

Abstract
The crustacean Hyperglycemic Hormone (cHH) is a neuropeptide present in many decapods. Two different chiral isomers are simultaneously present in Astacid crayfish and their specific biological functions are still poorly understood. The present study is aimed at better understanding the potentially different effect of each of the isomers on the hepatopancreatic gene expression profile in the crayfish Pontastacus leptodactylus, in the context of short term hyperglycemia. Hence, two different chemically synthesized cHH enantiomers, containing either L- or D-Phe3, were injected to the circulation of intermolt females following removal of their X organ-Sinus gland complex. The effects triggered by the injection of the two alternate isomers were detected after one hour through measurement of circulating glucose levels. Triggered changes of the transcriptome expression profile in the hepatopancreas were analyzed by RNA-seq. A whole transcriptome shotgun sequence assembly provided the assumedly complete transcriptome of P. leptodactylus hepatopancreas, followed by RNA-seq analysis of changes in the expression level of many genes caused by the application of each of the hormone isomers. Circulating glucose levels were much higher in response to the D-isoform than to the L-isoform injection, one hour from injection. Similarly, the RNA-seq analysis confirmed a stronger effect on gene expression following the administration of D-cHH, while just limited alterations were caused by the L-isomer. These findings demonstrated a more prominent short term effect of the D-cHH on the transcription profile and shed light on the effect of the D-isomer on specific functional gene groups. Another contribution of the study is the construction of a de novo assembly of the hepatopancreas transcriptome, consisting of 39,935 contigs, that dramatically increases the molecular information available for this species and for crustaceans in general, providing an efficient tool for studying gene expression patterns in this organ.
Introduction
The freshwater astacid Pontastacus leptodactylus, commonly called narrow-clawed crayfish, inhabits Western Asian and Eastern European lakes and watercourses. Its reproductive season includes mating from December to January and spawning in January. Eggs are incubated glued to the female pleopods till July [1]. Ovarian development takes place from June to November [2]. Induced molt through chirurgical intervention in P. leptodactylus maintained at 19°C, led to a premolt period of 17 days ending with ecdysis (personal observations of the present authors). Adults naturally shed their exoskeleton in summer just after hatching, even though some of them molt also in autumn [3].
Crustacean hyperglycemic hormones (cHHs) are a pleiotropic crustacean-specific neuropeptide family, functioning in a variety of physiological processes, recently reviewed by several authors [4], [5], [6], [7], [8]. The cHH family is divided into two subfamilies on the basis of their primary structure: (a) the cHH subfamily and (b) the molt-inhibiting hormone (MIH), the mandibular organ inhibiting hormone (MOIH) and the vitellogenesis/gonad-inhibiting hormone (V/GIH) subfamily. The translated neuropeptides in the tissues as well as their isoforms derived from post-translational modifications and their modes of action have only been partially described. Hence, a neurohormone name does not necessarily imply its entire range of functions. Several cHH variants occasionally co-exist in a single species. The variability can emerge either from a different primary sequence or from different post-translational modifications [9], [10]. Recently, chirality was observed also in a lobster VIH due to L to D alteration in the fourth N terminal amino acid, a tryptophan residue [11] demonstrating larger extent of the phenomenon in crustaceans. cHHs are produced in the neurosecretory perikarya sited in the medulla terminalis of the optic ganglion, located in the crustacean eyestalk and named X-organ. The X-organ secretes the neuropeptides into the hemal sinus gland and the entire neuroendocrine complex is abbreviated XOSG. Structurally, the cHH prepropeptide is composed of a signal peptide, a cHH precursor related peptide (CPRP) and a mature peptide of 72 amino acids. The role of the CPRP is still unknown, but CPRP structures, post-translational modifications and individual-related distribution have already been described [12], [13]. The mature cHH contains six cysteine residues that form three disulfide bridges and potentially possesses an amidated C-terminus and a pyroglutamate blocked N-terminus [6]. Documented physiological processes influenced by the eyestalk ablation are vitellogenesis [14], food intake, digestion, and nutrient transport [15], molting [16], metabolism of lipids [17], [18], regulation of glucose and proteins in hemolymph [17], [19], hydromineral balance, regeneration and pigment regulation [20]. Several cHH family members generally have an inhibitory effect, as the removal of the XOSG causes induction of both molt and reproduction. cHHs are produced also in other tissues: the pericardial organ, the subesophageal ganglia, and the fore- and hindguts. The more comprehensively studied action of the cHH is the regulation of carbohydrate metabolism. Its secretion follows a circadian rhythm, with a low concentration during the day which increases in the first hours of the night, and it is correlated to a similar daily pattern of the glycemia [21]. The injection of cHH induces a fast pharmacological hyperglycemic response in treated animals. Apart from glucose metabolism, cHH mediates other metabolic functions of the hepatopancreas, the site of synthesis and secretion of digestive enzymes (amylases, proteases, lipases and others) [22], [23], [24]. It is also involved in metabolism of proteins, lipids, and carbohydrates [25], as well as in the catabolism of organic compounds and in detoxification [26], [27]. Indeed, cHH stimulates amylase secretion [28], and the release of free fatty acids and phospholipids [18] from the midgut gland. D-cHH is also involved in the control of molt, exerting its function by inhibiting the synthesis of ecdysone in the Y-organ and having an activity 10 times higher than L-cHH [29]. Contrasting activities were reported for the CHH regulation of reproduction, probably due to species specificity [30], [31], [32], [33].
The structure of the cHH genes, the derived precursors and peptides have been recently reviewed by Webster and colleagues [7], while the dynamics of biosynthesis and release of cHH isoformes in Orconectes limosus have been clarified by Ollivaux and Soyez [34]. cHH peptide sequences were de novo sequenced through a multifaceted mass spectrometry approach by Jia and colleagues [35].
Although P. leptodactylus is not considered a model organism, it is one of the most studied species among decapods with about 143 published papers regarding its physiology [1], [36], [37], [38], its resistance to different types of pollution and stress conditions [39], [40], [41] and concerning the cHH and its variety of functions [42], [43], [44], [45]. Unfortunately this variety of studies does not reflect the fast progress of genomics and transcriptomics. Back in 1988 Sedlmeier [28] demonstrated that cHH increased amylase secretion from the hepatopancreas of O. limosus. He also identified the involvement of the second messengers: cAMP, cGMP and calcium ion and a participating adenylate cyclase. However, no effect on gene expression was examined and no distinction was made between the two O. limosus cHH isomers, discovered later [34]. The present study is aimed at defining the effect of cHH at the hepatopancreas transcriptome level distinguishing the putatively different effect of the two stereomers.The effect of the two isomers of cHH on gene expression in relation to their variety of assigned functions in the relevant tissues is still lacking. The working hypothesis for this study assumed two alternate or complementary effects of each of the two hormones: changes in gene expression or activation/inactivation of participating proteins. Elevation of circulating glucose level upon application of D and L-cHH to eyestalkless P.leptodactylus even one hour post-injection is well documented [42]. Hence, the study is aimed at examining the two alternatives during this short time span by simultaneously measuring the circulating glucose levels and changes of gene expression in the hepatopancreas applying a transcriptome approach. Beside this gene expression experiment, the construction of a de novo assembly of the hepatopancreas transcriptome, dramatically increased the molecular information available for this species and undoubtedly provides a useful tool for further gene expression experiments in this organ.
Materials and Methods
Crayfish maintenance and experimental design
Adult P. leptodactylus were bought from a local dealer on July, 2011. They were kept for two weeks before the experiment in 120 L tanks provided with closed circuit filtered and thoroughly aerated tap water at ∼18°C and were fed by fish pellets (Sera granular, Heisenberg, Germany) three times per week. Only females with no abdominally incubated ova were taken for the induction experiment. The females were at the end of the ova incubation period.
The chemical synthesis of the peptides and the glucose level induction protocol were accomplished according to Mosco et al. [42] with a few modifications detailed below. The two synthetic cHH hormone isomers, D-cHH and L-cHH, were injected to the circulation. Thirty-two P. leptodactylus females were divided into four groups each composed of 8 females. Two of the groups were injected with D- and L-cHH, respectively (0.5 µg/female in 100 µl PBS). A control group was sham-injected by the hormone carrier (S). Bilateral ablation of the XOSG, aimed at prevention of any possible interference due to endogenous cHH was performed 48 hours before the injections, and a fourth group of naïve females (N) was added to the experiment as control just before injections. All 32 females were sacrificed one hour post-injection. The incubation period included ten minutes of anesthesia in ice before sacrifice and all efforts were made to minimize the suffering of the animal. Immediately prior to the hormonal injection the females were bled for the evaluation of the pre-induction hemolymphatic glucose level. A second bleeding was performed just before sacrifice. Hemocytes were pelleted from the sampled hemolymph and the serum was kept on ice for later glucose measurement, which was performed using a glucose oxidase method (glucose liquid mono reagent, Hospitex diagnostics, Italy). The statistical analyses of glucose levels recorded in the four groups and females' morphometric parameters were performed using R software, version 2.14.1 [46] as follows: the normality of data was checked with a Shapiro-Wilk test and homogeneity of variance across groups was checked with a Bartlett test. The null hypotheses of both tests could not be rejected. Hence, differences of glucose levels among the experimental groups were tested using non-parametric statistics, Kruskal-Wallis rank sum test with post-hoc Wilcoxon rank sum test pairwise comparisons with Bonferroni correction. Box and whiskers plots were drawn with the boxplot command of R. Glucose levels are expressed as mean ± standard error. Carapace length was measured from its posterior edge to the base of the eye cavity just before sacrifice and a sample of hepatopancreatic tissue of ∼5×5×5 mm was dissected out and immediately snap-frozen in liquid nitrogen. The gastroliths were also taken for molt stage evaluation according to Shechter et al. [47] using the molt mineralization index (MMI) (gastrolith width/crayfish carapace length).
Ethical Note
The experiments comply with the current laws of Italy, the country in which they were done. No specific permits were required for the studies that did not involve endangered or protected species. Individuals were maintained in appropriate laboratory conditions to guarantee their welfare and responsiveness. After the experiments were completed, crayfish were sacrificed by hypothermia.
RNA extraction and sequencing
Twelve out of 32 females were randomly chosen for the RNA sequencing. RNA was extracted from frozen tissues, homogenized in TriReagent RNA isolation solution (Sigma-Aldrich, Cat UN2821) following the manufacturer's instructions. The resulted RNAs were further purified using the RNeasy kit (Qiagen, manufacturer's instructions). The RNA level was quantified by spectrophotometer and its quality was examined using capillary electrophoresis (BioAnalyzer 2100, Agilent).
RNA sequencing was carried out at the Applied Genomics Institute (IGA, Udine, Italy), on an Illumina sequencer HiSeq2000. The hepatopancreas reference transcriptome assembly was derived from a 2×100 bp paired-end sequencing performed on a cDNA library obtained from equimolar amounts of RNA taken from all experimental females. Gene expression was evaluated using the single-end 50 bp Illumina sequencing from the 12 distinct RNAs.
Sequences analysis
The processing and analysis of the obtained raw sequences was carried out using the CLC Genomics Workbench 4.5 software (CLC Bio, Aarhus, Denmark). Raw sequence reads were trimmed according to base calling quality. The resulting 2×100 bp sequence reads were assembled assuming a paired reads distance between 100 and 600 base pairs, and setting the penalties for mismatches to 2, insertions and deletions to 3 and similarity and length fraction to 0.9 and 0.5, respectively. The minimum allowed assembled contig length was set to 200 bp. The obtained contig assembly served as a comprehensive reference for the functional genomics RNA-seq analysis [48] of the P. leptodactylus hepatopancreas. We determined the presumptive amount of conceptual full length transcripts by using the Full-lengther Next webtool [49] considering alignments starting before the 10th aa and with an e-value below 1e-04. Filtered contigs displaying an average coverage lower than 25× of mapped 50 bp reads were discarded prior to the RNA-seq analysis for creating a robust set of contigs not subject to random expression fluctuations, corresponding to the 95th percentile of the genes expressed in the hepatopancreas throughout the experiment.
The 50 bp sequencing reads from the 12 different samples (namely, N1, N2, N3, S1, S2, S3, L1, L2, L3, D1, D2 and D3) were individually mapped on the filtered reference set using the RNA-seq procedure, considering a maximum number of mismatches cost of 3 and a maximum of 10 hits for a read. The expression values were calculated based on unique gene reads. The Baggerly's test [50] was used to identify statistically significant differential expression using S as a reference group; a FDR corrected p-value<0.01 was set as threshold of significant differential expression [51] and an additional minimum threshold of fold change of 2 was also used. The similarity among the profiles obtained from this study was examined by hierarchical clustering (complete linkage, average linkage and single linkage) using the Pearson correlation coefficient as a distance measure. An alternative clustering was performed by Principal Component Analysis (PCA). The raw Illumina reads were stored at the NCBI Sequence Read Archive (SRA: SRR650486), whereas 39,935 assembled contigs were deposited at NCBI Transcriptome Shotgun Assembly (TSA: GAFY00000000).
Transcripts annotation and their expression pattern
Differentially expressed transcripts were characterized with the Blast2Go platform [52], [53]. The characterized parameters were: resemblance to genes with known function using BLASTx algorithm [54] against the NCBI non-redundant protein databases applying an e-value cut-off of 106. The default Blast2Go similarity annotations were also checked manually against the list of resemblances obtained for each contig by BLASTx, to conform to the UniProt nomenclature guidelines and to select the annotations in a more educated manner, based on the GO terms and domain characterization described below. Blast2Go was used also to assign Gene Ontology (GO) functional terms [55] and Interpro domains annotations [56] to the contigs, with default settings. The GO and Interpro annotation outputs were modified with scripts developed in-house and imported into the CLC Genomics Workbench 4.5 environment for implementing the hypergeometric test [57] on the annotations. Significantly altered GO terms and Interpro domains were detected with this test considering a p-value threshold of 0.01 and a difference between observed and expected >1.
Results and Discussion
Morphological and physiological state of the animals
The morphometric characteristics and the molt stage of the experimental females as well as their hemolymphatic glucose levels before injection and at their sacrifice are presented in Table 1 . The glucose level of all the 3 groups of eyestalk-less females was highly significantly lower than the native ones (Wilcoxon rank sum test: p<0.01; Figure 1A ), but after an hour the injection of both cHHs was able to restore glycemia to almost normal levels (D-cHH and L-cHH injected animals vs native animals, Wilcoxon rank sum test: p = 1; Figure 1B ). On the contrary, the sham-injected animals still presented a significantly lower glycemia compared to all other groups (Wilcoxon rank sum test: p<0.02). The carapace length was similar among the four experimental groups, namely N, S, L and D (Wilcoxon rank sum test: p>0.05). All females were at intermolt with MMI<0.01 and all ovaries were immature according to Hubenova et al. [2].
Table 1. Morphometric characteristics of the experimental females, their hemolymphatic glucose levels before injection and at their sacrifice.
| Experimental group [n = 8 in each group] | Carapace length [mm] | *MMI – Gastrolith width/Carapace length [mm/mm] | Glucose levels [mg/dL] | |
| 0 min. | 60 min | |||
| Native | 39.4±1.2 | 0.0067±0.0048 | 9.14±0.75 | 14.78±1.93 |
| Sham-injected | 39.4±2.4 | 0.0094±0.0086 | 1.94±0.33 | 1.01±0.59 |
| L-cHH-injected | 41.5±2.0 | 0.0081±0.0065 | 2.36±1.10 | 13.84±3.13 |
| D-cHH-injected | 39.9±1.7 | 0.0025±0.0001 | 2.49±0.83 | 11.45±2.32 |
MMI – molt mineralization index.
Figure 1. Box plot of the glycemia values obtained in the four groups S – sham injected, N – intact, L – L-cHH-injected, D- D-cHH-injected at the beginning of the experiment and before sacrifice, one hour post-injection.

De novo assembly of the P. leptodactylus hepatopancreas transcriptome
The Illumina 2×100 bp sequencing of the hepatopancreas of adult P. leptodactylus females, generated 296,112,296 nucleotide reads. The number was reduced to 289,731,590 after quality trimming. The de novo assembly by the CLC Genomic Workbench produced 42,187 contigs with an average length of 802 bp. This assembly was used as a reference transcript library for subsequent analyses. A total of 9,474 contigs longer than 1 Kb was obtained, while the longest contig almost reached 15 Kb in length. Table 2 summarizes the trimming and assembly statistics. The raw Illumina reads were stored at the NCBI Sequence Read Archive (SRA: SRR650486), whereas 39,935 assembled contigs were deposited at NCBI Transcriptome Shotgun Assembly (TSA: GAFY00000000). The reduction in number of deposited contigs was obliged by the TSA quality control process which includes additional trimming of primers, shortening of long N rows artificially produced by the definition of the assembly parameters which rendered some contigs shorter than 200 bps. Moreover, some genes suspected to be introduced by contamination were deleted as well. Through the Full-Lengther Next webtool [49] we determined the presumptive percentage of the full length transcripts present in our library. Hence, 1/3rd (32.2%) of the transcripts were putatively assembled to their full length, 1/3rd (32.6%) covered either the N-terminal or the C-terminal region, whereas the remaining 35.2% consisted of internal fragments.
Table 2. Trimming statistics and assembly summary of the Reference dataset.
| Trimming statistics | |
| Number of reads before trimming | 296,112,296 |
| Number of reads after trimming | 289,731,590 |
| Paired-end reads after trimming | 283,857,480 |
| Single reads after trimming | 5,874,110 |
| Sequences discarded during trimming | 6,380,706 |
| Average length before trimming | 101.0 |
| Average length after trimming | 97.2 |
N50, 80 and 90 are the quantiles corresponding to the 50, 80 and 90 percentiles, respectively.
From the output of Blast2Go we checked the thirteen Top-hits species sharing similarity with the P. leptodactylus reference library. The two most represented species were Daphnia pulex (Crustacea) and Tribolium castaneum (Insecta). The first thirteen BLAST top-hits species are reported in Figure S1. To date, the only crustacean whose genome has been fully sequenced is the branchiopod Daphnia pulex [58]. Even though a few deep transcriptome sequencing approaches were applied to decapods [59], [60], no study has ever targeted Astacidae. An astacoid epithelial assembly from Cherax quadricarinatus, performed by 454 was simultaneously prepared by the present co-authors and their collaborators and will be soon released for the public (TSA: GADE00000000). At present, only 275 nucleotide sequences belonging to the genus Pontastacus are stored at NCBI, while only 67 originated from Pontastacus leptodactylus. Next generation sequencing greatly simplifies large-scale molecular studies of non-model organisms, and this transcriptome sequencing with the assembly of 39,935 contigs, represents the first large-scale sequencing approach in this genus, as well as a remarkable contribution to the genetic knowledge of decapods, paving the way to straightforward comparative molecular studies on non-model crustaceans.
RNA-seq analysis of cHH isomer effects
An average of 12.7±5.2 million short reads were obtained from the sequencing of RNAs extracted from the hepatopancreas of each sampled female. An average of 50.8±6.9% reads were mapped to the reference transcriptome. Table 3 reports in detail the number of reads obtained from each adult female. The RNA-seq mappings of the 12 sequenced samples (N1, N2, N3, S1, S2, S3, L1, L2, L3, D1, D2 and D3) were used for the gene expression analysis. Only a selected set of 6,860 contigs with an average coverage >25× was used for the analysis, reducing the noise potentially caused by the high number of fragmented or poorly expressed transcripts, not strictly related to the hepatopancreas, as highlighted by the Full-lengther analysis. This selected set of transcripts accounts for about 96% of the total expression in this organ during the present experiment. The stringency applied to the contigs selection (FDR corrected p-value <0.01 and a weighted fold change at least of 2), identified the prominently differentially expressed transcripts in the pairwise comparisons, by using the S group as reference for all the analyses. Table 4 summarizes the numbers of differentially expressed genes while the complete list of differentially expressed genes is available in Table S1. The expression profiles of the various females, namely the list of mapped reads per each gene in each sample were clustered by two methods, hierarchical clustering (Figure S2A) and a principal component analysis (Figure S2B), which highlighted a large difference between D-cHH-injected animals and all the other groups, which in turn clustered together and did not display detectable differences in this preliminary analysis.
Table 3. Total number of sequencing reads obtained per animal and the number of the reads mapped and the corresponding percentage.
| Sample | Total number of reads | Mapped reads (%) |
| N1 | 23,784,636 | 13,086,802 (55.0%) |
| N2 | 10,985,623 | 5,738,820 (52.2%) |
| N3 | 17,933,507 | 9,093,453 (50.7%) |
| S1 | 8,589,134 | 4,239,139 (49.4%) |
| S2 | 12,303,830 | 5,904,274 (48.0%) |
| S3 | 14,403,132 | 6,722,862 (46.7%) |
| L1 | 10,579,155 | 6,740,537 (63.7%) |
| L2 | 13,929,531 | 7,154,589 (51.4%) |
| L3 | 11,168,774 | 6,111,228 (54.7%) |
| D1 | 6,239,023 | 2,596,219 (41.6%) |
| D2 | 12,781,748 | 4,431,787 (34.7%) |
| D3 | 7,379,961 | 3,173,378 (43.0%) |
N = native group, S = sham-injected, L = L-cHH injected and D = D-cHH injected.
Table 4. Number of differentially expressed transcripts in each of the experimental groups.
| Group | Number of differentially expressed transcripts | Up regulated | Down regulated |
| S vs N | 214 | 147 | 67 |
| L vs S | 45 | 30 | 15 |
| D vs S | 917 | 60 | 857 |
FDR corrected p-value <0.01, minimum difference in fold change = 2. S = sham-injected, L = L-cHH injected and D = D-cHH injected.
Effects of XOSG removal on the hepatopancreas gene expression
The comparison between the S and N groups was useful to depict the effects derived solely from the XOSG removal procedure. XOSG extirpation resulted in the differential expression of 214 transcripts, with up-regulation (147 sequences, 68.7%) exceeding down-regulation (67 sequences, 31.3%). It is likely that the transcripts characterized by an increase of expression in respect to the naïve state are connected to two main events, the injury and the reduced production of several hormones caused by the XOSG removal. 63% of the up-regulated and 67% of the down-regulated genes showed no resemblance to known sequences. Among the genes which demonstrated similarity we identified in particular some transcripts connected to glycolysis process, such as transcript containing enolase domain (IPR000941), probably connected to the removal of cHHs production sites.
The hypergeometric test highlighted the carbohydrate metabolism (GO:0005975) and the chitinase activity (GO:0004568) as the most relevant GO terms repressed by XOSG extirpation; and many associated Interpro domains, e.g. glycoside hydrolase, family 9 (IPR001701), six-hairpin glycosidase (IPR012341) and chitinase II (IPR011583) were significantly repressed. Many other GO terms and Interpro signatures were found to be up-regulated, even though most of them could not be easily linked to any reported physiological effect of the extirpation. The most studied process in many crustacean species triggered by XOSG extirpation is undoubtedly vitellogenesis [61], [62], [63], but there is no uniform vitellogenesis-related physiological response to the extirpation among the studied species.
As for other genes, in pandalid shrimp, eyestalk ablation down-regulated the hepatopancreatic chitinase expression and also in P. leptodactylus we observed a significant repression of the IPR domain of the chitinase II (IPR011583) within the S group. This transcript pertains to the group 1 reported in Salma et al. [65] that represent chitinases produced in hepatopancreatic tissues that may function in the digestion of ingested chitin and the modification of peritrophic membrane in the intestine. Other studies [15], [64] showed that XOSG extirpation caused an increase in respiratory rate and a decrease in metabolic activity with respect to intact shrimps. Similarly, in P. leptodactylus the repression of six transcripts associated to metabolic process (GO:0008152) was observed in the S group: differently from Marsupenaeus japonicus [65] in which fructose-1,6-bisphosphatase was not significantly changed after XOSG removal, we observed an up-regulation of a transcript containing the fructose-1,6-bisphosphatase domain (IPR000146). This gene is involved in gluconeogenesis and cHH could affect also this metabolic pathway.
A similar although somewhat different experiment was carried out in the crayfish Cherax quadricarinatus by Yudkovski et al. [66]. Hepatopancreatic gene expression was compared between eyestalk extirpated and intact males. This experiment made use of a microarray analysis and differs from the present one by the time elapsed from extirpation to sacrifice (nine days vs two days here) and by the number of the reference genes which participated in the analysis (1865 vs 6,860 selected contigs here). Bearing in mind these differences, chitinases and carbohydrate metabolism-related enzymes were down-regulated in both studies. In addition, proteolytic enzymes and several hemocyanins were attenuated in the C. quadricarinatus experiment in contrary to the present experiment in which they were not differentially expressed. In general, the reduction in the hepatopancreas metabolic activity was more pronounced nine days after ablation than in P. leptodactylus two days after ablation.
XOSG removal affects also the circulating levels of hormones other than the cHH. These include the cHH type II hormones [7], the Molt Inhibiting Hormone (MIH), the Gonad/Vitellogenesis Inhibitng Hormone (GIH/VIH) and the Mandibular Organ-Inhibiting Hormone (MOIH). Hence, pathways controlled by these hormones may potentially be affected by the extirpation, an effect that would be difficult to discriminate due to the pleiotropic nature of the cHH members, affecting multiple processes [66].
Differential effects of the two cHH-isomers on hepatopancreatic gene expression
The injection of the two cHHs, distinguished only by chirality, showed different effects on gene expression. The overall difference can be clearly demonstrated by the results of the clustering procedures revealing relatively different expression profile of the D-cHH-treated group in comparison to the other three groups, and in turn elucidated much more similar expression profiles with each other (Fig. S2). More in detail, comparison with the S group revealed only 45 differentially expressed transcripts in response to the administration of the L-isomer, whereas 917 transcripts were responsive to the D-isomer. Among the transcripts differentially up-regulated after L-cHH injection we depicted an increase of aminopeptidase n-like and trypsin 4, both correlated to proteolytic activity (GO:0006508), while all the down-regulated ones were without similarity. Moreover no functional or structural feature (GO term or domain) could be significantly linked to L-cHH injection by the hypergeometric test.
To examine a possible overlap in the effects exerted by L- and D-cHH we identified differentially expressed transcripts shared by the two treatments. Six transcripts elucidated differential expression caused by both treatments, but the majority of them had different trends of expression. No transcripts shared up-regulation caused by both treatments, while 2 non-annotated transcripts were mutually down-regulated (Pastle_hepa_c8804 and Pastle_hepa_c22565). Three transcripts were up-regulated after L-cHH, but down-regulated following D-cHH injection; among them a trypsin 4, a transcript containing chitinase II domain (IPR011583) and one without similarity. Only one non annotated transcript resulted to be up-regulated in D and down-regulated in the L group.
In the light of the results described above, the analysis was mainly focused on the variations between the S and the D groups.
Table 5 presents the Interpro domains and the GO terms altered in the in D group in comparison to the S group as revealed by the hypergeometric test.
Table 5. List of the Interpro domains and GO terms differentially regulated by XOSG removal (S) or by D-cHH-injection (D), detected by the Hypergeometric test.
| Group | Trend | Annotation | p-value | Description |
| S vs N | + | IPR006195 | 0.000117 | Aminoacyl-tRNA synthetase, class II |
| S vs N | + | IPR002930 | 0.000979 | Glycine cleavage H-protein |
| S vs N | + | IPR004365 | 0.000979 | Nucleic acid binding, OB-fold, tRNA/helicase-type |
| S vs N | + | IPR020809 | 0.000979 | Enolase, conserved site |
| S vs N | + | IPR020810 | 0.000979 | Enolase, C-terminal |
| S vs N | + | IPR000537 | 0.00288 | UbiA prenyltransferase family |
| S vs N | + | IPR000941 | 0.00288 | Enolase |
| S vs N | + | IPR004364 | 0.00288 | Aminoacyl-tRNA synthetase, class II (D/K/N) |
| S vs N | + | IPR003604 | 0.00288 | Zinc finger, U1-type |
| S vs N | + | IPR018150 | 0.00288 | Aminoacyl-tRNA synthetase, class II (D/K/N)-like |
| S vs N | + | IPR011053 | 0.00288 | Single hybrid motif |
| S vs N | + | IPR001790 | 0.00564 | Ribosomal protein L10/acidic P0 |
| S vs N | + | IPR016027 | 0.00681 | Nucleic acid-binding, OB-fold-like |
| S vs N | + | IPR012340 | 0.00681 | Nucleic acid-binding, OB-fold |
| S vs N | + | IPR012675 | 0.00921 | Beta-grasp domain |
| S vs N | + | GO:0005960 | 0.000979 | Glycine cleavage compleX |
| S vs N | + | GO:0004659 | 0.000979 | Prenyltransferase activity |
| S vs N | + | GO:0030414 | 0.00288 | Peptidase inhibitor activity |
| S vs N | + | GO:0000287 | 0.00847 | Magnesium ion binding |
| S vs N | + | GO:0004634 | 0.00921 | Phosphopyruvate hydratase activity |
| S vs N | + | GO:0004867 | 0.00921 | Serine-type endopeptidase inhibitor activity |
| S vs N | − | IPR001701 | 1.36E-05 | Glycoside hydrolase, family 9 |
| S vs N | − | IPR012341 | 1.89E-05 | Six-hairpin glycosidase |
| S vs N | − | IPR008928 | 1.89E-05 | Six-hairpin glycosidase-like |
| S vs N | − | IPR017853 | 0.000453 | Glycoside hydrolase, superfamily |
| S vs N | − | IPR011583 | 0.0023 | Chitinase II |
| S vs N | − | IPR001223 | 0.0023 | Glycoside hydrolase, family 18, catalytic domain |
| S vs N | − | IPR013781 | 0.00314 | Glycoside hydrolase, catalytic domain |
| S vs N | − | GO:0005975 | 0.000496 | Carbohydrate metabolic process |
| S vs N | − | GO:0043169 | 0.000584 | Cation binding |
| S vs N | − | GO:0003824 | 0.000858 | Catalytic activity |
| S vs N | − | GO:0004568 | 0.0023 | Chitinase activity |
| D vs S | + | IPR013128 | 0.0000548 | Peptidase C1A, papain |
| D vs S | + | IPR013201 | 0.000777 | Proteinase inhibitor I29, cathepsin propeptide |
| D vs S | + | IPR000668 | 0.0024 | Peptidase C1A, papain C-terminal |
| D vs S | + | GO:0008234 | 0.000113 | Cysteine-type peptidase activity |
| D vs S | + | GO:0004197 | 0.00544 | Cysteine-type endopeptidase activity |
| D vs S | − | IPR000941 | 0.0058 | Enolase |
| D vs S | − | IPR011042 | 0.0058 | Six-bladed beta-propeller, TolB-like |
| D vs S | − | GO:0006096 | 0.000818 | Glycolysis |
| D vs S | − | GO:0030234 | 0.00444 | Enzyme regulator activity |
+ designates up-regulation and − down-regulation.
As previously shown by the PCA analysis, the D-cHH administration caused pronounced alterations in comparison to the sham injected females, dominated by down-regulated genes (857 transcripts, accounting for 93.5% out of the total). The hypergeometric test elucidated a few D-cHH up-regulated functions, and in particular several GO terms related to peptidases, including cysteine-type peptidase (GO:0008234) and endopeptidase activity (GO:0004197), the papain domain of peptidase C1A, (IPR013128) and the cathepsin propeptide domain of proteinase inhibitor I29 (IPR013201), which is usually found in the N-terminus of many proteinases.
On the contrary, glycolysis (GO:0006096) appeared to be the GO term most significantly repressed in the D-cHH-injected female crayfish, which may be related to the return of the glycemic level to the native state, and suggests a modulation activity of the D-cHH on the glucose and energy metabolism in the hepatopancreas.
Given the absence of the source of endogenous cHH, it was of interest to assess whether the injection of exogenous hormone was able to restore, even partially, the native conditions. Obviously, XOSG-ablation does not only remove the primary organ of cHH synthesis (cHH type I), but also prevents the secretion of other cHH superfamily hormones (cHH type II), possibly causing a series of gene expression changes not solely related to the deprivation of cHH itself. Therefore, the administration of synthetic cHH was only expected to partially restore the gene expression profile observed prior to XOSG ablation. In fact, about 1/3th (50 out of 147) of the up-regulated transcripts in the S group were restored at lower level following the D-cHH injection ( Figure 2A ). Among them transcripts related to several different cellular processes and molecular functions were identified, including cellular organization (i. e. actin and tubulin beta 2), transcripts related to RNA processing and protein synthesis, (i.e, mRNA turnover protein 4-like protein), a transcript containing Fibrillarin domain (IPR000692) that plays a role in ribosomal RNA processing and lysyl-tRNA synthetase. Two cuticle proteins were also detected, playing a role in calcium ion binding; an armet-like protein precursor containing EF-Hand 1, calcium-binding site domain (IPR018247) and crustin 1 antimicrobial peptide were also restored by the D-cHH injection. Figure 2A shows two transcripts with an expression fold-change higher than the others, one encoding a C-reactive protein (CRP) containing a pentaxin domain (IPR001759) and the other transcript coding for a cuticle protein which contains the Rebers and Riddiford arthropod chitin binding domain (IPR000618). CPR is a fundamental player in the inflammatory response, in particular binding to the phosphocholine expressed on the surface of dead or dying cells in order to activate the complement system via the C1Q complex. Eyestalk ablation did not cause an increase in the level of CPR mRNA in M. japonicus in the ovary [67]. In the present study and in a different tissue an up-regulation of about 9.5-fold was depicted following the XOSG-ablation, subsequently restored by the D-cHH administration. But most notably, the expression of four transcripts crucially involved in glycolysis, which were increased in the S group, returned to lower levels after the D-cHH administration. Namely, phosphoglycerate mutase 1, two highly similar enolases catalyzing the eighth and ninth steps of glycolysis respectively, and fructose-1,6-bisphosphatase, an allosteric regulator of pyruvate kinase, the final step of glycolysis, all followed the same expression trends. This supports the hypothesis that D-cHH could have mainly an inhibitory effect on transcripts up-regulated following XOSG ablation and a minor effect on those silenced by the ablation, which probably required other molecules and centers of control located in the removed eyestalk. In fact, only 5% of the repressed genes upon ablation (3 out of 60) returned to the native levels of expression after D-cHH administration ( Figure 2B ). One of them resembles the 3-hydroxybutyrate dehydrogenase, related to oxydoreductase and metabolic processes and the two remaining transcripts have an unknown function.
Figure 2. Expression trend of transcripts altered by XOSG ablation and restoring their native level following D-cHH injection.
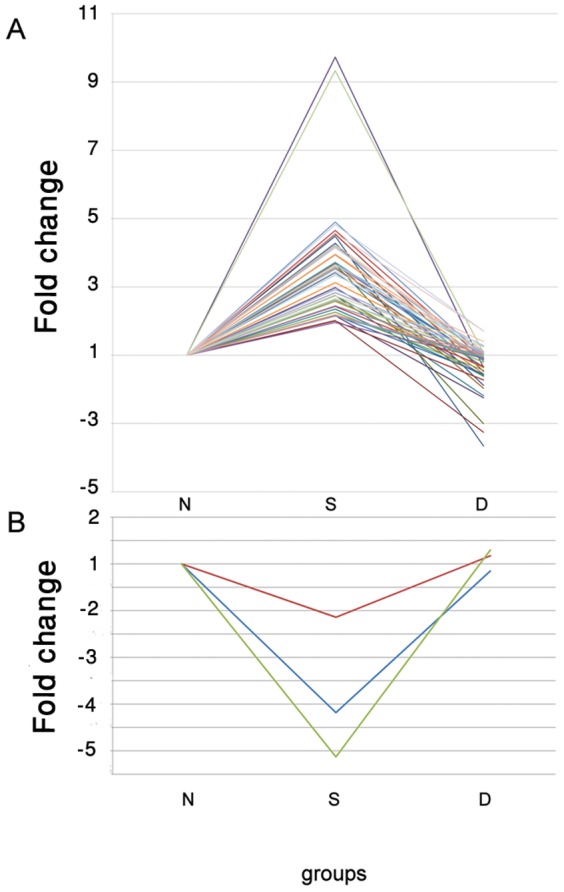
A Up-regulated transcripts following XOSG ablation. B Down-regulated transcripts following XOSG ablation.
The L-configuration provides the peptide with a strictly hyperglycemic activity whereas the D-cHH may exhibit other functions, in addition to a strong hyperglycemic potency. The finding of several transcripts containing a sodium symporter domain (IPR002657) suggests that D-cHH may also regulate glucose absorption by the hepatopancreas, a process that occurs through a Na+/D-glucose co-transport [68].
It has to be emphasized that the experiment was not intended to mimic natural situation in which the two hormonal species may be present in different levels, not necessarily reflected the injected amounts, but to apply rather pharmacological dose aimed at distinguishing the effects of the two isoforms. The data all together suggest that, despite being apparently able to restore glycemia to almost normal levels in eye-ablated animals one hour post injection (see Table 1 ), L-cHH does not significantly influence gene expression in the hepatopancreas. Therefore the apparent positive effect of the two neuropeptides on glycemia, at least after a short period, is either provided by a specific action of different tissues, or more likely still on the hepatopancreas, but through different mechanisms, one involving gene expression (D-cHH) and the other involving post translational enzyme activation (L-cHH). It is also possible that the effect of L-cHH on gene expression would be only visible at longer periods from its injection, after more than one hour. An additional experiment, similar to the one described here, but implementing a prolonged time-course and sampling of additional tissues has already been performed in our laboratory, and is currently being analyzed to provide better insight to the presented alternative hypotheses.
Conclusions
The effects of two cHH enantiomers on a multi-gene expression profile of P. leptodactylus were examined here. The presence of both cHH isomers has been demonstrated only in Astacoidea [69]. The scarcity of P. leptodactylus sequenced transcripts required the construction of a comprehensive transcriptomic library of its hepatopancreas, comprising 39,935 contigs obtained through Illumina sequencing. This library was used as a reference gene assembly in a RNA-seq gene expression experiment. The short term effect of two enantiomers of the peptidic hyperglycemic hormone (D-cHH and L-cHH) of P. leptodactylus on hepatopancreatic gene expression was examined at two levels, the specific gene level and the functional group level.
The overall effect of the different treatments was demonstrated by clustering the 12 individual transcript profiles, which resulted with prominent difference of the transcription profiles of the D-cHH treated females in respect to all other samples.
This overall effect is detailed as follows: The XOSG synthesizes and secretes multiple peptidic hormones, including D- and L-cHH. The effect of its removal on gene expression profiles revealed the differential expression of 214 transcripts, 147 up-regulated and 67 down-regulated. The carbohydrate metabolic activity was repressed as expected from the elimination of the glycemia inducing hormone.
The L-cHH-injected individuals revealed an expression profile similar to sham injected individuals. In contrary, D-cHH caused a considerable short term change in the hepatopancreatic gene expression profile. 917 transcripts were responsive to this isomer: 857 out of them were down regulated and only 60 were up-regulated. The functions mainly affected were the up-regulation of the proteolytic activity and the down-regulation of glycolysis. L-cHH caused much less pronounced effects, with only 45 differentially expressed genes (30 up-regulated and 15 down-regulated), without the detectable alteration of any specific function.
The hypothesized glycemia-related mechanisms of L- and D- cHH may involve protein activation with no changes in gene expression for both isomers and an additional D-cHH mechanism which involves changes in gene expression, which may be related or not to the increase of glucose levels in the circulation.
The D-cHH effect on gene expression was also demonstrated by the hypergeometric test, which distinguished an opposite trend related to the glycolysis which was revealed in the XOSG ablated group (S). The later rend was characterized by the up regulation of the enolase (IPR000941) that resulted to be significantly repressed after the D-cHH administration. Moreover, this short-time study showed that D-cHH was apparently able to restore the expression of a certain number of transcripts to the level of the naïve state, and it was also able to affect molecular pathways not altered by the XOSG ablation.
Supporting Information
The distribution of BLAST 13 Top Hit Species matching P. leptodactylus contigs.
(TIF)
Clustering the 12 mapping profiles of individual females using hierarchical and principal component analyses. A: Hierarchical clustering applying three clustering protocols, complete and single linkages and group average. (S) – sham injected, (N) – intact, (L) – L-cHH-injected, (D) – D-cHH-injected. B: principal component analysis. Green – native, Red – sham, Blue – L-cHH-injected and Yellow – D-cHH-injected.
(TIF)
Acknowledgments
The authors acknowledge the computer resources and technical support provided by the Plataforma Andaluza de Bioinformática of the University of Málaga, Spain.
Funding Statement
This work was supported by PRIN (Progetti di Ricerca di Interesse Nazionale) 2010-11 Prot. 20109XZEPR_002, Ministero dell'Istruzione, dell'Università e della Ricerca. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. No additional external funding received for this study.
References
- 1. Balik I, Çubuk H, Özkök R, Uysal R (2005) Some biological characteristics of crayfish (Astacus leptodactylus, Eschscholtz, 1823) in Lake Eǧirdir. Turkish Journal of Zoology 29: 295–300. [Google Scholar]
- 2. Hubenova TA, Vasileva PL, Zaikov AN (2009) Histological analysis of ovary development in narrow-clawed crayfish Astacus leptodactylus Esch. 1823 (crustaceae, decapoda, astacidae), reared in ponds in South Bulgaria. AACL Bioflux 2: 261–270. [Google Scholar]
- 3. Van Herp F, Bellon-Humbert C (1978) Setal development and molt prediction in the larvae and adults of the crayfish, Astacus leptodactylus (Nordmann, 1842). Aquaculture 14: 289–301. [Google Scholar]
- 4. Hopkins PM (2012) The eyes have it: A brief history of crustacean neuroendocrinology. General and Comparative Endocrinology 175: 357–366. [DOI] [PubMed] [Google Scholar]
- 5. Fanjul-Moles ML (2006) Biochemical and functional aspects of crustacean hyperglycemic hormone in decapod crustaceans: Review and update. Comparative Biochemistry and Physiology - C Toxicology and Pharmacology 142: 390–400. [DOI] [PubMed] [Google Scholar]
- 6.Giulianini P, Edomi P (2006) Neuropeptides controlling reproduction and growth in Crustacea: a molecular approach. In: Satake H, editor. Invertebrate Neuropeptides and Hormones: Basic Knowledge and Recent Advances Kerala, India: Research Signpost. pp. 225–252. [Google Scholar]
- 7. Webster SG, Keller R, Dircksen H (2012) The CHH-superfamily of multifunctional peptide hormones controlling crustacean metabolism, osmoregulation, moulting, and reproduction. General and Comparative Endocrinology 175: 217–233. [DOI] [PubMed] [Google Scholar]
- 8. Chung JS, Zmora N, Katayama H, Tsutsui N (2010) Crustacean hyperglycemic hormone (CHH) neuropeptidesfamily: Functions, titer, and binding to target tissues. General and Comparative Endocrinology 166: 447–454. [DOI] [PubMed] [Google Scholar]
- 9. Hsu YWA, Weller JR, Christie AE, de la Iglesia HO (2008) Molecular cloning of four cDNAs encoding prepro-crustacean hyperglycemic hormone (CHH) from the eyestalk of the red rock crab Cancer productus: Identification of two genetically encoded CHH isoforms and two putative post-translationally derived CHH variants. General and Comparative Endocrinology 155: 517–525. [DOI] [PubMed] [Google Scholar]
- 10. Lee KJ, Doran RM, Mykles DL (2007) Crustacean hyperglycemic hormone from the tropical land crab, Gecarcinus lateralis: Cloning, isoforms, and tissue expression. General and Comparative Endocrinology 154: 174–183. [DOI] [PubMed] [Google Scholar]
- 11. Ollivaux C, Gallois D, Amiche M, Boscaméric M, Soyez D (2009) Molecular and cellular specificity of post-translational aminoacyl isomerization in the crustacean hyperglycaemic hormone family. FEBS Journal 276: 4790–4802. [DOI] [PubMed] [Google Scholar]
- 12. Fu Q, Goy MF, Li L (2005) Identification of neuropeptides from the decapod crustacean sinus glands using nanoscale liquid chromatography tandem mass spectrometry. Biochemical and Biophysical Research Communications 337: 765–778. [DOI] [PubMed] [Google Scholar]
- 13. Stemmler EA, Hsu YWA, Cashman CR, Messinger DI, de la Iglesia HO, et al. (2007) Direct tissue MALDI-FTMS profiling of individual Cancer productus sinus glands reveals that one of three distinct combinations of crustacean hyperglycemic hormone precursor-related peptide (CPRP) isoforms are present in individual crabs. General and Comparative Endocrinology 154: 184–192. [DOI] [PubMed] [Google Scholar]
- 14. Palacios E, Carreño D, Rodríguez-Jaramillo MC, Racotta IS (1999) Effect of eyestalk ablation on maturation, larval performance, and biochemistry of white pacific shrimp, Penaeus vannamei, Broodstock. Journal of Applied Aquaculture 9: 1–23. [Google Scholar]
- 15. Rosas C, Fernandez I, Brito R, Diaz-Iglesia E (1993) The effect of eyestalk ablation on the energy balance of the pink shrimp, Penaeus notialis. Comparative Biochemistry and Physiology - A Physiology 104: 183–187. [Google Scholar]
- 16. Chang ES (1985) Hormonal control of molting in decapod crustacea. Integrative and Comparative Biology 25: 179–185. [Google Scholar]
- 17. Teshima SI, Kanazawa A, Koshio S, Horinouchi K (1989) Lipid metabolism of the prawn Penaeus japonicus during maturation: Variation in lipid profiles of the ovary and hepatopancreas. Comparative Biochemistry and Physiology – Part B: Biochemistry and 92: 45–49. [Google Scholar]
- 18. Santos EA, Nery LEM, Keller R, Gonçalves AA (1997) Evidence for the involvement of the crustacean hyperglycemic hormone in the regulation of lipid metabolism. Physiological Zoology 70: 415–420. [DOI] [PubMed] [Google Scholar]
- 19. Santos EA, Keller R (1993) Regulation of circulating levels of the crustacean hyperglycemic hormone: evidence for a dual feedback control system. Journal of Comparative Physiology B 163: 374–379. [Google Scholar]
- 20.Keller R, Sedlmeier D (1988) A metabolic hormone in crustaceans: the hyperglycemic hormone. In: Laufer H, Downer, FGH, editor. Endocrinology of selected invertebrates. New York: Alan R Liss Inc. pp. 315–326. [Google Scholar]
- 21. Kallen JL, Abrahamse SL, Van Herp F (1990) Circadian rhythmicity of the crustacean hyperglycemic hormone (CHH) in the hemolymph of the crayfish. Biol Bull 179: 351–357. [DOI] [PubMed] [Google Scholar]
- 22. Rivera-Pérez C, Navarrete del Toro M, García-Carreño FL (2010) Digestive lipase activity through development and after fasting and re-feeding in the whiteleg shrimp Penaeus vannamei. Aquaculture 300: 163–168. [Google Scholar]
- 23. Vogt G, Stfcker W, Storeh V, Zwilling R (1989) Biosynthesis of Astacus protease, a digestive enzyme from crayfish. Histochemistry 91: 373–381. [DOI] [PubMed] [Google Scholar]
- 24. Castro PF, Freitas Jr ACV, Santana WM, Costa HMS, Carvalho LB Jr, et al. (2012) Comparative study of amylases from the midgut gland of three species of penaeid shrimp. Journal of Crustacean Biology 32: 607–613. [Google Scholar]
- 25.Ceccaldi H (1989) Anatomy and physiology of digestive tract of Crustaceans Decapods reared in aquaculture. Advances in tropical aquaculture. Tahiti. pp. 243–259. [Google Scholar]
- 26. James M (1989) Cytochrome P450 monooxygenases in crustaceans. Xenobiotica 19: 1063–1076. [DOI] [PubMed] [Google Scholar]
- 27. Ahearn GA, Mandal PK, Manda A (2004) Mechanisms of heavy-metal sequestration and detoxification in crustaceans: a review. Journal of Comparative Physiology B 174: 439–452. [DOI] [PubMed] [Google Scholar]
- 28. Sedlmeier D (1988) The crustacean hyperglycemic hormone (CHH) releases amylase from the crayfish midgut gland. Regul Peptides 20: 91–98. [DOI] [PubMed] [Google Scholar]
- 29. Yasuda A, Yasuda Y, Fujita T, Naya Y (1994) Characterization of crustacean hyperglycemic hormone from the crayfish (Procambarus clarkii): multiplicity of molecular forms by sterioinversion and diverse function. Gen Comp Endocrinol 95: 387–398. [DOI] [PubMed] [Google Scholar]
- 30. Khayat M, Yang W, Aida K, Nagasawa H, Tietz A, et al. (1998) Hyperglycemic hormones inhibit protein and mRNA synthesis in in vitro-incubated ovarian fragments of the marine shrimp Penaeus semisulcatus. Gen Comp Endocrinol 110: 307–318. [DOI] [PubMed] [Google Scholar]
- 31. Avarre JCKM, Michelis R, Nagasawa H, Tietz A, Lubzens E (2001) Inhibition of de novo synthesis of a jelly layer precursor protein by crustacean hyperglycemic hormone family peptides and posttranscriptional regulation by sinus gland extracts in Penaeus semisulcatus ovaries. Gen Comp Endocrinol 124: 257–268. [DOI] [PubMed] [Google Scholar]
- 32. Tsutsui NKH, Ohira T, Nagasawa H, Wilder MN, Aida K (2005) The effects of crustacean hyperglycemic hormone-family peptides on vitellogenin gene expression in the kuruma prawn, Marsupenaeus japonicus. Gen Comp Endocrinol 144: 232–239. [DOI] [PubMed] [Google Scholar]
- 33. de Kleijn D, Janssen KPC, Waddy SL, Hegeman R, Lai WY, et al. (1998) Expression of the crustacean hyperglycaemic hormones and the gonad-inhibiting hormone during the reproductive cycle of the female American lobster Homarus americanus. Journal of Endocrinology 156: 291–298. [DOI] [PubMed] [Google Scholar]
- 34. Ollivaux C, Soyez D (2000) Dynamics of biosynthesis and release of crustacean hyperglycemic hormone isoforms in the X-organ-sinus gland complex of the crayfish Orconectes limosus. European Journal of Biochemistry 267: 5106–5114. [DOI] [PubMed] [Google Scholar]
- 35. Jia C, Hui L, Cao W, Lietz CB, Jiang X, et al. (2012) High-definition de novo sequencing of Crustacean Hyperglycemic Hormone (CHH)-family neuropeptides. Molecular and Cellular Proteomics 11: 1951–1964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Susanto G, Charmantier G (2001) Crayfish freshwater adaptation starts in eggs: ontogeny of osmoregulation in embryos of Astacus leptodactylus. J Exp Zool 289: 433–440. [DOI] [PubMed] [Google Scholar]
- 37. Harlioglu A, Aydin S, Yilmaz O (2012) Fatty acid, cholesterol and fat-soluble vitamin composition of wild and captive freshwater crayfish (Astacus leptodactylus). Food Sci Technol Int 18: 93–100. [DOI] [PubMed] [Google Scholar]
- 38. Giulianini P, Bierti M, Lorenzon S, Battistella S, Ferrero EA (2007) Ultrastructural and functional characterization of circulating hemocytes from the freshwater crayfish Astacus leptodactylus: cell types and their role after in vivo artificial non-self challenge. Micron 38: 49–57. [DOI] [PubMed] [Google Scholar]
- 39. Tunca E, Ucuncu E, Ozkan AD, Ulger ZE, Tekinay T (2013) Tissue Distribution and Correlation Profiles of Heavy-Metal Accumulation in the Freshwater Crayfish Astacus leptodactylus. Arch Environ Contam Toxicol Epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 40. Barim O, Karatepe M (2010) The effects of pollution on the vitamins A, E, C, beta-carotene contents and oxidative stress of the freshwater crayfish, Astacus leptodactylus. Ecotoxicol Environ Saf 73: 138–142. [DOI] [PubMed] [Google Scholar]
- 41. Malev O, Srut M, Maguire I, Stambuk A, Ferrero EA, et al. (2010) Genotoxic, physiological and immunological effects caused by temperature increase, air exposure or food deprivation in freshwater crayfish Astacus leptodactylus. Comp Biochem Physiol C Toxicol Pharmacol 152: 433–443. [DOI] [PubMed] [Google Scholar]
- 42. Mosco A, Zlatev V, Guarnaccia C, Pongor S, Campanella A, et al. (2012) Novel protocol for the chemical synthesis of crustacean hyperglycemic hormone analogues - an efficient experimental tool for studying their functions. PLoS ONE 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Mosco A, Edomi P, Guarnaccia C, Lorenzon S, Pongor S, et al. (2008) Functional aspects of cHH C-terminal amidation in crayfish species. Regul Pept 147: 88–95. [DOI] [PubMed] [Google Scholar]
- 44. Lebaupain F, Boscameric M, Pilet E, Soyez D, Kamech N (2012) Natural and synthetic chiral isoforms of crustacean hyperglycemic hormone from the crayfish Astacus leptodactylus: hyperglycemic activity and hemolymphatic clearance. Peptides 34: 65–73. [DOI] [PubMed] [Google Scholar]
- 45. Serrano L, Grousset E, Charmantier G, Spanings-Pierrot C (2004) Occurrence of L- and D-crustacean hyperglycemic hormone isoforms in the eyestalk X-organ/sinus gland complex during the ontogeny of the crayfish Astacus leptodactylus. J Histochem Cytochem 52: 1120–1140. [DOI] [PubMed] [Google Scholar]
- 46.Team RDC (2011) R: A language and environment for statistical computing. R Foundation for Statistical Computing. [Google Scholar]
- 47. Shechter A, Tom M, Yudkovski Y, Weil S, Chang SA, et al. (2007) Search for hepatopancreatic ecdysteroid-responsive genes during the crayfish molt cycle: From a single gene to mutagenicity. Journal of Experimental Biology 210: 3525–3537. [DOI] [PubMed] [Google Scholar]
- 48. Wang ZY, Gerstein M, Snyder M (2009) RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet 57–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Fernández-Pozo N, Guerrero-Fernández D, Bautista R, Gonzalo Claros M Full-LengtherNext: A tool for fine-tuning de novo assembled transcriptomes of non-model organisms. Paper in preparation, personal communication. [Google Scholar]
- 50. Baggerly KA, Deng L, Morris JS, Aldaz CM (2003) Differential expression in SAGE: Accounting for normal between-library variation. Bioinformatics 19: 1477–1483. [DOI] [PubMed] [Google Scholar]
- 51. Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: A practical and powerful approach to multiple testing. J R Stat Soc 57: 289–300. [Google Scholar]
- 52. Götz S, García-Gómez JM, Terol J, Williams TD, Nagaraj SH, et al. (2008) High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Research 36: 3420–3435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Conesa A, Götz S, García-Gómez JM, Terol J, Talón M, et al. (2005) Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21: 3674–3676. [DOI] [PubMed] [Google Scholar]
- 54. Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, et al. (1997) Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Research 25: 3389–3402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Ashburner M, Lewis S, Subramaniam, Noble, Kanehisa (2002) On ontologies for biologists: The gene ontology - Untangling the web. pp. 66–83. [PubMed] [Google Scholar]
- 56. Zdobnov EM, Apweiler R (2001) InterProScan - An integration platform for the signature-recognition methods in InterPro. Bioinformatics 17: 847–848. [DOI] [PubMed] [Google Scholar]
- 57. Falcon S, Gentleman R (2007) Using GOstats to test gene lists for GO term association. Bioinformatics 23: 257. [DOI] [PubMed] [Google Scholar]
- 58. Colbourne JK, Pfrender ME, Gilbert D, Thomas WK, Tucker A, et al. (2011) The ecoresponsive genome of Daphnia pulex. Science 331: 555–561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Kawahara-Miki R, Wada K, Azuma N, Chiba S (2011) Expression profiling without genome sequence information in a non-model species, pandalid shrimp (Pandalus latirostris), by next-generation sequencing. PLoS ONE 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Jung H, Lyons RE, Dinh H, Hurwood DA, McWilliam S, et al. (2011) Transcriptomics of a giant freshwater prawn (macrobrachium rosenbergii): De Novo assembly, annotation and marker discovery. PLoS ONE 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Uawisetwathana U, Leelatanawit R, Klanchui A, Prommoon J, Klinbunga S, et al. (2011) Insights into eyestalk ablation mechanism to induce ovarian maturation in the black tiger shrimp. PLoS ONE 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Chaves AR (2000) Effect of x-organ sinus gland extract on [35S] methionine incorporation to the ovary of the red swamp crawfish Procambarus clarkii. Comparative Biochemistry and Physiology - A Molecular and Integrative Physiology 126: 407–413. [DOI] [PubMed] [Google Scholar]
- 63. Sudha K, Anilkumar G (2007) Elevated ecdysteroid titer and precocious molt and vitellogenesis induced by eyestalk ablation in the estuarine crab, Metopograpsus messor (Brachyura: Decapoda). Journal of Crustacean Biology 27: 304–308. [Google Scholar]
- 64. Venkitraman PR, Jayalakshmy KV (2008) Effect of eyestalk ablation on the metabolic activities of two penaeid prawns: Metapenaeous monoceros and M. dobsoni (De Man). Italian Journal of Zoology 75: 345–360. [Google Scholar]
- 65. Nagai C, Nagata S, Nagasawa H (2011) Effects of crustacean hyperglycemic hormone (CHH) on the transcript expression of carbohydrate metabolism-related enzyme genes in the kuruma prawn, Marsupenaeus japonicus. General and Comparative Endocrinology 172: 293–304. [DOI] [PubMed] [Google Scholar]
- 66. Zmora N, Trant J, Zohar Y, Chung JS (2009) Molt-inhibiting hormone stimulates vitellogenesis at advanced ovarian developmental stages in the female blue crab, Callinectes sapidus 1: An ovarian stage dependent involvement. Saline Systems 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Okumura T, Kim YK, Kawazoe I, Yamano K, Tsutsui N, et al. (2006) Expression of vitellogenin and cortical rod proteins during induced ovarian development by eyestalk ablation in the kuruma prawn, Marsupenaeus japonicus. Comp Biochem Physiol A Mol Integr Physiol 143: 246–253. [DOI] [PubMed] [Google Scholar]
- 68. Vilella S, Zilli L, Ingrosso L, Schiavone R, Zonno V, et al. (2003) Differential expression of Na+/D-glucose cotransport in isolated cells of Marsupenaeus japonicus hepatopancreas. J Comp Physiol B 173: 679–686. [DOI] [PubMed] [Google Scholar]
- 69.Soyez D (2003) Recent data on the crustacean hyperglycemic hormone family; R FMN, editor. Enfield (NH), USA, Plymouth, UK: Science. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
The distribution of BLAST 13 Top Hit Species matching P. leptodactylus contigs.
(TIF)
Clustering the 12 mapping profiles of individual females using hierarchical and principal component analyses. A: Hierarchical clustering applying three clustering protocols, complete and single linkages and group average. (S) – sham injected, (N) – intact, (L) – L-cHH-injected, (D) – D-cHH-injected. B: principal component analysis. Green – native, Red – sham, Blue – L-cHH-injected and Yellow – D-cHH-injected.
(TIF)