

Abstract
Background and Purpose
Binge eating disorder (BED) is characterized by excessive food intake during short periods of time. Recent evidence suggests that alterations in the endocannabinoid signalling could be involved in the pathophysiology of BED. In this study, we investigated whether pharmacological manipulation of endocannabinoid transmission may be effective in modulating the aberrant eating behaviour present in a validated rat model of BED.
Experimental Approach
Binge-type eating was induced in female rats by providing limited access to an optional source of dietary fat (margarine). Rats were divided into three groups, all with ad libitum access to chow and water: control (C), with no access to margarine; low restriction (LR), with 2 h margarine access 7 days a week; high restriction (HR), with 2 h margarine access 3 days a week.
Key Results
Compared with the LR group, the HR group consumed more margarine and this was accompanied by an increase in body weight. The cannabinoid CB1/CB2 receptor agonist Δ9-tetrahydrocannabinol significantly increased margarine intake selectively in LR rats, while the fatty acid amide hydrolase inhibitor URB597 showed no effect. The CB1 receptor inverse agonist/antagonist rimonabant dose-dependently reduced margarine intake in HR rats. Notably, in HR rats, chronic treatment with a low dose of rimonabant induced a selective long-lasting reduction in margarine intake that did not develop tolerance, and a significant and persistent reduction in body weight.
Conclusions and Implications
Chronic pharmacological blockade of CB1 receptors reduces binge eating behaviour in female rats and may prove effective in treating BED, with an associated significant reduction in body weight.
Linked Articles
This article is part of a themed section on Cannabinoids. To view the other articles in this section visit http://dx.doi.org/10.1111/bph.2013.169.issue-4 & http://dx.doi.org/10.1111/bph.2012.167.issue-8
Keywords: binge eating, food intake, high-fat diet, weight loss, THC, URB597, rimonabant
Introduction
The endocannabinoid system, which comprises two cannabinoid receptor subtypes CB1 and CB2, their endogenous ligands (endocannabinoids, ECs) anandamide (AEA) and 2-arachidonoylglycerol, and corresponding biosynthesis and degradation pathways, is a homeostatic system involved in the regulation of several physiological functions, including cognition, reward, emotion, pain sensitivity and motor activity (Ameri, 1999; Solinas et al., 2008; Zanettini et al., 2011). There is a large body of evidence supporting the involvement of the endocannabinoid system in the modulation of energy balance by controlling food intake through central and peripheral mechanisms (Di Marzo and Matias, 2005). Systemic and local administrations of ECs and CB1 receptor agonists increase food intake in both animals and humans (Williams and Kirkham, 1999; Jamshidi and Taylor, 2001; Hart et al., 2002), whereas levels of ECs change during fasting and feeding in the rat hypothalamus and limbic brain areas (Kirkham et al., 2002). These hyperphagic actions are mediated by the CB1 receptor (Williams and Kirkham, 1999), as they can be selectively blocked by CB1 receptor antagonists, such as rimonabant and AM251 (Soria-Gòmez et al., 2007). In keeping with this, rimonabant and AM251 reduce food intake and feeding-associated behaviours in several rodent models (McLaughlin et al., 2003). Rimonabant also induces a significant decrease in food intake and body weight when chronically administered to either normal or obese rodents (Carai et al., 2006). Moreover, mice lacking the CB1 receptor are lean and hypophagic (Wiley et al., 2005), indicating that food intake and body weight gain depend upon the functional expression and activity of CB1 receptors. Conversely, an overactivity of the endocannabinoid system seems to be a key component in the pathophysiology of obesity (Di Marzo and Matias, 2005).
For these reasons, CB1 receptor blockade has long been considered a potential pharmacological tool to restore the normal endocannabinoid system tone under this pathological condition (Harrold and Williams, 2003). Rimonabant was the first selective CB1 receptor ligand to be used clinically and was approved worldwide as a treatment for obese and overweight individuals with metabolic complications (Scheen, 2008; Despres et al., 2009). However, due to the occurrence of psychiatric side effects after prolonged use (European Medicines Agency, 2009), in 2009 rimonabant was withdrawn from the European market, but still represents a reference drug in the search for new therapies for obesity (Cervino et al., 2009).
There is also converging evidence indicating that defects in the endocannabinoid system might be implicated not only in obesity but also in other eating disorders (Marco et al., 2011), such as binge eating disorder (BED). In the Diagnostic and Statistical Manual of Mental Disorders (fourth edition, text revision), BED is categorized as an ‘Eating Disorder Not Otherwise Specified’, and only listed in the appendix (American Psychiatric Association, 2000). Experts define ‘binge eating behaviour’ as an intermittent and uncontrollable consuming of an unusual amount of food (typically highly palatable foods rich in calories and with a high fat content) larger than that normally eaten by non-bingeing people in comparable periods of time. Binge eating episodes are not inevitably motivated by hunger or metabolic needs, and may or may not be followed by regular use of inappropriate compensatory behaviours, such as laxative use, vomiting, fasting, excessive exercise training, to purge the body of excess calories (American Psychiatric Association, 2000). Like other eating disorders, BED is more common in young people (Neumark-Sztainer et al., 2002) with a lifetime prevalence of 1.9 and 0.3% for women and men in six European countries (Preti et al., 2009) and of 3.5 and 2.0% for women and men in the USA (Hudson et al., 2007).
BED is of public and clinical interest due to its association with emotional distress and physic health problems (Johnson et al., 2001) and with the risk of excessive weight gain and obesity (Yanovski et al., 1993). Co-morbidity of obesity or abnormal eating behaviours marked by binge eating (Hudson et al., 2007; Grilo et al., 2009) and substance abuse disorders is well known (Wiederman and Pryor, 1996; Davis and Claridge, 1998). Compelling parallelisms exist between compulsive overeating and drug abuse, among which are the presence of craving and the loss of the inhibitory control on food intake and drug use. Interestingly, there is neuroanatomical and neurochemical overlapping between food and drug craving, with similar factors triggering relapse to overeating and drug use. Indeed, if drug-taking behaviour can be re-instated in abstinent rats and humans by a single exposure to the previously abused drug (Schmidt et al., 2005), a sustained intermittent exposure to sugar solution induces patterns of food intake and neurotransmitter changes comparable to those observed in animal models of drug abuse (Kelley et al., 2005; Avena et al., 2008). It is widely recognized that the endocannabinoid system is greatly involved in food intake (Li et al., 2011), drug addiction (Serrano and Parsons, 2011) and relapse (Fattore et al., 2007).
Overweight and obese subjects show increased frequency of the naturally occurring missense polymorphism of the gene that encodes the AEA hydrolyzing enzyme fatty acid amide hydrolase (FAAH), FAAH cDNA 385 A/A, which might potentiate the drive to eat by increasing endocannabinoid signalling (Sipe et al., 2005; Monteleone et al., 2008). On the other hand, a polymorphism of the CNR1 gene (encoding the human CB1 receptor) is thought to contribute to the vulnerability to anorexia nervosa (Siegfried et al., 2004). Moreover, women with anorexia nervosa or BED have elevated plasma levels of AEA (Monteleone et al., 2005), which could affect the rewarding aspect of eating that is compromised in these patients (Monteleone et al., 2008).
Although clinical studies indicate some improvement in the pharmacological management of bingeing-related eating disorders (Marazziti et al., 2011), treatment options are quite restricted and relapse incidences are still high. Limited progress in the development of strategies for the treatment of BED was mainly due to the fact that the neurobiological bases for repeatedly engaging in binge-type behaviour are not fully understood, and are not easy to study in humans. Examination of what is known about analogous behaviours in animals would help to elucidate the core components of BED.
In humans, accumulating data suggest that different situations may trigger compulsive eating leading to binge behaviour, psychological distress and dysphoria (Polivy, 1996). People typically binge on high palatable foods to which they have limited access (Kales, 1990; Fisher and Birch, 1999), after starvation and dietary restriction (American Psychiatric Association, 2000) or under stress (Oliver and Wardle, 1999).
Binge eating behaviour can be modelled in animal protocols to investigate neurobiological substrates and pharmacological determinants of human bingeing disorders (Berner et al., 2011). All animal models of BED available to date are isomorphic, and as such they mimic the human symptomatology by inducing similar behavioural states, that is, compulsive overeating (Hancock and Olmstead, 2010).
Based on previous studies showing that CB1 receptor blockade reduces feeding behaviour and the hedonic response to food, probably through the modulation of the mesolimbic dopaminergic pathway (Melis et al., 2007), this study was undertaken to investigate whether the CB1 receptor inverse agonist/antagonist rimonabant may be effective in modifying the aberrant eating behaviour present in a validated rat model of binge eating (Corwin et al., 1998). The effects of the natural CB1/CB2 receptor agonist Δ9-tetrahydrocannabinol (THC) were evaluated to compare the effects of CB1 receptor activation with that of CB1 receptor blockade. Furthermore, we tested the indirectly acting CB1 receptor synthetic agonist URB597 (a FAAH inhibitor) because it possesses anti-craving properties (Scherma et al., 2008b) and exerts anxiolytic-like and antidepressant-like effects in rodents (Piomelli et al., 2006; Gaetani et al., 2008; Scherma et al., 2008a) without evoking classical cannabinoid agonist-like effects (e.g. catalepsy, hypothermia, hyperphagia). Because craving as well as anxiety or depressive-like status may represent relevant behavioural traits related to BED, it was important to verify whether binge eating behaviour was affected by URB597 pretreatment.
Several studies have evaluated the effect of chronic blockade of CB1 receptors on eating behaviour and have reported a significant reduction in food consumption exclusively at the beginning of the treatment, thus demonstrating a transient reducing effect (Colombo et al., 1998; Carai et al., 2006; Martín-García et al., 2010). Therefore, it was important to verify in our experimental conditions whether rimonabant was able to maintain its acute effect on binge eating behaviour over time or whether such an effect underwent tolerance development.
Methods
Animals
Sprague–Dawley young adult female rats (Harlan Nossan, Udine, Italy) weighing 185–200 g at the start of the study (60–65 days old) were used. In our experiments, we used young female rats because in humans BED is more frequent in young females than in young males (Hudson et al., 2007). Following arrival, animals were individually housed in a climate-controlled animal room (21 ± 2°C temperature; 60% humidity) under a reversed 12 h light/dark cycle (lights on 24:00 h) with standard rat chow and water ad libitum. All experiments were approved by the local Animal Care Committee and carried out in strict accordance with the E.C. Regulations for Animal Use in Research (CEE No. 86/609). All studies involving animals are reported in accordance with the ARRIVE guidelines for reporting experiments involving animals (Kilkenny et al., 2010; McGrath et al., 2010).
Diets
Standard rat chow (Safe, France): 3% kcal from fat, 61% kcal from carbohydrate, 16% kcal from protein, 20% moisture, containing 2.9 kcal·g−1. High-fat diet (Margarine, Gradina Unilever Italia Mkt.): 70% kcal from fat, <1% kcal from carbohydrate, containing 6.5 kcal·g−1).
Experimental procedure
Our protocol is based on previous work showing that limiting access to an optional source of dietary fat induces a binge-type eating behaviour in rats that can be maintained for long periods of time, even though animals are never deprived of food (Corwin et al., 1998; Corwin and Buda-Levin, 2004; Hancock and Olmstead, 2010). Animals were given access to high-fat food during their low activity period, that is, close to the time of switching lights off, as this does not interfere with their normal circadian rhythm, as previously described by Corwin and Wojnicki (2006).
As illustrated in Figure 1, following 1 week of acclimatization rats were divided into three groups matched for body weight, which correspond to the following diet conditions (that were maintained for the entire period of the experimental study):
Figure 1.
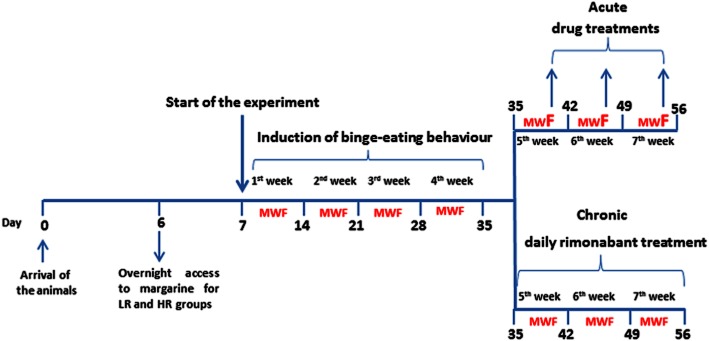
Schematic representation of the experimental design.
Low restriction (LR): had continuous access to standard chow and water. In addition, animals were given 2 h access to a separate bowl of margarine introduced into the home cage every day of the week, 3 h before the start of the dark cycle (Corwin and Wojnicki, 2006).
High restriction (HR): had continuous access to standard chow and water. In addition, animals were given 2 h access to a separate bowl of margarine introduced into the home cage every Monday, Wednesday and Friday, 3 h before the start of the dark cycle.
Control (C): had continuous access to standard chow and water. Margarine was not provided at any time of the study.
Margarine and/or standard chow were measured in all diet groups on Mondays, Wednesdays and Fridays (MWF) throughout the study by weighing them before and after the 2 h margarine access period (Figure 1). Before the start of the experiments, both LR and HR groups were given overnight access to a bowl of margarine to prevent neophobia.
Drug treatment commenced only once binge eating behaviour was firmly established (induction phase: 3–4 weeks). In our experiments, we included only animals that, at the end of the induction phase, displayed an intake of margarine within ±25% variation of the mean intake. Two separate batches of animals were used for acute and chronic treatments.
Acute treatment
Animals from each diet group (C, LR and HR, n = 24 per diet group) were randomly allocated into three different groups according to the pharmacological treatment assigned on the test day (Friday). Drug treatments (n = 8 per drug treatment group) were administered in a random sequence at weekly intervals. In keeping with previous studies (Koch JE, 2001; Fegley et al., 2005; Orio et al., 2009), a 1 week interval between each drug treatment proved to be a sufficient washout period. Treatments were the following: (i) vehicle, THC 0.125 and 0.250 mg·kg−1; (ii) vehicle, URB597 0.3 and 3 mg·kg−1; (iii) vehicle, rimonabant 0.3 and 3 mg·kg−1. Drug injections were given i.p. 30 min before the margarine access period, with the only exception of URB597, in which pretreatment time was 40 min based on previous studies (Solinas et al., 2006; Scherma et al., 2008a). In all diet groups, margarine and/or chow were weighed before and after the 2 h access period on the test day.
Chronic treatment
Animals from each diet group (n = 20) were randomly assigned to two different groups, which received either rimonabant 0.3 mg·kg−1 (n = 10) or vehicle i.p. (n = 10). Drugs were administered once a day for 21 consecutive days, 30 min before the margarine access period. In both groups, margarine and/or chow were weighed on MWF before and after the 2 h access period. Body weight was recorded once a week on Fridays.
Materials
THC (RTI International, Research Triangle Park, NC, USA), 50 mg·mL−1 in ethanol, and rimonabant (National Institute on Drug Abuse, NIH, Baltimore, MD, USA) were dissolved in 2% Tween 80, 2% ethanol, and saline. URB597 (Cayman Chemical Company, Ann Arbor, MI, USA) was dissolved in 20% DMSO and saline. All drugs were injected i.p. in a volume of 1 mL·kg−1.
Data analysis
Data from the induction of binge-type eating are expressed as mean kcal of margarine, chow and margarine + chow (total intake) (1-block week: MWF) ± SEM during the 2 h access period. Data were analysed by two-way anova for repeated measures with diet group and week as factors, and week as a repeated factor.
Data from each acute treatment (margarine, chow and total intake) are expressed as mean kcal ± SEM during the 2 h access period on the test day, and were analysed by two-way anova with diet group and treatment as factors. The effects of treatment within each diet group were analysed by one-way anova as treatment between-subjects factor.
Data from chronic treatment (margarine, chow and total intake) are expressed as mean kcal (1-block week: MWF) ± SEM during the 2 h access period, and were analysed by three-way anova with diet group, treatment and week as main factors, and week as a repeated factor. Significant differences within the diet group were further analysed by two-way anova with treatment and week as main factors and week as a repeated factor.
Data from body weight during the induction phase of binge eating are expressed as mean in g ± SEM and were analysed by two-way anova with diet groups and week as main factors and week as a repeated factor.
Data from body weight during chronic treatment were analysed by three-way anova with groups, treatment and week as main factors and week as a repeated factor. Significant differences within diet groups were further analysed by two-way anova with treatment and week as main factors and week as a repeated factor.
Post hoc comparisons, when appropriate, were performed by Newman–Keuls multiple comparison test or by Bonferroni test. In all cases, differences with a P < 0.05 were considered significant. Student's t-test was used when indicated and used to compare two conditions.
Results
Experiment 1. Acute drug treatments
Induction of binge-type eating
Consistent with previous findings (Corwin et al., 1998; Dimitriou et al., 2000), HR rats invariably consumed more margarine than LR rats during the 2 h limited access (Figure 2A). Two-way anova revealed a significant effect of diet group [F(1,138) = 12.77, P = 0.0008] and week [F(3,138) = 6.22, P = 0.0005], and a diet group × week significant interaction [F(3,138) = 3.78, P = 0.0121]. Post hoc analysis showed that margarine consumption was significantly greater in the HR than in the LR group by the third week of the study (3rd week: P < 0.001; 4th week: P < 0.01).
Figure 2.
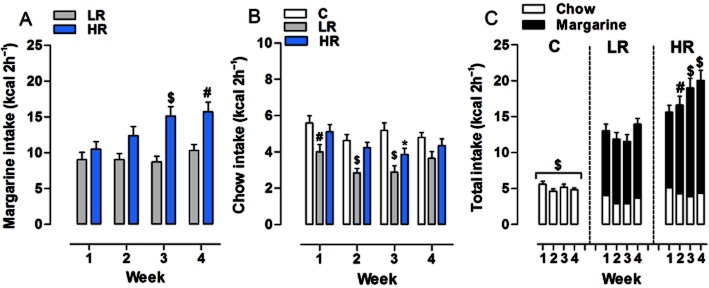
Induction of binge-type eating. All data are presented as mean kcal (1-block week: MWF) ± SEM during the limited (2 h) access. (A) Margarine intake: HR group with limited access to margarine 3 days a week consumed more margarine than LR group with daily access to margarine (3rd week: $P < 0.001; 4th week: #P < 0.01, Bonferroni post test). (B) Chow intake: LR group consumed less chow than C group (1st week: #P < 0.01; 2nd and 3rd weeks: $P < 0.001, Student's t-test) and HR group (2nd week: $P < 0.001, Student's t-test). HR group consumed less chow than C group only during the third week (*P < 0.05, Student's t-test). (C) Total intake: during the limited access HR group displayed higher total intake than LR group (2nd week: #P < 0.01; 3rd and 4th weeks: $P < 0.001, Bonferroni post test). In the C group, the total intake during the limited access was significantly less than that of both the LR and HR groups ($P < 0.001, Bonferroni post test).
Notably, chow consumption during the 2 h limited access period was affected by the schedule of margarine availability (Figure 2B), since two-way anova revealed a significant effect of diet group [F(2,207) = 11.80, P < 0.0001] and week [F(3,207) = 7.46, P < 0.0001]. LR group consumed significantly less chow than C group during the first 3 weeks of the study [Student's t-test: 1st week: t(46) = 2.814, P = 0.0072; 2nd week: t(46) = 4.387, P < 0.0001; 3rd week: t(46) = 4.234, P = 0.0001], but not on the last (4th) week. Moreover, LR rats consumed significantly less chow than HR rats on the second week only [Student's t-test: t(46) = 3.614, P = 0.0007], while a significant difference between C and HR groups was found on the third week [Student's t-test: t(46) = 2.459, P = 0.0178].
When looking at the cumulative amount of margarine and/or chow consumed by each diet group during the 2 h limited access (Figure 2C), two-way anova detected a main effect of diet group [F(2,207) = 82.92, P < 0.0001] and week [F(3,207) = 4.33, P = 0.0055] and a diet group × week significant interaction [F(6,207) = 3.81, P = 0.0013]. Post hoc analysis showed that HR group displayed higher total intake during the limited access than LR group by the second week of the study (2nd week: P < 0.01; 3rd and 4th weeks: P < 0.001). In both LR and HR groups, the total intake during the limited access was significantly higher than that of C group (P < 0.001).
Effect of THC on binge-type eating
On the test day, margarine consumption during the 2 h limited access period was affected by treatment with THC (0.125 and 0.250 mg·kg−1) (Figure 3A). Two-way anova revealed a significant effect of diet group [F(1,42) = 5.66, P = 0.0220] and treatment [F(2,42) = 4.60, P = 0.0156]. Subsequent individual one-way anova showed that THC significantly increased margarine intake in LR group [F(2,21) = 7.112, P = 0.0044]. Compared with vehicle-treated rats, post hoc analysis revealed that both doses of THC were effective in increasing margarine intake in LR rats (+101% and +121%, respectively, P < 0.01). THC did not affect margarine intake in the HR group [F(2,21) = 0.367, P = 0.697].
Figure 3.
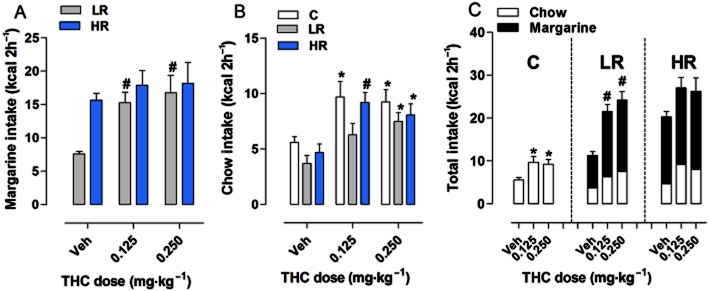
Effect of THC on binge-type eating. All data are presented as mean kcal ± SEM during the limited (2 h) access on the test day. (A) Margarine intake: THC doses of 0.125 and 0.250 mg·kg–1 increased margarine intake in the LR group (#P < 0.01 vs. Veh, Newman–Keuls multiple comparison test). No effect was found in the HR group. (B) Chow intake: THC was effective in increasing chow intake in all groups (C group: *P < 0.05 vs. Veh; LR group: *P < 0.05 vs. Veh; HR group: #P < 0.01 and *P < 0.05 vs. Veh, Newman–Keuls multiple comparison test). (C) Total intake: THC was effective in increasing the total intake in both C and LR groups (*P < 0.05 vs. Veh, #P < 0.01 vs. Veh, Newman–Keuls multiple comparison test).
On the other hand, THC also affected chow consumption during the 2 h limited access period, as two-way anova detected a significant effect of diet group [F(2,63) = 4.68, P = 0.0128] and treatment [F(2,63) = 14.41, P < 0.0001] (Figure 3B). Individual one-way anova revealed that THC significantly increased chow intake in all diet groups as compared with vehicle-treated rats [C group: F(2,21) = 4.258, P = 0.028; LR group: F(2,21) = 4.772, P = 0.0196; HR group: F(2,21) = 6.715, P = 0.0056, one-way anova]. Post hoc analysis revealed that both doses of THC were effective in increasing chow intake by 73 and 65%, respectively, in C rats (P < 0.05), and by 96 and 72%, respectively, in the HR group (P < 0.01 and P < 0.05). Conversely, only the highest dose of THC (0.250 mg·kg−1) was effective in increasing chow intake in LR rats by 102% (P < 0.05).
Finally, as shown in Figure 3C, THC affected the total food intake during the 2 h limited access period (margarine + chow for LR and HR groups, and only chow for the C group) in both C [F(2,21) = 4.258, P = 0.028, one-way anova] and LR [F(2,21) = 18.89, P < 0.0001, one-way anova] groups. Post hoc analysis showed that both doses of THC were effective in increasing total intake as compared with vehicle-treated rats (P < 0.05 and P < 0.01 respectively).
Effect of URB597 on binge-type eating
On the test day, treatment with URB597 (0.3 and 3 mg·kg−1) did not produce significant changes on margarine intake during the 2 h limited access (Figure 4A). Two-way anova revealed a significant effect of diet group [F(1,42) = 11.83, P = 0.00013], but not treatment [F(2,42) = 0.15, P = 0.8628]. Although chow intake was affected by URB597 treatment [F(2,63) = 4.46, P = 0.0155, two-way anova], one-way anova within each group did not show significant differences with respect to vehicle-treated rats during the 2 h limited access period [C group: F(2,21) = 2.218, P = 0.1337; LR group: F(2,21) = 1.546, P = 0.2365; HR group: F(2,21) = 1.629, P = 0.219, one-way anova] (Figure 4B).
Figure 4.
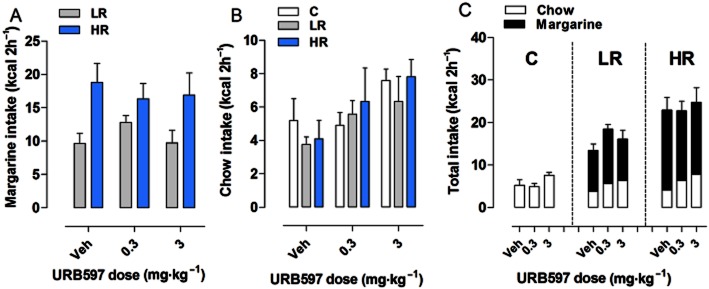
Effect of URB597 on binge-type eating. All data are presented as mean kcal ± SEM during the limited (2 h) access on the test day. (A) Margarine intake: URB597 (0.3 and 3 mg·kg−1) did not alter margarine intake in both LR and HR groups as compared with vehicle-treated rats. (B) Chow intake: no effect was found on chow intake in all experimental groups. (C) Total intake: URB597 treatment did not modify the total intake.
URB597 treatment did not modify the total food intake during the 2 h limited access period (margarine + chow for the LR and HR groups, only chow for the C group) either (Figure 4C), as revealed by individual one-way anova within each diet group [C: F(2,21) = 2.218, P = 0.133; LR: F(2,21) = 2.173, P = 0.138; HR: F(2,21) = 0.145, P = 0.865].
Effect of rimonabant on binge-type eating
Treatment with rimonabant (0.3 and 3 mg·kg−1) significantly affected margarine intake in both LR and HR groups on the test day (Figure 5A). Two-way anova revealed a significant effect of diet group [F(1,42) = 5.72, P = 0.0213] and treatment [F(2,42) = 14.18, P < 0.0001]. Individual one-way anova showed that both doses of rimonabant tested were effective in reducing margarine intake by 47 and 76%, respectively, as compared with vehicle-treated rats in HR group [F(2,21) = 10.82, P = 0.0006, (P < 0.01 and P < 0.001, post hoc test)]. Notably, the highest dose of rimonabant (3 mg·kg−1) also reduced (−41%) the intake of margarine in LR group as compared with vehicle-treated rats [F(2,21) = 3.709, P = 0.0418, (P < 0.05, post hoc test), one-way anova]. Treatment with rimonabant also affected chow consumption during the 2 h limited access period [two-way anova, significant effect of diet group (F(2,63) = 4.38, P = 0.0166) and treatment (F(2,63) = 28.89, P < 0.0001)]. As shown in Figure 5B, rimonabant decreased chow intake in all groups [C group: F(2,21) = 11.48, P = 0.0004; LR group: F(2,21) = 8.584, P = 0.0019; HR group: F(2,21) = 9.679, P = 0.001, one-way anova] at both doses tested as compared with vehicle-treated rats (C group: −73 and −83% with P < 0.001; LR group: −65 and −95% with P < 0.05 and P < 0.01 respectively; HR group: −64 and −83% with P < 0.01, post hoc test).
Figure 5.
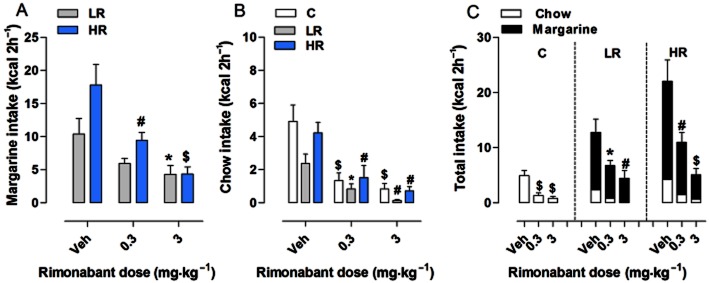
Effect of rimonabant on binge-type eating. All data are presented as mean kcal ± SEM during the limited (2 h) access on the test day. (A) Margarine intake: rimonabant doses of 0.3 and 3 mg·kg−1 decreased margarine intake in the HR group (#P < 0.01 and $P < 0.001 vs. Veh, Newman–Keuls multiple comparison test). The higher dose of 3 mg·kg−1 was also found to be effective in the LR group (*P < 0.05 vs. Veh, Newman–Keuls multiple comparison test). (B) Chow intake: both doses were effective in decreasing chow intake in all groups (C group: $P < 0.001 vs. Veh; LR group: *P < 0.05 and #P < 0.01 vs. Veh; HR group: #P < 0.01 vs. Veh, Newman–Keuls multiple comparison test). (C) Total intake: rimonabant was effective in decreasing the total intake in all groups (C group: $P < 0.001 vs. Veh; LR group: *P < 0.05 and #P < 0.01 vs. Veh; HR group: #P < 0.01 and $P < 0.001 vs. Veh, Newman–Keuls multiple comparison test).
Finally, rimonabant significantly decreased the total intake of food during the 2 h limited access period (margarine + chow for the LR and HR groups, and only chow for the C group) in all three diet groups as compared with vehicle-treated rats [C group: F(2,21) = 11.48, P = 0.0004; LR group: F(2,21) = 6.118, P = 0.0081; HR group: F(2,21) = 15.11, P < 0.0001, one-way anova] (Figure 5C). Post hoc analysis revealed a significant drug effect at both the 0.3 and 3 mg·kg−1 doses (C group: P < 0.001; LR group: P < 0.05 and P < 0.01 respectively; HR group: P < 0.01 and P < 0.001 respectively).
Experiment 2. Chronic rimonabant treatment
Effect of chronic rimonabant on binge-type eating
The effects of a chronic treatment (21 consecutive days) with rimonabant or its vehicle on margarine and/or chow intake during the 2 h limited access period were studied in a different set of animals showing binge-type eating behaviour that was induced in a similar manner to animals in Figure 2 (data not shown). Figure 6A shows the effect of chronic rimonabant and vehicle on margarine intake. Three-way anova showed a significant effect of diet group [F(1,36) = 71.630, P < 0.0001] and treatment [F(1,36) = 71.630, P < 0.0001], but not week [F(2,36) = 0.7592, P = 0.4713], nor a significant interaction among these three factors. Two-way anova performed within each diet group also revealed a significant effect of treatment [LR: F(1,36) = 18.50, P = 0.0004; HR: F(1,36) = 21.75, P = 0.0002]. Both LR and HR groups treated with rimonabant consumed significantly less margarine compared with vehicle-treated rats during the 3 weeks of treatment [Student's t-test: LR 1st week: t(18) = 4.518, P = 0.0003; 2nd week: t(18) = 4.816, P = 0.0001; 3rd week: t(18) = 2.622, P = 0.0173; HR 1st week: t(18) = 3.775, P = 0.0014; 2nd week: t(18) = 4.009, P = 0.0007; 3rd week: t(18) = 3.256, P = 0.0044].
Figure 6.
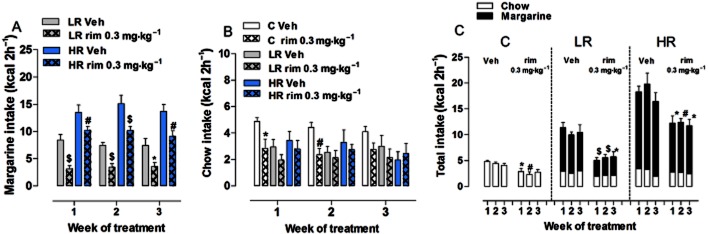
Effect of chronic rimonabant on binge-type eating. All data are presented as group mean kcal (1-block week: MWF) ± SEM during the limited (2 h) access. (A) Margarine intake: rimonabant (0.3 mg·kg−1) decreased margarine intake in both LR (1st and 2nd weeks: $P < 0.001; 3rd: *P < 0.05, Student's t-test) and HR (1st and 3rd weeks: #P < 0.01; 2nd week: $P < 0.001, Student's t-test) groups as compared with vehicle-treated rats. (B) Chow intake: chronic treatment with rimonabant significantly decreased chow intake only in C group during the first 2 weeks as compared with vehicle-treated rats (1st week: *P < 0.05; 2nd week: #P < 0.01, Student's t-test). (C) Total intake: chronic rimonabant was effective in decreasing the total intake in all groups as compared with vehicle-treated rats (C group: 1st week: *P < 0.05; 2nd week: #P < 0.01, Student's t-test; LR group: 1st and 2nd weeks: $P < 0.001; 3rd: *P < 0.05, Student's t-test; HR group: 1st and 3rd weeks: *P < 0.05; 2nd week: #P < 0.01, Student's t-test).
On the other hand, three-way anova showed that chow consumption was affected by treatment only [F(1,54) = 5.1504, P = 0.027], as no significant effect of diet group [F(2,54) = 2.5399, P = 0.088] or week [F(2,54) = 1.613, P = 0.203] was found, nor a significant interaction among these three factors (Figure 6B). Two-way anova performed within each diet group revealed a significant effect of treatment in the C group only [F(1,36) = 11.66, P = 0.0031]: rimonabant-treated animals consumed significantly less chow compared with vehicle-treated rats during the first 2 weeks of the treatment [Student's t-test: 1st week: t(18) = 2.716, P = 0.0142; 2nd week: t(18) = 3.369, P = 0.0034], but not on the last (3rd) week.
Finally, as shown in Figure 6C, rimonabant also affected the total intake (margarine + chow for the LR and HR groups, and only chow for the C group) in all three diet groups; two-way anova performed within each diet group detected a main effect of treatment [C: F(1,36) = 11.66, P = 0.0031; LR: F(1,36) = 20.97, P = 0.0002; HR: F(1,36) = 9.78, P = 0.0058]. The total food intake in both LR and HR groups treated with rimonabant was less than in vehicle-treated rats during all the 3 weeks of treatment [Student's t-test: LR 1st week: t(18) = 5.224, P < 0.0001; 2nd week: t(18) = 4.897, P = 0.0001; 3rd week: t(18) = 2.599, P = 0.0181; HR 1st week: t(18) = 2.826, P = 0.0112; 2nd week: t(18) = 3.247, P = 0.0045; 3rd week: t(18) = 2.748, P = 0.0132]. As mentioned above, rimonabant affected the total intake of the C group only during the first 2 weeks of treatment.
Effect of chronic rimonabant on body weight
As shown in Figure 7A, significant changes in the mean body weight were detected during the induction phase. Two-way anova revealed a significant effect of diet group [F(2,171) = 4.13, P = 0.0212] and week [F(3,171) = 529.55, P < 0.0001], and a significant diet group × week interaction [F(6,171) = 4.13, P = 0.0007]. Post hoc analysis indicated that HR rats weighed more than C rats by the third week of the study (3rd week: P < 0.05; 4th week P < 0.01). No significant differences were found between HR and LR rats, nor between C and LR rats.
Figure 7.
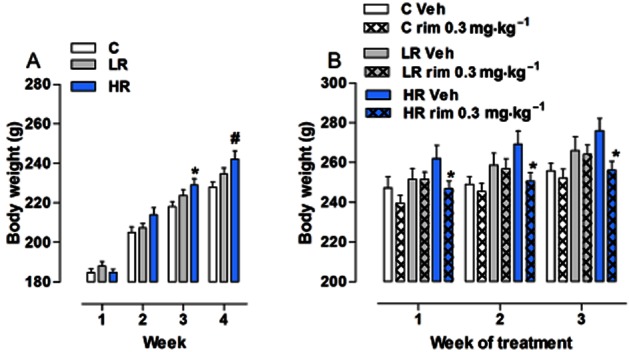
All data are presented as group means ± SEM weekly change in body weight. (A) Induction of binge-type eating: HR group weighed more than the C group (3rd week: *P < 0.05; 4th week: #P < 0.01, Bonferroni post test). Not significant differences were found between HR and LR groups, nor between C and LR groups. (B) Effect of chronic rimonabant on body weight: HR rats chronically treated with rimonabant had a significantly decreased body weight as compared with corresponding vehicle-treated rats (*P < 0.05, Student's t-test). Chronic treatment with rimonabant did not affect the body weight in either theC or LR groups when compared with corresponding vehicle-treated rats.
The effects of chronic treatment with rimonabant or its vehicle are shown in Figure 7B. Three-way anova showed a significant effect of diet group [F(2,54) = 4.72, P = 0.01294] and treatment [F(1,54) = 5.15, P = 0.0276] and week [F(2,54) = 51.38, P < 0.0001], but not a significant interaction among these three factors. Two-way anova performed within each group detected a main effect of treatment in the HR group only [F(1,36) = 5.58, P = 0.0296]. HR rats chronically treated with rimonabant significantly decreased body weight as compared with corresponding vehicle-treated rats [Student's t-test: 1st week: t(18) = 2.612, P = 0.0176; 2nd week: t(18) = 2.372, P = 0.0290; 3rd week: t(18) = 2.566, P = 0.0194].
Discussion
The aim of our study was to verify if the pharmacological manipulation of the endocannabinoid system could be effective in the modulation of abnormal eating behaviour developed by female rats in a confirmed rat model of BED, in which binge eating behaviour is induced in animals by giving them a sporadic (3 days week-1) and limited (2 h) access to a high-fat diet (margarine) in addition to a continuous access to chow and water (HR group). In these animals, the intake of margarine becomes significantly greater than those of animals with limited daily access to margarine (LR group), and remains stable over prolonged periods of time (Corwin and Buda-Levin, 2004; Corwin and Wojnicki, 2006).
As in other animal models of BED, in our limited access model binge eating was characterized by behavioural patterns similar to those seen in humans, as rats consumed a large quantity of food in a brief, defined period of time, and these quantities exceeded the amount typically consumed by control animals. Notably, our rats were never deprived of food, which is similar to bingeing humans who eat in the absence of hunger (Marcus and Kalarchian, 2003). Unfortunately, human subjective feelings of distress or loss of control found in some (Engel et al., 2007) but not in all bingeing subjects (Wegner et al., 2002) cannot be assessed easily in animals (Corwin and Buda-Levin, 2004). However, binge eating animal models exploit the precursory circumstances leading to binge eating in humans (e.g. dieting, exposure to palatable foods and fluids, stress) (Corwin and Buda-Levin, 2004).
In this study, treatment with the natural CB1/CB2 receptor agonist THC proved to be effective in increasing margarine intake exclusively in the LR group, in line with the orexigenic effects of THC in humans and rodents (Williams and Kirkham, 1999; Hart et al., 2002), and with the finding that our control (C) animals, in which margarine was not provided at any time, ate more standard chow after THC injection.
In LR rats, THC increased the total food intake with a specific effect on palatable food (Koch, 2001), where it was effective even at the lowest dose tested. However, THC did not affect margarine consumption in HR animals, although it significantly stimulated chow intake. We assume that the consumption of margarine was already the highest achievable in our bingeing animals (HR group), so that the CB1 receptor agonist was not able to increase it further, suggesting the possibility of an enhanced endocannabinoid tone in HR rats compared with the LR and C groups.
Treatment with URB597, which prevents intracellular inactivation of AEA by FAAH inhibition and prolongs its behavioural and neurochemical effects (Kathuria et al., 2003), did not induce a significant increase in the amount of margarine or for chow consumed, although we observed a positive trend. In contrast to our findings, Soria-Gòmez et al. (2007) reported that local infusion of N-arachidonoyl-serotonin (AA5-HT), another FAAH inhibitor (Bisogno et al., 1998), into the nucleus accumbens shell markedly stimulated the ingestion of standard chow. This effect was prevented by AM251, suggesting the involvement of the CB1 receptor in the orexigenic responses to AA5-HT administration. The FAAH inhibitor AA5-HT, infused in the parabrachial nucleus, also increased the consumption of a palatable high-fat/sucrose diet, an effect mediated by CB1 receptors (Dipatrizio and Simansky, 2008). After injection of doses similar to those used in this study (0.3 and 3 mg·kg−1), FAAH inhibition was reported to be rapid (<15 min), persistent (>16 h) and correlated with a threefold increase in brain AEA levels (Kathuria et al., 2003; Fegley et al., 2005). The lack of response in our animals could be ascribed to the different route of administration used (i.p. rather than direct brain infusion), or to the fact that URB597 is unnable to further enhance the pre-existing endocannabinoid tone of our bingeing rats. Alternatively, the finding that the FAAH inhibitor had no effect in our bingeing animals can imply the involvement of other neurotransmitter systems. It should be noted that FAAH inhibition increases brain levels and magnifies and prolongs the effects of the non-cannabinoid fatty acid ethanolamides oleoylethanolamide (OEA) and palmitoylethanolamide, which are endogenous ligands for the PPARα (Fegley et al., 2005; O'Sullivan, 2007). In contrast to AEA, OEA decreases food intake and body weight gain in lean and obese rats and mice through a CB1 receptor-independent mechanism (Fu et al., 2003; Lo Verme et al., 2005).
In contrast, the inverse agonist/antagonist rimonabant, when given acutely significantly and dose-dependently decreased margarine intake in HR rats; yet, it only reduced margarine intake in LR animals at high doses (3 mg·kg−1). In line with this, Parylak et al. (2012) showed that administration of a different CB1 receptor antagonist, SR147778, dose-dependently attenuated binge-like intake of a sweet-fat diet in rats. In addition, both doses of rimonabant reduced consumption of standard chow in all three diet groups.
The most intriguing outcome of our study comes from the chronic treatment experiment with the low dose (0.3 mg·kg−1) of rimonabant, which showed that rimonabant preserves its selective reducing effect on fat food over time, as demonstrated by the finding that both HR and LR rats treated with the CB1 receptor inverse agonist/antagonist consumed less margarine but their intake of standard chow was unaltered. At present, as reviewed by Berner et al. (2011), the anorectic effect of CB1 receptor antagonists on palatable food consumed in a binge-like manner is not completely elucidated. Our results show for the first time that chronic treatment with rimonabant in rats that consumed the high-fat diet in a binge-type pattern resulted in a selective decrease in the consumption of palatable food, an effect that was maintained throughout the entire treatment period without the occurrence of tolerance, that instead develops towards standard chow intake (Colombo et al., 1998; Carai et al., 2006; Martín-García et al., 2010) or in other animal models of food intake (Mathes et al., 2008). Indeed, chronic rimonabant reduced food intake in the control rats with only access to standard chow, showing the development of tolerance from the second week of treatment. In line with this, preceding studies showed that CB1 receptor antagonists decrease energy intake by selectively reducing the consumption of palatable diets in normal rats (Arnone et al., 1997; Simiand et al., 1998; Mathes et al., 2008), and that the antagonistic effect might not be limited to palatable food (McLaughlin et al., 2003; Foltin and Haney, 2007). In our study, chronic treatment with rimonabant also reduced body weight in the HR group (which was increased at the end of the induction phase), but not in the C and LR groups. This finding is consistent with previous reports showing that CB1 receptor antagonism preferentially reduces the body weight of obese rats or rats with access to a sugar fat whip dessert (Mathes et al., 2008; Martín-García et al., 2010).
Although the mechanism through which rimonabant exerts its effects on binge-type eating behaviour is still to be clarified, previous findings suggest that it may be effective in suppressing the hedonic response to food rather than just hunger (Cota et al., 2006). Several lines of evidence point to the endocannabinoid system as an important constituent of neuronal substrates involved in brain reinforcement/reward processes implicated in both food consumption and the homeostatic and hedonic regulation of eating (Di Marzo and Matias, 2005). In keeping with this, CB1 receptors are expressed particularly in brain areas (nucleus accumbens, hippocampus and entopeduncular nucleus) that are either directly involved in the hedonic aspects of eating or are connected to reward-related brain areas (Herkenham et al., 1990). Administration of ECs into the nucleus accumbens exerts a potent CB1 receptor-selective hyperphagic action (Kirkham et al., 2002; Soria-Gòmez et al., 2007) and enhances sucrose hedonic impact with hotspot focus in the dorsal shell part of the nucleus (Mahler et al., 2007). It is well recognized that palatable food stimulates the mesocorticolimbic dopamine system in a way similar to that of drugs of abuse, by increasing dopamine release in the shell of the nucleus accumbens (Martel and Fantino, 1996). Importantly, the increase in dopamine induced by presentation of palatable foods is blocked by administration of rimonabant (Melis et al., 2007), which suggests that the hedonic response to food might depend on the endocannabinoid system, probably through modulation of the mesocorticolimbic system. It should be noted that the release of dopamine disappears with repeated access to palatable food (Bassareo and Di Chiara, 1999), an effect not found when the subject is exposed to drugs of abuse (Wise et al., 1995). On the other hand, it has been demonstrated that animals that binge to a 10% sucrose solution repeatedly release dopamine in the nucleus accumbens shell each time they binge (Rada et al., 2005). The same effect has been shown when animals binge high-fat diets (Liang et al., 2006). In rats, prolonged binge-like intake of sugar solution leads to increased D1 receptor binding in the accumbens core and shell and decreased D2 receptor binding in the dorsal striatum (Colantuoni et al., 2001). Also, restricted feeding with scheduled sucrose access is associated with increased dopamine membrane transporter protein density in the nucleus accumbens and in the ventral tegmental area (Bello et al., 2003) in addition to an increased dopamine turnover exclusively in the nucleus accumbens (Hajnal and Norgren, 2002).
In our study, the effect of rimonabant on the bingeing group could be related to its capacity to block dopamine release in the nucleus accumbens shell that might be induced by the consumption of margarine, and by a possible enhancement in the tone of the endocannabinoid system. Chronic exposure to high-fat palatable diet was found to decrease the expression of CB1 receptors in the nucleus accumbens (Harrold et al., 2002). Accordingly, Bello et al. (2012) reported a reduction in CB1 receptor density in the same central area in an animal model of BED. The reduction in CB1 receptor expression could be interpreted as the resulting effect of increased EC levels that in turn could induce dopamine release in this area (Bermudez-Silva et al., 2012). As mentioned before, CB1 receptors are expressed in the nucleus accumbens and are mainly located at the presynaptic level, and an important functional consequence of their activation is the inhibition of the release of other neurotransmitters (Schlicker and Kathmann, 2001). Activation of CB1 receptors on axon terminals of glutamatergic neurons in the nucleus accumbens would inhibit glutamate release, thus inhibiting the GABAergic neurotransmission in the ventral tegmental area, consequently disinhibiting VTA dopamine neurons (Melis et al., 2004; Riegel and Lupica, 2004). Thus, it is possible that rimonabant, by blocking the inhibitory effect of ECs, elicits stimulation of GABA release, resulting in reduced firing activity of dopaminergic neurons and reduced release of dopamine in terminal areas. On the other hand, clinical studies have found elevated EC plasma levels in women with BED, and this increase may drive the binge episodes and reinforce the rewarding effects of palatable foods, promoting the cycle of binge eating (Monteleone et al., 2005). However, our results do not exclude the possibility that rimonabant can produce its effect acting as an inverse agonist but also through one or more CB1 receptor-independent mechanisms (Pertwee, 2010).
An overlap between compulsive overeating and drug abuse has been established, including the occurrence of craving and the loss of the inhibitory control over food intake and drug use. Notably, the choice of using the low dose (0.3 mg·kg−1) of rimonabant for chronic treatment was due to the fact that the same dose is able to antagonize i.v. self-administration of the CB1 receptor agonist WIN 55 212-2 as well as the reinstatement of drug-seeking behaviour in abstinent rats (Fattore et al., 2001; 2005), including female rats (Fattore et al., 2010).
In conclusion, our results provide the first evidence that a chronic low dose of a CB1 receptor antagonist reduces fat intake in female rats showing binge-type eating behaviour, with a concomitant significant weight loss without development of any tolerance. In this respect, our data suggest the potential therapeutic utility of CB1 receptor antagonists in the treatment of binge-like eating disorders. Further studies are needed to characterize the exact mechanisms and brain areas mediating such effects.
Acknowledgments
This work was partially supported by grant from the Italian Ministry of University and Scientific Research (FAR DM28141 del 21/11/2005) and by the Intramural Research Program of the National Institute on Drug Abuse, National Institutes of Health, Department of Health and Human Services, Baltimore, MD, USA.
Glossary
- AEA
anandamide
- AM251
1-(2, 4-dichlorophenyl)-5-(4-iodophenyl)-4-methyl-N-1-piperidinyl-1H-pyrazole-3-carboxamide
- BED
binge eating disorder
- CB1 receptor
subtype 1 cannabinoid receptor
- CB2 receptor
subtype 2 cannabinoid receptor
- ECs
endocannabinoids (endogenous ligands)
- FAAH
enzyme fatty acid amide hydrolase
- THC
Δ9-tetrahydrocannabinol
- URB597
cyclohexyl carbamic acid 3′-carbamoyl-3-yl ester
Conflict of interest
None.
References
- Ameri A. The effects of cannabinoids on the brain. Prog Neurobiol. 1999;58:315–348. doi: 10.1016/s0301-0082(98)00087-2. [DOI] [PubMed] [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders. 4th ed., Text Revision (DSM-IV-TR) Washington, DC: American Psychiatric Association; 2000. [Google Scholar]
- Arnone M, Maruani J, Chaperon F, Thiebot M, Poncelet M, Soubrie P, et al. Selective inhibition of sucrose and ethanol intake by SR 141716, an antagonist of central cannabinoid (CB1) receptors. Psychopharmacology. 1997;132:104–106. doi: 10.1007/s002130050326. [DOI] [PubMed] [Google Scholar]
- Avena NM, Rada P, Hoebel BG. Sugar and fat bingeing have notable differences in addictive-like behavior. J Nutr. 2008;139:623–628. doi: 10.3945/jn.108.097584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassareo V, Di Chiara G. Modulation of feeding-induced activation of mesolimbic dopamine transmission by appetitive stimuli and its relation to motivational state. Eur J Neurosci. 1999;11:4389–4397. doi: 10.1046/j.1460-9568.1999.00843.x. [DOI] [PubMed] [Google Scholar]
- Bello NT, Sweigart KL, Lakoski JM, Norgren R, Hajnal A. Restricted feeding with scheduled sucrose access results in an upregulation of the rat dopamine transporter. Am J Physiol Regul Integr Comp Physiol. 2003;284:1260–1268. doi: 10.1152/ajpregu.00716.2002. [DOI] [PubMed] [Google Scholar]
- Bello NT, Coughlin JW, Redgrave GW, Ladenheim EE, Moran TH, Guarda AS. Dietary conditions and highly palatable food access alter rat cannabinoid receptor expression and binding density. Physiol Behav. 2012;105:720–726. doi: 10.1016/j.physbeh.2011.09.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bermudez-Silva FJ, Cardinal P, Cota D. The role of the endocannabinoid system in the neuroendocrine regulation of energy balance. J Psychopharmacol. 2012;26:114–124. doi: 10.1177/0269881111408458. [DOI] [PubMed] [Google Scholar]
- Berner LA, Bocarsly ME, Hoebel BG, Avena NM. Pharmacological interventions for binge eating: lessons from animal models, current treatments, and future directions. Curr Pharm Des. 2011;17:1180–1187. doi: 10.2174/138161211795656774. [DOI] [PubMed] [Google Scholar]
- Bisogno T, Melck D, De Petrocellis L, Bobrov MY, Gretskaya NM, Bezuglov VV, et al. Arachidonoylserotonin and other novel inhibitors of fatty acid amide hydrolase. Biochem Biophys Res Commun. 1998;248:515–522. doi: 10.1006/bbrc.1998.8874. [DOI] [PubMed] [Google Scholar]
- Carai MA, Colombo G, Maccioni P, Gessa GL. Efficacy of rimonabant and other cannabinoid CB1 receptor antagonists in reducing food intake and body weight: preclinical and clinical data. CNS Drug Rev. 2006;12:91–99. doi: 10.1111/j.1527-3458.2006.00091.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cervino C, Vicennati V, Pasquali R, Pagotto U. Feeding disorders and obesity. Curr Top Behav Neurosci. 2009;1:373–385. doi: 10.1007/978-3-540-88955-7_15. [DOI] [PubMed] [Google Scholar]
- Colantuoni C, Schwenker J, McCarthy J, Rada P, Ladenheim B, Cadet JL, et al. Excessive sugar intake alters binding to dopamine and mu-opioid receptors in the brain. Neuroreport. 2001;12:3549–3552. doi: 10.1097/00001756-200111160-00035. [DOI] [PubMed] [Google Scholar]
- Colombo G, Agabio R, Diaz G, Lobina C, Reali R, Gessa GL. Appetite suppression and weight loss after the cannabinoid antagonist SR 141716. Life Sci. 1998;63:PL113–PL117. doi: 10.1016/s0024-3205(98)00322-1. [DOI] [PubMed] [Google Scholar]
- Corwin RL, Buda-Levin A. Behavioral models of binge-type eating. Physiol Behav. 2004;82:123–130. doi: 10.1016/j.physbeh.2004.04.036. [DOI] [PubMed] [Google Scholar]
- Corwin RL, Wojnicki FH. Binge eating in rats with limited access to vegetable shortening. 2006. Curr Protoc Neurosci Chapter 9: Unit 9.23B. [DOI] [PubMed]
- Corwin RL, Wojnicki FH, Fisher JO, Dimitriou SG, Rice HB, Young MA. Limited access to a dietary fat option affects ingestive behavior but not body composition in male rats. Physiol Behav. 1998;65:545–553. doi: 10.1016/s0031-9384(98)00201-7. [DOI] [PubMed] [Google Scholar]
- Cota D, Tschop MH, Horvath TL, Levine AS. Cannabinoids, opioids and eating behavior: the molecular face of hedonism? Brain Res Rev. 2006;51:85–107. doi: 10.1016/j.brainresrev.2005.10.004. [DOI] [PubMed] [Google Scholar]
- Davis C, Claridge G. The eating disorders as addiction: a psychobiological perspective. Addict Behav. 1998;23:463–475. doi: 10.1016/s0306-4603(98)00009-4. [DOI] [PubMed] [Google Scholar]
- Despres JP, Ross R, Boka G, Almeras N, Lemieux I. Effect of rimonabant on the high-triglyceride/low-HDL-cholesterol dyslipidemia, intraabdominal adiposity, and liver fat: the ADAGIO-Lipids trial. Arterioscler Thromb Vasc Biol. 2009;29:416–423. doi: 10.1161/ATVBAHA.108.176362. [DOI] [PubMed] [Google Scholar]
- Di Marzo V, Matias I. Endocannabinoid control of food intake and energy balance. Nat Neurosci. 2005;8:585–589. doi: 10.1038/nn1457. [DOI] [PubMed] [Google Scholar]
- Dimitriou SG, Rice HB, Corwin RL. Effects of limited access to a fat option on food intake and body composition in female rats. Int J Eat Disord. 2000;28:436–445. doi: 10.1002/1098-108x(200012)28:4<436::aid-eat12>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- Dipatrizio NV, Simansky KJ. Inhibiting parabrachial fatty acid amide hydrolase activity selectively increases the intake of palatable food via cannabinoid CB1 receptors. Am J Physiol Regul Integr Comp Physiol. 2008;295:1409–1414. doi: 10.1152/ajpregu.90484.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engel SG, Boseck JJ, Crosby RD, Wonderlich SA, Mitchell JE, Smyth J, et al. The relationship of momentary anger and impulsivity to bulimic behavior. Behav Res Ther. 2007;45:437–447. doi: 10.1016/j.brat.2006.03.014. [DOI] [PubMed] [Google Scholar]
- European Medicines Agency (London, UK) 2009. The European Medicines Agency recommends suspension of the marketing authorization of Acomplia. Available at: http://www.emea.europa.eu/humandocs/Humans/EPAR/acomplia/acomplia.htn (accessed 1/30/2009)
- Fattore L, Cossu G, Martellotta CM, Fratta W. Intravenous self-administration of the cannabinoid CB1 receptor agonist WIN 55,212-2 in rats. Psychopharmacology. 2001;156:410–416. doi: 10.1007/s002130100734. [DOI] [PubMed] [Google Scholar]
- Fattore L, Spano S, Cossu G, Deiana S, Fadda P, Fratta W. Cannabinoid CB1 antagonist SR 141716A attenuates reinstatement of heroin self-administration in heroin-abstinent rats. Neuropharmacology. 2005;48:1097–1104. doi: 10.1016/j.neuropharm.2005.01.022. [DOI] [PubMed] [Google Scholar]
- Fattore L, Fadda P, Fratta W. Endocannabinoid regulation of relapse mechanisms. Pharmacol Res. 2007;56:418–427. doi: 10.1016/j.phrs.2007.09.004. [DOI] [PubMed] [Google Scholar]
- Fattore L, Melis M, Fadda P, Pistis M, Fratta W. The endocannabinoid system and nondrug rewarding behaviours. Exp Neurol. 2010;224:23–36. doi: 10.1016/j.expneurol.2010.03.020. [DOI] [PubMed] [Google Scholar]
- Fegley D, Gaetani S, Duranti A, Tontini A, Mor M, Tarzia G, et al. Characterization of the fatty acid amide hydrolase inhibitor cyclohexylcarbamicacid 3_-carbamoyl-biphenyl-3-yl ester (URB597): effects on anandamide and oleoylethanolamide deactivation. J Pharmacol Exp Ther. 2005;313:352–358. doi: 10.1124/jpet.104.078980. [DOI] [PubMed] [Google Scholar]
- Fisher JO, Birch LL. Restricting access to palatable foods affects children's behavioral response, food selection, and intake. Am J Clin Nutr. 1999;69:1264–1272. doi: 10.1093/ajcn/69.6.1264. [DOI] [PubMed] [Google Scholar]
- Foltin RW, Haney M. Effects of the cannabinoid antagonist SR141716 (Rimonabant) and d-amphetamine on palatable food and food pellet intake in non-human primates. Pharmacol Biochem Behav. 2007;86:766–773. doi: 10.1016/j.pbb.2007.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu J, Gaetani S, Oveisi F, Lo Verme J, Serrano A, Rodríguez De Fonseca F, et al. Oleylethanolamide regulates feeding and body weight through activation of the nuclear receptor PPAR-alpha. Nature. 2003;425:90–93. doi: 10.1038/nature01921. [DOI] [PubMed] [Google Scholar]
- Gaetani S, Kaye WH, Cuomo V, Piomelli D. Role of endocannabinoids and their analogues in obesity and eating disorders. Eat Weight Disord. 2008;13:e42–e48. [PubMed] [Google Scholar]
- Grilo CM, White MA, Masheb RM. DSM-IV psychiatric disorder comorbidity and its correlates in binge eating disorder. Int J Eat Disord. 2009;42:228–234. doi: 10.1002/eat.20599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajnal A, Norgren R. Repeated access to sucrose augments dopamine turnover in the nucleus accumbens. Neuroreport. 2002;13:2213–2216. doi: 10.1097/00001756-200212030-00010. [DOI] [PubMed] [Google Scholar]
- Hancock SD, Olmstead MC. Animal models of eating disorders. In: Olmstead MC, editor. Animal Models of Drug Addiction. Totowa, NJ: Humana Press Inc; 2010. pp. 207–234. [Google Scholar]
- Harrold JA, Williams G. The cannabinoid system: a role in both the homeostatic and hedonic control of eating? Br J Nutr. 2003;90:729–734. doi: 10.1079/bjn2003942. [DOI] [PubMed] [Google Scholar]
- Harrold JA, Elliott JC, King PJ, Widdowson PS, Williams G. Down-regulation of cannabinoid-1 (CB-1) receptors in specific extrahypothalamic regions of rats with dietary obesity: a role for endogenous cannabinoids in driving appetite for palatable food? Brain Res. 2002;952:232–238. doi: 10.1016/s0006-8993(02)03245-6. [DOI] [PubMed] [Google Scholar]
- Hart L, Ward AS, Haney M, Comer SD, Foltin R, Fishman MW. Comparison of smoked marijuana and oral D9-tetrahydrocannabinol in humans. Psychopharmacology. 2002;164:407–415. doi: 10.1007/s00213-002-1231-y. [DOI] [PubMed] [Google Scholar]
- Herkenham M, Lynn AB, Little MD, Johnson MR, Melvin LS, de Costa BR, et al. Cannabinoid receptor localization in brain. Proc Natl Acad Sci USA. 1990;87:1932–1936. doi: 10.1073/pnas.87.5.1932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hudson JI, Hiripi E, Pope HG, Jr, Kessler RC. The prevalence and correlates of eating disorders in the National Comorbidity Survey Replication. Biol Psychiatry. 2007;61:348–358. doi: 10.1016/j.biopsych.2006.03.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jamshidi N, Taylor DA. Anandamide administration into the ventromedial hypothalamus stimulates appetite in rats. Br J Pharmacol. 2001;134:1151–1154. doi: 10.1038/sj.bjp.0704379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson JG, Spitzer RL, Williams JB. Health problems, impairment and illnesses associated with bulimia nervosa and binge eating disorder among primary care and obstetric gynaecology patients. Psychol Med. 2001;31:1455–1466. doi: 10.1017/s0033291701004640. [DOI] [PubMed] [Google Scholar]
- Kales EF. Macronutrient analysis of binge eating in bulimia. Physiol Behav. 1990;48:837–840. doi: 10.1016/0031-9384(90)90236-w. [DOI] [PubMed] [Google Scholar]
- Kathuria S, Gaetani S, Fegley D, Valino F, Duranti A, Tontini A, et al. Modulation of anxiety through blockade of anandamide hydrolysis. Nat Med. 2003;9:76–81. doi: 10.1038/nm803. [DOI] [PubMed] [Google Scholar]
- Kelley AE, Schiltz CA, Landry CF. Neural systems recruited by drug- and food-related cues: studies of gene activation in corticolimbic regions. Physiol Behav. 2005;86:11–14. doi: 10.1016/j.physbeh.2005.06.018. [DOI] [PubMed] [Google Scholar]
- Kilkenny C, Browne W, Cuthill IC, Emerson M, Altman DG. NC3Rs Reporting Guidelines Working Group. Br J Pharmacol. 2010;160:1577–1579. doi: 10.1111/j.1476-5381.2010.00872.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirkham TC, Williams CM, Fezza F, Di Marzo V. Endocannabinoid levels in rat limbic forebrain and hypothalamus in relation to fasting, feeding and satiation: stimulation of eating by 2-arachidonoyl glycerol. Br J Pharmacol. 2002;136:550–557. doi: 10.1038/sj.bjp.0704767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koch JE. Delta(9)-THC stimulates food intake in Lewis rats: effects on chow, high-fat and sweet high-fat diets. Pharmacol Biochem Behav. 2001;68:539–543. doi: 10.1016/s0091-3057(01)00467-1. [DOI] [PubMed] [Google Scholar]
- Li C, Jones PM, Persaud SJ. Role of the endocannabinoid system in food intake, energy homeostasis and regulation of the endocrine pancreas. Pharmacol Ther. 2011;129:307–320. doi: 10.1016/j.pharmthera.2010.10.006. [DOI] [PubMed] [Google Scholar]
- Liang NC, Hajnal A, Norgren R. Sham feeding corn oil increases accumbens dopamine in the rat. Am J Physiol Regul Integr Comp Physiol. 2006;291:1236–1239. doi: 10.1152/ajpregu.00226.2006. [DOI] [PubMed] [Google Scholar]
- Lo Verme J, Gaetani S, Fu J, Oveisi F, Burton K, Piomelli D. Regulation of food intake by oleoylethanolamide. Cell Mol Life Sci. 2005;62:708–716. doi: 10.1007/s00018-004-4494-0. [DOI] [PubMed] [Google Scholar]
- McGrath J, Drummond G, McLachlan E, Kilkenny C, Wainwright C. Guidelines for reporting experiments involving animals: the ARRIVE guidelines. Br J Pharmacol. 2010;160:1573–1576. doi: 10.1111/j.1476-5381.2010.00873.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McLaughlin PJ, Winston K, Swezey L, Wisniecki A, Aberman J, Tardif DJ, et al. The cannabinoid CB1 antagonists SR 141716A and AM 251 suppress food intake and food-reinforced behavior in a variety of tasks in rats. Behav Pharmacol. 2003;14:583–588. doi: 10.1097/00008877-200312000-00002. [DOI] [PubMed] [Google Scholar]
- Mahler SV, Smith KS, Berridgem KC. Endocannabinoid hedonic hotspot for sensory pleasure: anandamide in nucleus accumbens shell enhances ‘liking’ of a sweet reward. Neuropsychopharmacology. 2007;32:2267–2278. doi: 10.1038/sj.npp.1301376. [DOI] [PubMed] [Google Scholar]
- Marazziti D, Rossi L, Baroni S, Consoli G, Hollander E, Catena-Dell'Osso M. Novel treatment options of binge eating disorder. Curr Med Chem. 2011;18:5159–5164. doi: 10.2174/092986711797636045. [DOI] [PubMed] [Google Scholar]
- Marco EM, García-Gutiérrez MS, Bermúdez-Silva FJ, Moreira FA, Guimarães F, Manzanares J, et al. Endocannabinoid system and psychiatry: in search of a neurobiological basis for detrimental and potential therapeutic effects. Front Behav Neurosci. 2011;5:63. doi: 10.3389/fnbeh.2011.00063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marcus MD, Kalarchian MA. Binge eating in children and adolescents. Int J Eat Disord. 2003;34:S47–S57. doi: 10.1002/eat.10205. [DOI] [PubMed] [Google Scholar]
- Martel P, Fantino M. Mesolimbic dopaminergic system activity as a function of food reward: a microdialysis study. Pharmacol Biochem Behav. 1996;53:221–226. doi: 10.1016/0091-3057(95)00187-5. [DOI] [PubMed] [Google Scholar]
- Martín-García E, Burokas A, Martín M, Berrendero F, Rubí B, Kiesselbach C, et al. Central and peripheral consequences of the chronic blockade of CB1 cannabinoid receptor with rimonabant or taranabant. J Neurochem. 2010;112:1338–13351. doi: 10.1111/j.1471-4159.2009.06549.x. [DOI] [PubMed] [Google Scholar]
- Mathes CM, Ferrara M, Rowland NE. Cannabinoid CB1 receptor antagonists reduce caloric intake by decreasing palatable diet selection in a novel dessert protocol in female rats. Am J Physiol Regul Integr Comp Physiol. 2008;295:67–75. doi: 10.1152/ajpregu.00150.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melis M, Pistis M, Perra S, Muntoni AL, Pillolla G, Gessa GL. Endocannabinoids mediate presynaptic inhibition of glutamatergic transmission in rat ventral tegmental area dopamine neurons through activation of CB1 receptors. J Neurosci. 2004;24:53–62. doi: 10.1523/JNEUROSCI.4503-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Melis MR, Succu S, Mascia MS, Sanna F, Melis T, Castelli MP, et al. The cannabinoid receptor antagonist SR-141716A induces penile erection in male rats: involvement of paraventricular glutamic acid and nitric oxide. Neuropharmacology. 2007;50:219–228. doi: 10.1016/j.neuropharm.2005.09.009. [DOI] [PubMed] [Google Scholar]
- Monteleone P, Matias I, Martiadis V, De Petrocellis L, Maj M, Di Marzo V. Blood levels of the endocannabinoid anandamide are increased in anorexia nervosa and in binge-eating disorder, but not in bulimia nervosa. Neuropsychopharmacology. 2005;30:1216–1221. doi: 10.1038/sj.npp.1300695. [DOI] [PubMed] [Google Scholar]
- Monteleone P, Tortorella A, Martiadis V, Di Filippo C, Canestrelli B, Maj M. The cDNA 385C to A missense polymorphism of the endocannabinoid degrading enzyme fatty acid amide hydrolase (FAAH) is associated with overweight/obesity but not with binge eating disorder in overweight/obese women. Psychoneuroendocrinology. 2008;33:546–550. doi: 10.1016/j.psyneuen.2008.01.004. [DOI] [PubMed] [Google Scholar]
- Neumark-Sztainer D, Story M, Hannan PJ, Perry CL, Irving LM. Weight-related concerns and behaviors among overweight and nonoverweight adolescents: implications for preventing weight-related disorders. Arch Pediatr Adolesc Med. 2002;156:171–178. doi: 10.1001/archpedi.156.2.171. [DOI] [PubMed] [Google Scholar]
- O'Sullivan SE. Cannabinoids go nuclear: evidence for activation of peroxisome proliferator-activated receptors. Br J Pharmacol. 2007;52:576–582. doi: 10.1038/sj.bjp.0707423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oliver G, Wardle J. Perceived effects of stress on food choice. Physiol Behav. 1999;66:511–515. doi: 10.1016/s0031-9384(98)00322-9. [DOI] [PubMed] [Google Scholar]
- Orio L, Edwards S, George O, Parsons LH, Koob GF. A role for the endocannabinoid system in the increased motivation for cocaine in extended-access conditions. J Neurosci. 2009;29:4846–4857. doi: 10.1523/JNEUROSCI.0563-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parylak SL, Cottone P, Sabino V, Rice KC, Zorrilla E. Effects of CB(1) and CRF(1) receptor antagonists on binge-like eating in rats with limited access to a sweet fat diet: lack of withdrawal-like responses. Physiol Behav. 2012;107:231–242. doi: 10.1016/j.physbeh.2012.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pertwee RG. Receptors and channels targeted by synthetic cannabinoid receptor agonists and antagonists. Curr Med Chem. 2010;17:1360–1381. doi: 10.2174/092986710790980050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piomelli D, Tarzia G, Duranti A, Tontini A, Mor M, Compton TR, et al. Pharmacological profile of the selective FAAH inhibitor KDS-4103 (URB597) CNS Drug Rev. 2006;12:21–38. doi: 10.1111/j.1527-3458.2006.00021.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polivy J. Psychological consequences of food restriction. J Am Diet Assoc. 1996;96:589–592. doi: 10.1016/S0002-8223(96)00161-7. [DOI] [PubMed] [Google Scholar]
- Preti A, Girolamo G, Vilagut G, Alonso J, Graaf R, Bruffaerts R, et al. ESEMeD-WMH Investigators. The epidemiology of eating disorders in six European countries: results of the ESEMeD-WMH project. J Psychiatr Res. 2009;43:1125–1132. doi: 10.1016/j.jpsychires.2009.04.003. [DOI] [PubMed] [Google Scholar]
- Rada P, Avena NM, Hoebel BG. Daily bingeing on sugar repeatedly releases dopamine in the accumbens shell. Neuroscience. 2005;134:737–744. doi: 10.1016/j.neuroscience.2005.04.043. [DOI] [PubMed] [Google Scholar]
- Riegel AC, Lupica CR. Independent presynaptic and postsynaptic mechanisms regulate endocannabinoid signaling at multiple synapses in the ventral tegmental area. J Neurosci. 2004;24:11070–11078. doi: 10.1523/JNEUROSCI.3695-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scheen AJ. CB1 receptor blockade and its impact on cardiometabolic risk factors: overview of the RIO programme with rimonabant. J Neuroendocrinol. 2008;20:139–146. doi: 10.1111/j.1365-2826.2008.01681.x. [DOI] [PubMed] [Google Scholar]
- Scherma M, Medalie J, Fratta W, Vadivel SK, Makriyannis A, Piomelli D, et al. The endogenous cannabinoid anandamide has effects on motivation and anxiety that are revealed by fatty acid amide hydrolase (FAAH) inhibition. Neuropharmacology. 2008a;54:129–140. doi: 10.1016/j.neuropharm.2007.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scherma M, Panlilio LV, Fadda P, Fattore L, Gamaleddin I, Le Foll B, et al. Inhibition of anandamide hydrolysis by cyclohexyl carbamic acid 3′-carbamoyl-3-yl ester (URB597) reverses abuse-related behavioral and neurochemical effects of nicotine in rats. J Pharmacol Exp Ther. 2008b;327:482–490. doi: 10.1124/jpet.108.142224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlicker E, Kathmann M. Modulation of transmitter release via presynaptic cannabinoid receptors. Trends Pharmacol Sci. 2001;22:565–572. doi: 10.1016/s0165-6147(00)01805-8. [DOI] [PubMed] [Google Scholar]
- Schmidt HD, Anderson SM, Famous KR, Kumaresan V, Pierce RC. Anatomy and pharmacology of cocaine priming-induced reinstatement of drug seeking. Eur J Pharmacol. 2005;526:65–76. doi: 10.1016/j.ejphar.2005.09.068. [DOI] [PubMed] [Google Scholar]
- Serrano A, Parsons LH. Endocannabinoid influence in drug reinforcement, dependence and addiction-related behaviors. Pharmacol Ther. 2011;132:215–241. doi: 10.1016/j.pharmthera.2011.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siegfried Z, Kanyas K, Latzer Y, Karni O, Bloch M, Lerer B, et al. Association study of cannabinoid receptor gene (CNR1) alleles and anorexia nervosa: differences between restricting and binging/purging subtypes. Am J Med Genet B Neuropsychiatr Genet. 2004;125:126–130. doi: 10.1002/ajmg.b.20089. [DOI] [PubMed] [Google Scholar]
- Simiand J, Keane M, Keane PE, Soubrie P. SR 141716, a CB1 cannabinoid receptor antagonist, selectively reduces sweet food intake in marmoset. Behav Pharmacol. 1998;9:179–181. [PubMed] [Google Scholar]
- Sipe JC, Waalen J, Gerber A, Beutler E. Overweight and obesity associated with a missense polymorphism in fatty acid amide hydrolase (FAAH) Int J Obes. 2005;29:755–759. doi: 10.1038/sj.ijo.0802954. [DOI] [PubMed] [Google Scholar]
- Solinas M, Justinova Z, Goldberg SR, Tanda G. Anandamide administration alone and after inhibition of fatty acid amide hydrolase (FAAH) increases dopamine levels in the nucleus accumbens shell in rats. J Neurochem. 2006;98:408–419. doi: 10.1111/j.1471-4159.2006.03880.x. [DOI] [PubMed] [Google Scholar]
- Solinas M, Goldberg SR, Piomelli D. The endocannabinoid system in brain reward processes. Br J Pharmacol. 2008;154:369–383. doi: 10.1038/bjp.2008.130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soria-Gòmez E, Matias I, Rueda-Orozco PE, Cisneros M, Petrosino S, Navarro L, et al. Pharmacological enhancement of the endocannabinoid system in the nucleus accumbens shell stimulates food intake and increases c-Fos expression in the hypothalamus. Br J Pharmacol. 2007;151:1109–1116. doi: 10.1038/sj.bjp.0707313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wegner KE, Smyth JM, Crosby RD, Wittrock D, Wonderlich SA, Mitchell JE. An evaluation of the relationship between mood and binge eating in the natural environment using ecological momentary assessment. Int J Eat Disord. 2002;32:352–361. doi: 10.1002/eat.10086. [DOI] [PubMed] [Google Scholar]
- Wiederman MW, Pryor T. Substance use among women with eating disorders. Int J Eat Disord. 1996;20:163–168. doi: 10.1002/(SICI)1098-108X(199609)20:2<163::AID-EAT6>3.0.CO;2-E. [DOI] [PubMed] [Google Scholar]
- Wiley JL, Burston JJ, Leggett DC, Alekseeva OO, Razdan RK, Mahadevan A, et al. CB1 cannabinoid receptor-mediated modulation of food intake in mice. Br J Pharmacol. 2005;145:293–300. doi: 10.1038/sj.bjp.0706157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams CM, Kirkham TC. Anandamide induces overeating: mediation by central cannabinoid (CB1) receptors. Psychopharmacology. 1999;143:315–317. doi: 10.1007/s002130050953. [DOI] [PubMed] [Google Scholar]
- Wise RA, Newton P, Leeb K, Burnette B, Pocock D, Justice JB. Fluctuations in nucleus accumbens dopamine concentration during intravenous cocaine self-administration in rats. Psychopharmacology. 1995;120:10–20. doi: 10.1007/BF02246140. [DOI] [PubMed] [Google Scholar]
- Yanovski SZ, Nelson JE, Dubbert BK, Spitzer RL. Association of binge eating disorder and psychiatric comorbidity in obese subjects. Am J Psychiatry. 1993;150:1472–1479. doi: 10.1176/ajp.150.10.1472. [DOI] [PubMed] [Google Scholar]
- Zanettini C, Panlilio LV, Alicki M, Goldberg SR, Haller J, Yasar S. Effects of endocannabinoid system modulation on cognitive and emotional behavior. Front Behav Neurosci. 2011;5:57. doi: 10.3389/fnbeh.2011.00057. [DOI] [PMC free article] [PubMed] [Google Scholar]