

Abstract
The whipworm of humans, Trichuris trichiura, is responsible for a neglected tropical disease (NTD) of major importance in tropical and subtropical countries of the world. Whipworms also infect animal hosts, including pigs, dogs and non-human primates, cause clinical disease (trichuriasis) similar to that of humans. Although Trichuris species are usually considered to be host specific, it is not clear whether non-human primates are infected with T. trichiura or other species. In the present study, we sequenced the complete mitochondrial (mt) genome as well as the first and second internal transcribed spacers (ITS-1 and ITS-2) of Trichuris from the François’ leaf-monkey (langur), and compared them with homologous sequences from human- and pig-derived Trichuris. In addition, sequence comparison of a conserved mt ribosomal gene among multiple individual whipworms revealed substantial nucleotide differences among these three host species but limited sequence variation within each of them. The molecular data indicate that the monkey-derived whipworm is a separate species from that of humans. Future work should focus on detailed population genetic and morphological studies (by electron microscopy) of whipworms from various non-humans primates and humans.
Introduction
Neglected tropical diseases (NTDS) have a devastating effect on animal and human health and food production globally. For instance, it is estimated that more than two billion people are infected with geohelminths, including the Ascaris (common roundworm), Necator, Ancylostoma (hookworms) and Trichuris (whipworm), mainly in underprivileged areas of the world [1]. Trichuris trichiura is a very common parasite of humans in developing countries, and causes trichuriasis in ∼ 600 million people worldwide, mainly in children aged between 5 and 15 years [2]. Trichuriasis can be associated with intestinal symptoms, such as abdominal pain, dysentery, nausea, vomiting, anorexia, constipation and chronic appendiceal syndrome [2]. Whipworms also infect a broad range of other hosts, including pigs (T. suis), dogs (T. vulpis), sheep (T. ovis), goats (T. skrjabini), rats (T. muris) and non-human primates, and can cause clinical disease similar to trichuriasis of humans [3]–[7].
Trichuris infects non-human primates in many countries, including Belgium [7], China [8], Ethiopia [9], Kenya [10], [11], Peru [12], South Africa [13]. In spite of the high prevalence of Trichuris sometimes reported in non-human primates [13], it is not clear whether the non-human primates harbour T. trichiura or other congeners. Based on morphological features of adult worms, Trichuris of non-human primates (including Trichuris cynocephalus and T. rhinopithecus) have been regarded as T. trichiura [14], [15]. However, the identification of Trichuris to species using morphological criteria alone is not reliable. Moreover, neither larval or egg stages of Trichuris from humans, pigs and non-human primates can be identified or differentiated unequivocally to species using classical diagnostic approaches [14], [16]. Therefore, there is a need for suitable molecular approaches to accurately identify and distinguish closely-related Trichuris species from different hosts.
Molecular tools, using genetic markers in mitochondrial (mt) DNA and in the internal transcribed spacer (ITS) regions of nuclear ribosomal DNA (rDNA), have been used effectively to identify nematode species [17]–[21]. For whipworms, mtDNA has been used in China to show clear genetic distinctiveness between human- and pig-derived Trichuris [22], and between T. ovis and T. discolor from ruminant hosts [23]. Using ITS rDNA, recent studies of Trichuris specimens obtained from humans and pigs [24], [25] also indicate that T. trichiura and T. suis are separate species. Cutillas et al. [26] used the ITS rDNA to infer the existence of two separate Trichuris species in murid and arvicolid rodents. In other studies from Spain, ITS rDNA has also been employed to distinguish among T. suis from swine, T. vulpis from dogs [27] and T. trichiura from the non-human primates (i.e. Pan troglodytes, Colobus guereza kikuyensis and Nomascus gabriellae) [15]. Although a recent investigation has shown two distinct Trichuris genotypes infecting both humans and non-human primates [28], there is still a paucity of information on Trichuris from different species of primates and countries around the world. Therefore, in the present study, we characterized the mt genomic and ITS rDNA sequences of Trichuris from the endangered François’ leaf-monkey (Trachypithecus (Presbytis) françoisi), which usually lives in close proximity to human populations in southern China [29], and we compared them with homologous sequences of human- and pig-derived Trichuris, and then tested the hypothesis that this monkey-derived Trichuris is a separate species.
Materials and Methods
Ethics Statement
This study did not require approval by an ethics committee. Two François’ leaf-monkeys, from which Trichuris specimens were collected from their caeca post-mortem, were handled and housed in a zoo in strict accordance with good animal practices required by the Animal Ethics Procedures and Guidelines of the People’s Republic of China. The monkeys were caged, and there were two rooms in a cage; one was indoor and the other was outdoor. They were fed fruits and vegetables. The monkeys were under the care and treatment of a licensed veterinarian at the zoo, and were euthanized due to acute gastric dilation.
Parasites and Isolation of Total Genomic DNA
Two adult specimens of Trichuris (designated “monkey-Trichuris”) were collected from each of the two François’ leaf-monkeys, and were washed in physiological saline, identified morphologically [14], fixed in 70% (v/v) ethanol and stored at –20°C until use. Total genomic DNA was isolated separately from four individual worms (coded TH1-TH4) using an established method [30].
Long-range PCR-based Sequencing of mt DNA
To obtain some mt sequence data for primer design, we PCR-amplified regions (400–500 bp) of the cox1 gene by using a (relatively) conserved primer pair JB3-JB4.5 [31], and nad5 gene was amplified using primers NAD5F (forward; 5′- CAAGGATTTTTTTGAGATCTTTTTC-3′) and NAD5R (reverse; 5′- TAAACCGAATTGGAGATTTTTGTTT-3′) designed to conserved regions of sequence between human- and pig-derived Trichuris. The amplicons were sequenced in both directions using BigDye terminator v.3.1, ABI PRISM 3730. We then designed primers (Table 1) to regions within cox1, nad5 and rrnL (based on sequences conserved between human- and pig-derived Trichuris) and amplified from total genomic DNA (from an individual worm, coded TH1) the entire mt genome in three overlapping fragments (each ∼ 5 kb): cox1-nad5, nad5-rrnL and rrnL-cox1. The cycling conditions used were 92°C for 2 min (initial denaturation), then 92°C/10 s (denaturation), 48–52°C/30 s (annealing), and 60°C/10 min (extension) for 10 cycles, followed by 92°C/10 s, 58–52°C/30 s, and 60°C/10 min for 20 cycles, with a cycle elongation of 10 s for each cycle and a final extension at 60°C/10 min. Each amplicon, which represented a single band in a 0.8% (w/v) agarose gel, following electrophoresis and ethidium-bromide staining, was column-purified and then sequenced using a primer walking strategy [32].
Table 1. Sequences of primers used to amplify mitochondrial DNA regions from monkey-Trichuris.
| Primer | Sequence (5′ to 3′) |
| THCO1F | GTTTTCTCTCTGGACCGATTACCTA |
| THND5R | TACTTTAGTAGTTGCAGGGGTTATC |
| THND5F | TAGGAGCAGCCATAGCGATAGGTAA |
| TH16SR | AATCACGTAATGTTCCATCGTCGAA |
| TH16SF | TAAACGAGAAGACCCTAGGAACTTG |
| THCO1R | CGAAAAGTATGTATATGGTGCCAAT |
Sequence Analyses
Sequences were assembled manually and aligned against the complete mt genome sequences of T. trichiura [22] using the computer program Clustal X 1.83 [33] to infer gene boundaries. The open reading frames (ORFs) were identified using ORFFinder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html) employing the invertebrate mitochondrial code, and subsequently compared with that of T. trichiura [22]. Translation initiation and termination codons were identified based on comparison with those reported previously [22]. The secondary structures of 22 tRNA genes were predicted using tRNAscan-SE [34] and/or manual adjustment [35], and rRNA genes were identified by comparison with those known for Trichuris [22].
Sequencing of ITS rDNA and mt rrnL
The full ITS rDNA region including primer flanking 18S and 28S rDNA sequences was PCR-amplified from individual DNA samples using universal primers NC5 (forward; 5′-GTAGGTGAACCTGCGGAAGGATCATT-3′) and NC2 (reverse; 5′-TTAGTTTCTTTTCCTCCGCT-3′) described previously [36]. The primers rrnLF (5′-AAAACTCGGCAAATCGCATACTAAT-3′) and rrnLR (5′- CGAGCCACAAGACAGTAATGATAAG -3′) designed to conserved mt genome sequences within the rrnL gene were employed for PCR amplification and subsequent sequencing of a portion (∼ 600 bp) of this gene from multiple individuals of monkey-derived Trichuris.
Phylogenetic Analyses
Amino acid sequences inferred from the 12 protein-coding genes (i.e. not atp-8) common among all of the nematodes included here were concatenated into a single alignment, and then aligned with those of four other enoplid nematodes (GenBank accession nos. GU385218, GU070737, JQ996232 and JQ996231 for T. trichiura, T. suis, T. ovis and T. discolor, respectively), using T. spiralis (accession no. NC_002681) [37] as an outgroup. Ambiguous sites and regions in the alignment were excluded using Gblocks (http://molevol.cmima.csic.es/castresana/Gblocks_server.html) [38] using default parameters. The rrnL sequences determined here and those of human- and pig-derived Trichuris [22] were aligned and subjected to phylogenetic analysis using Trichinella spiralis (accession no. NC_002681) [37] as an outgroup (cf. [39]). Phylogenetic analyses were conducted using Bayesian inference (BI), as described previously [40]. Phylograms were drawn using the program Tree View v.1.65 [41].
Results
Features of the Circular mt Genome of Trichuris from the François Leaf-monkey
The complete mt genome sequence was 14,147 bp in length (GenBank accession no. KC461179). The mt genome contains 13 protein-coding genes (cox1-3, nad1-6, nad4L, cytb, atp6 and atp8), 22 transfer RNA genes and two ribosomal RNA genes (rrnS and rrnL) (Table 2); the atp8 gene is encoded (Figure 1). The protein-coding genes are transcribed in different directions, as reported for T. trichiura and T. suis [22] (Table 2). Protein-coding genes were annotated by aligning sequences, and identifying translation initiation and termination codons by comparison with homologous sequences for other whipworms (Table 2).
Table 2. Positions and nucleotide sequence lengths of mitochondrial genome of monkey-Trichuris (MT) compared with that of human-Trichuris (T. trichiura = TT) and pig-Trichuris (T. suis = TS).
| Gene or region | Positions and nt sequence lengths (bp) | Strand | Initiation/termination codons | Anticodons | ||||
| MT | TT | TS | MT | TT | TS | MT/TT/TS | ||
| cox1 | 1–1536 | 1–1545 | 1–1542 | H | ATG/TAG | ATG/TAA | ATG/TAG | |
| cox2 | 1556 –2230 | 1560 –2234 | 1578 –2258 | H | ATG/TAA | ATG/TAA | ATG/TAA | |
| tRNA–LeuUUR (L2) | 2264–2328 (65) | 2251–2313 (63) | 2271–2332 (62) | H | TAA | |||
| tRNA–Glu (E) | 2339–2396 (58) | 2318–2374 (57) | 2337–2393 (57) | H | TTC | |||
| nad1 | 2413–3312 | 2397–3296 | 2415–3314 | H | ATT/TAA | ATA/TAA | ATT/TAG | |
| Non–coding region (NCR–L) | 3313–3436 (124) | 3297–3458 (162) | 3315–3458 (144) | H | ||||
| tRNA–Lys (K) | 3437–3501 (65) | 3459–3524 (66) | 3459–3521 (63) | H | TTT | |||
| nad2 | 3508–4407 | 4406–3522 | 4414–3533 | L | ATT/TAG | ATA/TAA | ATA/TAG | |
| tRNA–Met (M) | 4468–4408 (61) | 4479–4419 (61) | 4485–4424 (62) | L | CAT | |||
| tRNA–Phe (F) | 4463–4521 (59) | 4530–4474 (57) | 4546–4488 (59) | L | GAA | |||
| nad5 | 6070–4523 | 6078–4531 | 6094–4538 | L | ATA/TAA | ATA/TAA | ATA/TAG | |
| tRNA–His (H) | 6061–6122 (62) | 6128–6072 (57) | 6150–6094 (57) | L | GTG | |||
| tRNA–Arg (R) | 6188–6124 (65) | 6194–6130 (65) | 6218–6152 (67) | L | ACG | |||
| nad4 | 7401–6190 | 7406–6195 | 7432–6224 | L | ATG/TAA | ATG/TAA | ATA/TAA | |
| nad4L | 7420–7680 | 7682–7425 | 7901–7650 | L | ATA/TAA | ATA/TAA | ATA/TAG | |
| tRNA–Thr (T) | 7685–7744 (60) | 7687–7744 (58) | 7905–7962 (58) | H | TGT | |||
| tRNA–Pro (P) | 7796–7743 (54) | 7802–7744 (59) | 8023–7966 (58) | L | TGG | |||
| nad6 | 7798–8265 | 7795–8271 | 8016–8486 | H | ATG/TAA | ATT/TAA | ATT/TAA | |
| cytb | 8273–9379 | 8278–9384 | 8501–9613 | H | ATG/TAG | ATG/TAG | ATG/TAG | |
| tRNA–Ser AGN (S1) | 9378–9429 (52) | 9383–9432 (50) | 9612–9666 (55) | H | GCT | |||
| rrnS | 9422–10126 | 9425–10122 | 9664–10375 | H | ||||
| tRNA–Val (V) | 10129–10185 (57) | 10124–10180 (57) | 10375–10431 (57) | H | TAC | |||
| rrnL | 10192–11198 | 10180–11190 | 10440–11450 | H | ||||
| atp6 | 11175–11996 | 11173–12000 | 11422–12249 | H | ATA/TAA | ATA/TAA | ATA/TAA | |
| cox3 | 12001–12744 | 11975–12748 | 12259–13035 | H | ATG/TAA | ATG/TAA | ATG/TAA | |
| tRNA–Trp (W) | 12844–12779 (66) | 12817–12755 (63) | 13106–13040 (67) | L | TCA | |||
| tRNA–Gln (Q) | 12852–12908 (57) | 12821–12874 (54) | 13110–13166 (57) | H | TTG | |||
| tRNA–Ile (I) | 12992–12927 (66) | 12937–12871 (66) | 13234–13169 (66) | L | GAT | |||
| tRNA–Gly (G) | 13057–12996 (62) | 13003–12947 (57) | 13308–13253 (56) | L | TCC | |||
| tRNA–Asp (D) | 13063–13121 (59) | 13009–13067 (58) | 13302–13363 (62) | H | GTC | |||
| atp8 | 13109–13273 | 13055–13219 | 13360–13530 | H | ATA/TAG | ATA/TAG | TTG/TAA | |
| nad3 | 13286–13624 | 13229–13570 | 13555–13896 | H | ATT/TAA | ATA/TAA | ATA/TAA | |
| Non–coding region (NCR–S) | 13625–13729 (105) | 13571–13663 (93) | 13887–14003 (117) | H | ||||
| tRNA–Ser UCN (S2) | 13730– 13782 (53) | 13664–13715 (52) | 14004–14055 (52) | H | TGA | |||
| tRNA–Asn (N) | 13782–13836 (55) | 13715–13768 (54) | 14055–14113 (59) | H | GTT | |||
| tRNA–Leu CUN (L1) | 13841–13907 (67) | 13776–13842 (67) | 14131–14193 (63) | H | TAG | |||
| tRNA–Ala (A) | 13924–13983 (60) | 13845–13899 (57) | 14196–14250 (55) | H | TGC | |||
| tRN A–Cys (C) | 14065–14003 (63) | 13979–13925 (55) | 14274–14328 (55) | L | GCA | |||
| tRNA–Tyr (Y) | 14127–14068 (60) | 14046–13986 (50) | 14394–14336 (59) | L | TGT | |||
Figure 1. Structure of the mitochondrial genome for Trichuris from the François’ langur (Trichuris sp.).

Genes are designated according to standard nomenclature, except for the 22 tRNA genes, which are designated using one-letter amino acid codes, with numerals differentiating each of the two leucine- and serine-specifying tRNAs (L1 and L2 for codon families CUN and UUR, respectively; S1 and S2 for codon families AGN and UCN, respectively). “NCR-L” refers to a large non-coding region; “NCR-S” refers to a small non-coding region.
Twenty-two tRNA genes, which varied from 52 to 67 bp in length, were predicted from the mt genomes. The two ribosomal RNA genes (rrnL and rrnS) were inferred; rrnL is located between tRNA-Val and atp6, and rrnS is located between tRNA-Ser (AGN) and tRNA-Val. The lengths of rrnL and rrnS are 1,007 bp and 705 bp, respectively. The A+T contents of rrnL and rrnS are 69.02% and 70.21%, respectively.
Two AT-rich non-coding regions (NCRs) were inferred in the mt genome. For this genome, the long NCR (designated NCR-L; 124 bp in length) is located between the nad1 and tRNA-Lys (Figure 1), has an A+T content of 59.68%. The short NCR (NCR-S; 105 bp in length) is located between genes nad3 and tRNA-Ser (UCN) (Figure 1), with an A+T content of 79.25%.
Nuclear Ribosomal DNA Regions of Trichuris from the Monkey
The rDNA region including ITS-1, ITS-2 and intervening 5.8 rRNA gene sequenced from individual Trichuris samples (coded TH1-TH4) was 1,314 bp in length. Individual spacers were 570 bp (ITS-1) and 468 bp (ITS-2), and the 5.8S rRNA gene was 154 bp long.
Comparative Analyses Among Monkey-Trichuris, Human-Trichuris and Pig-Trichuris
The mt genome sequence of monkey-Trichuris (accession no. KC461179) was 14,147 bp in length, 101 bp longer than that of human-Trichuris, and 289 bp shorter than that of pig-Trichuris. The arrangement of the mt genes (i.e., 13 protein genes, 2 rrn genes and 22 tRNA genes) and NCRs were the same. A pairwise comparison of the nucleotide sequences of each mt gene and the amino acid sequences conceptually translated from individual protein genes was made among the three taxa of Trichuris (from the three host species) (Table 3). The sequence lengths of individual genes varied among these taxa, except for the nad1 gene, which was the same (Table 3). The magnitude of sequence variation in each gene among the three taxa of Trichuris ranged from 24.2–50.9% for nucleotide sequences and 13.6–62.5% for amino acid sequences (Table 3). The sequence difference across the entire mt genome between monkey- and human-Trichuris was 29.35% (a total of 4,152 nucleotide alterations). This difference across the entire mt genome between monkey- and pig-Trichuris was 33.49% (a total of 4835 nucleotide alterations). The greatest variation among the three taxa of Trichuris was in the atp8 gene (42.4–58.9%), whereas least differences (24.3%–31.5%) were detected in the rrnS and rrnL subunits, respectively (Table 3).
Table 3. Nucleotide and/or predicted amino acid (aa) sequence differences for mt protein-coding and ribosomal RNA genes among monkey-Trichuris (MT), human-Trichuris (Trichuris trichiura = TT) and pig-Trichuris (T. suis = TS).
| Gene/region | Nucleotide length (bp) | Nucleotide difference (%) | Number of aa | aa difference (%) | ||||||||
| MT | TT | TS | MT/TT | MT/TS | TT/TS | MT | TT | TS | MT/TT | MT/TS | TT/TS | |
| atp6 | 822 | 828 | 828 | 36.8 | 40.2 | 39.6 | 273 | 275 | 275 | 40.0 | 49.5 | 49.5 |
| nad1 | 900 | 900 | 900 | 29.6 | 32.8 | 33.1 | 299 | 299 | 299 | 28.1 | 32.4 | 33.1 |
| nad2 | 900 | 885 | 882 | 34.7 | 39.2 | 34.1 | 299 | 294 | 293 | 31.8 | 44.2 | 40.8 |
| nad3 | 339 | 342 | 342 | 32.7 | 38.6 | 33.6 | 112 | 113 | 113 | 27.4 | 35.4 | 31.9 |
| nad4 | 1212 | 1212 | 1209 | 31.3 | 41.2 | 40.9 | 403 | 403 | 402 | 31.3 | 55.1 | 58.3 |
| nad4L | 261 | 258 | 252 | 28.7 | 35.6 | 38.0 | 86 | 85 | 83 | 36.0 | 46.5 | 42.4 |
| nad5 | 1548 | 1548 | 1557 | 34.6 | 38.6 | 35.9 | 515 | 515 | 518 | 37.5 | 47.1 | 42.5 |
| nad6 | 468 | 477 | 471 | 32.3 | 35.7 | 33.1 | 155 | 158 | 156 | 35.4 | 45.6 | 38.6 |
| cox1 | 1536 | 1545 | 1542 | 24.2 | 25.0 | 25.4 | 511 | 514 | 513 | 13.6 | 16.6 | 13.6 |
| cox2 | 675 | 675 | 681 | 24.3 | 31.6 | 30.7 | 224 | 224 | 226 | 18.7 | 31.0 | 28.8 |
| cox3 | 774 | 774 | 777 | 29.2 | 31.0 | 35.5 | 257 | 257 | 258 | 25.7 | 30.6 | 34.5 |
| cytb | 1107 | 1107 | 1113 | 26.1 | 29.0 | 27.7 | 368 | 368 | 370 | 20.9 | 27.3 | 26.2 |
| atp8 | 165 | 165 | 171 | 42.4 | 50.9 | 47.4 | 54 | 54 | 56 | 55.6 | 58.9 | 62.5 |
| rrnS | 705 | 698 | 712 | 25.4 | 24.3 | 24.6 | – | – | – | – | – | – |
| rrnL | 1007 | 1011 | 1011 | 25.1 | 31.5 | 25.1 | – | – | – | – | – | – |
Amino acid sequences inferred from individual mt protein genes of monkey-Trichuris were compared with those of human- and pig-Trichuris. The difference across amino acid sequences of the 13 protein genes between the monkey- and human-Trichuris was 28.52% (a total of 1015 amino acid alterations) and 38.28% (a total of 1364 amino acid alterations) between the monkey- and pig-Trichuris, respectively. The amino acid sequence differences among three taxa of Trichuris ranged from 13.6–62.5%, with COX1 being the most conserved and ATP8 the least conserved protein. Phylogenetic analyses of concatenated amino acid sequence data sets, using T. spiralis as an outgroup, revealed that the monkey-Trichuris was more closely related to the human-Trichuris than to representative Trichuris species from porcine and ruminant hosts, with absolute support (pp = 1.00) (Figure 2).
Figure 2. Inferred phylogenetic relationship of Trichuris from the François’ langur (Trichuris sp.) with T. trichiura , T. suis, T. ovis and T. discolor.
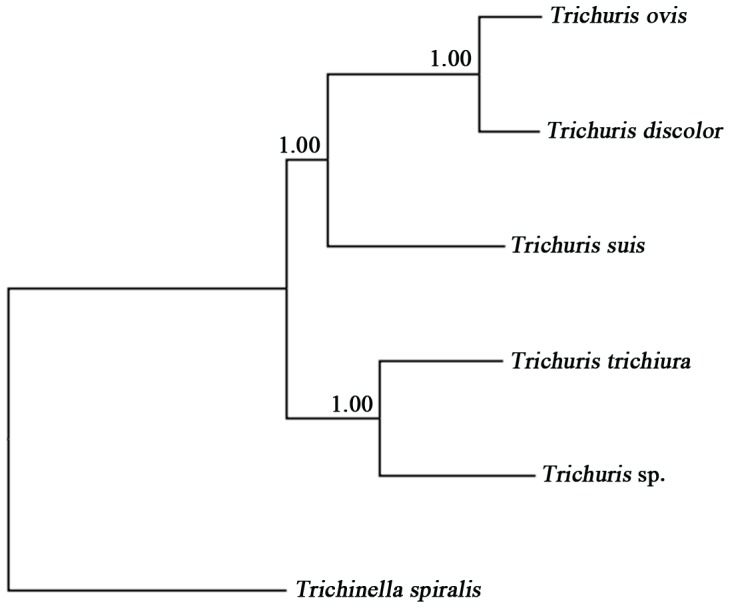
Analysis of the concatenated amino acid sequence data representing 12 protein-coding genes (with the exception of atp8) by Bayesian inference (BI), using Trichinella spiralis (NC_002681) as the outgroup.
Comparison of the mt genomes of monkey-Trichuris, human-Trichuris and pig-Trichuris showed that the rrnS and rrnL were the two most conserved genes (Table 3). Sequence variation in part of the rrnL gene was assessed among four individuals of Trichuris from monkeys. The rrnL sequences of the four monkey-Trichuris individuals (GenBank accession nos. KC481232-KC481235) were of the same length (616 bp). Nucleotide variation among the four monkey-Trichuris individuals was detected at 15 sites (15/616; 2.44%). The four monkey-Trichuris sequeces were aligned with 10 and six rrnL sequences (GenBank accession nos. AM993017-AM993032; [22]) reported previously for human- and pig- derived Trichuris, respectively. The alignment of the partial rrnL sequences revealed that all individuals of monkey-Trichuris differed at 140 nucleotide positions (140/430; 32.6%) when compared with human- and pig-Trichuris. Phylogenetic analysis of the rrnL sequence data from individual worms revealed strong support for the separation of monkey-Trichuris from human-Trichuris and pig-Trichuris (Figure 3).
Figure 3. Inferred genetic relationships of four individual specimens of from the François’ langur (Trichuris sp.) with those of Trichuris trichiura (n = 6) and T. suis (n = 10) from China.

The analyses of mitochondrial rrnL sequence data were carried out by Bayesian inference (BI), using Trichinella spiralis as the outgroup. Posterior probabilities (pp) values of <0.9 are not shown.
Sequence variation was examined for both ITS-1 and ITS-2 of monkey-Trichuris. The ITS-1 and ITS-2 sequences from four individual adults of monkey-Trichuris were compared with those of human- and pig-derived Trichuris [25]. Sequence variations were 0–0.7% (ITS-1) and 0–0.9% (ITS-2) among the three specimens of monkey-Trichuris. However, the sequence differences were 21.4–22.3% (ITS-1) and 22.4–23.7% (ITS-2) between the monkey- and human-Trichuris, and 56.5–57.0% (ITS-1) and 43.6–45.5% (ITS-2) between the monkey- and pig-Trichuris.
Discussion
To date, more than 20 Trichuris species have been described from various mammalian hosts based on the microscopic features of the adult worms [42]. Some studies (e.g., [43], [44]) have claimed that male spicule and body lengths are useful morphological parameters for the differentiation of Trichuris species. However, other studies have shown that these measurements are not necessarily reliable for specific identification [15]. For instance, Cutillas et al. (2009) [15] observed that the spicule lengths of T. trichiura and T. suis overlapped. While other workers considered that the presence of pericloacal papillae might be useful for species determination [45], also this criterion does not appear to allow accurate identification/delineation [15]. Clearly, these studies show that morphological characters or morphometrics should be interpreted with caution. For this reason, we employed here a molecular genetic approach, logically extending previous studies [22]–[26], so that comparative genetic analyses could be conducted.
The present investigation shows clear genetic distinctiveness between Trichuris from the François’ langur and Trichuris from humans and livestock animals (i.e., T. suis, T. ovis and T. discolor) (Figure 2). Our and previous findings [22]–[27] support the contention that each Trichuris species has a very specific affiliation with a particular host species [16], although, to date, only small numbers of adult worms have been studied molecularly. Clearly, larger population genetic and molecular epidemiological studies should be conducted using the mt and nuclear markers defined in this and previous studies [22]–[28] to further test this hypothesis.
The sequence difference in the inferred mt proteome between monkey- and human-Trichuris was 29.4%, and sequence variation among individual worms from each host species was low (0–2.4%), suggesting that these parasites are separate species. This proposal was further supported by phylogenetic analysis (cf. Figure 3). Previous studies [7]–[13] have indicated that many non-human primates, such as Colobus guereza, Macaca fascicularis, M. silenus, Papio anubis, P. hamadryas ursinus and Theropithecus gelada, can harbour Trichuris. However, the specific identity, host specificity and zoonotic potential of each operational taxonomic unit (OTU) [13] of Trichuris from each of these host species are unknown. Based on molecular findings to date for Trichuris from other animal species [22]–[28], [46]–[51], we anticipate that each primate species harbours its own species of Trichuris, but, clearly, this proposal requires rigorous testing.
Trachypithecus françoisi is a threatened/endangered species of primate nearing extinction [52]. Populations of this langur have been on the decline for the past 30 years. For instance, the populations in Guangxi province, China, crashed from 4000–5000 individuals in 1980 to a mere 307 in 2002–2003 [53]. The main factors linked to this decline have been hunting, habitat destruction and harvesting of langur organs for the preparation of traditional medicines [54], [55]. Another likely threat to the François’ langur, particularly in captive situations in conservation parks and zoos, is whipworm disease. This statement is supported by reports from China (e.g., [56]–[59]), indicating that Trichuris infection is common (14.3–100%) in this langur in zoos and conservation parks, as are clinical cases of trichuriasis. The direct life cycle of Trichuris, the accumulation of eggs in environments with relatively high population density of primates (animals) and the robustness and longevity of the infective stage (larvated eggs) in the environment [60] are all factors that contribute significantly to a gradual increase of trichuriasis in ‘closed’ environments, such as parks [61]. Although we expect the monkey-Trichuris studied herein to be specific to the François’ langur, there is a possibility that this parasite is transmissible to other primates, including humans. However, this proposal needs to be assessed.
In spite of molecular evidence for the existence of a unique Trichuris species in the endangered François’ langur, the interpretations from the present study are guarded, at this stage, until detailed population genetic investigations have been conducted. Future studies should include (i) exploring, in detail, nucleotide variation in rDNA and mtDNA within and among Trichuris populations from a range of different primate species and countries, and to establish whether more than one Trichuris species infect non-human primates, (ii) establishing, using accurate molecular tools, whether cross-host species infection occurs or not, (iii) undertaking detailed morphological studies, by scanning electron microscopy and field emission scanning electron microscopy, of whipworms from various non-humans primates. This focus is important because, traditionally, the diagnosis of Trichuris infection in animals has relied mainly on the morphological identification of adult and egg stages.
Funding Statement
This work was supported by the International Science & Technology Cooperation Program of China (Grant No. 2013DFA31840) to XQZ and RBG. This work was also supported by the Science Fund for Creative Research Groups of Gansu Province (Grant No. 1210RJIA006) to XQZ. RBG’s research is supported by the Australian Research Council (ARC), National Health and Medical Research Council (NHMRC) and Melbourne Water Corporation (MWC); the Alexander von Humboldt Foundation is also gratefully acknowledged. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Hotez PJ, Fenwick A, Savioli L, Molyneux DH (2009) Rescuing the bottom billion through control of neglected tropical diseases. Lancet 373: 1570–1575. [DOI] [PubMed] [Google Scholar]
- 2. Bethony J, Brooker S, Albonico M, Geiger SM, Loukas A, et al. (2006) Soil-transmitted helminth infections: ascariasis, trichuriasis, and hookworm. Lancet 367: 1521–1532. [DOI] [PubMed] [Google Scholar]
- 3. Roepstorff A, Mejer H, Nejsum P, Thamsborg SM (2011) Helminth parasites in pigs: new challenges in pig production and current research highlights. Vet Parasitol 180: 72–81. [DOI] [PubMed] [Google Scholar]
- 4. Traversa D (2011) Are we paying too much attention to cardio-pulmonary nematodes and neglecting old-fashioned worms like Trichuris vulpis? Parasit Vectors 4: 32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Khalafalla RE, Elseify MA, Elbahy NM (2011) Seasonal prevalence of gastrointestinal nematode parasites of sheep in Northern region of Nile Delta, Egypt. Parasitol Res 108: 337–340. [DOI] [PubMed] [Google Scholar]
- 6. Cliffe LJ, Grencis RK (2004) The Trichuris muris system: a paradigm of resistance and susceptibility to intestinal nematode infection. Adv Parasitol 57: 255–307. [DOI] [PubMed] [Google Scholar]
- 7. Levecke B, Dorny P, Geurden T, Vercammen F, Vercruysse J (2007) Gastrointestinal protozoa in non-human primates of four zoological gardens in Belgium. Vet Parasitol 148: 236–246. [DOI] [PubMed] [Google Scholar]
- 8. Lin K, Li Z, Li L, Zhou J, Zhang R, et al. (2011) Investigation on the intestinal parasites infection in conventional (CV) macaques. Fujian Anim Husbandry Vet Med 6: 1–4 (in Chinese).. [Google Scholar]
- 9. Legesse M, Erko B (2004) Zoonotic intestinal parasites in Papio anubis (baboon) and Cercopithecus aethiops (vervet) from four localities in Ethiopia. Acta Trop. 90: 231–236. [DOI] [PubMed] [Google Scholar]
- 10. Muriuki SM, Murugu RK, Munene E, Karere GM, Chai DC (1998) Some gastro-intestinal parasites of zoonotic (public health) importance commonly observed in old world non-human primates in Kenya. Acta Trop 71: 73–82. [DOI] [PubMed] [Google Scholar]
- 11. Munene E, Otsyula M, Mbaabu DA, Mutahi WT, Muriuki SM, et al. (1998) Helminth and protozoan gastrointestinal tract parasites in captive and wild-trapped African non-human primates. Vet Parasitol 78: 195–201. [DOI] [PubMed] [Google Scholar]
- 12. Michaud C, Tantalean M, Ique C, Montoya E, Gozalo A (2003) A survey for helminth parasites in feral New World non-human primate populations and its comparison with parasitological data from man in the region. J Med Primatol 32: 341–345. [DOI] [PubMed] [Google Scholar]
- 13. Ravasi DF, O’Riain MJ, Adams VJ, Appleton CC (2012) A coprological survey of the protozoan and nematode parasites of free-ranging chacma baboons (Papio ursinus) in the south-western Cape, South Africa. S Afr J Wildl Res 42: 35–44. [Google Scholar]
- 14. Ooi HK, Tenora F, Itoh K, Kamiya M (1993) Comparative study of Trichuris trichiura from non-human primates and from man, and their difference with T. suis. J Vet Med Sci. 55: 363–366. [DOI] [PubMed] [Google Scholar]
- 15. Cutillas C, Callejon R, Rojas MD, Tewes B, Ubeda JM (2009) Trichuris suis and Trichuris trichiura are different nematode species. Acta Trop 111: 299–307. [DOI] [PubMed] [Google Scholar]
- 16. Beer RJ (1976) The relationship between Trichuris trichiura (Linnaeus 1758) of man and Trichuris suis (Schrank 1788) of the pig. Res Vet Sci 20: 47–54. [PubMed] [Google Scholar]
- 17. Blouin MS (2002) Molecular prospecting for cryptic species of nematodes: mitochondrial DNA versus internal transcribed spacer. Int J Parasitol 32: 527–531. [DOI] [PubMed] [Google Scholar]
- 18. Gasser RB (2006) Molecular tools–advances, opportunities and prospects. Vet Parasitol 136: 69–89. [DOI] [PubMed] [Google Scholar]
- 19. Gasser RB, Bott NJ, Chilton NB, Hunt P, Beveridge I (2008) Toward practical, DNA-based diagnostic methods for parasitic nematodes of livestock–bionomic and biotechnological implications. Biotechnol Adv 26: 325–334. [DOI] [PubMed] [Google Scholar]
- 20. Jex AR, Waeschenbach A, Hu M, van Wyk JA, Beveridge I, et al. (2009) The mitochondrial genomes of Ancylostoma caninum and Bunostomum phlebotomum–two hookworms of animal health and zoonotic importance. BMC Genomics 10: 79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Jex AR, Littlewood DT, Gasser RB (2010) Toward next-generation sequencing of mitochondrial genomes–focus on parasitic worms of animals and biotechnological implications. Biotechnol Adv 28: 151–159. [DOI] [PubMed] [Google Scholar]
- 22. Liu GH, Gasser RB, Su A, Nejsum P, Peng L, et al. (2012) Clear genetic distinctiveness between human- and pig-derived Trichuris based on analyses of mitochondrial datasets. PLoS Negl Trop Dis 6: e1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Liu GH, Wang Y, Xu MJ, Zhou DH, Ye YG, et al. (2012) Characterization of the complete mitochondrial genomes of two whipworms Trichuris ovis and Trichuris discolor (Nematoda: Trichuridae). Infect Genet Evol 12: 1635–1641. [DOI] [PubMed] [Google Scholar]
- 24. Nissen S, Al-Jubury A, Hansen TV, Olsen A, Christensen H, et al. (2012) Genetic analysis of Trichuris suis and Trichuris trichiura recovered from humans and pigs in a sympatric setting in Uganda. Vet Parasitol 188: 68–77. [DOI] [PubMed] [Google Scholar]
- 25.Liu GH, Zhou W, Nisbet AJ, Xu MJ, Zhou DH, et al. (2013) Characterization of Trichuris trichiura from humans and T. suis from pigs in China using internal transcribed spacers of nuclear ribosomal DNA. J Helminthol. In press. [DOI] [PubMed]
- 26. Cutillas C, Oliveros R, de Rojas M, Guevara DC (2002) Determination of Trichuris muris from murid hosts and T. arvicolae (Nematoda) from arvicolid rodents by amplification and sequentiation of the ITS1–5.8S-ITS2 segment of the ribosomal DNA. Parasitol Res 88: 574–582. [DOI] [PubMed] [Google Scholar]
- 27. Cutillas C, de Rojas M, Ariza C, Ubeda JM, Guevara D (2007) Molecular identification of Trichuris vulpis and Trichuris suis isolated from different hosts. Parasitol Res 100: 383–389. [DOI] [PubMed] [Google Scholar]
- 28. Ravasi DF, O’Riain MJ, Davids F, Illing N (2012) Phylogenetic evidence that two distinct Trichuris genotypes infect both humans and non-human primates. PLoS One. 7: e44187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Hu G, Wei Y (2002) Population decline and habitats destruction of Francois’ langur, Trachypithecus francoisi in Fusui Nature Reserve, southwest Guangxi, China. In: Abstr 19th Congr Int Primatol Soc. Beijing, China, 74–75.
- 30. Gasser RB, Hu M, Chilton NB, Campbell BE, Jex AJ, et al. (2006) Single-strand conformation polymorphism (SSCP) for the analysis of genetic variation. Nat Protoc 1: 3121–3128. [DOI] [PubMed] [Google Scholar]
- 31. Gasser RB, Zhu XQ, McManus DP (1999) NADH dehydrogenase subunit 1 and cytochrome c oxidase subunit I sequences compared for members of the genus Taenia (Cestoda). Int J Parasitol 29: 1965–1970. [DOI] [PubMed] [Google Scholar]
- 32. Hu M, Jex AR, Campbell BE, Gasser RB (2007) Long PCR amplification of the entire mitochondrial genome from individual helminths for direct sequencing. Nature Protoc 2: 2339–2344. [DOI] [PubMed] [Google Scholar]
- 33. Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 24: 4876–4882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Lowe TM, Eddy SR (1997) tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res 25: 955–964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Hu M, Chilton NB, Gasser RB (2002) The mitochondrial genomes of the human hookworms, Ancylostoma duodenale and Necator americanus (Nematoda: Secernentea). Int J Parasitol 32: 145–158. [DOI] [PubMed] [Google Scholar]
- 36. Zhu X, Chilton NB, Jacobs DE, Boes J, Gasser RB (1999) Characterisation of Ascaris from human and pig hosts by nuclear ribosomal DNA sequences. Int J Parasitol 29: 469–478. [DOI] [PubMed] [Google Scholar]
- 37. Lavrov DV, Brown WM (2001) Trichinella spiralis mtDNA: A nematode mitochondrial genome that encodes a putative atp8 and normally structured tRNAs and has a gene arrangement relatable to those of coelomate metazoans. Genetics 157: 621–637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Talavera G, Castresana J (2007) Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst Biol 56: 564–577. [DOI] [PubMed] [Google Scholar]
- 39. Blaxter ML, De Ley P, Garey JR, Liu LX, Scheldeman P, et al. (1998) A molecular evolutionary framework for the phylum Nematoda. Nature 392: 71–75. [DOI] [PubMed] [Google Scholar]
- 40. Liu GH, Wang SY, Huang WY, Zhao GH, Wei SJ, et al. (2012) The complete mitochondrial genome of Galba pervia (Gastropoda: Mollusca), an intermediate host snail of Fasciola spp. PLoS One 7: e42172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Page RD (1996) TREEVIEW: an application to display phylogenetic trees on personal computers. Comput Appl Biosci 12: 357–358. [DOI] [PubMed] [Google Scholar]
- 42. Robles MR (2011) New species of Trichuris (Nematoda: Trichuridae) from Akodon montensis, Thomas, 1913, of the Paranaense Forest in Argentina. J Parasitol 97: 319–327. [DOI] [PubMed] [Google Scholar]
- 43. Chandler AC (1930) Specific characters in the genus Trichuris with a description of the new species Trichuris tenuis from a camel. J Parasitol 16: 198–206. [Google Scholar]
- 44. Knight RA (1971) Redescription of Trichuris discolor (von Linstow, 1906) and T. skrjabini (Baskakov, 1924) from domestic ruminants in the United States and comparisons with T. ovis (Abildgaard, 1795). J Parasitol 57: 302–310. [PubMed] [Google Scholar]
- 45. Spakulová (1994) Discriminant analysis as a method for the numerical evaluation of taxonomic characters in male trichurid nematode. Syst Parasitol 29: 113–119. [Google Scholar]
- 46. Cutillas C, Oliveros R, de Rojas M, Guevara DC (2004) Determination of Trichuris skrjabini by sequencing of the ITS1–5.8S-ITS2 segment of the ribosomal DNA: comparative molecular study of different species of trichurids. J Parasitol 90: 648–652. [DOI] [PubMed] [Google Scholar]
- 47. Callejón R, de Rojas M, Ariza C, Ubeda JM, Guevara DC, et al. (2009) Cytochrome oxidase subunit 1 and mitochondrial 16S rDNA sequences of Trichuris skrjabini (Tricocephalida: Trichuridae). Parasitol Res 104: 715–716. [DOI] [PubMed] [Google Scholar]
- 48. Callejón R, de Rojas M, Nieberding C, Foronda P, Feliú C, et al. (2010) Molecular evolution of Trichuris muris isolated from different Muridae hosts in Europe. Parasitol Res 107: 631–641. [DOI] [PubMed] [Google Scholar]
- 49. Callejón R, Halajian A, de Rojas M, Marrugal A, Guevara D, et al. (2012) 16S partial gene mitochondrial DNA and internal transcribed spacers ribosomal DNA as differential markers of Trichuris discolor populations. Vet Parasitol. 186: 350–363. [DOI] [PubMed] [Google Scholar]
- 50. Oliveros R, Cutillas C, De Rojas M, Arias P (2000) Characterization of four species of Trichuris (Nematoda: Enoplida) by their second internal transcribed spacer ribosomal DNA sequence. Parasitol Res 86: 1008–1013. [DOI] [PubMed] [Google Scholar]
- 51.Salaba O, Rylková K, Vadlejch J, Petrtýl M, Scháňková S, et al. (2013) The first determination of Trichuris sp. from roe deer by amplification and sequenation of the ITS1–5.8S-ITS2 segment of ribosomal DNA. Parasitol Res. In press. [DOI] [PubMed]
- 52. Duckworth JW, Boonratana R, Robichaud WG, Timmins RJ (2010) A review of François’ leaf monkey Trachypithecus francoisi (sensu lato) in Lao PDR. Primate Conserv (25): 1–19. [Google Scholar]
- 53. Li Y, Huang C, Ding P, Tang Z, Wood C (2007) Dramatic decline of Francois’ langur Trachypithecus francoisi in Guangxi Province, China. Oryx 41: 38–43. [Google Scholar]
- 54. Li HQ, Lian ZM (2007) A survey of resources and protection of Francois’ Langur (Trachypithecus francois francois) in China. J Yanan Univ (Natural Science Edition) 26: 60–67 (in Chinese).. [Google Scholar]
- 55. Ma Q, Su HL (2004) Francois’s Leaf Monkey (Trachypithecus francoisi). Chinese J Zool 39: 53–54 (in Chinese).. [Google Scholar]
- 56. Hu HG, Zhao GL, Huang H, Wu J (1993) The control of parasites in wild animals in Chongqing zoological garden. J Sichuan Teach Coll (Natural Science) 14: 62–69 (in Chinese).. [Google Scholar]
- 57. Qi M, Dong HJ, Hu L, Peng GN, Chen ZQ, et al. (2011) Investigation on the prevalence of intestinal parasites in wild animal zoo. China Anim Husbandry Vet Med 38: 188–191 (in Chinese).. [Google Scholar]
- 58. Wei LY, Jiang CF, Han JJ, Yang DJ, LIAO XF (1990) Investigation of gastrointestinal parasites of primate animals in Wuhan zoo, Hubei province. Chinese J Zool 25: 3–6 (in Chinese).. [Google Scholar]
- 59. Yang MH, Gong GJ (1998) Analyses and fecal example of parasites in Beijing zoo. Chinese J Vet Med 24: 19–20 (in Chinese).. [Google Scholar]
- 60. Stephenson LS, Holland CV, Cooper ES (2000) The public health significance of Trichuris trichiura . Parasitology 121: S73–95. [DOI] [PubMed] [Google Scholar]
- 61. Dubin S, Segall S, Martindale J (1975) Contamination of soil in two city parks with canine nematode ova including Toxocara canis: a preliminary study. Am J Public Health 65: 1242–1245. [DOI] [PMC free article] [PubMed] [Google Scholar]