

Abstract
STUDY QUESTION
Can time-lapse analysis of cell division timings [morphokinetics (MK)] in mouse embryos detect toxins at concentrations that do not affect blastocyst formation?
SUMMARY ANSWER
An MK algorithm enhances assay sensitivity while providing results 24–48 h sooner than the traditional mouse embryo assay (MEA).
WHAT IS KNOWN ALREADY
Current quality control testing methodology is sensitive but further improvements are needed to assure optimal culture conditions. MKs of embryo development may detect small variations in culture conditions.
STUDY DESIGN
Cross sectional—control versus treatment. Mouse embryo development kinetics of 466 embryos were analyzed according to exposure to various concentrations of toxins and toxic mineral oil.
MATERIALS, SETTING, METHODS
Cryopreserved 1-cell embryos from F1 hybrid mice were cultured with cumene hydroperoxide (CH) (0, 2, 4, 6 and 8 µM) and Triton X-100 (TX-100; 0, 0.0008, 0.0012, 0.0016 and 0.002%). Using the Embryoscope, time-lapse images were obtained every 20 min for 120 h in seven focal planes. End-points were timing and pattern of cell division and embryo development. The blastocyst rate (BR) was defined as the percentage of embryos that developed to the expanded blastocyst stage within 96 h.
MAIN RESULTS AND THE ROLE OF CHANCE
BR was not affected for embryos cultured in the three lowest concentrations of CH and the four lowest concentrations of TX-100. In contrast, a unique MK model detected all concentrations tested (P < 0.05). The MK model identified toxicity in two lots of toxic mineral oil that did not affect BR (P < 0.05).
LIMITATIONS, REASONS FOR CAUTION
A limited number of toxins were used so that the results may not apply to all potential embryo toxins. A larger sample size may also demonstrate other statistically significant developmental kinetic parameters.
WIDER IMPLICATIONS OF THE FINDINGS
MKs in mouse embryos are a sensitive and efficient method for quality control testing of in vitro culture conditions. BR, the end-point of traditional quality control assays, did not detect sublethal concentrations of toxins in the culture milieu in our study. This study demonstrates that temporal variation at key developmental stages reflects the quality of the culture environment. An MEA that incorporates MK will provide enhanced sensitivity and faster turn-around times.
STUDY FUNDING/COMPETING INTEREST(S)
The study was supported by Mayo Clinic Department of Obstetrics and Gynecology Small Grant Program. The authors have no competing interests to declare.
Keywords: time-lapse imaging, morphokinetics, embryo culture, quality control, toxicity
Introduction
The goal of an IVF program is a high live birth rate of healthy singletons. This success is dependent in part on a culture system that supports development of healthy embryos that are capable of realizing their implantation potential (Puissant et al., 1987; Hu et al., 1998; Strandell et al., 2000). Optimal culture conditions are the product of a good quality control system, which should maintain a stable culture environment and screen products for embryo-toxic substances (Gardner et al., 2005; Lane et al., 2008; Morbeck, 2012).
In order to optimize conditions and achieve consistent results, standardization and quality assessment of all culture components is essential. Bioassays employed by manufacturers and individual laboratories include the 1-cell and 2-cell mouse embryo assay (MEA) and the human sperm motility assay (HSMA). While the 1-cell MEA is more sensitive than the 2-cell MEA (Davidson et al., 1988; Scott et al., 1993; Hughes et al., 2010; Morbeck et al., 2010) or HSMA (Hughes et al., 2010), it is unclear if the sensitivity of these assays is sufficient to detect toxins relevant to the human IVF laboratory. Strain-to-strain variation provides evidence that the current standard for QC testing, which uses the 1-cell F1 hybrid mouse MEA, may not be sensitive enough to detect contaminants that affect human gametes and embryos (Khan et al., 2013). Recent cases where toxic mineral oils passed MEA testing by the manufacturer but reached clinical laboratories and were subsequently recalled support this hypothesis (Morbeck, 2012; Morbeck et al., 2012).
An inherent limitation of the MEA is the relative lack of sensitivity of the end-point: expanded blastocysts can form from few blastomeres and are not always associated with viability, a fact confirmed with a more objective end-point such as cell number (Lane and Gardner 1997). In contrast, time-lapse imaging is a continuous, objective record of embryo development that provides timing of cell division events (Gonzales et al., 1995; Arav et al., 2008; Lemmen et al., 2008). While timing of cell divisions has been promoted as an important variable for selecting embryos for transfer in the clinical IVF laboratory (Pribenszky et al., 2010; Wong et al., 2010; Azzarello et al., 2012; Cruz et al., 2012; Hlinka et al., 2012; Kirkegaard et al., 2012; Meseguer et al., 2012), timing as a QC tool in the MEA has not been adopted, likely due to limited evidence and lack of standardization. The development of time-lapse imaging technology provides a tool to test the hypothesis that timing of cleavage events is a sensitive marker for in vitro stress.
The aim of this study was to determine whether toxins at concentrations that do not affect blastocyst formation could be detected by morphokinetic (MK) analysis of cell division. The relative sensitivity of MKs was tested with two toxins to develop a quality control algorithm based on timing of cell divisions. These markers were then applied to mineral oil that passed standard quality control testing but had been recalled by the manufacturer due to potential toxicity to human embryos.
Materials and Methods
Culture of embryos
Cryopreserved 1-cell mouse embryos from F1 hybrid mice were obtained from Embryotech Laboratories (Haverhill, MA, USA). The embryos were thawed according to the manufacturer's instructions and allowed to equilibrate for 10 min at room temperature in HTF with HEPES (HTF-HEPES; Irvine Scientific, Irvine, CA, USA) containing 0.1 mg/ml polyvinyl alcohol (PVA) (Sigma Chemical, St. Louis, MO, USA). Following equilibration, embryos were placed in individual wells (n = 10/treatment per replicate) in an EmbryoSlide (Unisense Fertilitech, Aarhus, Denmark) and placed immediately into an EmbryoScope (Unisense Fertilitech) for time-lapse analysis. All tests were performed with HTF containing EDTA and glutamine (In Vitro Care, Frederick, MD, USA) supplemented with 0.1 mg/ml PVA. EmbryoSlides were prepared the day before thawing embryos and were equilibrated overnight in an incubator with 5% O2, 6.5% CO2 and balance nitrogen. Individual wells in the EmbryoSlide contained either 10 µl (for oil toxicity experiments) or 25 µl (for Triton X-100) HTF and the wells were covered with 1.2 ml mineral oil (Fisher Scientific, Pittsburgh, PA, USA).
Experiment one: cumene hydroperoxide
Cumene hydroperoxide (CH; Sigma-Aldrich) was used as a surrogate for peroxides that can accumulate naturally in mineral oil (Otsuki et al., 2007; Morbeck et al., 2010) and was prepared as previously described (Hughes et al., 2010). For each replicate (n = 3), an EmbyoSlide (n = 5/replicate) was prepared for each concentration of CH (0, 2, 4, 6 or 8 µM).
Experiment two: Triton X-100
Triton X-100 (TX-100) is a detergent that was previously identified as a contaminant of mineral oil (Morbeck et al., 2010). TX-100 was added directly to HTF and embryos were cultured in 25 µl in individual wells in an EmbryoSlide covered with 1.2 ml of mineral oil. For each replicate (n = 3), an EmbyoSlide (n = 6/replicate) was prepared for each concentration of TX-100 (0, 0.0004, 0.0008, 0.0012, 0.0016 or 0.002% v/v).
Experiment three: recalled oil lots
Two lots of US Pharmaceutical grade mineral oil that were recalled in 2010 due to suspicion of toxicity during human embryo culture were compared with control oil (Fisher). The two lots of oil (Lot 1 and Lot 2) were from different suppliers and passed standard mouse embryo testing. A sample size estimate was performed to determine the number of embryos needed to detect a difference using the MK model based on results from experiments 1 and 2. Assuming 60% of embryos in the control group fit the MK model, 38 embryos in each group would be needed to detect a decrease in the model fit rate to 40% (20% decrease) with 80% power and an α-error rate of 0.05. Therefore, four replicates (n = 40 embryos/treatment) were performed with 10 embryos per replicate in HTF.
Time-lapse system
Precise division kinetics were captured using time-lapse microscopy with an EmbryoScope incubator (Unisense Fertilitech). The embryos were placed in the incubator and seven planes of images were obtained every 20 min for 120 h.
The blastocyst rate (BR) was defined as the percentage of embryos that developed to the expanded blastocyst stage by 96 h. Cell division timings were determined manually and included t2, t3, t4, t5, t6, t7 and t8 (time from thawing to 2, 3, 4, 5, 6, 7 and 8 cells), time of compaction, tM, time to formation of a blastocoel, tB and time to an expanded blastocyst (tEB). Other parameters included cc2 (duration of the second cell cycle—the time an embryo is at the 2-cell stage), i3 (duration of the third interphase—the time an embryo is at the 4-cell stage), s2 (synchrony of the second cell division, which is the time an embryo is at the 3-cell stage) and s3 (synchrony of the third cell division, which is the time an embryo contains 5, 6 or 7 cells).
Data and statistical analysis
Data were analyzed using a one-way analysis of variance with Dunnett's test for comparison of timings to the control and the χ2 test for comparison of proportions. The timings followed a mostly normal distribution. Timings that showed significant treatment effects were used to develop a multivariate model. Optimal timing for these events (t5, cc2, i3 and tB) was defined as less than the upper 95% confidence interval (CI) of the control for each replicate. Statistical analyses were performed using JMP statistical software (SAS Institute, Cary, NC, USA).
Results
Toxin sensitivity
Timing of cleavage events, cleavage synchrony and cell cycle length for different concentrations of CH and TX-100 are presented in Tables I and II, respectively. The completion of the second cell cycle (t4) was the earliest event with a delay in response to CH at 4 µM, with a more profound effect at initiation of the third cell cycle (t5). The synchrony of the third cell cycle (s3) was not affected by CH at this concentration, indicating that the timings of t6, t7 and t8 are unchanged despite effects on t5 and thus do not provide additional information. CH had a strong effect on time of morula formation and tEB at 4 µM, though these variables are the least objective and most difficult to standardize. In contrast, blastocoel formation (tB), a feature of development that is distinct and easily assessed, was delayed at 4 µM CH. Duration of the second and third cell cycles was affected by CH at 6 µM.
Table I.
Division kinetics of 1-cell mouse embryos cultured with increasing concentrations of CH.
| CH |
|||||
|---|---|---|---|---|---|
| 0 µM | 2 µM | 4 µM | 6 µM | 8 µM | |
| Embryos | 31 | 31 | 30 | 32 | 29 |
| t2 (h) | 5.8 (5.3–6.3) | 5.7 (5.0–6.4) | 6.1 (5.5–6.7) | 5.6 (5.0–6.3) | 6.7 (5.9–7.6) |
| t3 (h) | 28.2 (27.3–29.1) | 27.3 (25.4–29.2) | 30.7 (29.1–32.3) | 30.9 (29.5–32.2)* | 33.4 (31.8–34.9)*** |
| t4 (h) | 29.0 (28.0–29.9 | 29.0 (27.1–30.9) | 33.0 (30.9–35.1)* | 32.5 (30.7–34.4)** | 34.4 (32.8–36.1)*** |
| t5 (h) | 38.3 (37.1–39.4) | 39.9 (38.5–41.2) | 44.4 (42.1–46.7)*** | 45.8 (44.1–47.5)*** | 49.7 (46.9–52.4)*** |
| t6 (h) | 38.8 (37.7–39.9) | 40.5 (39.1–41.9) | 45.8 (42.6–49.0)*** | 46.8 (44.8–48.7)*** | 51.1 (48.2–54.0)*** |
| t7 (h) | 39.5 (38.6340.7) | 42.0 (40.2–43.8) | 46.0 (43.7–48.2)*** | 47.4 (45.5–49.3)*** | 53.1 (49.7–56.5)*** |
| t8 (h) | 40.2 (38.9–41.5) | 43.1 (40.8–45.4) | 46.6 (44.3–48.9)*** | 48.0 (46.0–50.0)*** | 54.0 (50.3–57.6)*** |
| tM (h) | 52.8 (51.1–54.5) | 55.9 (54.4–57.5)* | 58.6 (56.7–60.6)*** | 59.5 (57.5–61.5)*** | 68.2 (61.6–74.8)*** |
| tB (h) | 67.1 (64.9–69.3) | 70.0 (67.5–71.6) | 73.6 (70.6–76.5)** | 74.9 (71.6–78.2)*** | 90.6 (90.6–90.6)** |
| tEB (h) | 75.7 (73.8–77.6) | 79.7 (77.2–82.2) | 84.0 (80.7–87.3)*** | 85.9 (82.1–89.7)*** | |
| cc2 (h) | 22.4 (21.7–23.1) | 21.6 (19.8–23.4) | 24.6 (23.2–26.0) | 25.3 (24.0–26.5)** | 26.7 (25.3–28.1)*** |
| s2 (h) | 0.8 (0.6–1.1) | 1.7 (0.7–2.7) | 2.3 (1.3–3.4)* | 1.6 (.6–2.6) | 1.3 (0.8–1.8) |
| i3 (h) | 9.3 (8.8–9.7) | 10.8 (9.1–12.6) | 11.4 (9.5–13.2) | 13.3 (11.9–14.6)*** | 16.7 (14.4–18.9)*** |
| s3 (h) | 1.9 (1.4–2.4) | 3.2 (1.9–4.5) | 3.4 (2.4–4.4) | 2.7 (2.1–3.3) | 4.4 (2.5–6.3)*** |
Values are mean (95% CI). t2, t3, t4, t5, t6, t7 and t8, cleavage times from a zygote to a 2–8-cell embryo; tM, time to morula formation; tB, time to formation of blastocoel cavity; tEB, time to expanded blastocyst; cc2, duration of 2-cell stage (t3–t2); i3, duration of 4-cell stage (t5–t4); s2, synchrony second cell cycle (t4–t3); s3, synchrony of third cell cycle (t8–t5).
*P < 0.05, **P < 0.01, ***P < 0.001 (Dunnett's test).
Table II.
Division kinetics of 1-cell mouse embryos cultured with increasing concentrations of Triton X-100 (TX-100).
| Triton X-100 |
||||||
|---|---|---|---|---|---|---|
| 0 | 0.0004% | 0.0008% | 0.0012% | 0.0016% | 0.0020% | |
| Embryos | 29 | 31 | 32 | 31 | 32 | 32 |
| t2 (h) | 5.6 (4.9–6.3) | 6.6 (5.8–7.4) | 6.5 (5.8–7.2) | 6.8 (6.2–7.4) | 7.4 (6.6–8.1)** | 7.9 (7.0–8.7)*** |
| t3 (h) | 27.3 (26.4–28.2) | 27.8 (26.1–29.5) | 29.2 (28.0–30.4) | 30.0 (29.2–30.8)** | 31.4 (30.4–32.4)*** | 33.5 (32.5–34.6)*** |
| t4 (h) | 28.2 (27.2–29.2) | 29.7 (27.6–31.7) | 30.7 (29.6–31.9)* | 30.9 (30.0–31.8)* | 32.6 (31.5–33.7)*** | 35.5 (33.8–37.2)*** |
| t5 (h) | 36.8 (35.7–37.8) | 38.1 (36.7–39.6) | 39.0 (37.7–40.2) | 39.5 (38.5–40.4)* | 41.4 (40.3–42.3)*** | 45.0 (43.2–46.7)*** |
| t6 (h) | 37.2 (36.1–38.3) | 38.7 (37.1–40.2) | 39.5 (38.3–40.8) | 40.0 (39.0–41.0)* | 41.9 (40.7–43.1)*** | 45.7 (43.9–47.4)*** |
| t7 (h) | 37.9 (36.7–39.0) | 40.3 (38.3–42.2) | 40.9 (39.3–42.5)* | 40.9 (39.8–42.0)* | 42.9 (41.5–44.3)*** | 46.6 (45.1–48.1)*** |
| t8 (h) | 38.3 (37.1–39.4) | 40.3 (38.6–42.0) | 41.5 (40.0–43.1)** | 41.6 (40.4–42.8)** | 43.5 (42.0–45.0)*** | 47.5 (45.8–49.1)*** |
| tM (h) | 51.2 (49.6–52.8) | 54.2 (52.2–56.3) | 56.3 (54.4–58.3) | 56.2 (54.7–57.6)*** | 60.4 (58.4–62.3)*** | 66.1 (63.9–68.3)*** |
| tB (h) | 63.9 (62.3–65.6) | 68.7 (65.9–71.5)* | 72.3 (69.3–75.3)*** | 72.3 (70.3–74.4)*** | 78.9 (76.2–81.5)*** | 84.0 (81.4–86.4)*** |
| tEB (h) | 75.7 (72.7–78.7) | 80.6 (80.0–84.2) | 83.4 (79.9–86.9)** | 85.6 (82.2–89.1)*** | 89.2 (85.9–92.5)*** | 95.8 (92.0–99.5)*** |
| cc2 (h) | 21.7 (21.1–22.3) | 21.2 (19.8–22.6) | 22.7 (21.8–23.5) | 23.2 (22.7–23.7)* | 24.0 (23.5–24.6)*** | 25.7 (25.1–26.2)*** |
| s2 (h) | 0.9 (0.6–1.1) | 1.8 (0.8–2.9) | 1.6 (0.7–2.4) | 0.9 (0.6–1.1) | 1.2 (0.9–1.5) | 2.0 (1.1–2.8) |
| i3 (h) | 8.6 (8.2–8.9) | 8.5 (7.4–9.6) | 8.2 (7.8–8.7) | 8.5 (8.1–9.0) | 8.8 (8.5–9.1) | 9.5 (8.6–10.3) |
| s3 (h) | 1.5 (1.1–1.9) | 2.4 (1.8–3.0) | 2.6 (1.8–3.4) | 2.1 (1.7–2.6) | 2.1 (1.5–2.7) | 3.1 (2.3–3.9) |
Values are mean (95% CI). t2, t3, t4, t5, t6, t7 and t8, cleavage times from a zygote to 2–8-cell embryo; tM, time to morula formation; tB, time to formation of blastocoel cavity; tEB, time to expanded blastocyst; cc2, duration of 2-cell stage (t3–t2); i3, duration of 4-cell stage (t5–t4); s2, synchrony second cell cycle (t4–t3); s3, synchrony of third cell cycle (t8–t5).
*P < 0.05, **P < 0.01, ***P < 0.001.
Formation of a blastocoel cavity (tB) was very sensitive to TX-100 (Table II), with a delay of nearly 5 h at the lowest concentration tested (0.0004%; P < 0.05). Early cleavage events were not consistently affected by TX-100 at the two lowest concentrations tested. However, an effect of treatment was observed at 0.0012% TX-100 on most of the early timings, including t5 and cc2.
Based on these results, the percentage of embryos that exhibited optimal timings based on the 95% CI for the control for each replicate were compared across concentrations within treatment for t5, cc2 and tB (Fig. 1). Optimal timings relative to the control's 95% CI provides an objective standardization per replicate that accounts for timing variation among replicates that is inherent with the MEA. Individually, optimal timings for the three variables detected all but the lowest concentration of CH tested (Fig. 1A). In contrast, tB but not t5 and cc2 detected the two lowest concentrations of TX-100 (Fig. 1B), illustrating a difference in the effect of these two toxins on early versus late stages of embryo development.
Figure 1.
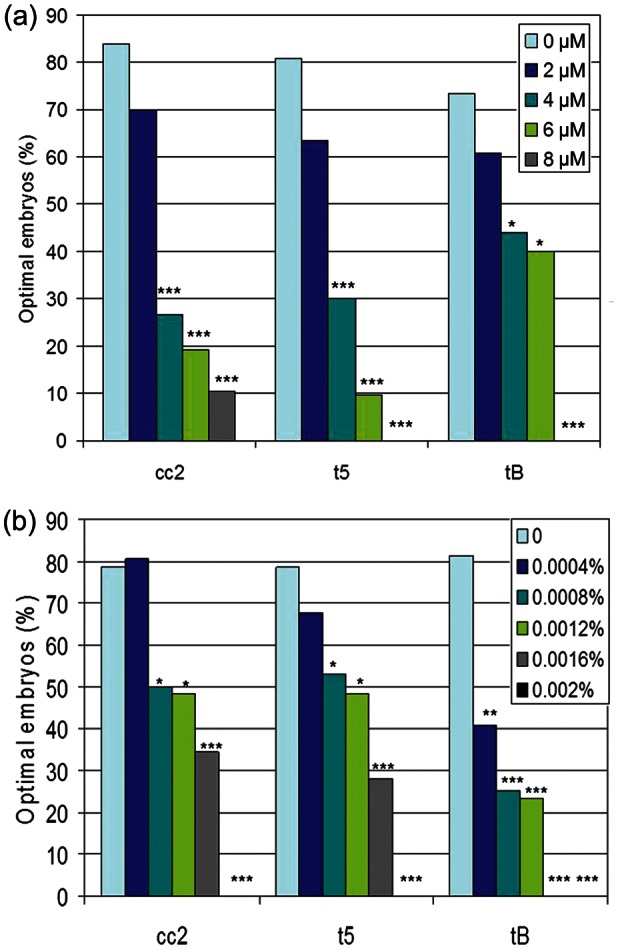
Percentage of embryos meeting optimal criteria (<95% CI of controls) for cc2, t5 and tB in response to (a) CH or (b) Triton X-100. t5 = time to 5-cell embryo; cc2 = second cell cycle length; tB = time to formation of blastocoel cavity *P < 0.05, **P < 0.01, ***P < 0.001 versus the control (χ2).
A powerful feature of time-lapse technology is the ability to combine several objective measures into one analysis model. In this phase of the study, t5, cc2 and tB were combined into one optimized model using the control group's 95% CI (MK Model), a model that provides results by 72 h of culture. In this model, the lowest concentration of both CH (Fig. 2A) and TX-100 (Fig. 2B) were detected by MKs. In contrast, the traditional end-point of expanded BR at 96 h was able to detect only the highest concentrations of either toxin (Fig. 2A and B).
Figure 2.
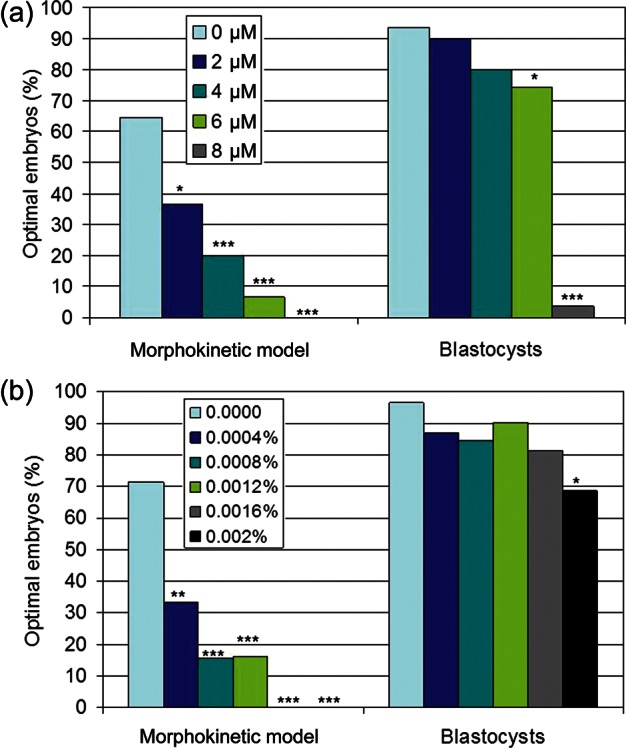
Percentage of embryos meeting optimal criteria (<95% CI of controls) for the morphokinetic (MK) model or BR in response to (a) CH or (b) Triton X-100. MK Model = combined optimal t5, cc2, tB; *P < 0.05, **P < 0.01, ***P < 0.001 versus the control (χ2).
Mineral oil for IVF
Timing of cleavage events, cleavage synchrony and cell cycle length for control, Lot 1 and Lot 2 oils are presented in Table III. Similar to CH, the initiation of the third cell cycle (t5) was affected by the two suspect oil lots. In contrast to both CH and TX-100, duration of the third interphase (i3) was longer (P < 0.001) for both oils, indicating an effect on mitosis at this stage of development that was not evident in the toxin studies. When compared with the control's 95% upper CI, fewer embryos met the criteria for t5 and i3 in the two lots of affected oil, whereas cc2 detected a difference only for Lot 1 and tB only for Lot 2 (Fig. 3A).
Table III.
Division kinetics of 1-cell mouse embryos cultured with different lots of mineral oil.
| Control | Lot 1 | Lot 2 | |
|---|---|---|---|
| Embryos | 42 | 42 | 42 |
| t2 (h) | 10.2 (9.8–10.7) | 10.7 (10.3–11.1) | 11.0 (9.9–12.2) |
| t3 (h) | 34.1 (33.4–34.7) | 35.6 (34.8–36.4)* | 35.2 (34.2–36.2) |
| t4 (h) | 34.9 (34.2–35.6) | 36.6 (35.7–37.5)* | 35.9 (34.8–37.0) |
| t5 (h) | 44.7 (43.8–45.5) | 48.4 (47.3–49.5)*** | 47.4 (45.7–49.1)*** |
| t8 (h) | 47.9 (46.6–49.3) | 51.8 (50.3–53.2)** | 49.7 (47.6–51.8) |
| tM (h) | 59.7 (58.1–61.2) | 65.4 (64.0–66.9)*** | 62.0 (60.1–63.9) |
| tB (h) | 73.1 (71.0–75.2) | 77.6 (75.1–80.1)* | 76.9 (73.7–80.1) |
| tEB (h) | 91.1 (88.4–93.7) | 97.1 (94.4–99.9)** | 94.3 (92.0–96.6) |
| cc2 (h) | 23.8 (23.4–24.2) | 24.9 (24.3–25.4) | 24.2 (22.8–25.6) |
| s2 (h) | 0.8 (0.6–1.1) | 1.0 (0.7–1.3) | 0.7 (.5–1.0) |
| i3 (h) | 9.8 (9.4–10.2) | 11.8 (11.1–12.4)*** | 11.4 (10.6–12.2)*** |
| s3 (h) | 3.3 (2.4–4.1) | 3.4 (2.7–4.0) | 2.8 (2.2–3.4) |
Values are mean (95% CI). t2, t3, t4, t5, t6, t7 and t8, cleavage times from a zygote to 2–8-cell embryo; tM, time to morula formation; tB, time to formation of blastocoel cavity; tEB, time to expanded blastocyst; cc2, duration of 2-cell stage (t3–t2); i3, duration of 4-cell stage (t5–t4); s2, synchrony second cell cycle (t4–t3); s3, synchrony of third cell cycle (t8–t5).
*P < 0.05, **P < 0.01, ***P < 0.001 (Dunnett's test).
Figure 3.
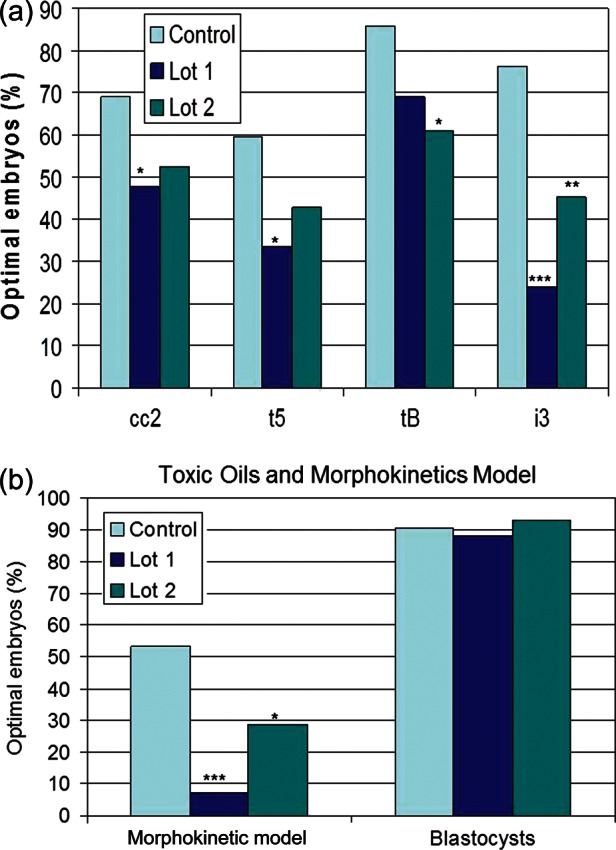
Percentage of embryos meeting optimal criteria (<95% CI of controls) for (a) individual parameters or (b) a morphokinetic (MK) model or BR for two lots of embryotoxic mineral oil (Lot 1 and Lot 2). t5 = time to 5-cell embryo; cc2 = second cell cycle length; tB = time to formation of blastocoel cavity; i3 = time at 4-cell stage; MK Model = combined optimal t5, cc2, tB and cc3; *P < 0.05, **P < 0.01, ***P < 0.001 versus the control oil (χ2).
When a new model was developed that included the four variables identified in both studies (t5, cc2, i3 and tB), an adverse effect of both Lot 1 and Lot 2 was identified (Fig. 3B; P < 0.05). Similar to results of manufacturer's quality control testing, neither of these lots affected BRs and all three lots would be considered acceptable with rates >80%.
Discussion
In this study we demonstrated that timing of early cleavage events of mouse embryos identified with time-lapse imaging are sensitive markers of toxins at concentrations that do not affect blastocyst development, the primary end-point of standard quality control testing. The sensitivity of MK analysis was confirmed when it successfully identified mineral oil that had passed manufacturer's quality control testing but was recalled due to questions of toxicity to human embryos. In addition to improved sensitivity over the 96 h 1-cell MEA, the cell cycle events that were most sensitive to toxicity occurred within the first 72 h of culture. These results present a new paradigm for quality control testing: a subjective MEA with improved sensitivity that provides results at least 24 h sooner than the standard MEA.
While time-lapse analysis of embryo development has been used in research (Bavister, 1995) and was recently introduced into clinical IVF (Lemmen et al., 2008; Basile et al., 2013), this is the first study of the utility of time-lapse for quality control of products used in the IVF laboratory. Previous work with animal models provide evidence that timing of cleavage events could be useful for determining viability of embryos, and therefore may provide a tool for quality control (McKiernan and Bavister 1994; Gardner et al., 2005; Taft 2008). Additional studies indicated that the duration of the third cell cycle (4–8 cells) showed a significant correlation with blastocyst formation for hamster embryos (Gonzales et al., 1995). In contrast, timing of the first cell division (Arav et al., 2008; Pribenszky et al., 2010) and the duration of the second cell cycle (Pribenszky et al., 2010) in mouse embryos were correlated with blastocyst formation in vitro. While species differences exist and these may depend on timing of activation of the embryonic genome, the timing of the first three cell divisions is dynamic and reflects the developmental capacity of the early embryo. Unlike blastocyst formation, which is known to be a poor measure of viability (Lane and Gardner 1996, 1997), our results demonstrate that time-lapse imaging provides subjective timings of distinct developmental events that are responsive to quality of culture conditions.
Development of a bioassay with sensitivity similar to a human embryo is fraught with challenges. Few studies have compared the sensitivities of the various bioassays reported in the literature, and a direct comparison of toxin sensitivity with human embryos and toxins like TX-100 and CH has not been performed. Though a direct comparison of bioassays to human embryos is unlikely, validation of bioassay sensitivity using a robust scientific method is critical to assure that manufacturers and IVF laboratories are using the most sensitive and appropriate assays available. Most studies report toxicity of individual laboratory items without an identification of the toxin (Naz et al., 1986; Fleming et al., 1987; Claassens et al., 2000); however, reports with concentrations of known or suspected toxins have consistently demonstrated that the 1-cell MEA is superior to the 2-cell MEA or the sperm survival assay (Davidson et al., 1988; Scott et al., 1993; Hughes et al., 2010; Morbeck et al., 2010). Even though the 1-cell MEA is the industry standard, mineral oil that passed manufacturer QC illustrates that the assay, with BR as an end-point, can fail to detect clinically relevant toxins. This relative insensitivity of the MEA may be strain dependent, and at least one manufacturer uses an outbred mouse strain in their QC program. Using the same toxins (TX-100 and CH), we have confirmed that embryos from outbred mice are significantly more sensitive to in vitro culture stress than either inbred or F1/hybrid embryo (Khan et al., 2013). Embryos from outbred mice were four times more sensitive to CH than other strains and detected the lowest concentration tested (2 µM), a concentration that was also detected by the MK model in this study. The MK model also detected TX-100 at concentrations similar to and lower than the concentration detected by outbred mouse embryos. Application of the MK model to an outbred MEA may provide even further sensitivity, though there are limitations inherent with outbred mouse embryos that have limited their use as a routine QC tool.
The importance of an optimal environment for IVF and preimplantation embryo development cannot be overstated. Recent reports of adverse effects of mineral oil on pregnancy rates illustrate the importance of effective supply management in the IVF laboratory (Morbeck, 2012). Beyond the immediate consequences of suboptimal culture are the concerns with long-term health of children conceived through IVF. Several animal models have illuminated a direct role of culture environment on many facets of embryo viability and long-term development. For instance, mouse embryos cultured under suboptimal conditions demonstrated differences in birthweight (Banrezes et al., 2011), imprinting (Fernandez-Gonzalez et al., 2004; Rivera et al., 2008) and gene expression (Ecker et al., 2004; Rinaudo and Schultz 2004; Giritharan et al., 2007). Errors in imprinting in domestic animals have been linked to culture conditions as well (Lonergan et al., 2003). Adverse effects of in vitro culture may also affect human development, with evidence that shorter culture times yield better long-term outcomes (Kallen et al., 2010; Kalra et al., 2012) and that culture media composition can affect birthweights (Dumoulin et al., 2010; Nelissen et al., 2012).
The results of these studies provide a new approach to quality control testing for products used in the clinical IVF laboratory. A desirable quality control method should be reproducible, sensitive and require a minimum investment of time and labor. An MEA with MKs incorporates these elements and with the prospect of automated cell division annotation and data analysis will likely provide an even more robust and effective quality control tool for both manufacturers and IVF laboratories.
Authors' roles
H.S.W., J.R.F. and D.L.W. performed the experiments and collected the data. D.E.M. designed the study and analyzed the data with input from H.S.W. H.S.W. wrote the manuscript with input from D.E.M.
Funding
The study was supported by Mayo Clinic Department of Obstetrics and Gynecology Small Grant Program. Funding to pay the Open Access publication charges for this article was provided by Unisense FertiliTech A/S.
Conflict of interest
None declared.
References
- Arav A, Aroyo A, Yavin S, Roth Z. Prediction of embryonic developmental competence by time-lapse observation and ‘shortest-half’ analysis. RBM Online. 2008;17:669–675. doi: 10.1016/s1472-6483(10)60314-8. [DOI] [PubMed] [Google Scholar]
- Azzarello A, Hoest T, Mikkelsen AL. (2012) The impact of pronuclei morphology and dynamicity on live birth outcome after time-lapse culture. Hum Reprod. 2012;27:2649–2657. doi: 10.1093/humrep/des210. [DOI] [PubMed] [Google Scholar]
- Banrezes B, Sainte-Beuve T, Canon E, Schultz RM, Cancela J, Ozil JP. Adult body weight is programmed by a redox-regulated and energy-dependent process during the pronuclear stage in mouse. PloS One. 2011;6:e29388. doi: 10.1371/journal.pone.0029388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Basile N, Morbeck D, Garcia-Velasco J, Bronet F, Meseguer M. (2013) Type of culture media does not affect embryo kinetics: a time-lapse analysis of sibling oocytes. Hum Reprod. 2013;28:634–641. doi: 10.1093/humrep/des462. [DOI] [PubMed] [Google Scholar]
- Bavister BD. Culture of preimplantation embryos: facts and artifacts. Hum Reprod Update. 1995;1:91–148. doi: 10.1093/humupd/1.2.91. [DOI] [PubMed] [Google Scholar]
- Claassens OE, Wehr JB, Harrison KL. Optimizing sensitivity of the human sperm motility assay for embryo toxicity testing. Hum Reprod. 2000;15:1586–1591. doi: 10.1093/humrep/15.7.1586. [DOI] [PubMed] [Google Scholar]
- Cruz M, Garrido N, Herrero J, Perez-Cano I, Munoz M, Meseguer M. Timing of cell division in human cleavage-stage embryos is linked with blastocyst formation and quality. RBM Online. 2012;25:371–381. doi: 10.1016/j.rbmo.2012.06.017. [DOI] [PubMed] [Google Scholar]
- Davidson A, Vermesh M, Lobo R, Paulson R. Mouse embryo culture as quality control for human in vitro fertilization: the one-cell versus the two-cell model. Fertil Steril. 1988;49:516–521. doi: 10.1016/s0015-0282(16)59783-0. [DOI] [PubMed] [Google Scholar]
- Dumoulin JC, Land JA, Van Montfoort AP, Nelissen EC, Coonen E, Derhaag JG, Schreurs IL, Dunselman GA, Kester AD, Geraedts JP, et al. Effect of in vitro culture of human embryos on birthweight of newborns. Hum Reprod. 2010;25:605–612. doi: 10.1093/humrep/dep456. [DOI] [PubMed] [Google Scholar]
- Ecker DJ, Stein P, Xu Z, Williams CJ, Kopf GS, Bilker WB, Abel T, Schultz RM. Long-term effects of culture of preimplantation mouse embryos on behavior. Proc Natl Acad Sci USA. 2004;101:1595–1600. doi: 10.1073/pnas.0306846101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez-Gonzalez R, Moreira P, Bilbao A, Jimenez A, Perez-Crespo M, Ramirez MA, Rodriguez De Fonseca F, Pintado B, Gutierrez-Adan A. Long-term effect of in vitro culture of mouse embryos with serum on mRNA expression of imprinting genes, development, and behavior. Proc Natl Acad Sci USA. 2004;101:5880–5885. doi: 10.1073/pnas.0308560101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleming TP, Pratt HP, Braude PR. The use of mouse preimplantation embryos for quality control of culture reagents in human in vitro fertilization programs: a cautionary note. Fertil Steril. 1987;47:858–860. doi: 10.1016/s0015-0282(16)59179-1. [DOI] [PubMed] [Google Scholar]
- Gardner DK, Reed L, Linck D, Sheehan C, Lane M. Quality control in human in vitro fertilization. Semin Reprod Med. 2005;23:319–324. doi: 10.1055/s-2005-923389. [DOI] [PubMed] [Google Scholar]
- Giritharan G, Talbi S, Donjacour A, Di Sebastiano F, Dobson AT, Rinaudo PF. Effect of in vitro fertilization on gene expression and development of mouse preimplantation embryos. Reproduction. 2007;134:63–72. doi: 10.1530/REP-06-0247. [DOI] [PubMed] [Google Scholar]
- Gonzales DS, Pinheiro JC, Bavister BD. Prediction of the developmental potential of hamster embryos in vitro by precise timing of the third cell cycle. J Reprod Fertil. 1995;105:1–8. doi: 10.1530/jrf.0.1050001. [DOI] [PubMed] [Google Scholar]
- Hlinka D, Kalatova B, Uhrinova I, Dolinska S, Rutarova J, Rezacova J, Lazarovska S, Dudas M. Time-lapse cleavage rating predicts human embryo viability. Physiol Res. 2012;61:513–525. doi: 10.33549/physiolres.932287. [DOI] [PubMed] [Google Scholar]
- Hu Y, Maxson WS, Hoffman DI, Ory SJ, Eager S, Dupre J, Lu C. Maximizing pregnancy rates and limiting higher-order multiple conceptions by determining the optimal number of embryos to transfer based on quality. Fertil Steril. 1998;69:650–657. doi: 10.1016/s0015-0282(98)00024-7. [DOI] [PubMed] [Google Scholar]
- Hughes PM, Morbeck DE, Hudson S, Fredrickson J, Walker DL, Coddington CC. Peroxides in mineral oil used for in vitro fertilization: defining limits of standard quality control assays. J Assist Reprod Genet. 2010;27:87–92. doi: 10.1007/s10815-009-9383-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kallen B, Finnström O, Lindam A, Nilsson E, Nygren K, Olausson PO. Blastocyst versus cleavage stage transfer in in vitro fertilization: differences in neonatal outcome? Fertil Steril. 2010;94:1680–1683. doi: 10.1016/j.fertnstert.2009.12.027. [DOI] [PubMed] [Google Scholar]
- Kalra SK, Ratcliffe SJ, Barnhart KT, Coutifaris C. Extended embryo culture and an increased risk of preterm delivery. Obstet Gynecol. 2012;120:69–75. doi: 10.1097/AOG.0b013e31825b88fc. [DOI] [PubMed] [Google Scholar]
- Khan Z, Wolff HS, Fredrickson JR, Walker DL, Daftary GS, Morbeck DE. Mouse strain and quality control testing: improved sensitivity of the mouse embryo assay with embryos from outbred mice. Fertil Steril. 2013;99:847–854. doi: 10.1016/j.fertnstert.2012.10.046. [DOI] [PubMed] [Google Scholar]
- Kirkegaard K, Agerholm IE, Ingerslev HJ. Time-lapse monitoring as a tool for clinical embryo assessment. Hum Reprod. 2012;27:1277–1285. doi: 10.1093/humrep/des079. [DOI] [PubMed] [Google Scholar]
- Lane M, Gardner DK. Selection of viable mouse blastocysts prior to transfer using a metabolic criterion. Hum Reprod. 1996;11:1975–1978. doi: 10.1093/oxfordjournals.humrep.a019527. [DOI] [PubMed] [Google Scholar]
- Lane M, Gardner DK. Differential regulation of mouse embryo development and viability by amino acids. J Reprod Fertil. 1997;109:153–164. doi: 10.1530/jrf.0.1090153. [DOI] [PubMed] [Google Scholar]
- Lane M, Mitchell M, Cashman KS, Feil D, Wakefield S, Zander-Fox DL. To QC or not to QC: the key to a consistent laboratory? Reprod Fertil Dev. 2008;20:23–32. doi: 10.1071/rd07161. [DOI] [PubMed] [Google Scholar]
- Lemmen JG, Agerholm I, Ziebe S. Kinetic markers of human embryo quality using time-lapse recordings of IVF/ICSI-fertilized oocytes. RBM Online. 2008;17:385–391. doi: 10.1016/s1472-6483(10)60222-2. [DOI] [PubMed] [Google Scholar]
- Lonergan P, Rizos D, Gutierrez-Adan A, Fair T, Boland MP. Effect of culture environment on embryo quality and gene expression—experience from animal studies. RBM Online. 2003;7:657–663. doi: 10.1016/s1472-6483(10)62088-3. [DOI] [PubMed] [Google Scholar]
- McKiernan SH, Bavister BD. Fertilization and early embryology: timing of development is a critical parameter for predicting successful embryogenesis. Hum Reprod. 1994;9:2123–2129. doi: 10.1093/oxfordjournals.humrep.a138403. [DOI] [PubMed] [Google Scholar]
- Meseguer M, Rubio I, Cruz M, Basile N, Marcos J, Requena A. Embryo incubation and selection in a time-lapse monitoring system improves pregnancy outcome compared with a standard incubator: a retrospective cohort study. Fertil Steril. 2012;98:1481–1489. doi: 10.1016/j.fertnstert.2012.08.016. [DOI] [PubMed] [Google Scholar]
- Morbeck DE. Importance of supply integrity for in vitro fertilization and embryo culture. Semin Reprod Med. 2012;30:182–190. doi: 10.1055/s-0032-1311520. [DOI] [PubMed] [Google Scholar]
- Morbeck DE, Khan Z, Barnidge DR, Walker DL. Washing mineral oil reduces contaminants and embryotoxicity. Fertil Steril. 2010;94:2747–2752. doi: 10.1016/j.fertnstert.2010.03.067. [DOI] [PubMed] [Google Scholar]
- Morbeck DE, Fredrickson JR, Walker DL. Factors that affect mineral oil toxicity: role of oxygen and protein supplement. Fertil Steril. 2012;98:S29. [Google Scholar]
- Naz RK, Janousek JT, Moody T, Stillman RJ. Factors influencing murine embryo bioassay: effects of proteins, aging of medium, and surgical glove coatings. Fertil Steril. 1986;46:914–919. doi: 10.1016/s0015-0282(16)49834-1. [DOI] [PubMed] [Google Scholar]
- Nelissen EC, Van Montfoort AP, Coonen E, Derhaag JG, Geraedts JP, Smits LJ, Land JA, Evers JL, Dumoulin JC. Further evidence that culture media affect perinatal outcome: findings after transfer of fresh and cryopreserved embryos. Hum Reprod. 2012;27:1966–1976. doi: 10.1093/humrep/des145. [DOI] [PubMed] [Google Scholar]
- Otsuki J, Nagai Y, Chiba K. Peroxidation of mineral oil used in droplet culture is detrimental to fertilization and embryo development. Fertil Steril. 2007;88:741–743. doi: 10.1016/j.fertnstert.2006.11.144. [DOI] [PubMed] [Google Scholar]
- Pribenszky C, Losonczi E, Molnar M, Lang Z, Matyas S, Rajczy K, Molnar K, Kovacs P, Nagy P, Conceicao J, et al. Prediction of in-vitro developmental competence of early cleavage-stage mouse embryos with compact time-lapse equipment. RBM Online. 2010;20:371–379. doi: 10.1016/j.rbmo.2009.12.007. [DOI] [PubMed] [Google Scholar]
- Puissant F, Van Rysselberge M, Barlow P, Deweze J, Leroy F. Embryo scoring as a prognostic tool in IVF treatment. Hum Reprod. 1987;2:705–708. doi: 10.1093/oxfordjournals.humrep.a136618. [DOI] [PubMed] [Google Scholar]
- Rinaudo P, Schultz RM. Effects of embryo culture on global pattern of gene expression in preimplantation mouse embryos. Reproduction. 2004;128:301–311. doi: 10.1530/rep.1.00297. [DOI] [PubMed] [Google Scholar]
- Rivera RM, Stein P, Weaver JR, Mager J, Schultz RM, Bartolomei MS. Manipulations of mouse embryos prior to implantation result in aberrant expression of imprinted genes on day 9.5 of development. Hum Mol Genet. 2008;17:1–14. doi: 10.1093/hmg/ddm280. [DOI] [PubMed] [Google Scholar]
- Scott LF, Sundaram SG, Smith S. The relevance and use of mouse embryo bioassays for quality control in an assisted reproductive technology program. Fertil Steril. 1993;60:559–568. doi: 10.1016/s0015-0282(16)56176-7. [DOI] [PubMed] [Google Scholar]
- Strandell A, Bergh C, Lundin K. Selection of patients suitable for one-embryo transfer may reduce the rate of multiple births by half without impairment of overall birth rates. Hum Reprod. 2000;15:2520–2525. doi: 10.1093/humrep/15.12.2520. [DOI] [PubMed] [Google Scholar]
- Taft RA. Virtues and limitations of the preimplantation mouse embryo as a model system. Theriogenology. 2008;69:10–16. doi: 10.1016/j.theriogenology.2007.09.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong CC, Loewke KE, Bossert NL, Behr B, De Jonge CJ, Baer TM, Reijo Pera RA. Non-invasive imaging of human embryos before embryonic genome activation predicts development to the blastocyst stage. Nat Biotechnol. 2010;28:1115–1121. doi: 10.1038/nbt.1686. [DOI] [PubMed] [Google Scholar]