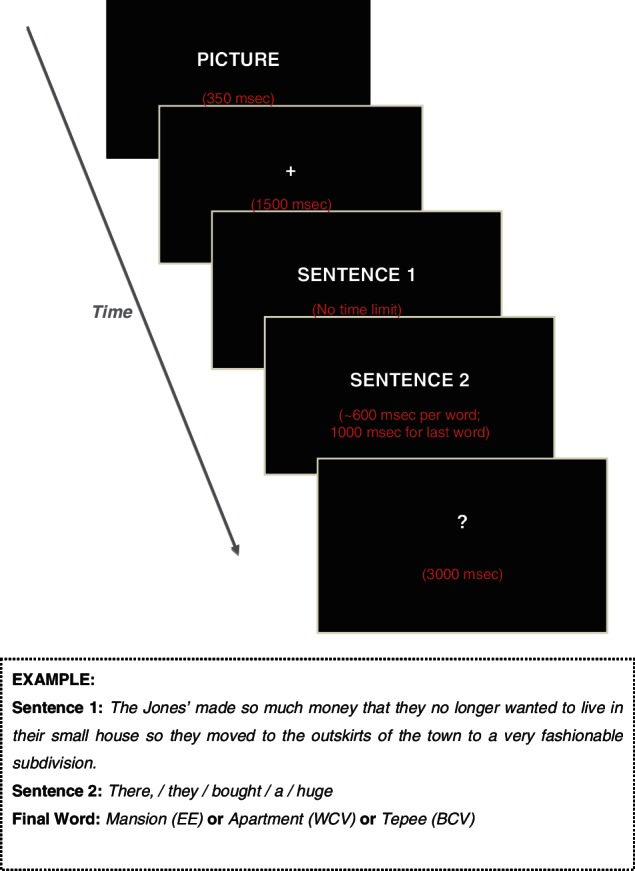
Schematic illustration of an experimental trial.
At the end of the experiment, in addition to the ‘Profile of Mood States’ questionnaire, participants answered a written questionnaire, designed by the authors, asking them to report: (i) how much they liked the set of pictures; (ii) how much seeing the pictures influenced the way they felt; and (iii) how much the pictures influenced their decision whether the sentences made sense or not.
At the end of the three sessions, participants were also asked to rate the valence and arousal of each picture presented, by using a computerized version of the Self-Assessment Manikin (SAM) (Lang et al., 2008).
EEG data acquisition
Sixty-four channel EEG was collected using custom designed electrode caps from BioSemi system (Active 2). EEG was acquired in a continuous mode at a digitization rate of 512 Hz, with a bandpass of 0.01–100 Hz, and stored for later analysis. Blinks and eye movements were monitored via electrodes placed on left and right temples and one above the left eye. Electrode impedances were kept below 5 KΩ.
ERP data analysis
Analysis of raw EEG data was conducted using Analyzer software package (Brain Products, Inc, 2000). EEG was re-referenced off-line to the average of the left and right mastoids. Individual ERP epochs for each target type were constructed with 150 ms pre-stimulus baseline and 1000 ms epoch after the stimulus onset. Eye blink and movement artifacts were corrected by the method of Gratton et al. (1983, BrainVision Analyzer package). Trials containing excessive eye movements, blinks, muscle activity or amplifier blocking were rejected off-line before averaging: single-trial epochs with voltage exceeding ±80 μV were rejected. Individual averages were considered for further analysis if at least 80% (i.e. 29) of segments available for a given condition passed the artifact rejection. On average, 33.24 segments were accepted in the neutral mood, 33.60 for the positive mood and 30.02 segments in the negative mood condition.
Visual inspection of grand averages localized the effects of mood and sentence condition to central and parietal electrode sites consistent with previous reports of centro-parietal distribution of N400 in visual paradigms (see Kutas and Federmeier, 2011). In addition to grand averages to each word type (Figures 5–7), we have also constructed difference waveforms that illustrate the N400 effect or the difference between the N400 to expected endings and the types of incongruous endings—WCV and BCV (Figures 8–10)—as well as the amplitude maps of these effects (Figure 11). They confirm the centro-parietal distribution of the N400 in this paradigm. Following previous studies (e.g. Federmeier et al., 2005), N400 amplitude was measured as mean voltage within 300–500 ms post-stimulus latency window to each sentence ending type, in each mood, at frontal (Fz, F1/2, F3/4), central (Cz, C1/2, C3/4) and parietal (Pz, P1/2, P3/4) regions.
Fig. 5.

Grand average waveforms for EW, WCV and BCV, under neutral mood.
Fig. 6.

Grand average waveforms for EW), WCV and BCV, under positive mood.
Fig. 7.

Grand average waveforms for EW, WCV and BCV, under negative mood.
Fig. 8.

Grand average difference waveforms for unexpected final endings (WCV, BCV), under neutral mood. Notes: LH = left hemisphere (average of amplitude of electrodes located in the left hemisphere that were included in the MANOVA analysis—frontal: F1/3; central: C1/3; parietal: P1/3); RH = right hemisphere (average of amplitude of electrodes located in the right hemisphere that were included in the MANOVA analysis—frontal: F2/4; central: C2/4; parietal: P2/4); midline = Fz, Cz, Pz.
Fig. 9.

Grand average difference waveforms for unexpected final endings (WCV, BCV), under positive mood. Notes: LH = left hemisphere (average of amplitude of electrodes located in the left hemisphere that were included in the MANOVA analysis—frontal: F1/3; central: C1/3; parietal: P1/3); RH = right hemisphere (average of amplitude of electrodes located in the right hemisphere that were included in the MANOVA analysis—frontal: F2/4; central: C2/4; parietal: P2/4); midline = Fz, Cz, Pz.
Fig. 10.

Grand average difference waveforms for unexpected final endings (WCV, BCV), under negative mood. Notes: LH = left hemisphere (average of amplitude of electrodes located in the left hemisphere that were included in the MANOVA analysis—frontal: F1/3; central: C1/3; parietal: P1/3); RH = right hemisphere (average of amplitude of electrodes located in the right hemisphere that were included in the MANOVA analysis—frontal: F2/4; central: C2/4; parietal: P2/4); midline = Fz, Cz, Pz.
Fig. 11.
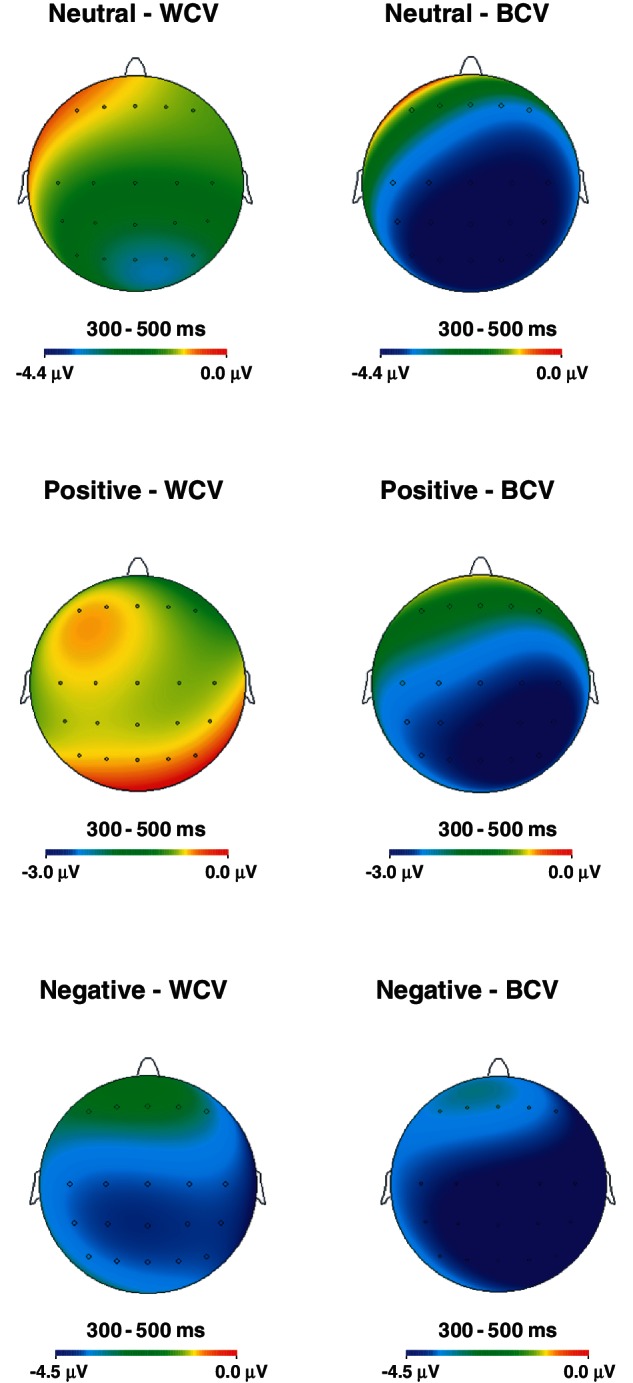
Topographic maps depicting voltage distribution across the scalp for WCV and BCV, under the three moods.
Statistical analyses
Behavioral data
In order to explore the effectiveness of the mood induction procedure, paired sample t-tests were computed for Profile of Mood States scale (POMS) scores before and after mood induction for each experimental session. In addition, the participants’ responses to the short post-experiment questionnaire were subjected to repeated-measures ANOVA with mood (3 levels) and question (3 levels) as dependent variables. Accuracy to target words was subjected to repeated-measures ANOVA, with mood (neutral, positive and negative), and sentence condition (EW, WCV and BCV) as dependent variables. Since responses were delayed until the question mark appeared, the reaction time data were not analyzed.
N400 ERP analyses
For the N400 amplitude analyses, the initial omnibus MANOVA with four within-factors of mood (3 levels), sentence condition (3 levels), region (3 levels) and electrodes (5 levels) was followed by separate MANOVAs for each mood (neutral, positive and negative), with sentence condition (EW, WCV and BCV), region (frontal, central and parietal), and electrodes (Fz, F1, F2, F3, F4, Cz, C1, C2, C3, C4; Pz, P1, P2, P3, P4) as within-subjects factors. Main effects were followed with planned comparisons with Bonferroni correction.
We note that we focused our analyses on the effects of each mood on the three sentence conditions. Thus, our interest was in how a given mood type, for example, positive, influences the processing of targets that share either many or few overlapping features with the best word fit and in their interactions with the context, rather than in how each word type is influenced by the three moods (neutral, positive and negative). Even though related, these are separate questions. Since each mood induction and the related experimental session were conducted on separate days, the question on how different moods influence the processing of the same information type, here EW, WCV and BCV, is harder to address. Nonetheless, we have also examined it (Figures 12–15) and we believe that the results supplement our main analyses in a noteworthy way.
Fig. 12.

Mean amplitude for each sentence condition, across the three moods.
Fig. 13.

Grand average waveforms for expected endings in the three mood conditions. Notes: LH = left hemisphere (average of amplitude of electrodes located in the left hemisphere that were included in the MANOVA analysis—frontal: F1/3; central: C1/3; parietal: P1/3); RH = right hemisphere (average of amplitude of electrodes located in the right hemisphere that were included in the MANOVA analysis—frontal: F2/4; central: C2/4; parietal: P2/4); midline = Fz, Cz, Pz.
Fig. 14.

Grand average difference waveforms for WCV in the three mood conditions. Notes: LH = left hemisphere (average of amplitude of electrodes located in the left hemisphere that were included in the MANOVA analysis—frontal: F1/3; central: C1/3; parietal: P1/3); RH = right hemisphere (average of amplitude of electrodes located in the right hemisphere that were included in the MANOVA analysis—frontal: F2/4; central: C2/4; parietal: P2/4); midline = Fz, Cz, Pz.
Fig. 15.

Grand average difference waveforms for BCV in the three mood conditions. Notes: LH = left hemisphere (average of amplitude of electrodes located in the left hemisphere that were included in the MANOVA analysis—frontal: F1/3; central: C1/3; parietal: P1/3); RH = right hemisphere (average of amplitude of electrodes located in the right hemisphere that were included in the MANOVA analysis—frontal: F2/4; central: C2/4; parietal: P2/4); midline = Fz, Cz, Pz.
RESULTS
Effectiveness of mood induction
POMS questionnaire
Paired t-tests did not show any changes after ‘neutral mood’ induction. A trend for reduced depression–dejection was observed after ‘positive mood’ induction [t(14) = 2.039, P = 0.061], as well as a trend for decreased happiness after ‘negative mood’ induction [t(14) = 1.871, P = 0.082].
Post-experiment questionnaire
A significant mood × question interaction [F(6,9) = 24.763, P < 0.001] was found for the post-experimental questionnaire. Participants liked the positive pictures more than the neutral (P = 0.011) and negative pictures (P < 0.001), and they liked the neutral pictures more than the negative ones (P < 0.001). In addition, negative pictures affected the way participants felt more than neutral pictures (P = 0.011), and positive pictures affected the way they felt more than the neutral ones at trend level (P = 0.079).
IAPS pictures ratings
Table 3 shows participants ratings (means ± s.d.) of valence and arousal of each picture presented for mood induction purposes. Very similar ratings were observed between these ratings and norms published by Lang et al. (2008).
Table 3.
Participants’ (n = 15) ratings of IAPS pictures’ valence and arousal
| Mood condition | Valence Mean (s.d.) | Arousal Mean (s.d.) |
|---|---|---|
| Neutral | 5.08 (0.72) | 2.78 (1.59) |
| Positive | 7.28 (1.09) | 5.12 (2.60) |
| Negative | 2.49 (0.77) | 4.70 (2.60) |
Behavioral results
Accuracy
A main effect of sentence condition was found [F(2,13) = 45.228, P < 0.001]: more correct responses were observed for EW sentences, followed by BCV, and then by WCV (Figure 4). The number of correct responses differed significantly between EW and WCV (P < 0.001), between EW and BCV (P = 0.025), and between WCV and BCV (P < 0.001). However, the number of correct responses did not differ as a function of mood [F(4, 11) = 2.303, P = 0.124].
Fig. 4.

Mean number of correct responses for sentences ending with EW, WCV and BCV, under neutral, positive and negative moods. Note. Error bars show s.d.
ERP results
N400 amplitude
The omnibus MANOVA revealed a main effect of sentence condition [F(2,13) = 26.631, P < 0.001]. Planned comparisons showed significantly less negative N400 amplitude for EW relative to both WCV (P < 0.01) and BCV (P < 0.001) (see Table 4). In addition, N400 to WCV was significantly less negative than to BCV (P < 0.01).
Table 4.
N400 mean amplitude at central and parietal regions for each sentence ending under neutral, positive and negative moods
| Mood | Sentence ending | Region |
|
|---|---|---|---|
| Central | Parietal | ||
| Neutral | EW | 3.83 (2.12) | 4.63 (2.33) |
| WCV | 1.87 (1.89) | 1.95 (1.90) | |
| BCV | 0.42 (2.66) | 0.71 (2.30) | |
| Positive | EW | 3.53 (3.31) | 3.89 (3.83) |
| WCV | 2.86 (2.70) | 3.31 (3.04) | |
| BCV | 1.08 (3.22) | 1.40 (3.65) | |
| Negative | EW | 5.61 (3.94) | 6.21 (4.20) |
| WCV | 2.53 (3.22) | 3.30 (3.40) | |
| BCV | 2.00 (3.12) | 2.93 (3.41) | |
Importantly, results revealed a significant mood × sentence condition × region interaction [F(8,7) = 3.770, P = 0.049]. In order to better understand the differential effects of mood on N400 amplitude for each sentence condition, we followed these interactions with MANOVAs for each mood separately in each of the regions.
Neutral Mood
No effect of sentence condition was observed at frontal electrode sites [F(2, 13) = 1.431, P > 0.05]. However, results showed a main effect of sentence condition at central electrode sites [F(2, 13) = 12.861, P < 0.01]. Pairwise follow-up analyses showed that N400 was significantly less negative for EW relative to both WCV (P = 0.021) and BCV (P < 0.01), and less negative for WCV relative to BCV (P = 0.012) (EW: mean = 3.83 µV; WCV: mean = 1.88 µV; BCV: mean = 0.42 µV). A sentence condition effect was also observed at parietal electrode sites [F(2, 13) = 14.219, P < 0.01]: N400 was significantly less negative for EW relative to both WCV (P < 0.01) and BCV (P < 0.001), and it was less negative for WCV relative to BCV (P = 0.025) (EW: mean = 4.63 µV; WCV: mean = 1.95 µV; BCV: mean = 0.71 µV).
Positive Mood
Again, no effect of sentence condition was observed at frontal electrode sites [F(2, 13) = 2.742, P > 0.05] but it was present in the central region [F(2, 13) = 14.910, P < 0.001]. Planned comparisons revealed that the impact of positive mood was reflected in a lack of N400 amplitude differences between EW and WCV (P = 0.949). N400 amplitude to EW and to WCV was significantly less negative than BCV N400 amplitude (EW: mean = 3.53 µV; WCV: mean = 2.86 µV; BCV: mean = 1.08 µV) (EW vs BCV: P < 0.001; WCV vs BCV: P = 0.079). An effect of sentence condition was also observed in the parietal region [F(2,13) = 12.426, P < 0.01]: no differences in N400 amplitude were observed between EW and WCV (P = 1.000). N400 amplitude to EW and WCV was significantly less negative than to BCV (EW: mean = 3.89 µV; WCV: mean = 3.31 µV; BCV: mean = 1.40 µV) (EW vs BCV: P < 0.01; WCV vs BCV: P = 0.037).
Negative Mood
Even though no effect of sentence condition was observed at frontal electrode sites [F(2, 13) = 1.084, P > 0.05], this effect was present in the central region [F(2, 13) = 21.007, P < 0.001]. N400 amplitude for EW differed significantly from WCV (P < 0.01) and BCV (P < 0.001). However, N400 for WCV did not significantly differ between WCV and BCV (P = 0.489) (EW: 5.61 µV; WCV: 2.53 µV; BCV: 2.01 µV).
An effect of sentence condition was also observed in the parietal region [F(2,13) = 19.710, P < 0.001]: N400 amplitude for EW differed significantly from WCV (P < 0.01) and BCV (P < 0.001). However, N400 for WCV did not significantly differ between WCV and BCV (P = 1.000) (EW: 6.21 µV; WCV: 3.30 µV; BCV: 2.93 µV).
The effects of mood on each sentence condition
A significant effect of mood for EW [F(2, 13) = 5.585, P = 0.018] revealed that N400 amplitude was more positive under negative mood than under positive mood (P = 0.012). No significant mood effect was observed for WCV, and a trend level mood effect was observed for BCV [F(2, 13) = 2.775, P = 0.099].
DISCUSSION
In this study, we explored the influence of mood on semantic memory organization in its interactions with sentential context, using both behavioral and electrophysiological data. We sought to examine how transient mood can contribute to the putative broadening (positive mood) or narrowing (negative mood) of the extent of activation of a set of semantic features activated by the available context and/or to the facilitation of semantic connections within long-term memory, to substantiate observations from behavioral literature that mood states can influence thinking style.
The results provided support for the concept of hierarchical organization of semantic memory according to categories, an observation made previously (Federmeier and Kutas, 1999a,b), the fact that access to these categories can be flexibly modulated by mood (Federmeier et al., 2001), and that, more broadly, mood states interact with language processing (Chwilla et al., 2011). Importantly, we demonstrated that both positive ‘and’ negative mood can exert these modulatory effects in unique ways.
For neutral mood, our results are consistent with those of Federmeier and Kutas (1999a, b): least negative N400 was associated with EW, followed by unexpected sentence endings from the same category as the EW (i.e. WCV), and then by unexpected sentence endings from a different, but neighboring category (i.e. BCV), where the most negative N400 was observed. This N400 pattern suggested that unexpected items that shared more features (e.g. ‘apartment’) with the predicted final word (e.g. ‘mansion’) were more easily accessed than unexpected items that belonged to a neighboring semantic category (e.g. ‘tent’) and shared fewer semantic features with the predicted final word. This finding suggests that N400 is sensitive to a degree of semantic feature overlap between an expected word and its semantic neighbors. If this fine-tuned neural response to semantic features did not exist, we would have observed similar N400 amplitudes for both types of unexpected sentence endings (i.e. WCV and BCV) in the neutral mood condition, that is the N400 would reflect plausibility of both endings relative to existing context (both WCV and BCV were implausible). Behavioral data also supported the differential status of WCV and BCV: more incorrect responses were observed for WCV relative to EW and BCV, under neutral mood. Thus, words from the same semantic category (i.e. sharing more semantic features with the best fit target) were perceived more frequently as appropriate endings than words from a neighboring category, which suggests that the structure of semantic memory (independently of sentence context) was affecting sentence processing, also at the behavioral level. This impact of semantic memory structure on ERP and behavioral effects observed in our study was present in spite of the fact that the categorical relationships between the items were never mentioned to the participants.
Importantly, our findings pointed to a differential impact of positive and negative mood induction on the way in which context interacted with category membership.
Under positive mood, unexpected words from the same category as the expected ones (e.g. ‘apartment’) became processed as if they were expected sentence endings (e.g. ‘mansion’). While the processing of the word ‘apartment’ was facilitated under positive mood leading to a reduced N400, the word ‘tent’ (a BCV) remained outside a pool of acceptable endings and was associated with a larger N400 relative to both EW and WCV, presumably because it shared few semantic features with the expected word. Thus, the N400 for sentence endings under positive mood reflected three sources of modulatory influences: (i) the context that guided the generation of probable word candidates and led to the inhibition of irrelevant semantic features and inappropriate situational representations (Debruille, 2007); (ii) the structure of semantic memory where words are connected along the gradient of experientially formed category membership understood as the extent of feature overlap; and (iii) mood. This result has been additionally informed by the comparison of different ending types as a function of mood. The N400 to EW changed little as a function of positive mood (4.2 µV under neutral mood and 3.7 µV under the positive mood), while the N400 to WCV increased from 1.9 µV under the neutral mood to 3.1 µV under positive mood; the N400 also increased for the BCV from 0.6 to 1.2 µV, but still remained significantly less positive relative to the two other ending types.
The present findings (decreased N400 amplitude for WCV resulting in a lack of a difference between EW and WCV) differ from the ones reported by Federmeier et al. (1999): in the latter, positive mood led to the decreased N400 amplitude to BCV, resulting in a lack of amplitude difference between WCV and BCV. Differences between the two studies may be attributed to: stimuli (different sentence contexts were used in both studies, even though the present study has followed the sentence design originally reported by Federmeier and Kutas, 1999a,b), experimental design (in the present study, pictures were briefly presented before each sentence context in order to keep mood constant), samples (the sample of the present study consisted of males only; of note, in the Federmeier et al., 2001 study, the modulatory effects of mood on semantic processing were primarily found in women but not in men), and task instructions (in the study of Federmeier et al., 2001, participants were only asked to read the sentences and memorize them, while in the present study they were instructed to decide if the two sentences together made sense or not).
The facilitative impact of positive mood may be mediated by two possible mechanisms, according to an integration account of N400. For example, mood may impact the predictive mechanisms leading to the generation of a larger pool of plausible endings. Since according to the prediction account of N400, context pre-activates features associated with the concept that is most likely to come next, positive mood may strengthen the facilitation of these words that share more features with the EW (even if they do not fit the sentence context well and were judged as implausible endings—i.e. WCV), leading to the absence of N400 amplitude difference between EW and WCV. The other possible way that mood can influence language processing is by changing the gradient of connections among words in semantic memory such that, for example, apartment and ‘mansion’ become more alike. Finally, since we have proposed a dynamic interaction between the context and the structure of semantic memory these two mechanisms may interact.
In addition, for the first time, our results offered electrophysiological evidence on the effects of negative mood on semantic processing. Our ERP findings suggest that inducing a negative affect biases the processing of WCV (e.g. ‘apartment’) in relation to BCV (e.g. ‘tent’): the N400 to WCV and BCV had similar amplitude. As for the positive mood, negative mood can impact both the predictive contextual processes and connections within semantic memory. Thus, an integration account of N400 would predict that the negative mood may influence a set of expected words predicted by context, leading to a narrower set, such that making a fit between a context and the final word is actually easier with both BCV and WCV being outside of the pool of plausible endings. Second, it may alter a gradient of connections among semantic associates such that ‘apartment’ and ‘mansion’ seem like not related rather than related concepts.
Again, comparisons of the N400 amplitude to each word type as a function of mood provided additional insights. Under the negative mood, the N400 to EW was significantly less negative than the N400 to positive or neutral mood. The N400 to WCV (2.9 µV) under the negative mood was similar to the N400 under the positive mood (3.1 µV), with both reduced relative to the neutral mood (1.9 µV) (although not different statistically). The reduction of the N400 amplitude to EW is revealing. Since EW were best context completions, its further reduction under the negative mood must be interpreted as further narrowing of the set of predictions likely via further constricting a set of appropriate endings in a process of inhibition, leading to an even smaller N400 amplitude. We assume that the same narrowed set of predictions lead to the reduction of the N400 amplitude both in the WCV and BCW; when compared to EW, the N400 to both WCV and BCW was significantly more negative than the N400 to EW. Thus, under the negative mood, the generation of the narrowed set of predictions seems to dominate linguistic processes such that the language comprehension system is sensitive only to contextual information and congruency effect, but not to the relationship between concepts in the long-term memory.
Even though, in contrast to positive mood, research on negative mood is less consistent, these narrowing effects of contextually guided predictions seem to be supported by previous behavioral studies that have suggested that negative mood can impair creativity (Vosburg, 1998) and is associated with a more analytic, localized or focused mode of processing (Bolte et al., 2003; Park and Banaji, 2000; see also Mitchell and Phillips, 2007 for a review). This more analytic processing style (more detail-oriented) translated into a heightened sensitivity to contextual information, which allowed subjects to create a strong expectation for a given word. The present results elucidate further theorizing related to the effects of negative mood based thus far on behavioral studies results: it appears that its restrictive effects on the activation of knowledge representations are realized through the narrowed set of predictions (inhibition mediated) generated by the context.
In summary, our results suggest a differential modulation of access to semantic memory as a function of mood, at the electrophysiological level, with each mood impacting linguistic processing in a unique way. While, we claim, each mood impacted the linguistic processes in a different way, the interaction between the contextual influences and the structure of working memory persisted across the two moods as indexed by the main effect of sentence condition and the behavioral findings of more errors to WCV. This seeming discrepancy between the behavioral effects and the N400 effect further underscores the functional significance of the N400 as an index of a dynamic construction of meaning that enters into interactions with context, the contents of semantic message, and, as in our case, mood states, but does not reflect the final interpretation of what is said and written (Debruille, 2007; Kutas and Federmeier, 2011).
Caution is needed when interpreting these findings, given that only males participated in this study and gender differences have been reported in studies of affective processing (e.g. Koch et al., 2007). However, since previous results suggest that mood influences may be more readily detected in females than males, our results suggest that similar if not greater effects should be observed in females. Changes in our induced mood changes were only observed at the trend level, although both IAPS ratings and responses to specifically designed postinduction questions suggest effective mood manipulation. In addition, we have induced mood in three separate sessions since our focus was on the effects of a particular mood on three types of contextually and semantically related word types. However, this design makes comparisons of the same word type across moods more difficult.
In spite of these limitations, and consistent with previous behavioral studies, our findings suggest that positive and negative mood differentially affect the way we make sense of the constant flow of information, in particular semantic information. Future studies are needed to further enhance our understanding of the effects of mood on sentence comprehension.
Conflict of Interest
None declared.
Acknowledgments
The authors would like to thank to all the participants of the study, as well as to Jenna Mezin and Elizabeth Thompson for their help with data collection. This work was supported by a Doctoral Grant from Fundação para a Ciência e a Tecnologia – Portugal (SFRH/BD/35882/2007 to A.P.P.) and by the National Institute of Mental Health (RO1 MH 040799 to R.W.M.; RO3 MH 078036 to M.A.N.).
We would like to thank Dr. B. Debruille for his contribution to the title of the paper.
Appendix 1 Examples of sentences used in the experiment
A. Mansion/apartment/tepee/tent
The Jones’ made so much money that they no longer wanted to live in their small house so they moved to the outskirts of the town to a very fashionable subdivision. There, they bought a huge mansion (EW)/apartment (WCV)/tent (BCV).
For summer vacation, Jack and his friends decided to spend a week camping and they planned on hiking the whole day. They would spend the night in their tents (EW)/tepees (WCV)/mansions (BCV).
Most Native American tribes did not build permanent wooden houses but instead they used animal skins and birch bark to build their dwellings. They were called tepees (EW)/tents (WCV)/apartments (BCV).
When Sam decided to move downtown, he had to find a new place to live but he did not have much money. So, he rented a small apartment (EW)/mansion (WCV)/tent (BCV).
B. Doctor/dentist/teacher/librarian
Louise was suffering from a toothache for several days, but she still refused to do anything about it. She has always been afraid of going to the dentist (EW)/doctor (WCV)/librarian (BCV).
Paul loved to watch all TV shows that were set in hospitals. He has always wanted to be a doctor (EW)/dentist (WCV)/teacher (BCV).
Michaela loved to read and she could not wait to check out a new set of books. She knew they were set aside for her by the friendly librarian (EW)/teacher (WCV)/doctor (BCV).
Mr Chase was demanding but he always knew how to inspire and motivate his students. By far, he was Cindy’s favorite teacher (EW)/librarian (WCV)/dentist (BCV).
Footnotes
1IAPS pictures included: Negative—2141, 2276, 2278, 2375, 2455, 2700, 2750, 2799, 2900, 3181, 3300, 6311, 6561, 9000, 9007, 9041, 9102, 9180, 9265, 9290, 9320, 9419, 9421, 9432, 9435, 9452, 9471, 9561, 9584, 9830; Positive —1440, 1441, 1460, 1610, 1750, 1811, 1920, 2040, 2050, 2057, 2070, 2080, 2150, 2154, 2160, 2165, 2170, 2260, 2311, 2332, 2340, 2530, 2540, 2550, 2660, 5600, 5780, 5830, 5831, 7580; Neutral —2038, 2880, 5130, 5510, 5520, 5530, 6150, 7000, 7002, 7004, 7006, 7010, 7020, 7034, 7035, 7041, 7050, 7053, 7055, 7056, 7059, 7080, 7090, 7179, 7187, 7217, 7233, 7235, 7490, 7950.
2For example, EW—‘Paul loved to watch all TV shows that were set in hospitals. He has always wanted to be a doctor’; WCV—‘Louise was suffering from a toothache for several days, but she still refused to do anything about it. She has always been afraid of going to the doctor’; BCV—‘Michaela loved to read and she could not wait to check out a new set of books. She knew they were set aside for her by the friendly docto’.
References
- Ashby FG, Isen AM, Turken AU. A neuropsychological theory of positive affect and its influence on cognition. Psychological Review. 1999;106(3):529–50. doi: 10.1037/0033-295x.106.3.529. [DOI] [PubMed] [Google Scholar]
- Bauml KH, Kuhbandner C. Remembering can cause forgetting–but not in negative moods. Psychological Science. 2007;18(2):111–15. doi: 10.1111/j.1467-9280.2007.01857.x. [DOI] [PubMed] [Google Scholar]
- Bohner G, Chaiken S, Hunyadi P. The role of mood and message ambiguity in the interplay of heuristic and systematic processing. European Journal of Social Psychology. 1994;24:207–21. [Google Scholar]
- Bolte A, Goschke T, Kuhl J. Emotion and intuition: effects of positive and negative mood on implicit judgments of semantic coherence. Psychological Science. 2003;14(5):416–21. doi: 10.1111/1467-9280.01456. [DOI] [PubMed] [Google Scholar]
- Bower GH. Mood and Memory. American Psychologist. 1981;36(2):129–48. doi: 10.1037//0003-066x.36.2.129. [DOI] [PubMed] [Google Scholar]
- Bradley MM, Lang PJ. Affective reactions to acoustic stimuli. Psychophysiology. 2000;37(2):204–15. [PubMed] [Google Scholar]
- Canli T, Amin Z, Haas B, Omura K, Constable RT. A double dissociation between mood states and personality traits in the anterior cingulate. Behavioral Neuroscience. 2004;118(5):897–904. doi: 10.1037/0735-7044.118.5.897. [DOI] [PubMed] [Google Scholar]
- Chwilla DJ, Virgillito D, Vissers CT. The relationship of language and emotion: N400 support for an embodied view of language comprehension. Journal of Cognitive Neuroscience. 2011;23(9):2400–414. doi: 10.1162/jocn.2010.21578. [DOI] [PubMed] [Google Scholar]
- Collins AM, Loftus EF. A spreading-activation theory of semantic processing. Psychological Review. 1975;82(6):407–28. [Google Scholar]
- Coltheart M. The MRC psycholinguistics database. Quarterly Journal of Experimental Psychology. 1981;33A:497–505. [Google Scholar]
- Cuthbert BN, Schupp HT, Bradley MM, Birbaumer N, Lang PJ. Brain potentials in affective picture processing: covariation with autonomic arousal and affective report. Biological Psychology. 2000;52(2):95–111. doi: 10.1016/s0301-0511(99)00044-7. [DOI] [PubMed] [Google Scholar]
- Deacon D, Hewitt S, Yang C, Nagata M. Event-related potential indices of semantic priming using masked and unmasked words: evidence that the N400 does not reflect a post-lexical process. Cognitive Brain Research. 2000;9(2):137–46. doi: 10.1016/s0926-6410(99)00050-6. [DOI] [PubMed] [Google Scholar]
- Debruille JB. The N400 potential could index a semantic inhibition. Brain Research Reviews. 2007;56(2):472–77. doi: 10.1016/j.brainresrev.2007.10.001. [DOI] [PubMed] [Google Scholar]
- Demirtas-Tatlidede A, Freitas C, Pascual-Leone A, Schmahmann JD. Modulatory effects of theta burst stimulation on cerebellar nonsomatic functions. Cerebellum. 2011;10(3):495–503. doi: 10.1007/s12311-010-0230-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dreisbach G. How positive affect modulates cognitive control: the costs and benefits of reduced maintenance capability. Brain and Cognition. 2006;60(1):11–19. doi: 10.1016/j.bandc.2005.08.003. [DOI] [PubMed] [Google Scholar]
- Dreisbach G, Goschke T. How positive affect modulates cognitive control: reduced perseveration at the cost of increased distractibility. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2004;30(2):343–53. doi: 10.1037/0278-7393.30.2.343. [DOI] [PubMed] [Google Scholar]
- Dry MJ, Storms G. Features of graded category structure: Generalizing the family resemblance and polymorphous concept models. Acta Psychologica. 2010;133:244–55. doi: 10.1016/j.actpsy.2009.12.005. [DOI] [PubMed] [Google Scholar]
- Federmeier KD, Kirson DA, Moreno EM, Kutas M. Effects of transient, mild mood states on semantic memory organization and use: an event-related potential investigation in humans. Neuroscience Letters. 2001;305(3):149–52. doi: 10.1016/s0304-3940(01)01843-2. [DOI] [PubMed] [Google Scholar]
- Federmeier KD, Kutas M. Aging in context: age-related changes in context use during language comprehension. Psychophysiology. 2005;42(2):133–41. doi: 10.1111/j.1469-8986.2005.00274.x. [DOI] [PubMed] [Google Scholar]
- Federmeier KD, Kutas M. A rose by any other name: Long-term memory structure and sentence processing. Journal and Memory and Language. 1999a;41:469–95. [Google Scholar]
- Federmeier KD, Kutas M. Right words and left words: electrophysiological evidence for hemispheric differences in meaning processing. Cognitive Brain Research. 1999b;8(3):373–92. doi: 10.1016/s0926-6410(99)00036-1. [DOI] [PubMed] [Google Scholar]
- Fiedler K. Emotional mood, cognitive style, and behavioral regulation. In: Fiedler K, Forgas J, editors. Affect, Cognition and Social Behavior. Toronto: Hogrefe; 1988. [Google Scholar]
- Fiedler K. Affective states trigger processes of assimilation and accommodation. In: Martin LL, Clore GL, editors. Theories of Mood and Cognition: A User’s Guidebook. Mahwah, NJ: Lawrence Erlbaum Associates; 2001. pp. 85–98. [Google Scholar]
- First MB, Spitzer RL, Gibbon M, Williams JBM. Structured Clinical Interview for DSM-IV Axis II Personality Disorders (SCID-II, version 2.0) New York, NY: Biometrics Research Department, New York State Psychiatric Institute; 1995. [Google Scholar]
- First MB, Spitzer RL, Gibbon M, Williams JBW. Structured Clinical Interview for DSM-IV Axis I Diagnosis-Patient Edition (SCID-I/P, version 2.0) New York, NY: Biometric Research Department, New York State Psychiatric Institute; 2002. [Google Scholar]
- Forgas JP, Bower GH. Mood effects on person-perception judgments. Journal of Personality and Social Psychology. 1987;53(1):53–60. doi: 10.1037//0022-3514.53.1.53. [DOI] [PubMed] [Google Scholar]
- Fredrickson BL, Branigan C. Positive emotions broaden the scope of attention and thought-action repertoires. Cognition and Emotion. 2005;19:313–32. doi: 10.1080/02699930441000238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gratton G, Coles MG, Donchin E. A new method for off-line removal of ocular artifact. Electroencephalography and Clinical Neurophysiology. 1983;55:468–84. doi: 10.1016/0013-4694(83)90135-9. [DOI] [PubMed] [Google Scholar]
- Gray JR. Emotional modulation of cognitive control: approach-withdrawal states double-dissociate spatial from verbal two-back task performance. Journal of Experimental Psychology: General. 2001;130(3):436–52. doi: 10.1037//0096-3445.130.3.436. [DOI] [PubMed] [Google Scholar]
- Holcomb PJ. Semantic priming and stimulus degradation: implications for the role of the N400 in language processing. Psychophysiology. 1993;30(1):47–61. doi: 10.1111/j.1469-8986.1993.tb03204.x. [DOI] [PubMed] [Google Scholar]
- Hurlemann R, Hawellek B, Matusch A, et al. Noradrenergic modulation of emotion-induced forgetting and remembering. Journal of Neuroscience. 2005;25(27):6343–349. doi: 10.1523/JNEUROSCI.0228-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Isen AM, Daubman KA. The influence of affect on categorization. Journal of Personality and Social Psychology. 1984;47:1206–217. [Google Scholar]
- Isen AM, Daubman KA, Nowicki GP. Positive affect facilitates creative problem solving. Journal of Personality and Social Psychology. 1987;52(6):1122–131. doi: 10.1037//0022-3514.52.6.1122. [DOI] [PubMed] [Google Scholar]
- Isen AM, Johnson MMS, Mertz E, Robinson GF. The influence of positive affect on the unusualness of word associations. Journal of Personality and Social Psychology. 1985;48:1413–426. doi: 10.1037//0022-3514.48.6.1413. [DOI] [PubMed] [Google Scholar]
- Isen AM, Niedenthal P, Cantor N. The influence of positive affect on social categorization. Motivation and Emotion. 1992;16:65–78. [Google Scholar]
- Khateb A, Pegna AJ, Landis T, Mouthon MS, Annoni JM. On the origin of the N400 effects: an ERP waveform and source localization analysis in three matching tasks. Brain Topography. 2010;23(3):311–20. doi: 10.1007/s10548-010-0149-7. [DOI] [PubMed] [Google Scholar]
- Koch K, Pauly K, Kellermann T, et al. Gender differences in the cognitive control of emotion: An fMRI study. Neuropsychologia. 2007;45(12):2744–754. doi: 10.1016/j.neuropsychologia.2007.04.012. [DOI] [PubMed] [Google Scholar]
- Kutas M, Federmeier KD. Electrophysiology reveals semantic memory use in language comprehension. Trends in Cognitive Science. 2000;4(12):463–470. doi: 10.1016/s1364-6613(00)01560-6. [DOI] [PubMed] [Google Scholar]
- Kutas M, Federmeier KD. Finding meaning in the N400 component of the event-related brain potential (ERP) Annual Review of Psychology. 2011;62:621–47. doi: 10.1146/annurev.psych.093008.131123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang PJ, Bradley MM, Cuthbert BN International Affective Picture System (IAPS) Technical Report A-8. Gainesville, FL: University of Florida; 2008. Affective Ratings of Pictures and Instruction Manual. [Google Scholar]
- Larson MJ, Perlstein WM, Stigge-Kaufman D, Kelly KG, Dotson VM. Affective context-induced modulation of the error-related negativity. Neuroreport. 2006;17(3):329–33. doi: 10.1097/01.wnr.0000199461.01542.db. [DOI] [PubMed] [Google Scholar]
- Maratos EJ, Allan K, Rugg MD. Recognition memory for emotionally negative and neutral words: an ERP study. Neuropsychologia. 2000;38(11):1452–465. doi: 10.1016/s0028-3932(00)00061-0. [DOI] [PubMed] [Google Scholar]
- McNair DM, Lorr M, Droppleman LF. Profile of Mood States. San Diego, CA: Educational & Industrial Testing Service; 1971/1981. [Google Scholar]
- McNeely HE, Dywan J, Segalowitz SJ. ERP indices of emotionality and semantic cohesiveness during recognition judgments. Psychophysiology. 2004;41(1):117–29. doi: 10.1111/j.1469-8986.2003.00137.x. [DOI] [PubMed] [Google Scholar]
- Mitchell RL, Phillips LH. The psychological, neurochemical and functional neuroanatomical mediators of the effects of positive and negative mood on executive functions. Neuropsychologia. 2007;45(4):617–29. doi: 10.1016/j.neuropsychologia.2006.06.030. [DOI] [PubMed] [Google Scholar]
- Niznikiewicz M, Mittal MS, Nestor PG, McCarley RW. Abnormal inhibitory processes in semantic networks in schizophrenia. International Journal of Psychophysiology. 2010;75(2):133–40. doi: 10.1016/j.ijpsycho.2009.10.006. [DOI] [PubMed] [Google Scholar]
- Niznikiewicz MA, O'Donnell BF, Nestor PG, et al. ERP assessment of visual and auditory language processing in schizophrenia. Journal of Abnormal Psychology. 1997;106(1):85–94. doi: 10.1037//0021-843x.106.1.85. [DOI] [PubMed] [Google Scholar]
- Oldfield RC. The assessment and analysis of handedness: The Edinburgh Inventory. Neuropsychologia. 1971;9:97–113. doi: 10.1016/0028-3932(71)90067-4. [DOI] [PubMed] [Google Scholar]
- Park J, Banaji MR. Mood and heuristics: The influence of happy and sad states on sensitivity and bias in stereotyping. Journal of Personality and Social Psychology. 2000;78:1005–023. doi: 10.1037//0022-3514.78.6.1005. [DOI] [PubMed] [Google Scholar]
- Paulmann S, Kotz SA. Early emotional prosody perception based on different speaker voices. Neuroreport. 2008;19(2):209–13. doi: 10.1097/WNR.0b013e3282f454db. [DOI] [PubMed] [Google Scholar]
- Phillips LH, Bull R, Adams E, Fraser L. Positive mood and executive functions: evidence from stroop and fluency tasks. Emotion. 2002;2:12–22. doi: 10.1037/1528-3542.2.1.12. [DOI] [PubMed] [Google Scholar]
- Rosch E, Mervis CB. Family resemblances - Studies in internal structure of categories. Cognitive Psychology. 1975;7(4):573–605. [Google Scholar]
- Rosch E, Mervis CB, Gray WD, Johnson DM, Boyes-Braem P. Basic objects in natural categories. Cognitive Psychology. 1976;8:382–439. [Google Scholar]
- Rowe G, Hirsh JB, Anderson AK. Positive affect increases the breadth of attentional selection. Proceedings of National Academy of Sciences U S A. 2007;104(1):383–88. doi: 10.1073/pnas.0605198104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schupp HT, Junghofer M, Weike AI, Hamm AO. Attention and emotion: an ERP analysis of facilitated emotional stimulus processing. Neuroreport. 2003;14(8):1107–110. doi: 10.1097/00001756-200306110-00002. [DOI] [PubMed] [Google Scholar]
- Simon-Thomas ER, Knight RT. Affective and cognitive modulation of performance monitoring: behavioral and ERP evidence. Cognitive, Affective, and Behavioral Neuroscience. 2005;5(3):362–72. doi: 10.3758/cabn.5.3.362. [DOI] [PubMed] [Google Scholar]
- Steele KM, Bass KE, Crook MD. The mystery of the Mozart effect: failure to replicate. Psychological Science. 1999;10(4):366–69. [Google Scholar]
- Storbeck J, Clore GL. With sadness comes accuracy; with happiness, false memory: mood and the false memory effect. Psychological Science. 2005;16(10):785–91. doi: 10.1111/j.1467-9280.2005.01615.x. [DOI] [PubMed] [Google Scholar]
- Vissers CT, Virgillito D, Fitzgerald DA, et al. The influence of mood on the processing of syntactic anomalies: evidence from P600. Neuropsychologia. 2010;48(12):3521–531. doi: 10.1016/j.neuropsychologia.2010.08.001. [DOI] [PubMed] [Google Scholar]
- Vosburg SK. The effect of positive and negative mood on divergent-thinking performance. Creativity Research Journal. 1998;11:165–72. [Google Scholar]
- Wang L, LaBar KS, McCarthy G. Mood alters amygdala activation to sad distractors during an attentional task. Biological Psychiatry. 2006;60(10):1139–146. doi: 10.1016/j.biopsych.2006.01.021. [DOI] [PubMed] [Google Scholar]
- Wechsler D. Wechsler Adult Intelligence Scale: Administration and Scoring Manual. 3rd edn. San Antonio, TX: The Psychological Corporation; 1997. [Google Scholar]
- Wilson MD. The MRC psycholinguistic database: machine readable dictionary, version 2. Behavior Research Methods, Instruments, & Computers. 1998;20:6–11. [Google Scholar]
- Wiswede D, Munte TF, Goschke T, Russeler J. Modulation of the error-related negativity by induction of short-term negative affect. Neuropsychologia. 2009;47(1):83–90. doi: 10.1016/j.neuropsychologia.2008.08.016. [DOI] [PubMed] [Google Scholar]