

Abstract
Background
Factor VIII (FVIII) and von Willebrand factor (VWF) circulate in plasma in a tight non-covalent complex, being critical to hemostasis. Although structurally unrelated, both share the presence of sialylated glycan-structures, making them potential ligands for sialic-acid-binding-immunoglobulin-like-lectins (Siglecs).
Design and Methods
We explored the potential interaction between FVIII/VWF and Siglec-5, a receptor expressed in macrophages using various experimental approaches, including binding experiments with purified proteins and cell-binding studies with Siglec-5 expressing cells. Finally, Siglec-5 was overexpressed in mice via hydrodynamic gene transfer.
Results
In different systems using purified proteins, saturable, dose-dependent and reversible interactions between a soluble Siglec-5 fragment and both hemostatic proteins were found. Sialidase treatment of VWF resulted in a complete lack of Siglec-5 binding. In contrast, sialidase treatment left interactions between FVIII and Siglec-5 unaffected. FVIII and VWF also bound to cellsurface exposed Siglec-5, as was visualized by classical immunostaining as well as by Duolinkproximity ligation assays. Co-localization of FVIII and VWF with early endosomal markers further suggested that binding to Siglec-5 is followed by endocytosis of the proteins. Finally, overexpression of human Siglec-5 in murine hepatocytes following hydrodynamic gene transfer resulted in a significant decrease in plasma levels of FVIII and VWF in these mice.
Conclusions
Our data indicate that FVIII and VWF may act as a ligand for Siglec-5, and that Siglec-5 may contribute to the regulation of plasma levels of the FVIII/VWF complex.
Key words: factor VIII, von Willebrand factor, ligand, Siglec-5, interaction
Introduction
Coagulation factor VIII (FVIII) and von Willebrand factor (VWF) circulate in plasma in a tight non-covalent complex. Both proteins are essential elements of the hemostatic system, which is highlighted by the severe bleeding tendency that is associated with the functional deficiency of each protein, bleeding disorders known as hemophilia A and von Willebrand disease (VWD), respectively. FVIII and VWF are the products of two different genes, and their mature protein forms present in plasma display a distinct domain structure: A1-a1-A2-a2-B-a3-A3-C1-C2 for FVIII and D′-D3-A1-A2-A3-D4-B-C1-C2-CK for VWF.1,2 Another characteristic of FVIII and VWF is that they are both heavily decorated with carbohydrate-structures. The presence of these glycans is critical to the various steps in the lifecycle of both proteins, including biosynthesis/secretion, function and clearance.3
The predominant N-linked carbohydrate structure found on both FVIII and on VWF consists of a complextype biantennary core-fucosylated oligosaccharide, a structure that is commonly found on secreted proteins.4,5 In addition, tri- and tetra-antennary structures, as well as high mannose structures, have been identified.4,5 The Olinked glycans mainly consist of the sialylated T-antigen.3,6 Interestingly, FVIII and VWF molecules that are isolated from plasma are characterized by the presence of bloodgroup glycan structures. It has been estimated that approximately 10% of the N-linked glycans on FVIII (corresponding to 1-2 per molecule) and approximately 13% of the N-linked glycans on VWF (corresponding to 1-2 per subunit) contain ABO-determinants.4,5 Recently, also the O-linked glycans on VWF have been found to carry blood-group structures, albeit to a minor extent (approx. 1% corresponding to 1 per 10 subunits).6
As for many secreted glycoproteins, the vast majority (>80%) of the N- and O-linked carbohydrate structures are capped by sialic acids.4,5,7 Furthermore, approximately 25% of the O-linked T-antigens on VWF contain di-sialyl structures, indicating that terminal galactose or N-acetylgalactoside residues are capped with two rather than one sialic acid.6 The presence of sialic acids in the glycomes of FVIII and VWF makes both proteins potential ligands for a family of sialic-acid recognizing receptors: sialic-acid binding immunoglobulin-like lectins (Siglecs).
The human proteome contains 14 different Siglecs, each of which displays its own preference for the various sialic acid structures and conformations.8 Siglecs can be divided into two different subfamilies: CD22-related and CD33-related Siglecs. CD22-related Siglecs encompass four different Siglecs (including the archetype of this family, Sialoadhesin, now named Siglec-1), which are relatively well conserved between species. The human CD33-related Siglec subfamily contains 10 different members, which are poorly conserved in other species.9 Siglecs are selectively expressed in cells of hematopoietic origin, such as neutrophils, B cells, monocytes, dendritic cells and macrophages. However, the expression of each Siglec is restricted to a limited number of cells.9 For instance, expression of the CD33-related Siglec-5 includes monocytes/macrophages, neutrophils, and B cells but not T cells or NK cells.9 Siglec-5 is also weakly expressed on monocytic cell lines such as THP-1 and U937.10 The cloning of Siglec-5 was first reported in 1998 by Cornish et al. who isolated a full-length cDNA encoding Siglec-5 from a human activated monocyte cDNA library.10 Siglec-5 consists of 4 extracellular immunoglobulin-like domains and a single transmembrane domain that links the extracellular part to a cytoplasmic tail.10 The binding site for sialic acid is located in the N-terminal V-set domain.10,11 Compared to other Siglecs, Siglec-5 displays the least linkage specificity in sialic acid recognition, and is able to bind sialic acids in their α2-3, α2-6 and α2-8 linkage conformation.12 In addition, the two most common mammalian sialic acid variants (N-acetylneuraminic acid and N-glycolylneuraminic acid) are recognized by Siglec-5.12 Interestingly, Siglec-5 was recently found to interact with a bacterial protein in a non-sialic acid dependent manner, suggesting that ligand binding is not restricted per se to glycan-mediated interactions.13 The physiological function of Siglec-5 seems to be related to three different tasks: cell-cell interactions, signaling and endocytosis of its ligands.8,10,14-17
We explored the potential of Siglec-5 to interact with FVIII and/or VWF. There were three reasons to do so: 1) Siglec-5 is expressed on macrophages, a cell type that we have found to be dominant in the clearance of the FVIII/VWF complex;18 2) both FVIII and VWF contain sialic acid-capped glycans that (given the broad specificity of Siglec-5) could be recognized by this receptor; 3) preliminary experiments revealed that purified soluble Siglec-5 could bind efficiently to FVIII and VWF. In the present study, we used different experimental approaches to explore the interaction between Siglec-5 and these proteins. We found that both FVIII and VWF can bind to Siglec-5. Furthermore, overexpression of Siglec-5 in murine hepatocytes was associated with reduced plasma levels of FVIII and VWF, indicating that Siglec-5 contributes to the catabolism of the FVIII/VWF complex.
Design and Methods
Mice
Wild-type mice C57Bl/6 were purchased from Janvier (Le Genest Saint Isle, France). Housing and experiments were managed as recommended by French regulations and the experimental guidelines of the European Union. The animal experiments were approved by the Animal Care and Use Committee of INSERM.
Siglec-5 expressing cells
The cDNA encoding full-length human Siglec-5 was assembled synthetically (GeneArt, Regensburg, Germany) and cloned into pcDNA3.1. This plasmid was used to transfect human HEK293 cells, and after selection with geneticin (0.5 mg/mL) surviving cells were tested for the cell-surface expression of Siglec-5. To do this, cells were incubated with polyclonal goat-anti-Siglec-5 antibodies (R&D Systems, Minneapolis, MN, USA) and FITC-labeled rabbitanti-goat antibodies. Interestingly, these polyclonal anti-Siglec-5 antibodies display crossreaction with Siglec-14 and were, therefore, only used in systems in which Siglec-14 is absent. Positively stained clones were identified via flow cytometry, and expression was verified via immunostaining of the cells. Plasmid pcDNA3.1-Siglec-5 was used as a template for the generation of a construct encoding a protein consisting of soluble Siglec-5 fragments fused to an HPC4-tag (sSiglec-5/HPC4). Via polymerase chain reaction (PCR), the sequence encoding amino acids 1-434 (residues 1-16 represent signal peptide and residues 17-434 the ectodomain) was fused to a sequence encoding the HPC4-antibody recognition sequence (amino acid sequence EDQVDPRLIDGK), and cloned into pcDNA3.1 (designated pcDNA3.1-sSiglec-5/HPC4). HEK293 cells were transfected with pcDNA3.1-sSiglec-5/HPC4, and after selection with geneticin (0.5 mg/mL) surviving cells were tested for the release of sSiglec-5/HPC4 in the medium using an in-house ELISA for this protein. Primary monocytes were isolated from human blood via standard Ficoll gradient centrifugation.19 Following purification, adherent monocytes were stimulated with 100 nM phorbol myristate acetate (PMA) for 1 h at 37°C to induce a macrophage-like phenotype.
Proteins
Plasma-derived VWF was purified from therapeutic VWF-concentrates (Wilfactin, LFB Biomédicaments, Les Ulis, France) via size-exclusion chromatography (110 U VWF antigen/mg protein; FVIII: <0.1 U/mg protein). Plasma-derived FVIII was isolated from therapeutic FVIII concentrates (Factane, LFB Biomédicaments, Les Ulis, France) via immunoaffinity chromatography using VIII Select-affinity medium that was obtained from GE Healthcare, Auinay sous Bois, France (3500 U FVIII activity/mg protein; <0.01 U VWF/mg protein). Plasma-derived VWF and plasma-derived FVIII were used throughout the study, unless indicated otherwise. Refacto (Wyeth Pharmaceuticals, Paris, France) was used as a source for recombinant B-domainless FVIII (BDD-FVIII). Dimeric soluble Siglec-5 consisting of the extracellular region of Siglec-5 (residues 17-434) fused to Fc-portion of human IgG1 (designated sSiglec-5/Fc) was obtained from R&D Systems. Monomeric soluble Siglec-5 consisting of the extracellular region of Siglec-5 (residues 17-434) fused to the HPC4-antibody recognition sequence (designated sSiglec-5/HPC4) was purified from conditioned medium from stably transfected HEK293 cells expressing sSiglec-5/HPC4 via immunoaffinity chromatography employing antibody HPC4. Sialidase was purchased from Roche Diagnostics, Meylan, France. Bovine serum albumin (BSA) was from Sigma-Aldrich, Saint-Quentin Fallavier, France. FVIII and FX activity were measured using two-stage clotting assays, while FVIII and VWF antigen were measured using in-house immunosorbent assays. Normal pooled plasma was used as a reference.
Information on non-equilibrium binding assays, equilibrium binding assays, cellular binding of FVIII and VWF, hydrodynamic injection, immunostaining, Duolink-proximity ligation assay (Duolink-PLA) and microscopic imaging is available in the Online Supplementary Appendix.
Results
sSiglec-5/HPC4 binds to both FVIII and VWF in immunosorbent assays
In order to explore the potential interaction between Siglec-5 and the FVIII/VWF complex, we first analyzed binding of Siglec-5 to FVIII and VWF in a system using purified proteins. Highly purified FVIII and VWF were immobilized separately in microtiter-wells (2.5 μg/mL) and incubated with various concentrations of monomeric soluble Siglec-5 (sSiglec-5/HPC4; 0-1.3 μM). Bound sSiglec5/HPC4 was monitored using peroxidase-labeled monoclonal anti-HPC4 antibody. For both VWF and FVIII, a saturable and dose-dependent binding was observed (half-maximal binding being 0.54±0.20 and 0.48±0.21 μM, respectively; Figure 1), whereas no binding to BSA-coated control wells was observed. To rule out the contribution of potential trace contaminants in the VWF and FVIII preparations that could support Siglec-5 binding, we also tested binding of sSiglec-5/HPC4 to FVIII and VWF that were first caught via an immobilized antibody rather than being immobilized directly. A similar saturable and dose-dependent binding of sSiglec-5/HPC4 was observed, suggesting that sSiglec-5/HPC4 bound specifically to FVIII and VWF, respectively (data not shown). In addition, complementary experiments showed dose-dependent binding of FVIII or VWF to immobilized sSiglec-5/HPC4 (data not shown). To further assess the specificity of the interaction, binding was determined in the presence of anti-VWF, anti-FVIII or anti-Siglec-5 antibodies. As shown in Figure 1C, binding of sSiglec-5/HPC4 to FVIII was significantly reduced in the presence of anti-FVIII or anti-Siglec-5 antibodies, while binding to VWF was significantly reduced in the presence of anti-VWF or anti-Siglec-5 antibodies. To investigate the sialic acid dependence of the interactions, immobilized VWF or FVIII was incubated with sialidase (0.1 U/mL for 16 h at 37°C) prior to incubation with sSiglec-5/HPC4. Sialidase incubation resulted in an over 90% reduction in binding in case of VWF (Figure 1C). Surprisingly, no effect of sialidase treatment was observed for FVIII (Figure 1C). We considered the possibility that FVIII was not well de-sialylated, and tested the efficacy of sialidase-treatment via incubation with lectins ECL and WGA, which recognize terminal galactose and sialic acids, respectively. Sialidase-treatment resulted in increased ECL (82%) and reduced WGA (75%) binding, confirming that less sialic acids and increased numbers of galactose residues were exposed upon sialidase treatment. Using another approach, we compared plasma-derived full-length FVIII to recombinant BDD-FVIII, a variant that lacks 85% of the glycans present in full-length FVIII. No difference in binding of sSiglec5/Fc to anti-FVIII antibody immobilized FVIII variants was observed (Figure 1D). Half-maximal binding was calculated to be 79.0±26.2 nM and 69.8±15.5 nM for the binding of sSiglec-5/Fc to plasma-derived full-length FVIII and recombinant BDD-FVIII, respectively. These data suggest that binding of Siglec-5 to FVIII proceeds independently of sialic acids.
Figure 1.

Binding of soluble Siglec-5 to immobilized FVIII or VWF. (A and B) sSiglec-5/HPC4 (0-1.3 mM) was incubated with highly purified plasma-derived FVIII (A, closed circles), VWF (B, closed circles) or BSA (open circles) immobilized onto microtiter wells (all 2.5 μg/mL) in 0.15 M NaCl, 5 mM CaCl2, 0.1% Tween-20, 0.5% PVP, 25 mM Tris-HCl (pH 7.4). After 3 h of incubation, bound sSiglec-5/HPC4 was probed with peroxidase-labeled antibody HPC4 for 2 h at 37°C and detected by peroxidase hydrolysis of TMB. Presented is the absorbance at 450 nm versus sSiglec-5/HPC4 concentration. (C) Binding of sSiglec-5/HPC4 to immobilized FVIII or VWF was performed in the absence or presence of anti-FVIII, anti-VWF or anti-Siglec-5 antibodies. Alternatively, immobilized FVIII or VWF was incubated in the absence or presence of sialidase (0.1 U/mL) for 16 h at 37°C prior to incubation with Siglec-5/HPC4. Residual sSiglec-5/HPC4 binding compared to the binding in the absence of antibodies or sialidase is shown. (D) Plasma-derived full-length FVIII (closed circles) or recombinant BDD-FVIII (open circles) were adsorbed onto microtiter wells coated with monoclonal anti-FVIII antibody D4H1, and subsequently incubated with sSiglec-5/Fc (0-90 nM). Bound sSiglec-5/Fc was probed with peroxidase-labeled anti-human Fc antibody and detected via peroxidase hydrolysis of TMB. Shown is the absorbance at 450 nm versus sSiglec-5/Fc concentration. (E) VWF (2.5 μg/mL) was immobilized and incubated in the absence or presence of sialidase (0.1 U/mL) for 16 h at 37°C. Immobilized VWF was then incubated in the absence or presence of FVIII (5 μg/mlL for 1 h at 37°C. Finally, the wells were incubated with sSiglec-5/HPC4 (0.33 μM) and bound sSiglec-5/HPC4 was probed as described for panels A and B. The data present the mean±SEM of 3-5 independent experiments. The drawn lines (A,B and D) were obtained by fitting the data to an equation describing the binding of soluble Siglec-5 to a single class of binding sites, and were used to calculate half-maximal binding. **P<0.001.
VWF interferes with Siglec-5 binding to FVIII
Knowing that VWF is able to generally interfere with FVIII-receptor interactions,20 we were interested in testing the effect of VWF on FVIII-Siglec-5 interactions. To this end, VWF was immobilized and treated with sialidase to remove Siglec-5 binding sites. Sialidase-treatment did not affect FVIII binding (data not shown). The immobilized complex of FVIII and de-sialylated VWF was then incubated with sSiglec-5/HPC4. However, no binding of sSiglec-5/HPC4 to the de-sialylated VWF/FVIII complex could be detected (Figure 1E). In contrast, binding of sSiglec-5/HPC4 to the non-treated VWF/FVIII complex was unaffected when compared to non-treated VWF alone (Figure 1E). These data suggest that FVIII-bound to VWF is inaccessible for binding by Siglec-5.
Both VWF and FVIII bind to sSiglec-5/Fc in BLI-assays
Binding was further tested via BLI-assays using Octet-QK-equipment. Dimeric sSiglec-5/Fc was immobilized via protein A-coated biosensors. Subsequently, sSiglec-5/Fc-coated biosensors were incubated with various concentrations of VWF or FVIII. A saturable, dose-dependent and reversible binding of both proteins to sSiglec-5/Fc was observed (Online Supplementary Figure S1). Human IgG-coated control sensors did not identify binding (data not shown). Analysis of the binding data revealed that both proteins bound with relatively high affinity to sSiglec-5/Fc, with half-maximal binding values being 8.0±1.2 nM and 13.8±7.1 nM for VWF and FVIII, respectively (Online Supplementary Figure S1). Taken together, our results indicate that both FVIII and VWF are able to interact with the extracellular region of Siglec-5.
FVIII and VWF bind to Siglec-5 expressing cells
In a second series of tests, we examined whether FVIII and VWF are able to interact with Siglec-5 expressing cells. To this end, human HEK293 cells were stably transfected to express full-length Siglec-5. A Siglec-5-expressing clone was selected via flow-cytometric analysis using anti-human Siglec-5 antibodies (data not shown). Consistency of Siglec-5 expression was verified via immunofluorescence staining, which revealed that over 90% of the cells stained positive for Siglec-5 (data not shown). Siglec-5-expressing HEK293 cells and non-transfected control cells were grown on glass cover slips until 70-80% confluency. Cells were then incubated with purified VWF or FVIII (10 μg/mL) for 1 h at 4°C. After washing the cells, bound protein was monitored using anti-VWF or anti-FVIII antibodies. No staining for FVIII and VWF was observed when non-transfected HEK293 cells were used. In contrast, Siglec-5-expressing cells stained positive for VWF and for FVIII. Analysis of individual microscopic fields revealed that 23±16% (mean±SD; n=14) and 73±23% (n=25) of the Siglec-5 cells stained positive for VWF or FVIII, respectively. The relative fluorescence intensities were 846±266 RU/cell and 1731±109 RU/cell for VWF and FVIII, respectively. Given previous reports that part of cell-surface expressed Siglecs may be blocked by neighboring sialic-acid containing glycoproteins,8 we considered the possibility that binding of VWF and FVIII to cell-exposed Siglec-5 was suboptimal due to this inhibition. This was tested via pre-incubation of cells with sialidase in order to free Siglec-5 from potential inhibiting glycoproteins at the cell-surface. As for FVIII, binding appeared to be similar to non-treated and sialidase-treated Siglec-5 cells in terms of percentage positive cells (73±33% vs. 78±25%), even though sialidase-treatment resulted in slightly enhanced fluorescence intensity (1924±184 RU/cell; P<0.0001 compared to non-treated cells; Figure 2). No binding of FVIII to sialidase-treated non-transfected control cells could be detected (Figure 2). Similarly, sialidase treatment did not induce binding of VWF to non-transfected HEK293 cells (Figure 2). In contrast, binding of VWF to sialidase-treated Siglec-5 cells was remarkably more efficient. Not only the number of positively-stained cells was increased (74±33% vs. 23±16%), but also the intensity of positively-stained cells was increased compared to non-treated cells (1267±401 RU/cell; P<0.001 compared to non-treated cells; Figure 2). These data indicate that both FVIII and VWF are able to bind to cell-surface exposed Siglec-5. Furthermore, we observe for VWF that sialidase-treatment results in more positive cells and an increase in fluorescence intensity/cell, whereas for FVIII the number of positive cells remains unchanged with the intensity/cell being slightly increased.
Figure 2.

Binding of FVIII and VWF to Siglec-5 expressing cells. Non-transfected HEK293 cells and HEK293-Siglec-5 cells were grown on glass cover slips and incubated in the absence or presence of sialidase (0.1 U/mL for 1 h at 37°C) prior to incubation with FVIII or VWF (both 10 μg/mL) for 1 h at 4°C. After removing excess of unbound protein, cells were fixed by the addition of methanol. Bound FVIII or VWF was probed using mouse monoclonal antibodies and subsequently detected using AlexaFluor-488 conjugated F(Ab')2 fragments of goat-anti-mouse IgG. Cover slips were then embedded in DAPI-containing mounting medium. DAPI-stained nuclei are presented in blue, while FVIII or VWF are visualized in green. Images were collected using an AxioImager A1 microscope and a Plan-Apochromat 63x/NA 1.4 oil-immersion objective. For control, cells were incubated in the absence of VWF or FVIII.
FVIII and VWF co-localize with Siglec-5
To investigate whether binding of FVIII and VWF to Siglec-5-expressing cells involves Siglec-5, we performed Duolink-PLA analysis. This approach allows the detection of proteins that are co-localized within a radius of less than 40 nm, visualized as red spots via immunofluorescence microscopy. No red spots could be detected upon incubation of non-transfected cells in the presence of FVIII or VWF (Figure 3A). Also other controls (non-transfected cells in the absence of FVIII or VWF, or Siglec-5 expressing cells in the absence of FVIII or VWF) remained negative (data not shown). However, red spots representing co-localization of FVIII or VWF with Siglec-5 were selectively detected following incubation at 4°C of Siglec-5-expressing cells with FVIII or VWF (Figure 3A). Confocal analysis of positively-stained cells revealed that the co-localization was predominantly at the cell-surface, compatible with the absence of endocytosis at 4°C. These findings show that FVIII and VWF associate with the cell surface within a radius of 40 nm of Siglec-5, suggesting that both proteins may directly interact with this cell-surface exposed receptor.
Figure 3.
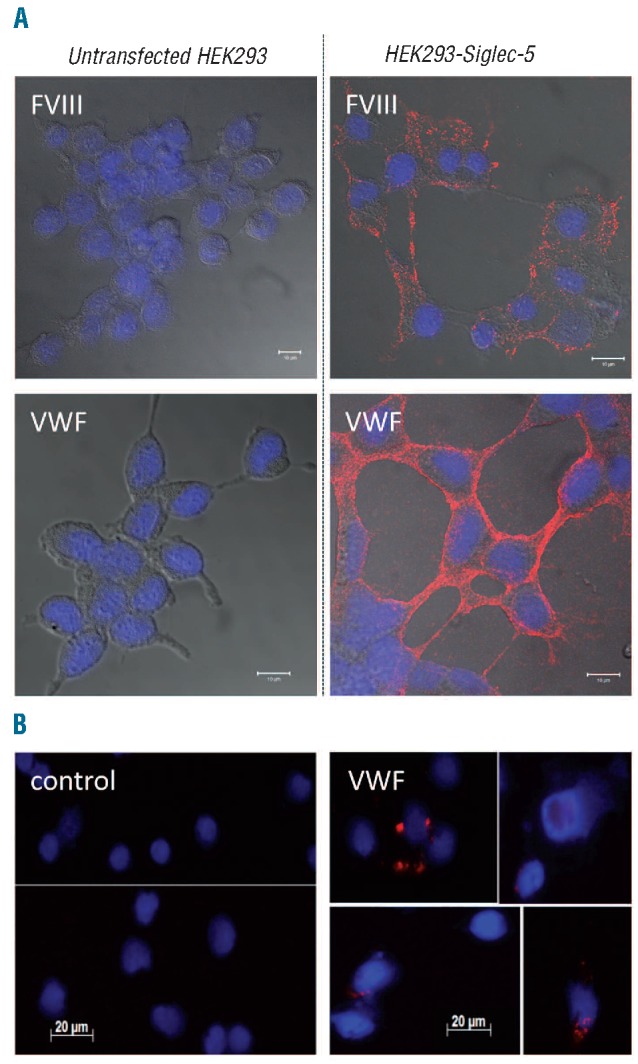
FVIII and VWF co-localize with Siglec-5. (A) Sialidase-treated HEK293-Siglec-5 or non-transfected cells were incubated with FVIII or VWF (10 μg/mL) for 1 h at 4°C. (B) Isolated human monocytes were stimulated with PMA (100 nM) for 1 h incubated in the absence or presence of VWF (10μg/mL) for 1 h at 4°C. (A and B) After removing excess of unbound protein, cells were fixed and incubated with a mixture of mouse monoclonal antibodies against FVIII or VWF in combination with polyclonal goat antibodies against Siglec-5. Bound antibodies were detected via Duolink-PLA analysis by the application of oligonucleotide-coupled secondary antibodies, the sequence of which was complementary. Following amplification, amplified oligonucleotides were highlighted using fluorescent-labeled probes, generating discrete red fluorescent spots for each FVIII/Siglec-5 or VWF/Siglec-5 complex. Cover slips were embedded in DAPI-containing mounting medium to allow blue nucleus staining. Cellular contours are visualized by merging in DIC-images. Confocal-images (A) were obtained with an Axiovert 200M microscope and a Zeiss LSM510-meta confocal system. Regular images (B) were obtained using an AxioImager A1 microscope. In both cases, a Plan-Apochromat 63x/NA 1.4-oil immersion objective was used.
Binding of VWF to Siglec-5 on PMA-stimulated primary monocytes
To test whether VWF is also able to bind to Siglec-5 on primary cells, monocytes were isolated from human blood and stimulated with PMA for 1 h to induce a macrophage-like phenotype. Cells were then incubated with VWF, and co-localization with endogenous Siglec-5 was assessed via Duolink-PLA analysis using anti-VWF and anti-Siglec-5 antibodies. Again, no red spots could be observed in PMA-stimulated monocytes that were incubated in the absence of VWF (Figure 3B). In contrast, multiple red spots were visible for about 20% of the PMA-stimulated monocytes. This suggests that VWF is able to co-localize with endogenous Siglec-5 at the surface of primary Siglec-5-expressing cells.
Siglec-5-bound FVIII and VWF are endocytosed into early endosomes
We then tested whether binding of FVIII and VWF to Siglec-5 expressing cells could result in endocytosis of both proteins. Following incubation of FVIII or VWF with Siglec-5-expressing cells at 4°C for 1 h, incubation was continued at 37°C potentially allowing internalization of the proteins. Subsequently, cells were stained using Duolink-PLA analysis with antibodies against FVIII or VWF and the early endosomal marker EEA-1. Using this approach, only FVIII or VWF molecules trapped in early endosomes will be detected, as they should be within 40 nm radius of the EEA-1 marker. No red spots could be detected in cells that were incubated at 4°C (Figure 4), or in cells that do not express Siglec-5 (data not shown). However, distinct red spots were observed in Siglec-5 expressing cells incubated at 37°C for both FVIII and VWF (Figure 4). Confocal analysis revealed that all of the spots where located inside the cell rather than at the cellular surface, confirming that FVIII and VWF were taken up by the cell.
Figure 4.
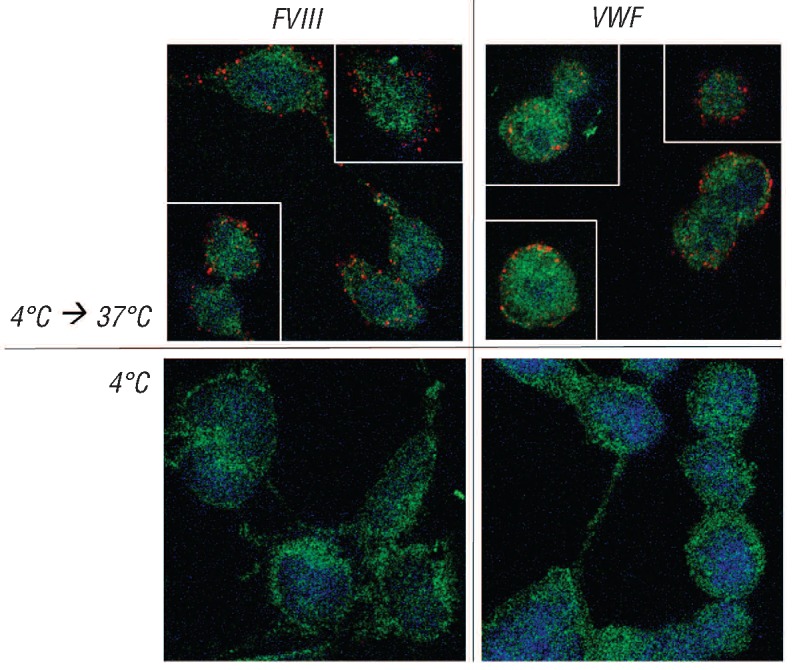
FVIII and VWF are targeted to early endosomes in Siglec-5 expressing cells. Sialidase-treated HEK293-Siglec-5 cells were incubated with FVIII or VWF (10μg/mL) for 1 h at 4°C. After removing excess of unbound protein, cells were put at 37°C for 30 min in case of FVIII and 15 min in case of VWF, allowing endocytosis of both proteins. Cells were then fixed and incubated with a mixture of antibodies against FVIII or VWF in combination with polyclonal rabbit antibodies against EEA1, a marker for early endosomes. Bound antibodies were detected via Duolink-PLA analysis as described in the legend of Figure 5. Red spots represent FVIII or VWF being within a radius of 40 nM of EEA1. Blue staining represents nuclei. Green staining represents auto-fluorescence of the cells, and is added to visualize the cellular contours.
In vivo expression of Siglec-5
Given the poor homology between human and murine Siglecs,9 we decided to express full-length human Siglec-5 in mice to study the interaction between this receptor and the FVIII/VWF complex in vivo. The full-length cDNA of Siglec-5 was subcloned in the pLIVE-expression vector. This plasmid contains a murine albumin promotor, allowing stable hepatic expression. The pLIVE-Siglec-5 plasmid or an empty pLIVE control vector were introduced via hydrodynamic gene transfer. Since Siglec-5 is not secreted as a soluble protein, livers were isolated four days after gene transfer in order to analyze for Siglec-5 expression. As shown in Figure 5A, no Siglec-5 could be detected in livers injected with empty pLIVE. In contrast, 20-30% of the liver hepatocytes stained positive for Siglec-5 following transfer with the pLIVE-Siglec-5 plasmid (Figure 5B). We also tested whether human Siglec-5 could interact with murine VWF. This was addressed in an immunosorbent-assay, in which murine plasma was incubated in wells coated with polyclonal anti-VWF antibodies. Plasma originated from normal C57B6-mice or from VWF-deficient mice. A specific and dose-dependent binding of sSiglec-5/HPC4 to wells incubated with VWF-containing plasma was observed (data not shown), suggesting that murine VWF is recognized by human Siglec-5.
Figure 5.
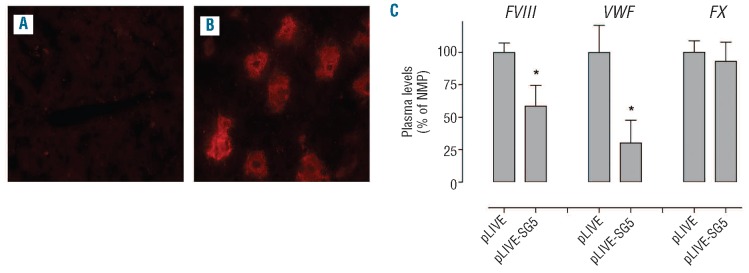
Effect of in vivo Siglec-5 expression of FVIII and VWF levels. C57B6 mice were injected hydrodynamically with an empty pLIVE plasmid of pLIVE-Siglec-5 (pLIVE-SG5), and after four days blood samples were taken to prepare plasma and livers were isolated. Liver sections of mice injected with pLIVE (A) or pLIVE-Siglec-5 (B) were analyzed for the expression of Siglec-5 using polyclonal anti-human Siglec-5 antibodies. Plasma samples were analyzed for levels of FVIII activity, VWF antigen and factor X (FX) activity (C). Levels of the various proteins in pLIVE-treated mice are relative to those in normal pooled plasma, which were arbitrarily set at 100%.
Hepatic Siglec-5 modulates plasma levels of the FVIII/VWF complex
Wild-type C57B6 mice were given an empty pLIVE-plasmid or pLIVE-Siglec-5 via hydrodynamic injection. Subsequently, plasma samples were taken from Siglec-5 expressing mice or control mice four days later, and analyzed for levels of FVIII, VWF and coagulation factor X. As shown in Figure 5C, plasma levels of factor X in Siglec-5 expressing mice were similar to those in control mice (93±15% vs. 100%; P>0.05). In contrast, levels of both FVIII and VWF were significantly lower in Siglec-5 positive mice compared to control mice (59±16% vs. 100%; P=0.015 for FVIII and 30±18% vs. 100%; P=0.011 for VWF; Figure 5C). This suggests that the presence of Siglec-5 in murine hepatocytes is associated with a downregulation of FVIII and VWF levels.
Discussion
Both FVIII and VWF are covered with sialylated glycans, making these proteins potential ligands for protein-receptors that recognize sialic acids, i.e. Siglecs. In the present study, we have found evidence that FVIII and VWF are able to interact with one of the members of the Siglec-family, Siglec-5. Our conclusion that Siglec-5 may function as a receptor for FVIII and VWF is based on the finding that purified FVIII and VWF specifically bind to recombinant soluble Siglec-5 variants in a dose-dependent and saturable manner (Figure 1 and Online Supplementary Figure S1). Furthermore, binding could be inhibited using antibodies against FVIII, VWF or Siglec-5 (Figure 1C). Also cellular binding studies using stable cell lines expressing Siglec-5 revealed that both FVIII and VWF are able to interact with cell-surface expressed Siglec-5 (Figures 2 and 3). Furthermore, Siglec-5 proved able to bind and transport both proteins to the intracellular early endosomes, indicating that Siglec-5 contributes to the endocytosis of FVIII and VWF (Figure 4). Finally, overexpression of Siglec-5 in mice resulted in a significant decrease in FVIII and VWF levels, suggesting that Siglec-5 contributes to the regulation of VWF/FVIII levels in vivo (Figure 5).
With regard to the specificity of their interactions, we considered that VWF and FVIII circulate in plasma in a tight non-covalent complex (Kd<1 nM). Therefore, it is possible that the purified plasma-derived protein preparations could contain traces of the other protein (VWF could be present in the FVIII preparations or vice versa). However, antigen-specific ELISAs were unable to detect such traces in the respective protein preparations (<1 ng FVIII/10 μg VWF and <8 ng VWF/10 μg FVIII). In addition, recombinant variants of FVIII and VWF (therefore lacking VWF and FVIII, respectively) displayed similar dose-dependent binding to Siglec-5, demonstrating that each protein individually can interact with this lectin.
Binding of soluble Siglec-5 to VWF was considerably reduced (>90%) upon pre-treatment of VWF with sialidase, suggesting that interactions are mediated by sialic acid moieties on the VWF molecule. In the immunosorbent assays, half-maximal binding was obtained at 0.5 μM of the monomeric sSiglec-5/HPC4. Half maximal binding was found to be 8 nM in our BLI-assays using a dimeric soluble Siglec-5 variant (Siglec-5/Fc). This 40-fold difference in half-maximal binding may be due to the use of different techniques (immunosorbent assays require washing steps, which could promote dissociation of the ligand-receptor complex). In addition, the use of an immobilized dimeric Siglec-5 variant results in a higher local concentration and higher avidity in comparison to the use of a monomeric variant in solution, and may, therefore, favor VWF binding. Interestingly, Siglec-5 is expressed at the cellular surface as a disulphide-linked homodimer10 indicating that the experiments with the dimeric Siglec-5/Fc variant seem to be of a more physiological relevance. We would like to emphasize that it is difficult, if not impossible, to determine a true affinity constant for this interaction, given that VWF is a heterogenous protein consisting of multiple covalently connected subunits. Interestingly, the multivalency of VWF may also contribute to the relatively efficient binding of Siglec-5, since a clustered surface of glycans can be presented to this receptor. Indeed, clustering of glycans promotes glycan-protein interactions21 that are often of relatively low affinity.22
FVIII also displayed efficient binding of soluble Siglec-5 variants: half-maximal binding being 0.5 μM in immunosorbent assays using monomeric sSiglec-5/HPC4 and 14 nM using dimeric Siglec-5/Fc in BLI-assays. This latter value is in the same order of magnitude as the apparent affinity for the interaction between FVIII and LDL-receptor related protein, a receptor known to affect FVIII plasma levels in vivo.23,24 This relatively high apparent affinity may be surprising in that FVIII (in contrast to VWF) is not a multimeric protein. The efficient interaction may be explained by the observation that sialidase-mediated de-sialylation of FVIII left binding of Siglec-5 unaffected (Figure 1C). This may point to the binding of FVIII to Siglec-5 being independent of low affinity sialic acid-Siglec interactions. Instead, our data suggest that Siglec-5 interacts with a protein portion of FVIII. In support of this view is our finding that both plasma-derived full-length FVIII and recombinant BDD-FVIII are similar in their interaction with Siglec-5 (Figure 1D), despite the fact that BDD-FVIII lacks 85% of the carbohydrate structures that are present in full-length FVIII.27 Although unusual, it is not without precedent that a protein interacts with Siglec-5 in a sialic acid-independent manner. Recently, it has been shown that the Group B streptococcus b-protein interacts with the aminoterminal V-set domain of Siglec-5 in a sialic acid-independent manner.13 The complementary interactive sites within FVIII remain to be determined.
Apart from their interaction with purified protein-fragments, FVIII and VWF also bound to stably transfected HEK293 cells expressing full-length Siglec-5. Again, VWF and FVIII behaved in a slightly different manner. For VWF, we observed that 23±16% of the Siglec-5 expressing cells stained positive for VWF following incubation at 4°C. Not only the number of positively stained cells increased to 74±33% when cells were pre-treated with sialidase, but also the fluorescence intensity was markedly increased (Figure 2). It has previously been suggested that the sialic acid binding pocket of cell-surface exposed Siglecs are often occupied by sialic acids that are present on neighboring cell surface proteins.8 It seems reasonable to assume that the presence of such sialylated proteins compete with VWF for binding to the sialic acid binding site of Siglec-5, explaining why increased binding of VWF is observed following de-sialylation of cell surface proteins. The number of positively-stained cells was not increased for FVIII, with the fluorescence intensity being increased, but to a minor extent (Figure 2). This suggests that for FVIII there is only a slight increase in the number of available Siglec-5 receptors/cell following sialidase treatment. It seems possible that this is due to FVIII being able to interact with Siglec-5 in a sialic acid-independent manner.
Cellular binding was also investigated via immunofluorescent confocal microscopy in combination with Duolink-PLA analysis. When binding was performed at 4°C, staining at the surface of the cells was observed (Figure 3). Given the way in which the Duolink-PLA technique works, this indicates that both FVIII and VWF locate at the cellular surface in the vicinity (<40 nm) of Siglec-5. Subsequent exposure of the cells to 37°C (which initiates the endocytic machinery) resulted in a positive Duolink-PLA staining for FVIII or VWF with an early endosomal marker (EEA1). This positive staining was exclusively observed for Siglec-5 expressing cells exposed to 37°C, and not for cells being maintained at 4°C or for cells lacking Siglec-5 (Figure 4). Apparently, the presence of Siglec-5 is associated with the uptake of FVIII and VWF, and their redistribution to the early endosomes. This observation is in agreement with other studies that have established the endocytic potential of Siglec-5, which allows the cellular uptake of Siglec-5 binding elements.8,16,17
Despite the capacity of FVIII to interact with Siglec-5 by itself, it is important to realize that this interaction seems regulated by VWF. As for the interaction with many other receptors,20 we observe that FVIII is inaccessible for Siglec-5 when bound to VWF. This suggests that in the circulation the majority of the FVIII will not interact with Siglec-5 itself, but presumably binding will be mediated via its carrier-protein VWF.
In our search for the physiological relevance of the interaction between FVIII/VWF and Siglec-5, we were limited by the fact that Siglec-5 is not well conserved between species,9 and there is no appropriate homolog in mice. To overcome this limitation, we decided to express Siglec-5 in mouse liver via hydrodynamic gene transfer. Analysis of liver tissue sections taken from mice after the gene transfer procedure revealed that 20-30% of the liver hepatocytes stained positive for Siglec-5 (Figure 5B). Interestingly, levels of the circulating FVIII/VWF complex were markedly lower in Siglec-5 expressing mice compared to mock-treated mice, whereas levels of another coagulation protein (Factor X) remained similar (Figure 5C). These data indicate that Siglec-5 may contribute to the regulation of FVIII/VWF levels. At this point, a number of options should be considered that could contribute to the mechanism by which hepatic Siglec-5 expression regulates VWF/FVIII plasma levels. Firstly, we cannot exclude the possibility that hepatic expression of Siglec-5 results in a reduced expression of VWF and FVIII in endothelial cells. Secondly, the reduced levels of FVIII/VWF might be explained by the expression of Siglec-5 resulting in an increased capacity of macrophages to clear the FVIII/VWF complex. Thirdly, it is possible that Siglec-5-expression converts these hepatocytes in cells that are able to clear the complex from the circulation. Obviously, additional studies are needed in this regard. Preliminary experiments revealed that, apart from Siglec-5, also other Siglecs seem able to interact with FVIII and/or VWF. The relative contribution (if any) of these other Siglecs in relation to Siglec-5 remains to be established. Another issue that needs to be resolved is whether Siglec-5 can bind VWF or FVIII irrespective of the cell type in which it is expressed, or whether cell specific elements are present that regulate binding to this receptor. It also remains to be determined to what extent Siglec-5 contributes to the regulation of FVIII/VWF levels with respect to other receptors for this protein complex, such as LDL-receptor related protein-1, LDL-receptor and asialoglycoprotein receptor.26
Supplementary Material
Acknowledgments:
we thank Philipe Leclerc (Institut Fédératif de Recherche 93, Le Kremlin-Bicêtre) for excellent support in the microscopic analysis.
Funding: this work was funded via grant ANR-08-CEXC-018-01 from the Agence Nationale de la Recherche, a PhD-thesis research grant from Insitut Servier & Association Nationale de la Recherche Technique and a grant from Fondation pour la Recherche Médicale (FRM-SPF20101220866).
Footnotes
The online version of this article has a Supplementary Appendix.
Authorship and Disclosures: The information provided by the authors about contributions from persons listed as authors and in acknowledgments is available with the full text of this paper at www.haematologica.org.
Financial and other disclosures provided by the authors using the ICMJE (www.icmje.org) Uniform Format for Disclosure of Competing Interests are also available at www.haematologica.org.
References
- 1.Lenting PJ, van Mourik JA, Mertens K. The life cycle of coagulation factor VIII in view of its structure and function. Blood 1998;92(11):3983-96 [PubMed] [Google Scholar]
- 2.Sadler JE. Biochemistry and genetics of von Willebrand factor. Annu Rev Biochem 1998; 67:395-424 [DOI] [PubMed] [Google Scholar]
- 3.Lenting PJ, Pegon JN, Christophe OD, Denis CV. Factor VIII and von Willebrand factor--too sweet for their own good. Haemophilia 2010;16(Suppl 5):194-9 [DOI] [PubMed] [Google Scholar]
- 4.Hironaka T, Furukawa K, Esmon PC, Fournel MA, Sawada S, Kato M, et al. Comparative study of the sugar chains of factor VIII purified from human plasma and from the culture media of recombinant baby hamster kidney cells. J Biol Chem 1992;267(12):8012-20 [PubMed] [Google Scholar]
- 5.Matsui T, Titani K, Mizuochi T. Structures of the asparagine-linked oligosaccharide chains of human von Willebrand factor. Occurrence of blood group A, B, and H(O) structures. J Biol Chem 1992;267(13):8723-31 [PubMed] [Google Scholar]
- 6.Canis K, McKinnon TA, Nowak A, Panico M, Morris HR, Laffan M, et al. The plasma von Willebrand factor O-glycome comprises a surprising variety of structures including ABH antigens and disialosyl motifs. J Thromb Haemost 2010;8(1):137-45 [DOI] [PubMed] [Google Scholar]
- 7.Schilow W F, Schoerner-Burkhardt E, Seitz R. Charge analysis of N-glycans from human recombinant coagulation factor VIII and human FVIII standards. Thromb Haemost 2004;92(2):427-8 [PubMed] [Google Scholar]
- 8.Lock K, Zhang J, Lu J, Lee SH, Crocker PR. Expression of CD33-related siglecs on human mononuclear phagocytes, monocyte-derived dendritic cells and plasmacytoid dendritic cells. Immunobiology 2004; 209(1-2):199-207 [DOI] [PubMed] [Google Scholar]
- 9.Jandus C, Simon HU, von Gunten S. Targeting siglecs--a novel pharmacological strategy for immuno- and glycotherapy. Biochem Pharmacol 2011;82(4):323-32 [DOI] [PubMed] [Google Scholar]
- 10.Cornish AL, Freeman S, Forbes G, Ni J, Zhang M, Cepeda M, et al. Characterization of siglec-5, a novel glycoprotein expressed on myeloid cells related to CD33. Blood 1998;92(6):2123-32 [PubMed] [Google Scholar]
- 11.Zhuravleva MA, Trandem K, Sun PD. Structural implications of Siglec-5-mediated sialoglycan recognition. J Mol Biol 2008;375(2):437-47 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Brinkman-Van der Linden EC, Varki A. New aspects of siglec binding specificities, including the significance of fucosylation and of the sialyl-Tn epitope. Sialic acid-binding immunoglobulin superfamily lectins. J Biol Chem 2000;275(12):8625-32 [DOI] [PubMed] [Google Scholar]
- 13.Carlin AF, Chang YC, Areschoug T, Lindahl G, Hurtado-Ziola N, King CC, et al. Group B Streptococcus suppression of phagocyte functions by protein-mediated engagement of human Siglec-5. J Exp Med 2009;206(8):1691-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Erickson-Miller CL, Freeman SD, Hopson CB, D'Alessio KJ, Fischer EI, Kikly KK, et al. Characterization of Siglec-5 (CD170) expression and functional activity of anti-Siglec-5 antibodies on human phagocytes. Exp Hematol 2003;31(5):382-8 [DOI] [PubMed] [Google Scholar]
- 15.Avril T, Freeman SD, Attrill H, Clarke RG, Crocker PR. Siglec-5 (CD170) can mediate inhibitory signaling in the absence of immunoreceptor tyrosine-based inhibitory motif phosphorylation. J Biol Chem. 2005; 280(20):19843-51 [DOI] [PubMed] [Google Scholar]
- 16.Jones C, Virji M, Crocker PR. Recognition of sialylated meningococcal lipopolysaccharide by siglecs expressed on myeloid cells leads to enhanced bacterial uptake. Mol Microbiol 2003;49(5):1213-25 [DOI] [PubMed] [Google Scholar]
- 17.Rapoport EM, Sapot'ko YB, Pazynina GV, Bojenko VK, Bovin NV. Sialoside-binding macrophage lectins in phagocytosis of apoptotic bodies. Biochemistry (Mosc.) 2005;70(3):330-8 [DOI] [PubMed] [Google Scholar]
- 18.van Schooten CJ, Shahbazi S, Groot E, Oortwijn BD, van den Berg HM, Denis CV, et al. Macrophages contribute to the cellular uptake of von Willebrand factor and factor VIII in vivo. Blood 2008;112(5):1704-12 [DOI] [PubMed] [Google Scholar]
- 19.Casale TB, Kaliner M. A rapid method for isolation of human mononuclear cells free of significant platelet contamination. J Immunol Methods 1982;55(3):347-53 [DOI] [PubMed] [Google Scholar]
- 20.Lenting PJ, Christophe OD, Gueguen P. The disappearing act of factor VIII. Haemophilia 2010;16(102):6-15 [DOI] [PubMed] [Google Scholar]
- 21.Dam TK, Brewer C F. Effects of clustered epitopes in multivalent ligand-receptor interactions. Biochemistry 2008;47(33):8470-6 [DOI] [PubMed] [Google Scholar]
- 22.Varki A. Natural ligands for CD33-related Siglecs¿̣ Glycobiology 2009;19(8):810-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lenting PJ, Neels JG, van den Berg BM, Clijsters PP, Meijerman DW, Pannekoek H, et al. The light chain of factor VIII comprises a binding site for low density lipoprotein receptor-related protein. J Biol Chem. 1999;274(34):23734-9 [DOI] [PubMed] [Google Scholar]
- 24.Bovenschen N, Herz J, Grimbergen JM, Lenting PJ, Havekes LM, Mertens K, et al. Elevated plasma factor VIII in a mouse model of low-density lipoprotein receptorrelated protein deficiency. Blood 2003;101(10):3933-9 [DOI] [PubMed] [Google Scholar]
- 25.Pipe SW. Functional roles of the factor VIII B domain. Haemophilia 2009;15(6):1187-96 [DOI] [PubMed] [Google Scholar]
- 26.Lenting PJ, Van Schooten CJ, Denis CV. Clearance mechanisms of von Willebrand factor and factor VIII. J Thromb Haemost. 2007;5(7):1353-60 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.