

Abstract
The high frequency of recurrence and poor survival rate of bladder cancer demand exploration of novel strategies. Gene therapy via adenovirus has shown promising potential for the treatment of tumors. We constructed a bladder cancer-specific adenovirus carrying E1A-androgen receptor (AR) under the control of UPII promoter and prostate stem cell antigen enhancer (PSCAE), designated as Ad/PSCAE/UPII/E1A-AR, and investigated its antitumor effects in vitro and in vivo. We demonstrated that Ad/PSCAE/UPII/E1A-AR could be selectively replicated in bladder tumor cell lines (5637, BIU87, EJ and T24) when compared with control adenovirus Ad/PSCAE/UPII/Luc. However, there was no evidence of cytotoxicity for normal human bladder cell line SV-HUC-1 and hepatoma cell line SMMC7721. AR agonist R1881 could strengthen the oncolytic effect of Ad/PSCAE/UPII/E1A-AR in bladder cancer cells. In addition, we demonstrated that intratumoral injection of Ad/PSCAE/UPII/E1A-AR into established subcutaneous human EJ tumors in nude mice could significantly regress the growth of tumor and markedly prolong survival for tumor-bearing mice; on the other hand, saline-treated tumors continued to grow rapidly. Our studies indicate that Ad/PSCAE/UPII/E1A-AR could effectively treat bladder cancer in vitro and in vivo. Furthermore, our findings provide a promising therapeutic modality for the treatment of bladder cancer.
Keywords: adenovirus, androgen receptor, bladder cancer
INTRODUCTION
In 2008, 68 810 new cases of bladder cancer were diagnosed and 14 100 deaths occurred in the United States.1 There is a high frequency of recurrence of noninvasive bladder cancer and a very poor 5-year survival rate for invasive bladder cancer,2 which makes the management of bladder cancer extremely challenging and the prognosis for patients with advanced disease dismal.3 Hence, there is urgent need to develop more effective novel treatment strategies to improve therapeutic outcome in patients with bladder cancer. Gene therapy is a promising and attractive strategy for cancer biotherapy.
However, the low specificity and efficacy of gene therapy is a hurdle yet to be overcome. In this study, we generated an oncolytic adenovirus that expresses fused E1A-androgen receptor (E1A-AR) controlled by human UPII promoter, and prostate stem cell antigen enhancer (PSCAE) was inserted upstream of the promoter to kill bladder tumor cells preferentially. E1A protein is the backbone of Ad5, and it encodes 243 and 289 amino-acid proteins; the former is involved in transcriptional activation and the latter in transcriptional repression. Besides, E1A protein could reprogram transcription in tumor cells.4 Hubberstey et al.5 reported that helper-dependent Ad vectors encoding E1A could reduce the tumorigenic potential of the cells and increase the cells’ sensitivity to anticancer drugs. In our study, 289 amino acids encoded by 13s mRNA were used to generate E1A-AR. Yang et al.6 reported that preferential expression of the 289 amino-acid protein in transgenic mice resulted in proapoptotic injury or in proinflammation in their lungs.
AR, 110kDa, a member of the nuclear receptor superfamily, regulates gene expression by interacting with androgen response elements correlated with androgen-regulated genes and functions as a transcription factor that mediates the biological effects of androgens in a ligand-dependent manner.7 Like other steroid receptors (glucocorticoid and mineralocorticoid), AR remains in the cytoplasm until it is activated by ligand binding.8 Researchers used some cell-based imaging models and found that the agonist initiates nuclear translocation9 and that AR may have a close relationship with androgen-insensitive syndromes and abnormalities in the male reproductive system.10,11 In addition, AR plays an important role in prostate cell differentiation, proliferation and survival. Studies indicate that androgen-independent prostate cancer is associated with AR gene mutations.12
Furthermore, AR has been detected in skeletal muscle, bone, adipose tissue and normal human bladder tissues,13 and it has a close relationship with the pathological tumor stage of bladder cancer.14 Studies focusing on the function of AR in androgen-dependent and androgen-independent prostate have found considerable success.15,16 Discovering the benefits of using androgen deprivation therapy and AR antagonists in bladder cancer has confirmed the essential role of AR in bladder cancer,17 and downregulation of AR expression could inhibit the growth of bladder tumor.18 This suggests that targeting AR in bladder tumors can be of tremendous importance.
Our previous studies have demonstrated that specific oncolytic adenoviruses have remarkable antitumor effect against prostate cancer19 and bladder cancer.20 Owing to this, AR is conceived to play an important role in bladder cancer growth; furthermore, E1A, as an anti-oncogene, could induce apoptosis, stimulate the G1 phase in bladder cancer cells, reverse the transformed morphology of cultured cancer cells and suppress the proliferation and transition of tumor cells. In addition, it could boost the susceptibility to chemotherapy and radiotherapy. Hence, we constructed an oncolytic adenovirus carrying E1A-AR (Ad/PSCAE/UPII/E1A-AR) to target AR-positive human bladder tumor. On the basis of previous experimental observations, we hypothesized that the antitumor efficacy of the oncolytic adenovirus could be enhanced by expression of chimeric protein E1A-AR, and we deduced that PSCA enhancer could further strengthen the activity of UPII promoter in bladder cancer. In this study, we demonstrated that E1A could inhibit the proliferation of bladder cancer cells and the growth of bladder tumor in nude mice.
However, to our knowledge, this is the first study to show the efficacy of oncolytic adenovirus carrying E1A-AR for the treatment of human bladder cancer. To address our hypothesis, we assessed cell selectivity and cytotoxicity in a series of human cancer cell lines and normal bladder cell lines in vitro and evaluated the antitumor effects of the bladder cancer-specific adenovirus carrying E1A-AR on established subcutaneous tumors.
RESULTS
Construction of recombinant adenovirus
Ad/PSCAE/UPII/E1A-AR is a conditionally replicative recombinant Ad5 virus in which the hUPII promoter controls the expression of the E1A gene, which is essential for virus replication (Figure 1a). PSCAE was inserted upstream of the UPII promoter, and Ad/PSCAE/UPII/E1A was constructed by the same method. In addition, Ad/PSCAE/UPII/Luc was used as a control. Human bladder cancer cells were infected with Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A and Ad/PSCAE/UPII/Luc at a multiplicity of infection (MOI) of 20 for 72 h, and mRNA was extracted for reverse-transcription-polymerase chain reaction (RT-PCR) to confirm the correct modification to the adenovirus genome. As Figure 1b shows, the human UPII promoter is 314bp, PSCA enhancer is 327 bp, E1A is 541bp and AR is 870bp, which verified the successful construction of Ad/PSCAE/UPII/E1A-AR and ensured it for further studies.
Figure 1.
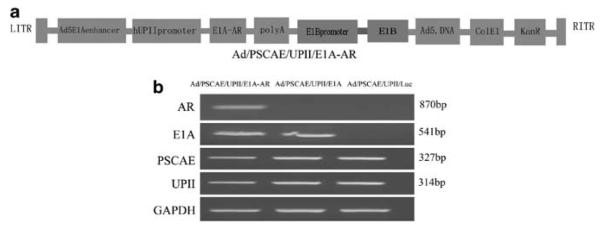
Construction and expression of Ad/PSCAE/UPII/E1A-AR. (a) Schematic diagram of the organization elements in the recombinant adenoviruses. Ad/PSCAE/UPII/E1A-AR is a conditionally replicative Ad5 virus, in which the human UPII promoter controls E1A gene expression. (b) RT-PCR analysis of AR, E1A, PSCAE and UPII expression. Human bladder cancer cell line EJ was treated with Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A or Ad/PSCAE/UPII/Luc at an MOI of 20 for 72 h. mRNA was extracted from cells for RT-PCR analysis. The primers used are described in Materials and Methods. Glyceraldehyde 3-phosphate dehydrogenase was used as the internal standard.
Luciferase activity assay in vitro
Higher Luc activity was detected in EJ, 5637 and BIU87 cells infected with Ad/PSCAE/UPII/Luc than in T24 (P<0.05) and SV-HUC-1 (P<0.01). However, much weaker Luc activity was detected in the normal bladder cell line SV-HUC-1 (Figure 2).
Figure 2.
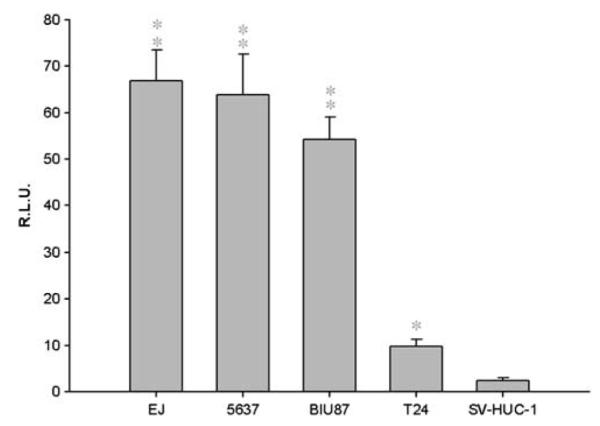
The detection of luciferase activity. Bladder cancer cell lines and normal bladder cell line SV-HUC-1 were infected with Ad/PSCAE/UPII/Luc at an MOI of 20 for 24 h, and luciferase activity was analyzed and represented as mean±s.d. **P<0.01 versus SV-HUC-1; *P<0.05 versus SV-HUC-1.
Ad/PSCAE/UPII/E1A-AR specifically inhibits the growth of bladder cancer cells and induced apoptosis in vitro
The cell viability of human bladder cancer cell lines (EJ, 5637, BIU-87 and T24), a normal bladder mucosa-derived cell line (SV-HUC-1) and a non-bladder cancer cell line (SMMC7721) after being infected with recombinant adenoviruses was depicted by a dose–response curve, which showed the increasing potency of adenovirus for spreading in tumor cells (Figure 3A). Compared with Ad/PSCAE/UPII/E1A and Ad/PSCAE/UPII/Luc, Ad/PSCAE/UPII/E1A-AR shows a more robust inhibition against bladder cancer cell lines EJ, 5637 or BIU87, and the inhibition effect is dose dependent. Although there is a much lower infection in T24, it is consistent with no evidence of CAR (Coxsackie virus-adenovirus receptor) expression and lower AR expression (Figures 4a and b). In addition, the cell viability for normal bladder cell line SV-HUC-1 and hepatoma cell line SMMC7721 did not change significantly after being treated with Ad/PSCAE/UPII/E1A-AR or Ad/PSCAE/UPII/E1A. However, Ad/PSCAE/UPII/Luc has no lytic activity in SV-HUC-1 and SMMC7721.
Figure 3.


Ad/PSCAE/UPII/E1A-AR inhibited the proliferation of bladder cancer cells and induced apoptosis in vitro. (A) Cell viability was examined after being treated with Ad/PSCAE/UPII/E1A-AR (◻), Ad/PSCAE/UPII/E1A (○) or Ad/PSCAE/UPII/Luc (▼) at an MOI of 0.1, 1, 10 and 100. Ad/CAE/UPII/E1A-AR suppressed the growth of bladder cancer cells, whereas it showed no inhibitory effect in normal bladder cells and non-bladder cancer cells. (B) Cell cycle was analyzed after treatment with Ad/PSCAE/UPII/E1A-AR or Ad/PSCAE/UPII/E1A using propidium iodide staining, and bladder cancer cells were infected with Ad/PSCAE/UPII/Luc as a control. (c) Quantitative assessment of the percentage of EJ, 5637, BIU87 and T24 cells in the sub-G1 phase. (D, E) Apoptotic cells were detected by transferase-mediated dUTP nick-end labeling. The number of apoptotic EJ, 5637 and BIU87 cells in the Ad/PSCAE/UPII/E1A-AR-treated group was significantly higher than that of T24 cells, and the number of apoptotic EJ, 5637 and BIU87 cells in the Ad/PSCAE/UPII/E1A-AR-treated group was significantly higher than that infected with Ad/PSCAE/UPII/E1A. Bladder cancer cells were infected with Ad/PSCAE/UPII/E1A-AR or Ad/PSCAE/UPII/E1A (20 MOI) for 72 h (× 100). (A, C) Ad/PSCAE/UPII/E1A-AR, (b, d) Ad/PSCAE/UPII/E1A and (e–h) control. (F) Caspase 3 activity was detected as described in Materials and methods, and the results are presented as mean±s.d. (G) Relative E1A gene expression levels of bladder cancer cells (BIU87, T24, 5637 and EJ) and normal bladder cells (SV-HUC-1) treated with Ad/PSCAE/UPII/E1A-AR were examined by semiquantitative RT-PCR with E1A-specific primers. The ΔΔCt method was used and glyceraldehyde 3-phosphate dehydrogenase was used for normalization. Results are presented as mean±s.d. **P<0.01 versus SV-HUC-1; *P<0.05 versus SV-HUC-1. (H) Virus particles in bladder cancer cells infected with Ad/PSCAE/UPII/E1A-AR or Ad/PSCAE/UPII/E1A (MOI = 10) for 72 h by TEM. (a) Bladder cancer cells without virus infection and (b) bladder cancer cells after infection with Ad/PSCAE/UPII/E1A-AR.
Figure 4.

CAR (a) and AR (b) protein expression levels in bladder cancer cells were determined by western blot. (a) The expression of CAR was detected in human bladder cancer cell lines 5637, T24, BIU-87 and EJ and in normal bladder cell line SV-HUC-1. Extracts from HEK293 were used as positive control, and β-actin was used as the internal standard. (b) The expression of AR was detected in human bladder cancer cell lines 5637, BIU-87, EJ and T24 and in normal bladder cell line SV-HUC-1. Extracts from LNCap, prostate cancer cell line, was used as positive control, and β-actin was used as the internal standard.
To explore whether apoptosis takes part in the process of cytotoxicity, cell cycle and caspase 3 activity were analyzed. Human bladder cancer cell lines were treated with recombinant adenoviruses before propidium iodide staining to analyze cell cycle by flow cytometry. As shown in Figures 3B and C, Ad/PSCAE/UPII/E1A-AR led to a significant increase in the number of cells in the G1 phase compared with Ad/PSCAE/UPII/Luc. Moreover, Ad/PSCAE/UPII/E1A-AR caused a high caspase 3 activity in bladder cancer cells in a time-dependent manner compared with SV-HUC-1 (P<0.05) (Figure 3F). More importantly, the number of apoptotic EJ, 5637 and BIU87 cells in the Ad/PSCAE/UPII/E1A-AR-treated group was much higher than that of T24 cells, and the number of apoptotic EJ, 5637 and BIU87 cells infected with Ad/PSCAE/UPII/E1A-AR was more than that infected with Ad/PSCAE/UPII/E1A (Figures 3D and F). The results of transmission electron microscopy (TEM) (Figure 3H) demonstrated that a mass of virus particles leading to, importantly, chromatin condensation and nucleoli fragmentation appeared in bladder cancer cells infected with Ad/PSCAE/UPII/E1A-AR, whereas bladder cancer cells without infection of virus showed no such signs of apoptosis. In addition, the expression level of E1A mRNA was detected by quantitative RT-PCR, and there is much higher expression of E1A mRNA in 5637 and BIU87 than in T24 (P<0.05) and SV-HUC-1 cells (P<0.01) (Figure 3G). The results further confirmed the fact that Ad/PSCAE/UPII/E1A-AR induced apoptosis in bladder tumor cells.
The expression of CAR and AR protein in bladder cancer cells by western blot analysis
To verify the expression level of AR and CAR in human bladder cancer cells, western blot analysis was carried out in this study. As shown in Figure 4a, there was a high expression of CAR in 5637, EJ and BIU-87 cells compared with T24 cells, which lacked detectable CAR protein. Human embryonic kidney 293 (HEK293) cells were used as a positive control. Figure 4b shows that the human bladder cancer cell lines 5637, EJ and BIU-87 have a higher expression of AR compared with T24 cells. LNCap, a prostate cancer cell line, was used as positive control.
The expression of E1A protein by western blot analysis and the inhibition effect of Ad/PSCAE/UPII/E1A-AR
Cell viability was examined as described in Materials and methods; the inhibition effect induced by Ad/PSCAE/UPII/E1A-AR was much higher for 5637, EJ and BIU-87 than for T24 and SV-HUC-1 (Figure 5a). However, flutamide has little impact on the effect of inhibition. To investigate the effect of AR agonist R1881 and AR antagonist flutamide on the activity of E1A protein, we analyzed the expression of E1A protein by western blot; there was a higher amount of E1A protein in 5637, EJ and BIU-87 cells infected with Ad/PSCAE/UPII/E1A-AR and a lower expression of E1A protein in 5637, EJ and BIU-87 cells treated with Ad/PSCAE/UPII/E1A (P<0.05). Furthermore, a weak expression of E1A was detected in T24 cells treated with both Ad/PSCAE/UPII/E1A and Ad/PSCAE/UPII/E1A-AR. As expected, E1A protein could not be detected in Ad/PSCAE/UPII/Luc-treated cells (Figure 5b). Meanwhile, the intensity of optical density of E1A protein was analyzed in Figure 5c. These observations indicate that R1881 enhanced higher expression of E1A in bladder cancer cell lines treated with Ad/PSCAE/UPII/E1A-AR in the presence of R1881, which could not be influenced by flutamide.
Figure 5.
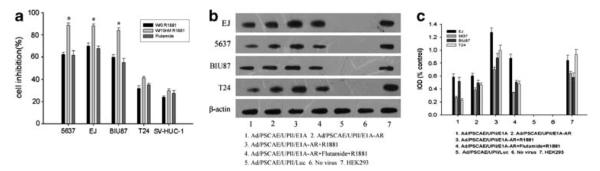
Expression of E1A protein and the effect of R1881 and flutamide on the activity of Ad/PSCAE/UPII/E1A-AR in bladder cancer cells. (a) R1881 significantly augmented the activity of Ad/PSCAE/UPII/E1A-AR in 5637, EJ and BIU87 cells, but not in T24 and SV-HUC-1 cells. Flutamide had no influence on the activity. Results are presented as mean±s.d. **P<0.01 versus the flutamide-treated group. (b) Expression of E1A protein by western blot analysis. Bladder cancer cells were treated with Ad/PSCAE/UPII/E1A or Ad/PSCAE/UPII/E1A-AR or Ad/PSCAE/UPII/Luc (20 MOI) for 72 h. Extracts from HEK293 were used as positive control, and β-actin was used as the internal standard. (c) Intensity of the optical density of E1A protein was analyzed.
Ad/PSCAE/UPII/E1A-AR suppressed EJ tumor growth and induced cell apoptosis in nude mice
To investigate the antitumor effect of Ad/PSCAE/UPII/E1A-AR in vivo, we established a xenograft bladder tumor model in nude mice by subcutaneous injection of bladder cancer cell line EJ. As shown in Figure 6A, compared with mice treated with saline or Ad/PSCAE/UPII/Luc, significant antitumor effect was observed in mice treated with Ad/PSCAE/UPII/E1A-AR (P<0.05). At the end of the experiment, the volume of tumor was 560 mm3 after intratumoral injection of Ad/PSCAE/UPII/E1A-AR, whereas the mean tumor volume was 2450 mm3 in the saline-treated group. This means that the bladder tumor growth was suppressed by 77%. Ad/PSCAE/UPII/E1A-AR could notably prolong the survival of tumor-bearing mice compared with Ad/PSCAE/UPII/Luc or saline as shown in Figure 6B (P<0.05). All the Ad/PSCAE/UP/E1A-AR-treated mice survived for more than 50 days, whereas all the control mice survived for o30 days. This was consistent with the morphology of tumors treated with Ad/PSCAE/UP/E1A-AR, as shown in Figure 6C.
Figure 6.

Ad/PSCAE/UPII/E1A-AR suppressed bladder tumor growth in vivo. (A) EJ cells were injected subcutaneously into nude mice, and 2 weeks later the tumor-bearing mice were treated with an intratumoral injection of Ad/PSCAE/UPII/E1A-AR (◻), Ad/PSCAE/UPII/E1A (○), Ad/PSCAE/UPII/Luc (▼) or saline (▽) every other day five times. At 28 days after virus injection, tumor-bearing mice were killed. Bars represent mean±s.d. (B) Survival curves for mice treated with Ad/PSCAE/UPII/E1A-AR (–), Ad/PSCAE/UPII/E1A (…), Ad/PSCAE/UPII/Luc (—) or saline (…). (C) Images of xenografts treated with recombinant adenovirus. (a) Ad/PSCAE/UPII/E1A-AR, (b) Ad/PSCAE/UPII/E1A, (c) Ad/PSCAE/UPII/Luc and (d) saline. Scale bar = 10 mm. (D) Hematoxylin and eosin staining of sections of tumor of the brain, liver and lung harvested from animals receiving intratumoral injection of Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A or saline. Arrows indicate the necrosis foci. (E) Particles arranged in the shape of a crystal lattice in tumor cells treated with Ad/PSCAE/UPII/E1A-AR under TEM ( × 30 000).
The tumors were excised and detected for the ability of viral replication. Hematoxylin and eosin staining revealed large areas of necrotic foci in the Ad/PSCAE/UPII/E1A-AR-treated group compared with the Ad/PSCAE/UPII/Luc- or saline-treated group (Figure 6D). As expected, there was no pathological change in the pathology sections of the liver, lung and brain extracted from mice treated with Ad/PSCAE/UPII/E1A-AR. Longer treatment period of mice by intramural injection was feasible. A mass of replicative adenovirus particles arranged in the shape of crystal lattices in the cytoplasm of bladder tumor cells was detected using TEM analysis (Figure 6E).
DISCUSSION
Conditionally replicative adenovirus (CRAD) is a promising therapy to combat cancer.21 Several reports have shown encouraging results using CRAD in vitro assay and also in animal models.22,23 In this study, we constructed an engineered adenovirus 5 type, Ad/PSCAE/UPII/E1A-AR, by inserting PSCAE into an engineered adenovirus Ad/UPII/E1A, which has shown potent antitumor activity for bladder cancer cells in our previous studies.20 Importantly, we replaced adenovirus E1A protein with chimeric protein E1A-AR, which has been verified in prostate cancer.19 Being of minimal toxicity to normal cells is critical for an oncolytic adenovirus. In our in vitro tests, we observed that almost 80% of bladder cancer cells were lysed even when infected with Ad/PSCAE/UPII/E1A-AR at an MOI of 20 in our in vitro tests, compared with no sign of cell killing observed in normal cell and non-bladder cancer cells infected with Ad/PSCAE/UPII/E1A-AR. We also revealed that the E1A gene is highly expressed in bladder cancer cells rather than in normal bladder cells. The luciferase assay in vitro revealed that Ad/PSCAE/UPII/Luc produced higher luciferase activity in bladder cancer cells than in SV-HUC-1, which further confirmed the fact that recombinant adenoviruses selectively replicate in bladder cancer cells.
The animal experiments suggest that Ad/PSCAE/UPII/E1A-AR replicated efficiently and induced marked cell killing in human bladder cancer cells; however, the replication and cytotoxicity were significantly attenuated in normal human bladder cells.
Genetically engineered viruses were used for selective replication in and for killing tumor cells, but sparing normal cells. This approach usually replaces the endogenous virus promoter sequence, for instance, E1A promoter, with a tissue-specific promoter.24 Numerous studies have shown promising therapeutic efficacy of the tissue promoter to control adenovirus E1A gene replication in midgut carcinoids, prostate cancer, ovarian cancer, colon cancer, hepatoma and osteosarcoma;22,23,25–29 in addition, some therapeutic agents using adenovirus vectors offer promise in bladder cancer gene therapy.30,31 UPII is a highly specific marker for human bladder cancer.32 In previous studies, we constructed a vector Rp-UPII-Luc and demonstrated that luciferase activity is much higher in bladder cancer than in other non-bladder cancers; that is, UPII promoter shows bladder tissue specificity.33 There is a broad perspective to treating bladder cancer by targeting UPII promoter and a powerful antitumor effect in vitro and in vivo.34–36
AR plays an important role in the development and growth of several cancers, including bladder cancer.37 Animal experiments have indicated that silencing AR by small interfering RNA could significantly repress bladder cancer progression.18 Although the expression of AR is downregulated on the basis of tumor grade and stage in bladder cancer,38 we found that fused protein E1A-AR could exhibit high activity in bladder cancer, regardless of the expression of AR. To address the role of AR in Ad/PSCAE/UPII/E1A-AR for the treatment of bladder cancer, AR agonist R1881 and antagonist flutamide were selected and western blotting was performed to examine the expression of E1A protein. The results indicated that R1881 could promote the replication and inhibition of Ad/PSCAE/UPII/E1A-AR, especially for AR-positive bladder cancer cells. However, when AR is blocked by flutamide, PSCAE could still show high activity, and it can be suggested that the activity of PSCAE is independent of androgen, consistent with our previous reports.33
Studies showed that the adenovirus death protein E3-11.6K is required for the efficient lysis of Ad-infected cells at very late stages of infection,39 whereas the primitive shutter vector used in our study is E3 deleted. The E3 region was found to be nonessential for viral replication.40 In addition, we used the Adeasy-1 system to generate recombinant adenoviruses. Importantly, in this study, we wanted to explore the function of favorite genes, such as E1A and AR, and investigate the antitumor effect of the CRAD constructed. In addition, there is an entire E1 region (E1A and E1B) in the vector, and UPII promoter and PSCA enhancer are also involved in the recombinant adenovirus we generated in our study; hence, it replicates not only in HEK293 but also in bladder cancer cell lines. Novel shutter vector involving the E3 region should be further studied to augment the oncolytic activity of adenovirus for the treatment of bladder cancer.
There are some differences in the inhibition of oncolytic adenovirus in different bladder cancer cell lines. These can be possibly explained by the differences in the expression of CAR or α-integrin receptors on different cancer cell lines. Immunohistochemistry observations suggest that decreased CAR expression has been associated with increased grade and stage of bladder cancer, and CAR has been found to be the primary cell surface receptor for adenovirus attachment during the infection process.41,42 Attempts have been made to enhance the oncolytic effect of viruses, and series studies have shown that CAR expression of bladder cancer cells could be markedly restored through the use of certain histone deacetylase inhibitors.43,44 Here, it should be mentioned that studies have also demonstrated that histone deacetylase inhibitor inhibits adenovirus replication.45 In our experiment, resistance to Ad/PSCAE/UPII/E1A-AR in T24 cells has a close relationship with poor infectivity and lack of CAR, which is consistent with a previous report.46 Some strategies of modifying adenovirus tropism have been explored to enhance the infectivity and specificity for CAR-deficient cell lines, such as chimeric Ad3/5 adenovirus,47,48 and studies have shown that CRADs expressing their own targeting adapter molecule are promising agents for cancer treatment.49 Novel strategies for the treatment of invasive bladder cancer should be further studied to augment the oncolytic activity of adenovirus.
Taking advantage of the anatomy of the bladder, intravesical instillation via the urethra using an oncolytic adenovirus is feasible and promising for superficial bladder cancer. Furthermore, complications could be reduced to a great extent compared with bacillus Calmette–Guerin or cytotoxic agents.50 Studies of intravesical administration of virus validated this view in preclinical trails and in animal experiments.31,51,52 Some phase I clinical trails have shown the safety and feasibility of intravesical instillation of adenovirus vectors.53,54 Published studies have examined the biodistribution following tail vein injection of adenovirus into immunocompetent mice or intraprostatic administration in mice.55,56 Pretreatment of the bladder with dodecyl-β-d-maltoside or sodium dodecyl sulfate could enhance adenovirus infection in a rat model.57
We also observed that Ad/PSCAE/UPII/E1A-AR could induce apoptosis, which was confirmed by the augmentation of caspase-3 activity, appearance of apoptotic cells, lattice-like bodies and appearance of a swollen endoplasmic reticulum by TEM. A previous study, including our study, revealed that the release and dissemination of virus particles of infected cells could be promoted by apoptosis.58
Although the antitumor effects of this strategy have been shown in our experiments, the potency is inadequate for monotherapy. The synergic therapeutic effect of adenovirus combined with radiation therapy or chemotherapy in cancers has been reported recently.59–62 Indeed, in our laboratory, continued experiments are being focused on the combination therapy of adenovirus gene therapy and radiation therapy or chemotherapy in in vitro tests (results not published). In addition, we have assessed the safety of oncolytic adenovirus in animals and did not find any toxic effects after intratumor administration. It should be pointed out that a subcutaneous bladder tumor model was used in our study; hence, a further study should be conducted to examine the efficacy and safety of the constructed adenoviruses in an orthotopic bladder tumor model.
In conclusion, our data showed that the constructed Ad/PSCAE/UPII/E1A-AR has a robust effect in the inhibition of proliferation of bladder cancer cells in vitro and in a marked regression of established bladder tumors in vivo. The results suggest the potential antitumor possibility of Ad/PSCAE/UPII/E1A-AR for clinical bladder cancer therapy.
MATERIALS AND METHODS
Cell culture
All cell lines were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). Human bladder transitional cell cancer cell lines (T24, 5637, EJ and BIU-87), human prostate cancer cell line (LNCap) and human hepatoma cell line (SMMC-7721) were cultured in RMPI1640 medium, whereas HEK293 cells were cultured in Dulbecco’s modified Eagle’s medium (Invitrogen, Grand Island, NY, USA), and normal human urinary cell line (SV-HUC-1) was cultured in Dulbecco’s modified Eagle’s medium/F12 (Invitrogen). The medium was supplemented with 10% fetal bovine serum, streptomycin (100 U ml−1) and penicillin (100 U ml−1). All cell lines were incubated in a humidified 37II incubator with 5% CO2 in air.
Generation of recombinant adenoviruses
We digested the vector RpS-TOAD-PSE-PBN-E1A to obtain the E1A fragment, after which we put a multiple cloning cassette into RpS-TOAD, which generated RpS-FROG. Thereafter, we inserted E1A into RpS-FROG to produce pCRAd-1. Human UPII promoter (312 bp) and human PSCA enhancer (327 bp) were amplified from human genomic DNA, and the primers used were as follows: UPII promoter forward, 5′-CGTAGCTAGCTGGCAACTTCAAGTGTGGGC-3′ and UPII promoter reverse, 5′-GGCAGGATCCT GGGAGGTGGAATAGGTGCT-3′; PSCA enhancer forward, 5′-GCTGACCGGT AGAGGCCAGCAGCACCCCTG-3′ and PSCA enhancer reverse, 5′-CGGTGCTA GCAACTGCTTCCGTGTGTGGCTGACAG-3′. The PCR products were cloned into pCRAd-1. We then digested RpS-TOAD-PSE-PBN-E1A-AR C685Y with BamHI/EcoRV and inserted E1A-ARC85Y into the vector backbone, which created Rp-PSCAE-UPII-E1A-AR (C685Y). Ad/PSCAE/UPII/E1A-AR was obtained through homologous recombination of Rp-PSCAE-UPII-E1A-AR (C685Y) and pAdEasy in Escherichia coli BJ5183 bacterial cells. We used similar strategies to generate Ad/PSCAE/UPII/E1A/Luc as a control. The recombination adenoviruses were packaged in HEK293 cells and purified by CsCl density gradient centrifugation. Extracellular versus intracellular virus titers were determined by TCID50.
Real-time-polymerase chain reaction
E1A gene expression was analyzed by RT-PCR, and cells were harvested after infection with adenovirus at an MOI of 10. Total RNA from different human cell lines was isolated by Trizol Reagent (Takara Biotechnology Co., Dalian, China) according to the manufacturer’s protocol. DNase was used to eliminate genomic DNA from RNA. cDNA was synthesized from 2 μg RNA using the PrimeScript RT reagent kit (Takara) according to the manufacturer’s protocol. Relative expression levels of E1A were assessed using the SYBR Premix Ex Taq kit (Takara). Primers for real-time PCR were as follows: E1A forward, 5′-CCCGAGTCTGTAATGTTGG-3′, E1A reverse, 5′-GTCGTCACTGGGTGGAAA-3′; and glyceraldehyde 3-phosphate dehydrogenase forward, 5′-GGATTTGGTCGTATTGGG-3′, glyceraldehyde 3-phosphate dehydrogenase reverse, 5′-GGAAGATGGTGATGGGATT-3′. PCR products were measured by rotor-gene 6000 (Roche, Shanghai, China), and we evaluated the data using the ΔΔCt method. Glyceraldehyde 3-phosphate dehydrogenase from each sample was used for normalization, and experiments were repeated three times.
Cell viability assay
Cell viability was examined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (Sigma, Ronkonkoma, NY, USA). Cells were plated on 96-well plates at 5 × 103 cells per well at 24 h before infection. Cells were infected with Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A or Ad/PSCAE/UPII/Luc at an MOI of 20 for 9 days; 20 μl 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (5 μg/ml) was added to each well and the plate was incubated at 37 1C for 4 h. Thereafter, the medium was removed before 150 μl dimethylsulfoxide was added to each well to solubilize the formazan crystals. In addition, bladder cancer cells were treated with Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A or Ad/PSCAE/UPII/Luc (20 MOI) for 24 h, followed by addition of fresh medium in the absence or presence of 10 nmol/l R1881 (a gift from John Hopkins University, Baltimore, MD, USA) or 10 nM flutamide for 48 h. Cell viability was analyzed using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. Plates were read at a wavelength of 490 nm using a Bioelisa Reader (EXL-800, BioTek Instruments, Inc., Winooski, VT, USA). The percentage of cell viability was calculated according to the formula 100% × (mean value A490 of infected cells)/(mean value A490 of uninfected cells).
Luciferase assay
Cells were plated on 6-well plates at 5 × 104 cells per well for 24 h and then infected with Ad/PSCAE/UPII/Luc at an MOI of 10. After 48 h, cells were extracted using the ONE-Glo Luciferase Assay System (Promega, Madison, WI, USA) according to the manufacturer’s protocol, using pCMV-β-gal as an internal control.
Flow cytometry analysis of the cell cycle
Human bladder cancer cells were seeded in 6-well plates at 5 × 104 cells per well and after 24 h were infected with Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A and Ad/PSCAE/UPII/Luc at an MOI of 10 for 48 h, respectively. Cells were trypsinized and washed once with phosphate-buffered saline and kept overnight at 4 1C in 70% ethanol, after which they were suspended with propidium iodide (50 μg ml−1; Sigma) and RNase A (100 μg ml−1). Cell cycle distribution was detected using flow cytometry (Beckman Coulter Epics XL, Ramsey, MN, USA).
Cytotoxicity assay
Cells were seeded in 96-well plates at 5 × 103 cells per well and infected with Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A and Ad/PSCAE/UPII/Luc at an MOI of 0.1, 1 and 10, respectively. After 96 h, the cells were fixed with 1% glutaraldehyde for 15 min, stained with 0.5% crystal violet (Sigma) for 15 min, washed with water for 10 min and dried overnight at room temperature. Sorenson’s solution (30 mM trisodium citrate, 0.06% HCl, 47.5% ethanol) was added to elute the crystal violet, and cytotoxicity was detected using a Bioelisa Reader (EXL-800) at 540 nm. Experiments were repeated at least three times.
Ultrastructural feature assays with TEM
To morphologically evaluate the selective replication of virus in cancer cells, bladder cancer cells were infected with Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A, Ad/PSCAE/UPII/Luc and phosphate-buffered saline for 72 h. The cells were fixed in 2.5% glutaraldehyde, postfixed in 1% osmium tetroxide, dehydrated using gradient ethanol and embedded in epoxy resin. Thereafter, 50-nm-thick ultrathin sections were cut, followed by staining with lead citrate and uranyl acetate. The sections were examined under JEM-1230 TEM (JOEL Ltd, Tokyo, Japan).
Western blot
The human bladder cancer cell lines and SV-HUC-1 were infected with or without Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A and Ad/PSCAE/UPII/Luc at 10 MOI in the presence or absence of 10 nmol l synthetic androgen R1881 and 10 nmol l flutamide and then harvested and lysed with RIPA supplemented with fresh protease inhibitor phenylmethanesulfonylfluoride on ice. Protein concentration was measured using a BCA Protein Assay Kit (Applygen Technologies Inc., Haidian, Beijing, China). Protein (20 μg) was boiled and loaded on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a PVDF membrane (Millipore, Bioprocess Technology Center, Billerica, MA, USA). The membranes were processed using the enhanced chemiluminescence detection system (TE 70 PWR Semi-Dry Transfer Unit, GE Healthcare, Piscataway, NJ, USA). Transfer efficiency was examined using a pre-stained protein marker. Equal loading and extract quality were manipulated by Coomassie staining.
Adenoviral E1A expression was detected using mouse monoclonal anti-Ad5 E1A (ab52523; Abcam, Cambridge, UK) as primary antibody, and coxsackie/adenovirus receptor and AR expression were detected via mouse monoclonal anti-CAR (05-644; Upstate Biotechnology, Lake Placid, NY, USA) or rat polyclonal anti-AR (PG-21; Millipore). Horseradish peroxidase-conjugated anti-mouse IgG or anti-rat IgG was used as a secondary antibody (Beijing Zhong Shan-Golden Bridge Biological Technology Co., Ltd, Beijing, China). Films were visualized using the enhanced chemiluminescence detection kit (Upstate Biotechnology).
Animal experiment
Male BALB/c nude mice, 4–6 weeks old, were purchased from the Animal Experimental Centre of Slaccas (Shanghai, China). All animals were housed under specific pathogen-free conditions, and experiments were carried out according to protocols approved by the Institutional Animal Care and Use Committee of Lanzhou University. A total of 5 × 106 EJ cells were subcutaneously incubated into the right flank of each mouse. When the volume of tumor reached 100–150 mm3, 40 mice were randomly divided into four groups. Ad/PSCAE/UPII/E1A-AR, Ad/PSCAE/UPII/E1A, Ad/PSCAE/UPII/Luc or saline was intratumorally injected at 5 × 108 PFU per mouse every other day five times. The tumor size was measured twice per week using a vernier caliper, and volume was calculated using the formula A × B2 × 0.5236, where A is the maximal length and B is the perpendicular width. Four weeks after the virus injection, all animals were killed by dislocation of the cervical vertebra, and the tumors were removed and weighed.
Histology of the xenografts was assessed using hematoxylin–eosin, and tumor tissues were weighed and fixed in 10% neutral formalin and finally embedded in paraffin. In all, 5 μm consecutive sections were obtained and treated with hematoxylin–eosin staining according to the manufacturer’s protocol. To further evaluate the selective replication of virus in tumor cells, tumor tissues were fixed in 2.5% glutaraldehyde, dehydrated using gradient ethanol, and embedded in epoxy resin. In all, 50 nm ultrathin sections were obtained and examined under JEM-1230 TEM (JOEL Ltd).
Statistical analysis
The results are presented as mean±s.d. One-way analysis of variance was used to determine the levels of difference between groups, and the Bonferroni test was used following the analysis of variance. Survival analysis was performed using the Kaplan–Meier method. The log-rank testwas used to analyze the survival rates. P-values <0.05 were considered statistically significant.
ACKNOWLEDGEMENTS
This work was supported by the National High-tech R&D Program of China (863 Program) (Grant No. 2008AA02Z421) and National Natural Science Foundation of China (Grant No. 30772177/H1619). We thank John Hopkins University for the gift of R1881, Xiang-dong He and De-gui Wang for helpful suggestions and comments, Hui-fang Zhang for carrying out the electron microscopic analysis (Lanzhou University Basic Medical College) and Jun Yan for carrying out the statistical analysis (Lanzhou University Basic Medical College).
Footnotes
CONFLICT OF INTEREST The authors declare no conflict of interest.
REFERENCES
- 1.Jemal A, Siegel R, Ward E, Hao Y, Xu J, Murray T, et al. Cancer statistics, 2008. CA Cancer J Clin. 2008;58:71–96. doi: 10.3322/CA.2007.0010. [DOI] [PubMed] [Google Scholar]
- 2.Voutsinas GE, Stravopodis DJ. Molecular targeting and gene delivery in bladder cancer therapy. J BUON. 2009;14(Suppl 1):S69–S78. [PubMed] [Google Scholar]
- 3.Kompier LC, Lurkin I, van der Aa MN, van Rhijn BW, van der Kwast TH, Zwarthoff EC. FGFR3, HRAS, KRAS, NRAS and PIK3CA mutations in bladder cancer and their potential as biomarkers for surveillance and therapy. PLoS One. 2010;5:e13821. doi: 10.1371/journal.pone.0013821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Frisch SM. Tumor suppression activity of adenovirus E1a protein: anoikis and the epithelial phenotype. Adv Cancer Res. 2001;80:39–49. doi: 10.1016/s0065-230x(01)80011-7. [DOI] [PubMed] [Google Scholar]
- 5.Hubberstey AV, Pavliv M, Parks RJ. Cancer therapy utilizing an adenoviral vector expressing only E1A. Cancer Gene Ther. 2002;9:321–329. doi: 10.1038/sj.cgt.7700436. [DOI] [PubMed] [Google Scholar]
- 6.Yang Y, McKerlie C, Borenstein SH, Lu Z, Schito M, Chamberlain JW, et al. Transgenic expression in mouse lung reveals distinct biological roles for the adenovirus type 5 E1A 243- and 289-amino-acid proteins. J Virol. 2002;76:8910–8919. doi: 10.1128/JVI.76.17.8910-8919.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Mangelsdorf DJ, Thummel C, Beato M, Herrlich P, Schutz G, Umesono K, et al. The nuclear receptor superfamily: the second decade. Cell. 1995;83:835–839. doi: 10.1016/0092-8674(95)90199-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lu ML, Schneider MC, Zheng Y, Zhang X, Richie JP. Caveolin-1 interacts with androgen receptor. A positive modulator of androgen receptor mediated transactivation. J Biol Chem. 2001;276:13442–13451. doi: 10.1074/jbc.M006598200. [DOI] [PubMed] [Google Scholar]
- 9.Marcelli M, Stenoien DL, Szafran AT, Simeoni S, Agoulnik IU, Weigel NL, et al. Quantifying effects of ligands on androgen receptor nuclear translocation, intranuclear dynamics, and solubility. J Cell Biochem. 2006;98:770–788. doi: 10.1002/jcb.20593. [DOI] [PubMed] [Google Scholar]
- 10.Tomura A, Goto K, Morinaga H, Nomura M, Okabe T, Yanase T, et al. The subnuclear three-dimensional image analysis of androgen receptor fused to green fluorescence protein. J Biol Chem. 2001;276:28395–28401. doi: 10.1074/jbc.M101755200. [DOI] [PubMed] [Google Scholar]
- 11.Szafran AT, Szwarc M, Marcelli M, Mancini MA. Androgen receptor functional analyses by high throughput imaging: determination of ligand, cell cycle, and mutation-specific effects. PLoS One. 2008;3:e3605. doi: 10.1371/journal.pone.0003605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Marcelli M, Ittmann M, Mariani S, Sutherland R, Nigam R, Murthy L, et al. Androgen receptor mutations in prostate cancer. Cancer Res. 2000;60:944–949. [PubMed] [Google Scholar]
- 13.Wilson CM, McPhaul MJ. A and B forms of the androgen receptor are expressed in a variety of human tissues. Mol Cell Endocrinol. 1996;120:51–57. doi: 10.1016/0303-7207(96)03819-1. [DOI] [PubMed] [Google Scholar]
- 14.Boorjian SA, Heemers HV, Frank I, Farmer SA, Schmidt LJ, Sebo TJ, et al. Expression and significance of androgen receptor coactivators in urothelial carcinoma of the bladder. Endocr Relat Cancer. 2009;16:123–137. doi: 10.1677/ERC-08-0124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Andersen RJ, Mawji NR, Wang J, Wang G, Haile S, Myung JK, et al. Regression of castrate-recurrent prostate cancer by a small-molecule inhibitor of the amino-terminus domain of the androgen receptor. Cancer Cell. 2010;17:535–546. doi: 10.1016/j.ccr.2010.04.027. [DOI] [PubMed] [Google Scholar]
- 16.Azuma K, Nakashiro K, Sasaki T, Goda H, Onodera J, Tanji N, et al. Anti-tumor effect of small interfering RNA targeting the androgen receptor in human androgen-independent prostate cancer cells. Biochem Biophys Res Commun. 2010;391:1075–1079. doi: 10.1016/j.bbrc.2009.12.024. [DOI] [PubMed] [Google Scholar]
- 17.Miyamoto H, Yang Z, Chen YT, Ishiguro H, Uemura H, Kubota Y, et al. Promotion of bladder cancer development and progression by androgen receptor signals. J Natl Cancer Inst. 2007;99:558–568. doi: 10.1093/jnci/djk113. [DOI] [PubMed] [Google Scholar]
- 18.Wu JT, Han BM, Yu SQ, Wang HP, Xia SJ. Androgen receptor is a potential therapeutic target for bladder cancer. Urology. 2010;75:820–827. doi: 10.1016/j.urology.2009.10.041. [DOI] [PubMed] [Google Scholar]
- 19.Hoti N, Li Y, Chen CL, Chowdhury WH, Johns DC, Xia Q, et al. Androgen receptor attenuation of Ad5 replication: implications for the development of conditionally replication competent adenoviruses. Mol Ther. 2007;15:1495–1503. doi: 10.1038/sj.mt.6300223. [DOI] [PubMed] [Google Scholar]
- 20.He XD, Wang ZP, Wei HY, Zhou Q, Wang DG, Tian JQ, et al. Construction of urothelium-specific recombinant adenovirus and its inhibition in bladder cancer cell. Urol Int. 2009;82:209–213. doi: 10.1159/000200802. [DOI] [PubMed] [Google Scholar]
- 21.Halloran CM, Ghaneh P, Shore S, Greenhalf W, Zumstein L, Wilson D, et al. 5-Fluorouracil or gemcitabine combined with adenoviral-mediated reintroduction of p16INK4A greatly enhanced cytotoxicity in Panc-1 pancreatic adenocarcinoma cells. J Gene Med. 2004;6:514–525. doi: 10.1002/jgm.540. [DOI] [PubMed] [Google Scholar]
- 22.Leja J, Dzojic H, Gustafson E, Oberg K, Giandomenico V, Essand M. A novel chromogranin–a promoter-driven oncolytic adenovirus for midgut carcinoid therapy. Clin Cancer Res. 2007;13:2455–2462. doi: 10.1158/1078-0432.CCR-06-2532. [DOI] [PubMed] [Google Scholar]
- 23.Xie X, Hsu JL, Choi MG, Xia W, Yamaguchi H, Chen CT, et al. A novel hTERT promoter-driven E1A therapeutic for ovarian cancer. Mol Cancer Ther. 2009;8:2375–2382. doi: 10.1158/1535-7163.MCT-09-0056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Campos SK, Barry MA. Current advances and future challenges in adenoviral vector biology and targeting. Curr Gene Ther. 2007;7:189–204. doi: 10.2174/156652307780859062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Rodriguez R, Schuur ER, Lim HY, Henderson GA, Simons JW, Henderson DR. Prostate attenuated replication competent adenovirus (ARCA) CN706: a selective cytotoxic for prostate-specific antigen-positive prostate cancer cells. Cancer Res. 1997;57:2559–2563. [PubMed] [Google Scholar]
- 26.Takahashi M, Sato T, Sagawa T, Lu Y, Sato Y, Iyama S, et al. E1B-55K-deleted adenovirus expressing E1A-13S by AFP-enhancer/promoter is capable of highly specific replication in AFP-producing hepatocellular carcinoma and eradication of established tumor. Mol Ther. 2002;5:627–634. doi: 10.1006/mthe.2002.0589. [DOI] [PubMed] [Google Scholar]
- 27.Li Y, Chen Y, Dilley J, Arroyo T, Ko D, Working P, et al. Carcinoembryonic antigen-producing cell-specific oncolytic adenovirus, OV798, for colorectal cancer therapy. Mol Cancer Ther. 2003;2:1003–1009. [PubMed] [Google Scholar]
- 28.Li X, Jung C, Liu YH, Bae KH, Zhang YP, Zhang HJ, et al. Anti-tumor efficacy of a transcriptional replication-competent adenovirus, Ad-OC-E1a, for osteosarcoma pulmonary metastasis. J Gene Med. 2006;8:679–689. doi: 10.1002/jgm.904. [DOI] [PubMed] [Google Scholar]
- 29.Li Y, Kacka M, Thompson M, Hsieh JT, Koeneman KS. Conditionally replicating adenovirus therapy utilizing bone sialoprotein promoter (Ad-BSP-E1a) in an in vivo study of treating androgen-independent intraosseous prostate cancer. Urol Oncol. 2011;29:624–633. doi: 10.1016/j.urolonc.2009.08.012. [DOI] [PubMed] [Google Scholar]
- 30.Shirakawa T, Hamada K, Zhang Z, Okada H, Tagawa M, Kamidono S, et al. A cox-2 promoter-based replication-selective adenoviral vector to target the cox-2-expressing human bladder cancer cells. Clin Cancer Res. 2004;10:4342–4348. doi: 10.1158/1078-0432.CCR-03-0267. [DOI] [PubMed] [Google Scholar]
- 31.Ramesh N, Ge Y, Ennist DL, Zhu M, Mina M, Ganesh S, et al. CG0070, a conditionally replicating granulocyte macrophage colony-stimulating factor–armed oncolytic adenovirus for the treatment of bladder cancer. Clin Cancer Res. 2006;12:305–313. doi: 10.1158/1078-0432.CCR-05-1059. [DOI] [PubMed] [Google Scholar]
- 32.Li SM, Zhang ZT, Chan S, McLenan O, Dixon C, Taneja S, et al. Detection of circulating uroplakin-positive cells in patients with transitional cell carcinoma of the bladder. J Urol. 1999;162:931–935. doi: 10.1097/00005392-199909010-00093. [DOI] [PubMed] [Google Scholar]
- 33.Wang D, Wang Z, Tian J, He X, Chowdhury WH, Zhang X, et al. Prostate stem cell antigen enhancer and uroplakin II promoter based bladder cancer targeted tissue-specific vector. Urol Oncol. 2010;28:164–169. doi: 10.1016/j.urolonc.2008.02.002. [DOI] [PubMed] [Google Scholar]
- 34.Zhang J, Ramesh N, Chen Y, Li Y, Dilley J, Working P, et al. Identification of human uroplakin II promoter and its use in the construction of CG8840, a urothelium-specific adenovirus variant that eliminates established bladder tumors in combination with docetaxel. Cancer Res. 2002;62:3743–3750. [PubMed] [Google Scholar]
- 35.Zhu H, Zhang ZA, Xu C, Huang G, Zeng X, Wei S, et al. Targeting gene expression of the mouse uroplakin II promoter to human bladder cells. Urol Res. 2003;31:17–21. doi: 10.1007/s00240-002-0294-x. [DOI] [PubMed] [Google Scholar]
- 36.Zhu HJ, Zhang ZQ, Zeng XF, Wei SS, Zhang ZW, Guo YL. Cloning and analysis of human uroplakin II promoter and its application for gene therapy in bladder cancer. Cancer Gene Ther. 2004;11:263–272. doi: 10.1038/sj.cgt.7700672. [DOI] [PubMed] [Google Scholar]
- 37.Liao X, Tang S, Thrasher JB, Griebling TL, Li B. Small-interfering RNA-induced androgen receptor silencing leads to apoptotic cell death in prostate cancer. Mol Cancer Ther. 2005;4:505–515. doi: 10.1158/1535-7163.MCT-04-0313. [DOI] [PubMed] [Google Scholar]
- 38.Boorjian S, Ugras S, Mongan NP, Gudas LJ, You X, Tickoo SK, et al. Androgen receptor expression is inversely correlated with pathologic tumor stage in bladder cancer. Urology. 2004;64:383–388. doi: 10.1016/j.urology.2004.03.025. [DOI] [PubMed] [Google Scholar]
- 39.Tollefson AE, Scaria A, Hermiston TW, Ryerse JS, Wold LJ, Wold WS. The adenovirus death protein (E3-11.6K) is required at very late stages of infection for efficient cell lysis and release of adenovirus from infected cells. J Virol. 1996;70:2296–2306. doi: 10.1128/jvi.70.4.2296-2306.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zhu M, Bristol JA, Xie Y, Mina M, Ji H, Forry-Schaudies S, et al. Linked tumor-selective virus replication and transgene expression from E3-containing oncolytic adenoviruses. J Virol. 2005;79:5455–5465. doi: 10.1128/JVI.79.9.5455-5465.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Sachs MD, Rauen KA, Ramamurthy M, Dodson JL, De Marzo AM, Putzi MJ, et al. Integrin alpha(v) and coxsackie adenovirus receptor expression in clinical bladder cancer. Urology. 2002;60:531–536. doi: 10.1016/s0090-4295(02)01748-x. [DOI] [PubMed] [Google Scholar]
- 42.Stoff-Khalili MA, Stoff A, Rivera AA, Mathis JM, Everts M, Wang M, et al. Gene transfer to carcinoma of the breast with fiber-modified adenoviral vectors in a tissue slice model system. Cancer Biol Ther. 2005;4:1203–1210. doi: 10.4161/cbt.4.11.2084. [DOI] [PubMed] [Google Scholar]
- 43.Okegawa T, Nutahara K, Pong RC, Higashihara E, Hsieh JT. Enhanced transgene expression in urothelial cancer gene therapy with histone deacetylase inhibitor. J Urol. 2005;174:747–752. doi: 10.1097/01.ju.0000164723.20555.e6. [DOI] [PubMed] [Google Scholar]
- 44.Sachs MD, Ramamurthy M, Poel H, Wickham TJ, Lamfers M, Gerritsen W, et al. Histone deacetylase inhibitors upregulate expression of the coxsackie adenovirus receptor (CAR) preferentially in bladder cancer cells. Cancer Gene Ther. 2004;11:477–486. doi: 10.1038/sj.cgt.7700726. [DOI] [PubMed] [Google Scholar]
- 45.Hoti N, Chowdhury W, Hsieh JT, Sachs MD, Lupold SE, Rodriguez R. Valproic acid, a histone deacetylase inhibitor, is an antagonist for oncolytic adenoviral gene therapy. Mol Ther. 2006;14:768–778. doi: 10.1016/j.ymthe.2006.07.009. [DOI] [PubMed] [Google Scholar]
- 46.El-Zawahry A, Lu P, White SJ, Voelkel-Johnson C. In vitro efficacy of AdTRAIL gene therapy of bladder cancer is enhanced by trichostatin A-mediated restoration of CAR expression and downregulation of cFLIP and Bcl-XL. Cancer Gene Ther. 2006;13:281–289. doi: 10.1038/sj.cgt.7700905. [DOI] [PubMed] [Google Scholar]
- 47.Haviv YS, Blackwell JL, Kanerva A, Nagi P, Krasnykh V, Dmitriev I, et al. Adenoviral gene therapy for renal cancer requires retargeting to alternative cellular receptors. Cancer Res. 2002;62:4273–4281. [PubMed] [Google Scholar]
- 48.Kanerva A, Wang M, Bauerschmitz GJ, Lam JT, Desmond RA, Bhoola SM, et al. Gene transfer to ovarian cancer versus normal tissues with fiber-modified adenoviruses. Mol Ther. 2002;5:695–704. doi: 10.1006/mthe.2002.0599. [DOI] [PubMed] [Google Scholar]
- 49.van Beusechem VW, Mastenbroek DC, van den Doel PB, Lamfers ML, Grill J, Wurdinger T, et al. Conditionally replicative adenovirus expressing a targeting adapter molecule exhibits enhanced oncolytic potency on CAR-deficient tumors. Gene Therapy. 2003;10:1982–1991. doi: 10.1038/sj.gt.3302103. [DOI] [PubMed] [Google Scholar]
- 50.Burke J. Virus therapy for bladder cancer. Cytokine Growth Factor Rev. 2010;21:99–102. doi: 10.1016/j.cytogfr.2010.02.003. [DOI] [PubMed] [Google Scholar]
- 51.Zhang X, Godbey WT. Preclinical evaluation of a gene therapy treatment for transitional cell carcinoma. Cancer Gene Ther. 2011;18:34–41. doi: 10.1038/cgt.2010.50. [DOI] [PubMed] [Google Scholar]
- 52.Tao Z, Connor RJ, Ashoori F, Dinney CP, Munsell M, Philopena JA, et al. Efficacy of a single intravesical treatment with Ad-IFN/Syn 3 is dependent on dose and urine IFN concentration obtained: implications for clinical investigation. Cancer Gene Ther. 2006;13:125–130. doi: 10.1038/sj.cgt.7700865. [DOI] [PubMed] [Google Scholar]
- 53.Kuball J, Wen SF, Leissner J, Atkins D, Meinhardt P, Quijano E, et al. Successful adenovirus-mediated wild-type p53 gene transfer in patients with bladder cancer by intravesical vector instillation. J Clin Oncol. 2002;20:957–965. doi: 10.1200/JCO.2002.20.4.957. [DOI] [PubMed] [Google Scholar]
- 54.Pagliaro LC, Keyhani A, Williams D, Woods D, Liu B, Perrotte P, et al. Repeated intravesical instillations of an adenoviral vector in patients with locally advanced bladder cancer: a phase I study of p53 gene therapy. J Clin Oncol. 2003;21:2247–2253. doi: 10.1200/JCO.2003.09.138. [DOI] [PubMed] [Google Scholar]
- 55.Wood M, Perrotte P, Onishi E, Harper ME, Dinney C, Pagliaro L, et al. Biodistribution of an adenoviral vector carrying the luciferase reporter gene following intravesical or intravenous administration to a mouse. Cancer Gene Ther. 1999;6:367–372. doi: 10.1038/sj.cgt.7700090. [DOI] [PubMed] [Google Scholar]
- 56.Paielli DL, Wing MS, Rogulski KR, Gilbert JD, Kolozsvary A, Kim JH, et al. Evaluation of the biodistribution, persistence, toxicity, and potential of germ-line transmission of a replication-competent human adenovirus following intraprostatic administration in the mouse. Mol Ther. 2000;1:263–274. doi: 10.1006/mthe.2000.0037. [DOI] [PubMed] [Google Scholar]
- 57.Ramesh N, Memarzadeh B, Ge Y, Frey D, VanRoey M, Rojas V, et al. Identification of pretreatment agents to enhance adenovirus infection of bladder epithelium. Mol Ther. 2004;10:697–705. doi: 10.1016/j.ymthe.2004.07.002. [DOI] [PubMed] [Google Scholar]
- 58.Mi J, Li ZY, Ni S, Steinwaerder D, Lieber A. Induced apoptosis supports spread of adenovirus vectors in tumors. Hum Gene Ther. 2001;12:1343–1352. doi: 10.1089/104303401750270995. [DOI] [PubMed] [Google Scholar]
- 59.Liu C, Zhang Y, Liu MM, Zhou H, Chowdhury W, Lupold SE, et al. Evaluation of continuous low dose rate versus acute single high dose rate radiation combined with oncolytic viral therapy for prostate cancer. Int J Radiat Biol. 2010;86:220–229. doi: 10.3109/09553000903419338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Onimaru M, Ohuchida K, Nagai E, Mizumoto K, Egami T, Cui L, et al. Combination with low-dose gemcitabine and hTERT-promoter-dependent conditionally replicative adenovirus enhances cytotoxicity through their crosstalk mechanisms in pancreatic cancer. Cancer Lett. 2010;294:178–186. doi: 10.1016/j.canlet.2010.01.034. [DOI] [PubMed] [Google Scholar]
- 61.Wang W, Sima N, Kong D, Luo A, Gao Q, Liao S, et al. Selective targeting of HPV-16 E6/E7 in cervical cancer cells with a potent oncolytic adenovirus and its enhanced effect with radiotherapy in vitro and vivo. Cancer Lett. 2010;291:67–75. doi: 10.1016/j.canlet.2009.09.022. [DOI] [PubMed] [Google Scholar]
- 62.Xie Y, Sheng W, Miao J, Xiang J, Yang J. Enhanced antitumor activity by combining an adenovirus harboring ING4 with cisplatin for hepatocarcinoma cells. Cancer Gene Ther. 2011;18:176–188. doi: 10.1038/cgt.2010.67. [DOI] [PMC free article] [PubMed] [Google Scholar]