

Abstract
Recent research has demonstrated that a population of stem cells can be isolated from amniotic fluid removed by amniocentesis that are broadly multipotent and nontumorogenic. These amniotic fluid-derived stem cells (AFSC) could potentially provide an autologous cell source for treatment of congenital defects identified during gestation, particularly cardiovascular defects. In this review, the various methods of isolating, sorting, and culturing AFSC are compared, along with techniques for inducing differentiation into cardiac myocytes and endothelial cells. Although research has not demonstrated complete and high-yield cardiac differentiation, AFSC have been shown to effectively differentiate into endothelial cells and can effectively support cardiac tissue. Additionally, several tissue engineering and regenerative therapeutic approaches for the use of these cells in heart patches, injection after myocardial infarction, heart valves, vascularized scaffolds, and blood vessels are summarized. These applications show great promise in the treatment of congenital cardiovascular defects, and further studies of isolation, culture, and differentiation of AFSC will help to develop their use for tissue engineering, regenerative medicine, and cardiovascular therapies.
Introduction
Birth defects are the number one cause of death in infants in the United States.1 Approximately 9200 invasive procedures were performed in infants with congenital heart disease in the United States in 2010.2 Many of these repairs require synthetic or acellular materials. Current myocardial patched repair, common for repair of septal defects and outflow tract obstructions, and conduit repair, or construction of a new conduit as in a Blalock-Taussig Shunt, generally uses Dacron, Teflon, or fixed acellular pericardium.3 However, these patches have been shown to increase the risk of sudden cardiac death by 25 to 100 times compared to a healthy population.4 Additionally, valve repair in infants can often make use of autologous valves (as in a Ross procedure, which uses the patient's pulmonary valve to replace a damaged aortic valve), but many times a mechanical valve is required as xenotropic (animal) valves are seldom used in children due to concerns of long-term durability.5 These mechanical valves do not grow with the patient and require a lifetime of anticoagulation therapy with many side effects.6 Tissue-engineered or regenerative therapies to treat these pathologies, which result in living tissue that grows with the child, could greatly benefit this population.
Many advances in biomaterials and decellularized tissues in the last decade have aimed to generate scaffolds for tissue engineering repair of congenital heart defects (see review in Ref.7). However, very few of these have used living cells. The use of live and functioning cardiac cells in tissue-engineered cardiac scaffolds could allow for correction of large, full-thickness defects, maintenance of the normal conduction pathways in the heart, the creation of highly patent valves that grow with pediatric patients, and many other transformative technologies that have been thoroughly reviewed elsewhere.7–9 Although some therapies have seeded constructs with bone marrow mesenchymal stem cells (MSC),10 these can be very difficult and risky to obtain autologously in neonates. Additionally, concern about tumorogenesis has limited use of human embryonic and induced pluripotent stem cells. Because of the risks of immunorejection and the high risks of immunosuppressive drugs in infants, nonautologous transplanted cells are not a desirable option.11 In this article, we will discuss the characterization and use of stem cells isolated from amniotic fluid, which can be obtained with relatively low risk when birth defects are diagnosed in utero and used to construct tissue-engineered and regenerative therapeutic tissues, using the fetus' own cells, for the correction of these defects in the newborn.
Amniotic Fluid
Amniotic fluid (AF) fills the amniotic cavity and acts to prevent adherence of the embryo to the amnion, absorb jolts to protect the embryo or fetus, and allow for fetal movement.12 Amniotic membrane cells produce some of the fluid, but the bulk of the liquid component comes from the maternal blood. Approximately 1 L of AF surrounds the fetus by birth. Beginning in the fifth month the fetus will consume an estimated 400 mL of AF a day, and fetal urine is then added back to the fluid. The fetal urine is mostly water, as metabolic waste is exchanged through the placenta.12
For research applications the most common method to obtain AF is through amniocentesis.13–17 Alternatively, fluid can be obtained from therapeutic amnioreductions in which large quantities of AF are removed for treatment of conditions such as twin–twin transfusion syndrome18 or AF may be collected during elective caesarean sections.19
Identification, Isolation, and Culture of Undifferentiated Stem Cells in AF
The cells contained in AF are a heterogeneous mixture of many cell types derived from and genetically matched to the fetus.15,20 The predominant phenotype observed is epithelial cells, but cells expressing mesenchymal, osteocyte, adipocyte, and neuronal markers are also present.20
AF was first found to contain Oct-4-positive cells in 2003.16 Oct-4 is a transcription factor necessary for embryonic stem cell (ESC) pluripotency.21 In 2007, AF was determined to contain a population of broadly multipotent stem cells.15 Adherent cells from amniocentesis were immunoselected for c-kit expression using magnetic microsphere-assisted cell sorting. C-kit, or CD117, is a tyrosine-kinase transmembrane receptor for stem cell factor, also known as mast cell growth factor, which is a pleiotropic factor involved in development and known to be required in hematopoiesis in adults.22 Clonal colonies were generated from adherent, c-kit-sorted cells and tracked by retroviral marking. It was found that some clonal colonies were able to differentiate toward adipogenic, osteogenic, myogenic, endothelial, neuronal, and hepatic lineages, representing cell types of all three germ layers.15 Even though AF-derived stem cells (AFSC) are broadly multipotent, they did not form teratomas when implanted in severe combined immunodeficiency mice.15 This nontumor forming property, which is a similarity AFSC share with adult stem cells, could be an advantage over human ESCs when medical applications are considered.
Despite this early promise with stem cells from AF, much work remains before their potential can be realized. Many differences still exist in the methods research labs use for culturing and isolating AFSC. While many researchers appear to use the same method as the Atala lab described above, variation does exist. A summary of isolation and culture conditions for maintaining undifferentiated AFSC currently published can be found in Table 1 (culture conditions used for AFSC tested for cardiovascular tissue engineering marked with *). In addition to the culture methods for maintaining undifferentiated stem cells, researchers are examining culture conditions to promote particular cell types, such as advanced Dulbecco's minimal essential medium with 5% fetal bovine serum resulting in the enrichment of epithelial cell population.14
Table 1.
Isolation Methods and Culture Medium for Undifferentiated Amniotic Fluid-Derived Stem Cell
| Sorting | Medium | References | |
|---|---|---|---|
| – | αMEM, 10% FBS | 14,38,40 | |
| c-kit | αMEM, 10% FBS | 14 | |
| * | – | αMEM, 15% FBS, 20% Chang (Irvine Scientific) | 23,52 |
| * | c-kit | αMEM, 15% FBS, 20% Chang (Irvine Scientific) | 15,18,32–34,36,37,45–48,58,65 |
| * | – | αMEM, 20% FBS, 4 ng/mL bFGF | 17,49,50,54 |
| – | M199 medium, 10% FBS | 20,51 | |
| – | F-10 nutrient medium, 10% FBS | 16,41,43 | |
| – | F-10 nutrient medium, 20% FBS | 44 | |
| – | DMEM, 20% FBS | 20,39 | |
| – | DMEM, 10% FBS, 10 ng/mL EGF | 29,99 | |
| – | DMEM, 10% FBS, 10 ng/mL bFGF | 30 | |
| – | DMEM, 20% FBS, 5–10 ng/mL bFGF | 26–28,31,97 | |
| – | DMEM, 20% FBS, 10 ng/mL bFGF, 10 ng/mL EGF | 19,24 | |
| * | c-kit | AmnioMax Medium (Invitrogen), 10% FBS | 35,51 |
| – | Mesencult medium (Stem Cell Technologies, Inc.) | 25 | |
| – | Amniomed (Euroclone) | 20 |
Culture conditions tested for cardiovascular tissue engineering.
AFSC, amniotic fluid-derived stem cells; αMEM, alpha modified Eagle's medium; FBS, fetal bovine serum; DMEM, Dulbecco's modified Eagle's medium; EGF, epidermal growth factor; bFGF, basic fibroblast growth factor.
AFSC are the subject of ongoing interest and research. Isolation, culture, and cryopreservation of AFSC are ongoing areas of investigation in the field.19,23–28 Additionally, some researchers are comparing AFSC to other fetal-derived tissues such as umbilical cord blood and amniotic membrane.27,29,30 AFSC are being explored for use in tissue engineering applications for a variety of tissue types. This review focuses on cardiovascular cell types and tissue engineering, which are addressed further below, but AFSC are also being investigated for osteogenic,13,31,32 renal,33,34 myogenic,35 epithelial,36,37 and neuronal14,38–45 differentiation and tissue engineering.
An area of contention that remains with AFSC is whether sorting for c-kit expression is necessary or appropriate. Approximately 1% of total AF cells are positive for c-kit.15,34 One research group studying neuronal differentiation investigated the ability of c-kit sorted cells compared to unsorted cells to differentiate toward a neuronal lineage.14 It was found that the unsorted population was better able to differentiate toward a neuronal lineage than the sorted population. However, the c-kit-sorted population was better able to differentiate toward adipogenic, osteogenic, and chondrogenic lineages.14 These findings, in combination with the lack of agreement in the field on isolation and culture procedures, indicate further research is needed to determine and validate the appropriate isolation procedures for cells from AF capable of differentiation into cardiovascular cell types. Furthermore, it may be necessary to distinguish distinct populations of AF cells that may be better suited for various therapeutic end points.
Analysis of the undifferentiated AFSC populations is necessary to understand the starting populations of cells in differentiation experiments. Furthermore, the different culture conditions used for the undifferentiated stem cells necessitates analysis of the undifferentiated stem cells to determine if the starting populations across experiments are similar and comparable. A summary of markers analyzed on undifferentiated AFSC can be found in Table 2. AFSC were found to be positive for markers common of MSCs: CD29 or integrin beta-1,17–19,24,25,30,40,41,46–51 CD44 or hyaluronate receptor,17,18,20,23,25,28–30,40,41,47–52 CD73 or ecto-5’-nucleotidase,17–20,23–25,28,40,46–50 CD90 or Thy-1,17–19,23,24,28,40,46–52 CD105 or endoglin,17–19,23–25,28,30,40,46–52 and CD146 or melanoma cell adhesion molecule.25,47,48 The stem cells were also found to be negative for endothelial markers CD31,18,19,24–26,47,49,50,53 CD144,25 and CD30925 and for hematopoietic markers CD11b,17 CD14,17,25,40,51 CD34,17,25,28,30,41,46–51 and CD45.18,19,23–25,28,40,41,46–52 AFSC were positive for vimentin,23,26,35,40,46,47,52–54 an intermediate filament commonly found in mesenchymal cells, and negative for desmin,26,53 an intermediate filament found in the Z-line of sarcomeres in muscle cells. AFSC were positive for major histocompatability complex (MHC) I,17,18,23,25,28,30,41,47,50 also known as HLA-ABC, and negative for MHC II,17,18,23,30,41,47,50,52 also known as HLA-DR. Stage-specific embryonic antigen (SSEA)-4, a surface antigen commonly found on ESCs, was positive on AFSC by flow cytometry and immunostaining.18,23,30,35,46,49–52,54 Results for expression of c-kit or CD117 in AFSC were mixed: some research demonstrated positive gene expression by PCR,37 but flow cytometry results were negative17,49,50 or slightly (∼25%) positive.18 Oct4, Sox2, and Nanog, transcription factors necessary for maintaining pluirpotency in ESCs, also displayed mixed results across different studies of AFSC.16,17,20,23,24,29–31,35,37,39–41,46,48–52,54
Table 2.
Characterization of Undifferentiated Amniotic Fluid-Derived Stem Cell
| Marker | Description | Results | References |
|---|---|---|---|
| Oct-4 | Transcription factor necessary for maintaining pluripotency of ESC | Positive expression, generally at low levels, by PCR, FC, and immunostaining. Found by some to be only expressed in some fluid samples. | 16,17,20,23,24,29–31,35,37,39–41,46,49–52,54 |
| Sox2 | Transcription factor necessary for maintaining pluripotency of ESC | Mixed results: both positive and negative expression by PCR reported, as well as positive immunostaining | 20,39,48 |
| Nanog | Transcription factor necessary for maintaining pluripotency of ESC | Positive expression at low levels by PCR, FC, and immunostaining | 39,40,49,50,54 |
| SSEA-4 | Stage-specific embryonic antigen 4; surface antigen found on ESC | Positive expression by FC and immunostaining | 18,23,30,35,46,49–52,54 |
| MHC I | Found on every nucleated cell in the body | Positive expression by PCR and FC | 17,18,23,25,28,30,41,47,50 |
| MHC II | Found only on antigen presenting cells | Negative by PCR and FC | 17,18,23,30,41,47,50,52 |
| CD29, CD44, CD73, CD90, CD105, CD146 | Markers common on MSC | Positive expression by PCR, FC, and immunostaining | 17–20,23–25,28–30,40,41,46–52 |
| Vimentin | Intermediate filament expressed in mesenchymal cells | Positive expression by PCR and immunostaining | 23,26,35,40,46,47,52–54 |
| CD11b, CD14, CD34, CD45 | Hematopoietic cell markers | Negative by PCR, FC, and/or immunostaining | 17–19,23–25,28,30,40,41,46–52 |
| CD117/c-kit | Stem cell growth factor receptor; expressed by hematopoietic stem cells as well as variety of other cell types | Mixed results: positive expression by PCR, but negative to only slight (∼25%) positive expression by FC | 17,18,37,29,50 |
| CD31, CD144, CD309 | Endothelial cell markers | Negative by FC and immunostaining | 18,19,24–26,47,49,50,53 |
| vWF | von Willebrand Factor; marker of endothelial cells | Mixed results: negative by FC but both positive and negative expression by immunostaining reported | 25,26,47,52 |
| Desmin | Intermediate filament found in the Z-line of sarcomeres in muscle | Negative by immunostaining | 26,53 |
ESC, embryonic stem cells; MSC, mesenchymal stem cells; CD, cluster of differentiation; vWF, von Willebrand factor; MHC, major histocompatability complex; SSEA, stage-specific embryonic antigen; PCR, polymerase chain reaction; FC, flow cytometry.
In Vitro Cardiac Myocyte Differentiation of AFSC
AFSC have been investigated for cardiac tissue engineering.55 AFSC applications to in vitro cardiac tissue engineering are summarized in this section and Table 3. The most common method tested for differentiation of AFSC toward a cardiac lineage is direct cocultures of AFSC and neonatal rat ventricular myocytes (NRVM), in which the two cell types are mixed and plated together.46–48,50,52 Additionally, some scientific groups have used AFSC for cell seeding of heart valves, such as seeding a fibrin gel as a cell carrier for AFSC for heart valve engineering.56
Table 3.
In Vitro Cardiac Differentiation of Amniotic Fluid-Derived Stem Cell
| Method | Length of culture (days) | Result | References |
|---|---|---|---|
| Human AFSC and NRVM coculture, 1:4 ratio | 10 | 5% of AFSC displayed cTnI protein expression and sarcomere banding; bi-nucleated cells observed | 47 |
| Human AFSC and NRVM coculture, 1:10 ratio | 7 or 10 | Functional gap junctions observed; Some cells observed to express human mitochondria as well as cTnT or α-actinin | 48,50 |
| Rat AFSC and NRVM coculture, 1:4 ratio | 9 | Cardiac differentiation observed in 3.5% to 16% of AFSC, determined by sarcomeric α-actinin, myosin heavy chain, cTnT, or cTnI protein expression; bi-nucleated cells observed | 46,52 |
| Cardiomyogenic differentiation medium | 15 | No differentiation observed | 47,52 |
| NRVM-conditioned medium | 10 | No differentiation observed | 46,47,52 |
| 10 μM 5-aza-2′deoxycytidine for 24 h | 10 | Increased cTnI and cTnT gene expression; decreased Sox2 gene expression; no mature sarcomeres or beating cells | 48 |
NRVM, neonatal rat ventricular myocytes; cTnI, cardiac troponin I; cTnT–cardiac troponin T.
Undifferentiated c-kit sorted AFSC have been shown to express some genes common in cardiac cells, including sarcomeric proteins myosin light chain 2v (MLC2v) and β-myosin heavy chain; transcription factors Nkx2.5, Gata4, Mef 2C, and Mef 2D; gap junction protein connexin 43; and adhesion proteins N- and H-cadherin.47,48 However, no protein expression was observed of MLC2v, β-myosin heavy chain, Nkx2.5, or Gata4.47 While connexin 43 and cadherins were observed at the protein level, they did not localize to the membrane in undifferentiated AFSC.47 It is important for researchers to understand the common gene expression between AFSC and cardiomyocytes because during differentiation gene expression is commonly examined to evaluate the extent of differentiation. Therefore, it is necessary to correctly understand the starting state of the AFSC in order to evaluate the effectiveness of tested differentiation methods. Additionally, expression of cardiac-specific genes, even in the absence of protein expression, may support the hypothesis that AFSC are capable of cardiomyocyte differentiation.48
In 2007, Chiavegato et al. studied cardiac differentiation of c-kit-sorted human AFSC.47 The AFSC were labeled with a fluorescent dye and mixed with NRVM at a ratio of 1:4, respectively. These cells were plated on gelatin-coated surfaces and cultured for 10 days, at which time immunofluorescence was performed. Approximately 5% of labeled AFSC displayed cardiac troponin I (cTnI) expression and showed A and I bands of sarcomeres at high magnification. However, at least some of these cells also appeared to have double nuclei, which the authors suggest supports a cell fusion explanation, rather than differentiation of the stem cells.47 Similarly, Iop et al. observed that in cocultures of rat AFSC with NRVM, ∼3.5% of rat AFSC expressed sarcomeric myosin or cardiac troponin T (cTnT). Again, many cells had more than one nuclei and the authors remained unsure if this occurrence of double nuclei is due to cell fusion or dividing cells.52
Further coculture studies of rat AFSC demonstrated additional promise of the use of AFSC in cardiac tissue engineering. In one example, rat AFSC were isolated by c-kit sorting from animals expressing green fluorescent protein (GFP), which facilitated separation of NRVM from GFP-negative rats. It was found that these undifferentiated rat AFSC had low levels of protein expression of both MHC I and MHC II.46 After 4 days of coculture with NRVM, some AFSC in cell clusters rich in NRVM expressed cTnI and MHC proteins, but only a minority of these clusters containing AFSC were beating. After 9 days, cardiac sarcomeric α-actinin was also detected in some AFSC. As in previously described coculture experiments, some bi-nucleated cells were detectable, suggesting that at least some of these events may be due to cell fusion. Furthermore, myocardial differentiation of the AFSC was found to be ∼16% of AFSC after 9 days of coculture. Electrical stimulation studies were also conducted to promote differentiation toward a cardiac lineage.46 Undifferentiated AFSC could not be excited through electrical stimulation, but after coculture they developed electrical excitability. Interestingly, only those AFSC that were in close contact with NRVM in the cocultures developed this excitability, suggesting that either cell fusion or short-range signals may be mediating this transformation.
A later study by Guan et al. supported that cocultures with NRVM could induce human AFSC to differentiate toward a cardiogenic lineage by observing observations that AFSC in coculture with NRVM could form functional gap junctions between the two cell types.48 Additionally, connexin 43 and N-cadherin were seen to localize to the membrane in AFSC in coculture with NRVM. Functional gap junctions and connexin 43 or N-cadherin localization to the membrane were not observed in cultures of AFSC alone or AFSC-fibroblast cocultures.48
AF cells cultured without a c-kit sorting step in a growth media of alpha-modified Eagle's medium, 20% FBS, and 4 ng/mL basic fibroblast growth factor have also been tested for ability to differentiate down a cardiac lineage.50 Again, the AF cells were directly mixed and plated with NRVM, and then cultured for 10 days. Cells were stained with antibodies specific for human mitochondria, as well as antibodies to α-actinin and cTnT. Some cells stained positive for human mitochondria, α-actinin, and cTnT. Connexin 43 was observed at the junctions between AFSC and NRVM.50
NRVM-conditioned media and a cardiomyogenic differentiation medium (60% Dulbecco's modified Eagle's medium (DMEM), 28% MCDB-201, 10−4 M ascorbate phosphate, 10−9 M dexamethasone, 10% fetal bovine serum, 1% antibiotics, and 1% ITS+medium supplement) previously used on MSC57 have also been tested on both human and rat AFSC.46,47,52 No changes were observed in the protein expression of these two conditions, compared to undifferentiated AFSC after a 10-day culture period.46,47,52 These results suggest that either fusion or specific features of NRVM, such as cell–cell contact or short-range soluble signals may be the cause of observed differentiation behavior in cocultures.
Cardiac differentiation of AFSC using treatment with 10 μM 5-aza-2′deoxycytidine for 24 h has also been tested. After the 24-h treatment, cell culture continued in DMEM with 10% horse serum, 10% fetal bovine serum, and antibiotics, AFSC became enlarged and developed organized stress fibers. At days 5 and 10, quantitative real-time polymerase chain reaction (qRT-PCR) showed progressive increases in cTnI and cTnT, as well as a reduction in Sox2, a transcription factor found in undifferentiated ESCs.48 Despite these results, no mature sarcomeres or spontaneous beating was observed, suggesting that additional cues may be necessary to obtain complete cardiac differentiation.
While these techniques have potential, they are also deficient in ways that merit future consideration. First, while there has been some success with c-kit-sorted cells, c-kit expression is not maintained after subsequent passages.18,47,58 This would seem to suggest that c-kit itself may not be the most important factor in determining cardiac differentiation potential and that future exploration and characterization of AFSC is necessary. Additionally, the differentiation of AFSC toward cardiomyogenic lineage versus cell fusion between AFSC and NRVM is an important issue that needs to be further addressed. This is particularly pertinent since, in other tissue systems, cell fusion has been shown to be the major reason for transdifferentiation of developmentally mature tissues from one tissue to another. The occurrence of cell fusion could limit the application of cocultures for immunogenic reasons.
Implantation of AFSC After Myocardial Infarct
In addition to the differentiation of AFSC toward a cardiac lineage, another area of interest has been the implantation of cells into cardiac tissue following myocardial infarction (MI). This work is motivated by similar studies performed using MSC, which have been shown to improve cardiac function when injected into the heart post-MI.59–62 A summary of AFSC implantation studies and results can be found in Table 4.
Table 4.
Amniotic Fluid-Derived Stem Cell Implantation in Myocardium
| Treatment | Observed results | References |
|---|---|---|
| Dissociated AFSC | Reduce fibrosis and enlargement of left ventricle compared to saline control but not as effective as aggregated AFSC structures | 49,50,54 |
| Proliferation of AFSC observed | 46,52 | |
| AFSC found expressing Nkx2.5, cTnT, α-actinin, smooth muscle α-actin, or vWF | 46,52 | |
| Majority of cells are quickly lost after injection | 46,49,52,54 | |
| Aggregated AFSC-ECM structures | Undifferentiated state of AFSC maintained prior to implantation | 49,54 |
| Cell sheet fragments - AFSC plated on thermo-responsive hydrogels to form confluent cells sheets, then separated into fragments | AFSC retained for 4 weeks in rats injected with aggregated structures | 49,54 |
| Cell bodies - AFSC plated in coated wells such that the cells were unable to attach and instead formed spherical cell bodies | Significant contractile function improvement over both saline controls and dissociated AFSC injections | 49,54 |
| Increased density of vasculature | 54 | |
| Short term effects of dissociated AFSC or AFSC-CM (2 h post-MI) | Reduced infarct size compared to saline treated control | 63 |
AFSC, amniotic fluid-derived stem cells; ECM, extracellular matrix; CM, conditioned medium; MI, myocardial infarction.
Injection of dissociated AFSC
Yeh et al. have previously demonstrated that injection of unsorted, undifferentiated human AFSC into rats at the border zone of infarct regions 1 week after MI caused by left coronary artery ligation led to superior cardiac function over saline injected controls.50 The AFSC-treated group had significantly less myocardial fibrosis and enlargement of the left ventricle than the saline control group. Some cells double-stained positive for human nuclei and cardiac markers Nkx2.5, α-actinin, or cTnT. Again in this instance, the authors point out that it is unknown whether these double-positive cells resulted from differentiation of the AFSC or cell fusion of the AFSC with the rat cardiomyocytes.50 Other researchers observed cells double positive for human-specific anti-platelet nonmuscle myosin (hptMyosin) antibody and cTnT after injection of AFSC into rat hearts. However, the hptMyosin+ cells still had protein expression of SSEA-4, so at best an incomplete transition to cardiomyocytes was achieved.47
Iop et al. have also compared rat bone marrow MSC (BM-MSC) to rat AFSC for cell therapy by injecting each into the periphery of cryoinjured areas on rat hearts.52 Fewer AFSC than BM-MSC were present in the injected area after 24 h, but approximately equal numbers were found after 30 days. Through analysis of mitotic marker phospho-Histone H3 and apoptotic markers, the researchers investigated this occurrence. The amount of apoptotic cells was consistent between the two cell types, but more mitotic cells, and therefore more proliferation, were observed in AFSC. This difference between the two cell types resulted from the superior proliferative capacity of AFSC. It was also found that 34.6% of BM-MSC and 49.6% of AFSC underwent differentiation in vivo to express cTnT, von Willebrand factor (vWF), or smooth muscle α-actin. BM-MSCs formed arterioles, but AFSC formed only capillaries, as AFSC appeared to be unable to give rise to SM α-actin+ cells.52 However, another study found that rat AFSC were observed to express smooth muscle α-actin and be located in both arterioles and capillaries several weeks after injection into a ischemia/reperfusion injury in rats.46 The conflicting results of differentiation of rat AFSC following injected into injured myocardium indicate that more work remains to be done on the differentiation potential and the in vivo beneficial effects of AFSC.
Injection of aggregated AFSC-extracellular matrix structures
While dilation of the left ventricle is significantly reduced in rats injected with dissociated AFSC post-MI, an even greater effect has been found by injecting fragmented cell sheets or cell bodies of AFSC.49,54 One week after acute MI, saline, dissociated AFSC, or aggregates of AFSC in the form of fragmented cell sheets or cell bodies were injected into the border of the infarct zone.49,54 Both forms of AFSC aggregates were compared to saline and dissociated AFSC, but not to the other type of aggregate.
To form injectable cell sheet fragments, human AFSC were plated on collagen I coated thermo-responsive methylcellulose hydrogels. Once a confluent cell layer was obtained, a sterile mesh compressed the cell sheet to separate it into fragments. At room temperature the hydrogel system liquefied and the cell sheet fragments were collected.54 To form AFSC aggregates, the cells were plated into a 96-well plate coated with methylcellulose hydrogel. Cells do not attach to the surface of the methylcellulose hydrogel and instead aggregate to form spherical cell bodies.49
In both the case of cell sheets and cell bodies, the undifferentiated state of AFSC was maintained, as evidenced by expression of vimentin, a common marker of MSC, and SSEA-4, a marker of ESC.49,54 Additionally, these methods allowed for collection of cell sheets with the endogenous extracellular matrix (ECM) maintained, which improved cell retention and survival following injection.49,54
It was found that dissociated AFSC were quickly lost after injection. Lee et al. tracked injected cells using bioluminescene, and while AFSC remained for 4 weeks in rats injected with cell bodies, the signal was barely detectable after 7 days in rats injected with dissociated cells.49 This finding is corroborated by the cell sheet fragment study, which found few AFSC by histological analysis in the dissociated AFSC injected rats at the end of the 4-week experiment compared to the large numbers of AFSC found in rats injected with cell sheet fragments.54 In both cases, the authors hypothesized that the improved engraftment was due to the large size of the cell-ECM aggregates, thus allowing for entrapment in interstitial spaces.49,54 Additionally, evidence of proliferation of AFSC was found in the cell sheet fragment group.54
While injection of dissociated AFSC demonstrated improved contractile function of the left ventricle, injection of AFSC aggregates led to a significant function improvement over both saline and dissociated AFSC injections. Improvement was seen in the reduced size of the infarct area and improved left ventricular function in the aggregate-treated groups compared to the dissociated cell and saline-treated groups.49,54 Density of vasculature was also higher in cell sheet fragment group than saline and dissociated AFSC-injected groups.54
AFSC are cardioprotective after MI
AFSC have been demonstrated to be cardioprotective if injected immediately following acute MI.63 In one experiment, immediately following MI caused by ligation of the left anterior descending coronary artery in adult rats, the hearts were injected with saline, human AFSC, or conditioned media from human AFSC (AFSC-CM), then reperfused for 2 h. Following this protocol, the infarct size was significantly smaller in the AFSC- and AFSC-CM-treated groups compared to the saline control.63 The infarct size was determined by treatment of the hearts with 2,3,5-triphenyltetrazolium chloride, which identifies necrotic areas.64
After 2 h of reperfusion, AFSC were found in the heart, spleen, and lungs of the rats, at increasing concentrations of cells in that order. Surprisingly, no AFSC were found in the liver. Furthermore, only 10% of AFSC in the cardiac tissue expressed apoptotic protein caspase-3, whereas ∼32% of AFSC in the heart stained for vWF, and ∼82% of cells stained for alpha-smooth muscle α-actin. Rats treated with AFSC or AFSC-CM had significantly reduced levels of myocardial apoptosis, as demonstrated by reduced TUNEL-positive apoptotic cardiac cells and reduced apoptotic active caspase-3-positive cells. AFSC were found to secrete the actin monomer-binding protein thymosin β4, which has been shown to be cardioprotective. Given the short time over which AFSC provide a benefit and the finding that they secrete thymosin β4, the authors hypothesize that the benefit is provided through an in situ paracrine effect.63
Immune response and pathologic differentiation of injected AFSC
In studies without the use of immunosuppressants, human c-kit-sorted AFSC were seen to be acutely rejected after injection into the infarct region of rats. An inflammatory response was evoked in the cell-injected region in rats with an induced MI or without, regardless of whether they were immuno-competent, immuno-suppressed, or immuno-deficient.47 This result was disappointing because researchers had hypothesized that AFSC would be immune privileged in much the same way as MSC. The researchers speculated that sorting for c-kit may have actually made the cells more immunogenic, though it appears the cells that were used lacked c-kit expression after passaging and at the time of injection.47 However, as this experiment was cross-species, with human AFSC injected into rats, it may still be possible to use AF cells in an allogenic fashion, in addition to a potential autologous use for congenital defects.
A key issue in using AFSC for the therapeutic applications is to ensure that the cells differentiate properly based on the needs of the clinical application. Interestingly, chondro-osteogenic structures were observed in some hearts after injection of AFSC.47,65 These masses were found in the subendocardial zone of infarct hearts, and the chondrogenic and osteogenic nature of the masses was confirmed with histology.65 It was originally hypothesized that the chondro-osteogenic masses resulted due to AFSC injection,47 but it was later found that the masses appeared in ∼50% of rat hearts with induced MI, regardless of AFSC injection, and infarct size was more correlated with formation of chondro-osteogenic masses than injection of AFSC.65 Injection of AFSC in the absence of MI did not lead to the formation of any chondro-osteogenic masses; therefore, it was concluded that chondro-osteogenic masses present independent of stem cell injection following MI.65
Heart Valve Tissue Engineering with AFSC
Disease or degeneration in the heart valves leads to morbidity and mortality66 in both young and adult patients. Heart valve diseases range from congenital malformations to age-dependent tissue degenerations, constituting a significant clinical challenge. Especially, there is a tremendous need to properly repair malfunctioning heart valves in pediatric tissue engineering since congenital heart defects usually require surgical procedures to correct the problem.67 Therefore, there has been a growing need for living autologous grafts as tissue replacements.67,68 Current replacement heart valves include mechanical and bioprosthetic types with certain limitations.67,69 For example, mechanical valves may cause thrombogenesis67 and are susceptible to infections.70 Similarly, bioengineered heart valves may succumb to calcification and possess poor durability.71 In addition, the existing strategies utilize prostheses without the capacity to grow.67 Fabrication of autologous heart valve replacements is preferred particularly for pediatric applications67,72,73 because clinically it is essential for an engineered tissue to have the ability to grow and remodel as the patient grows up.67 It is of vital importance to correct congenital defects early on to prevent further damage.67 Therefore, fabrication of prenatal autologous implants may be useful for hindering secondary damages in the newborn.68,72,74
The ideal tissue-engineered heart valves would allow for growth and remodeling, ensure complete closure, and be nonthrombogenic. Tissue-engineered heart valves can be fabricated using different approaches, such as seeding cells in decellularized tissues or synthetic scaffolds. For instance, autologous engineered heart valve constructs have been generated using the latter technique.75 In another attempt to create an autologous heart valve, biodegradable scaffolds were seeded with MSC and implanted in the pulmonary valve position in a juvenile sheep model.76 However, major limitations remain in the integrity of the ECM and functionality of the final valve construct.69 Calcification, stenosis, fibrosis, and inadequate elastin production in the engineered heart valves are additional major challenges.
Finding an ideal cell source for engineering heart valves can be challenging in pediatric tissue engineering.67 To avoid immunorejection problems, the use of autologous cells may be important. Although clinically important, a reliable cell source for generation of autologous grafts has not been identified yet.67 There are a number of different cell sources to engineer heart valves, such as AFSC,68,72–74 chorionic villi-derived cells,67,68,74 umbilical cord blood cells,67,68,74,77–81 endothelial progenitor cells (EPCs),74,82 BM-derived cells,74,81,83 adipose-derived cells,74,84 and pluripotent stem cells.74,85 Among these, adult stem cells have been reported to be promising cell sources to restore and maintain tissue function.68 Adult stem cells are usually isolated from blood68,86 or BM.68,86 Alternatively, fetal progenitor cells can be used as an autologous cell type for tissue growth and remodeling. The progenitor cells demonstrate plastic behavior with potential to differentiate into various lineages.73
The use of an appropriate cell type is important to generate functional biological substitutes to treat congenital tissue malformations. Moreover, the choice of 3D scaffold and biological or mechanical stimuli are critical features for engineered implants. Human cells that are isolated from fetal sources have been shown to be feasible to generate neo-tissues.68 With advanced imaging techniques, it is now possible to detect congenital disorders before birth. Harvesting cells from AF during pregnancy may create unique opportunities to fabricate autologous tissue-engineered heart valves, which could be ready by birth.67,68 Current strategies to repair congenital valvular defects require reoperations as somatic growth progresses. For this reason, AFSC that are isolated during pregnancy could be a great alternative to treat such congenital disorders. AF has previously been used as a powerful source to isolate progenitor cells.68,72,73 AFSC are readily accessible,87 easy to harvest, isolate, and expand.68,74,87 The protocol to isolate AF is a simple, routine, and relatively low-risk procedure to the mother and the child,67,68,73 making this cell type more advantageous than the other available sources. Additionally, AFSC contain multiple types of stem cells, which can maintain their proliferation ability while differentiating into different lineages and do not lead to the formation of tumors.87
Studies have demonstrated that heart valve leaflets can be engineered using AFSC (Fig. 1).56 In one report, AFSC were isolated from the AF by CD133-affinity magnetic beads.72 Synthetic nondegradable polymers were used as scaffolds and seeded with both CD133+ and CD133− AFSC to fabricate tissue-engineered valve leaflets. These valves were then conditioned in a bioreactor to mature the resulting constructs. It was found that the engineered construct was covered with AFSC and remained functional following 28 days of in vitro culture. In another study, AFSC were studied for their potential in adult heart valve tissue formation.73 Here the authors used cryopreserved AFSC, which were frozen at their seventh passage for 4 months. They demonstrated that both CD133+ and CD133− AFSC preserved their stem cell phenotypes after cryopreservation. This in vitro study demonstrated that heart valve leaflets could be fabricated by seeding synthetic polymers with cryopreserved AFSC, indicating their high potential to be used as a long-term autologous cell source for heart valve tissue engineering. In a similar study, AFSC were used to seed synthetic leaflet scaffolds to fabricate in vitro engineered heart valves.68 The valve leaflets were then endothelialized by EPCs, which were derived from AF and conditioned in a bioreactor. The resulting constructs were analyzed for their chemical, biological, and mechanical properties. These neo-tissues were shown to exhibit similar properties with native heart valves with functional endothelium demonstrating great potential for tissue engineering.
FIG. 1.
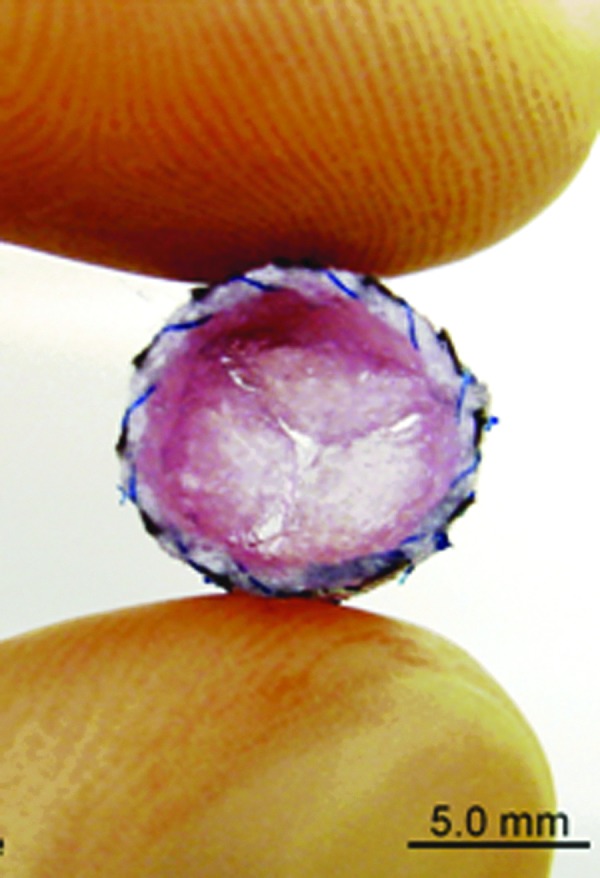
Amniotic fluid-derived stem cell (AFSC)-seeded heart valve leaflets. Nonwoven polyglycolic-acid scaffolds were dip-coated using poly-4-hydroxybutyric acid and seeded with AFSC. Reprinted from Weber et al.,56 with permission from Elsevier. Color images available online at www.liebertpub.com/teb
Collectively, AFSC have been illustrated to be an effective cell source for fabricating engineered heart valves. This could potentially enable the generation of heart valve constructs ready by birth to treat congenital heart malformations with important implications for pediatric tissue engineering research.
Vascular Therapies Utilizing AFSC
Cardiovascular diseases constitute a major focus in therapeutic and engineering approaches to treat vascular malfunctions.88,89 One of the main challenges in cardiovascular tissue engineering is to repair the damage in endothelium and further endothelialize artificial grafts or constructs for in vivo applications.77 Engineering strategies could be utilized for generation of artificial tissue replacements for damaged or diseased vascular tissues by controlling cell–material interactions.90,91 Endothelial cell lines may be used with biomaterials to treat vascular diseases in therapeutic and engineering approaches. Endothelialization could be achieved with different types of cells, such as ESC, AFSC, EPC, or human umbilical vein cord endothelial cells.
Differentiation of AFSC into an endothelial lineage has been successfully demonstrated by Benavides et al.18 In this example, the AF, originally isolated for therapeutic purposes, was enriched by an immunoselection for expression of c-kit. Subsequently, these c-kit+ cells were cultured in endothelial growth medium supplemented by different doses of vascular endothelial growth factor (VEGF) (1–100 ng/mL) for 14 days. AFSC-derived endothelial cells were shown to express endothelial markers, such as VEGF receptor-2 (VEGF-R2), von vWF, CD31, endothelial nitric oxide synthase (eNOS), and vascular endothelial cadherin. These cells also demonstrated endothelial morphology and formed interconnected networks on Matrigel coated culture plates. Furthermore, uptake of acetylated low-density lipoprotein (ac-LDL), and production of nitric oxide and VEGF were assayed to confirm functionality of resulting endothelial cells. Sample images of AFSC-derived endothelial cells are shown in Figure 2. This could have important implications in vascularization of artificial tissues.
FIG. 2.
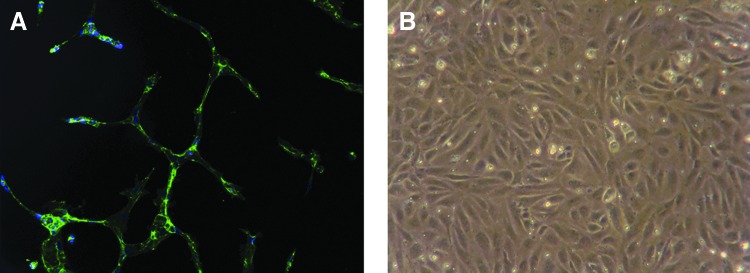
Morphology of AFSC-derived endothelial cells. Formation of network by AFSC-derived endothelial cells on Matrigel coating (A). Cobblestone-like morphology of confluent AFSC-derived endothelial cells (B). Color images available online at www.liebertpub.com/teb
There are a number of different factors, such as mechanical forces,92 oxygen tension,51 growth factors, or cytokines93 that can be used to promote differentiation of AFSC into an endothelial lineage. In one study, AFSC were cultured in endothelial media for 3 weeks to induce endothelial differentiation.51 Expressions of vWF, CD31, and eNOS were used to confirm endothelial lineage by RT-PCR. The function of the resulting endothelial cells was demonstrated by the uptake of ac-LDL and formation of vascular networks on a Matrigel coating. Endothelial cell markers were shown to be overexpressed when AFSC were further exposed to physiological shear stress conditions. Moreover, exposure to hypoxic environment induced the expression of angiogenic factors, such as hepatocyte growth factor, placental growth factor, and VEGF by endothelial cells. This study demonstrates that AFSC are a promising cell source, which can be differentiated into endothelial cells for potential applications in cardiovascular medicine.
Another major limitation in cardiovascular tissue engineering is the vascularization of injured myocardium.52 Cardiovascular type differentiation can be achieved by the use of isolated stem cells from different sources, such as adipose tissue,94 BM,95 embryonic,96 or AF.15,17,51,97–100 AFSC offer several advantages over embryonic or adult stem cells. For instance, they proliferate quickly, have the ability to differentiate into cells from all three germ layers,15 express pluripotency markers indicative of embryonic and adult stem cells,53,101,102 and do not form teratomas.103 For these reasons, AFSC are a promising cell source to vascularize cardiovascular tissues. This ability is of great importance for various applications. For example, it has been reported that the contractile function of the cardiac muscle could be restored through proper vascularization.104 In addition, it may be possible to prevent deterioration of the muscle tissue by inducing angiogenesis in ischemic cardiac injuries.
Limitations of AFSC
AFSC have many potential advantages over alternative cell sources for cardiovascular tissue engineering including broad multipotency,15 lack of tumor formation,15 and the possibility of using autologous cells for congenital heart defects.18 Despite the promise AFSC hold, there are still limitations to this cell source. The lack of agreement on isolation and culture method, as displayed in Table 1, is one limit on the use of AFSC as the differences in isolation and culture method make it uncertain if the same populations of cells are being compared across experiments. Another limitation of AFSC for cardiac muscle tissue engineering is that functionally mature contracting cardiomyocytes have not yet been obtained.48,50
Conclusion
AFSC are a promising cell source for cardiovascular therapies, regenerative medicine, and vascular tissue engineering. AFSC hold specific utility in the repair of congenital defects in a newborn infant or late-term fetus, because the fluid can be collected with relatively minimal risk, isolated stem cells have a greater differentiation potential than other adult stem cell sources, and generated cell-containing constructs are genetically matched to the fetus or newborn infant and can be used for repair without risk of immunorejection. Research studies have demonstrated that AFSC could induce or enhance the enothelialization and vascularization processes in engineered constructs, and can be used in cardiac muscle tissue, cardiac valve, and vessel tissue engineering applications. However, research into the characterization, processing, and use of these cells is limited, especially compared to other adult MSC and ESC sources. Further basic research in this area is necessary to lead us to these goals.
Acknowledgments
J.P.C. was funded by the NSF GRFP (0940902). J.G.J. was funded by the NSF CAREER and AHA BGIA. A.K. was funded by the NIH (EB012597, HL099073, DE019024) and the NSF CAREER award.
Disclosure Statement
No competing financial interests exist.
References
- 1.Kochanek K.D. Kirmeyer S.E. Martin J.A. Strobino D.M. Guyer B. Annual summary of vital statistics: 2009. Pediatrics. 2012;129:338. doi: 10.1542/peds.2011-3435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Lloyd-Jones D. Adams R.J. Brown T.M. Carnethon M. Dai S. De Simone G., et al. Heart disease and stroke statistics—2010 update: a report from the american heart association. Circulation. 2010;121:e46. doi: 10.1161/CIRCULATIONAHA.109.192667. [DOI] [PubMed] [Google Scholar]
- 3.Jonas R. London: Hodder Arnold; 2004. Comprehensive Surgical Management of Congenital Heart Disease. [Google Scholar]
- 4.Silka M.J. Hardy B.G. Menashe V.D. Morris C.D. A population-based prospective evaluation of risk of sudden cardiac death after operation for common congenital heart defects. J Am Coll Cardiol. 1998;32:245. doi: 10.1016/s0735-1097(98)00187-9. [DOI] [PubMed] [Google Scholar]
- 5.Brown J.W. Rodefeld M.D. Ruzmetov M. Eltayeb O. Yurdakok O. Turrentine M.W. Surgical valvuloplasty versus balloon aortic dilation for congenital aortic stenosis: are evidence-based outcomes relevant? Ann Thorac Surg. 2012;94:146. doi: 10.1016/j.athoracsur.2012.02.054. [DOI] [PubMed] [Google Scholar]
- 6.Whitlock R.P. Sun J.C. Fremes S.E. Rubens F.D. Teoh K.H. Antithrombotic and thrombolytic therapy for valvular disease: antithrombotic therapy and prevention of thrombosis, 9th ed: American college of chest physicians evidence-based clinical practice guidelines. Chest. 2012;141:e576S. doi: 10.1378/chest.11-2305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Pok S. Jacot J.G. Biomaterials advances in patches for congenital heart defect repair. J Cardiovasc Transl Res. 2011;4:646. doi: 10.1007/s12265-011-9289-8. [DOI] [PubMed] [Google Scholar]
- 8.Chien K.R. Domian I.J. Parker K.K. Cardiogenesis and the complex biology of regenerative cardiovascular medicine. Science. 2008;322:1494. doi: 10.1126/science.1163267. [DOI] [PubMed] [Google Scholar]
- 9.Vunjak-Novakovic G. Lui K.O. Tandon N. Chien K.R. Bioengineering heart muscle: a paradigm for regenerative medicine. Annu Rev Biomed Eng. 2011;13:245. doi: 10.1146/annurev-bioeng-071910-124701. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hibino N. McGillicuddy E. Matsumura G. Ichihara Y. Naito Y. Breuer C., et al. Late-term results of tissue-engineered vascular grafts in humans. J Thorac Cardiovasc Surg. 2010;139:431. doi: 10.1016/j.jtcvs.2009.09.057. [DOI] [PubMed] [Google Scholar]
- 11.Hogler W. Baumann U. Kelly D. Endocrine and bone metabolic complications in chronic liver disease and after liver transplantation in children. J Pediatr Gastroenterol Nutr. 2012;54:313. doi: 10.1097/MPG.0b013e31823e9412. [DOI] [PubMed] [Google Scholar]
- 12.Sadler T.W. 10th. Philadelphia: Lippincott Williams & Wilkins; 2006. Langman's Medical Embryology. [Google Scholar]
- 13.Antonucci I. Pantalone A. De Amicis D. D'Onofrio S. Stuppia L. Palka G., et al. Human amniotic fluid stem cells culture onto titanium screws: a new perspective for bone engineering. J Biol Regul Homeost Agents. 2009;23:277. [PubMed] [Google Scholar]
- 14.Arnhold S. Gluer S. Hartmann K. Raabe O. Addicks K. Wenisch S., et al. Amniotic-fluid stem cells: Growth dynamics and differentiation potential after a cd-117-based selection procedure. Stem Cells Int. 2011;2011:715341. doi: 10.4061/2011/715341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.De Coppi P. Bartsch G., Jr. Siddiqui M.M. Xu T. Santos C.C. Perin L., et al. Isolation of amniotic stem cell lines with potential for therapy. Nat Biotechnol. 2007;25:100. doi: 10.1038/nbt1274. [DOI] [PubMed] [Google Scholar]
- 16.Prusa A.R. Marton E. Rosner M. Bernaschek G. Hengstschlager M. Oct-4-expressing cells in human amniotic fluid: a new source for stem cell research? Hum Reprod. 2003;18:1489. doi: 10.1093/humrep/deg279. [DOI] [PubMed] [Google Scholar]
- 17.Tsai M.S. Lee J.L. Chang Y.J. Hwang S.M. Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol. Hum Reprod. 2004;19:1450. doi: 10.1093/humrep/deh279. [DOI] [PubMed] [Google Scholar]
- 18.Benavides O.M. Petsche J.J. Moise K.J. Johnson A. Jacot J. Evaluation of endothelial cells differentiated from amniotic fluid-derived stem cells. Tissue Eng Part A. 2012;18:1123. doi: 10.1089/ten.tea.2011.0392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.You Q. Tong X. Guan Y. Zhang D. Huang M. Zhang Y., et al. The biological characteristics of human third trimester amniotic fluid stem cells. J Int Med Res. 2009;37:105. doi: 10.1177/147323000903700112. [DOI] [PubMed] [Google Scholar]
- 20.Bossolasco P. Montemurro T. Cova L. Zangrossi S. Calzarossa C. Buiatiotis S., et al. Molecular and phenotypic characterization of human amniotic fluid cells and their differentiation potential. Cell Res. 2006;16:329. doi: 10.1038/sj.cr.7310043. [DOI] [PubMed] [Google Scholar]
- 21.Niwa H. Miyazaki J. Smith A.G. Quantitative expression of oct-3/4 defines differentiation, dedifferentiation or self-renewal of es cells. Nat Genet. 2000;24:372. doi: 10.1038/74199. [DOI] [PubMed] [Google Scholar]
- 22.Martin F.H. Suggs S.V. Langley K.E. Lu H.S. Ting J. Okino K.H., et al. Primary structure and functional expression of rat and human stem cell factor dnas. Cell. 1990;63:203. doi: 10.1016/0092-8674(90)90301-t. [DOI] [PubMed] [Google Scholar]
- 23.Seo J.M. Sohn M.Y. Suh J.S. Atala A. Yoo J.J. Shon Y.H. Cryopreservation of amniotic fluid-derived stem cells using natural cryoprotectants and low concentrations of dimethylsulfoxide. Cryobiology. 2011;62:167. doi: 10.1016/j.cryobiol.2011.02.003. [DOI] [PubMed] [Google Scholar]
- 24.You Q. Cai L. Zheng J. Tong X. Zhang D. Zhang Y. Isolation of human mesenchymal stem cells from third-trimester amniotic fluid. Int J Gynaecol Obstet. 2008;103:149. doi: 10.1016/j.ijgo.2008.06.012. [DOI] [PubMed] [Google Scholar]
- 25.Sessarego N. Parodi A. Podesta M. Benvenuto F. Mogni M. Raviolo V., et al. Multipotent mesenchymal stromal cells from amniotic fluid: solid perspectives for clinical application. Haematologica. 2008;93:339. doi: 10.3324/haematol.11869. [DOI] [PubMed] [Google Scholar]
- 26.Kaviani A. Guleserian K. Perry T.E. Jennings R.W. Ziegler M.M. Fauza D.O. Fetal tissue engineering from amniotic fluid. J Am Coll Surg. 2003;196:592. doi: 10.1016/s1072-7515(02)01834-3. [DOI] [PubMed] [Google Scholar]
- 27.Steigman S.A. Fauza D.O. Isolation of mesenchymal stem cells from amniotic fluid and placenta. Curr Protoc Stem Cell Biol. 2007;Chapter 1 doi: 10.1002/9780470151808.sc01e02s1. Unit 1E.2. [DOI] [PubMed] [Google Scholar]
- 28.Steigman S.A. Armant M. Bayer-Zwirello L. Kao G.S. Silberstein L. Ritz J., et al. Preclinical regulatory validation of a 3-stage amniotic mesenchymal stem cell manufacturing protocol. J Pediatr Surg. 2008;43:1164. doi: 10.1016/j.jpedsurg.2008.02.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Filioli Uranio M. Valentini L. Lange-Consiglio A. Caira M. Guaricci A.C. L'Abbate A., et al. Isolation, proliferation, cytogenetic, and molecular characterization and in vitro differentiation potency of canine stem cells from foetal adnexa: a comparative study of amniotic fluid, amnion, and umbilical cord matrix. Mol Reprod Dev. 2011;78:361. doi: 10.1002/mrd.21311. [DOI] [PubMed] [Google Scholar]
- 30.Lovati A.B. Corradetti B. Lange Consiglio A. Recordati C. Bonacina E. Bizzaro D., et al. Comparison of equine bone marrow-, umbilical cord matrix and amniotic fluid-derived progenitor cells. Vet Res Commun. 2011;35:103. doi: 10.1007/s11259-010-9457-3. [DOI] [PubMed] [Google Scholar]
- 31.Mauro A. Turriani M. Ioannoni A. Russo V. Martelli A. Di Giacinto O., et al. Isolation, characterization, and in vitro differentiation of ovine amniotic stem cells. Vet Res Commun. 2010;34:S25. doi: 10.1007/s11259-010-9393-2. [DOI] [PubMed] [Google Scholar]
- 32.Peister A. Deutsch E.R. Kolambkar Y. Hutmacher D.W. Guldberg R.E. Amniotic fluid stem cells produce robust mineral deposits on biodegradable scaffolds. Tissue Eng Part A. 2009;15:3129. doi: 10.1089/ten.tea.2008.0536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Perin L. Giuliani S. Jin D. Sedrakyan S. Carraro G. Habibian R., et al. Renal differentiation of amniotic fluid stem cells. Cell Prolif. 2007;40:936. doi: 10.1111/j.1365-2184.2007.00478.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Siegel N. Rosner M. Unbekandt M. Fuchs C. Slabina N. Dolznig H., et al. Contribution of human amniotic fluid stem cells to renal tissue formation depends on mtor. Hum Mol Genet. 2010;19:3320. doi: 10.1093/hmg/ddq236. [DOI] [PubMed] [Google Scholar]
- 35.Gekas J. Walther G. Skuk D. Bujold E. Harvey I. Bertrand O.F. In vitro and in vivo study of human amniotic fluid-derived stem cell differentiation into myogenic lineage. Clin Exp Med. 2010;10:1. doi: 10.1007/s10238-009-0060-2. [DOI] [PubMed] [Google Scholar]
- 36.Carraro G. Perin L. Sedrakyan S. Giuliani S. Tiozzo C. Lee J., et al. Human amniotic fluid stem cells can integrate and differentiate into epithelial lung lineages. Stem Cells. 2008;26:2902. doi: 10.1634/stemcells.2008-0090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Siegel N. Valli A. Fuchs C. Rosner M. Hengstschlager M. Induction of mesenchymal/epithelial marker expression in human amniotic fluid stem cells. Reprod Biomed Online. 2009;19:838. doi: 10.1016/j.rbmo.2009.09.015. [DOI] [PubMed] [Google Scholar]
- 38.Arnhold S. Post C. Gluer S. Hoopmann M. Wenisch S. Volpers C., et al. Neuronal characteristics of amniotic fluid derived cells after adenoviral transformation. Cell Biol Int. 2008;32:1559. doi: 10.1016/j.cellbi.2008.09.002. [DOI] [PubMed] [Google Scholar]
- 39.Jezierski A. Gruslin A. Tremblay R. Ly D. Smith C. Turksen K., et al. Probing stemness and neural commitment in human amniotic fluid cells. Stem Cell Rev. 2010;6:199. doi: 10.1007/s12015-010-9116-7. [DOI] [PubMed] [Google Scholar]
- 40.Mareschi K. Rustichelli D. Comunanza V. De Fazio R. Cravero C. Morterra G., et al. Multipotent mesenchymal stem cells from amniotic fluid originate neural precursors with functional voltage-gated sodium channels. Cytotherapy. 2009;11:534. doi: 10.1080/14653240902974024. [DOI] [PubMed] [Google Scholar]
- 41.McLaughlin D. Tsirimonaki E. Vallianatos G. Sakellaridis N. Chatzistamatiou T. Stavropoulos-Gioka C., et al. Stable expression of a neuronal dopaminergic progenitor phenotype in cell lines derived from human amniotic fluid cells. J Neurosci Res. 2006;83:1190. doi: 10.1002/jnr.20828. [DOI] [PubMed] [Google Scholar]
- 42.Orciani M. Emanuelli M. Martino C. Pugnaloni A. Tranquilli A.L. Di Primio R. Potential role of culture mediums for successful isolation and neuronal differentiation of amniotic fluid stem cells. Int J Immunopathol Pharmacol. 2008;21:595. doi: 10.1177/039463200802100312. [DOI] [PubMed] [Google Scholar]
- 43.Pfeiffer S. Boyle J. Daly S. Dowd E. Haase J. McLaughlin D. Human amniocytes regulate serotonin levels by active uptake and express genes suggestive of a wider role in facilitating neurotransmitter regulation in the fetal environment. Stem Cells Dev. 2011;20:341. doi: 10.1089/scd.2009.0500. [DOI] [PubMed] [Google Scholar]
- 44.Pfeiffer S. McLaughlin D. In vitro differentiation of human amniotic fluid-derived cells: augmentation towards a neuronal dopaminergic phenotype. Cell Biol Int. 2010;34:959. doi: 10.1042/CBI20090445. [DOI] [PubMed] [Google Scholar]
- 45.Donaldson A.E. Cai J. Yang M. Iacovitti L. Human amniotic fluid stem cells do not differentiate into dopamine neurons in vitro or after transplantation in vivo. Stem Cells Dev. 2009;18:1003. doi: 10.1089/scd.2008.0300. [DOI] [PubMed] [Google Scholar]
- 46.Bollini S. Pozzobon M. Nobles M. Riegler J. Dong X. Piccoli M., et al. In vitro and in vivo cardiomyogenic differentiation of amniotic fluid stem cells. Stem Cell Rev. 2011;7:364. doi: 10.1007/s12015-010-9200-z. [DOI] [PubMed] [Google Scholar]
- 47.Chiavegato A. Bollini S. Pozzobon M. Callegari A. Gasparotto L. Taiani J., et al. Human amniotic fluid-derived stem cells are rejected after transplantation in the myocardium of normal, ischemic, immuno-suppressed or immuno-deficient rat. J Mol Cell Cardiol. 2007;42:746. doi: 10.1016/j.yjmcc.2006.12.008. [DOI] [PubMed] [Google Scholar]
- 48.Guan X. Delo D.M. Atala A. Soker S. In vitro cardiomyogenic potential of human amniotic fluid stem cells. J Tissue Eng Regen Med. 2011;5:220. doi: 10.1002/term.308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lee W.Y. Wei H.J. Lin W.W. Yeh Y.C. Hwang S.M. Wang J.J., et al. Enhancement of cell retention and functional benefits in myocardial infarction using human amniotic-fluid stem-cell bodies enriched with endogenous ECM. Biomaterials. 2011;32:5558. doi: 10.1016/j.biomaterials.2011.04.031. [DOI] [PubMed] [Google Scholar]
- 50.Yeh Y.C. Wei H.J. Lee W.Y. Yu C.L. Chang Y. Hsu L.W., et al. Cellular cardiomyoplasty with human amniotic fluid stem cells: In vitro and in vivo studies. Tissue Eng Part A. 2010;16:1925. doi: 10.1089/ten.TEA.2009.0728. [DOI] [PubMed] [Google Scholar]
- 51.Zhang P. Baxter J. Vinod K. Tulenko T.N. Di Muzio P.J. Endothelial differentiation of amniotic fluid-derived stem cells: Synergism of biochemical and shear force stimuli. Stem Cells Dev. 2009;18:1299. doi: 10.1089/scd.2008.0331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Iop L. Chiavegato A. Callegari A. Bollini S. Piccoli M. Pozzobon M., et al. Different cardiovascular potential of adult- and fetal-type mesenchymal stem cells in a rat model of heart cryoinjury. Cell Transplant. 2008;17:679. doi: 10.3727/096368908786092739. [DOI] [PubMed] [Google Scholar]
- 53.Kaviani A. Perry T.E. Dzakovic A. Jennings R.W. Ziegler M.M. Fauza D.O. The amniotic fluid as a source of cells for fetal tissue engineering. J Pediatr Surg. 2001;36:1662. doi: 10.1053/jpsu.2001.27945. [DOI] [PubMed] [Google Scholar]
- 54.Yeh Y.C. Lee W.Y. Yu C.L. Hwang S.M. Chung M.F. Hsu L.W., et al. Cardiac repair with injectable cell sheet fragments of human amniotic fluid stem cells in an immune-suppressed rat model. Biomaterials. 2010;31:6444. doi: 10.1016/j.biomaterials.2010.04.069. [DOI] [PubMed] [Google Scholar]
- 55.Weber B. Emmert M.Y. Hoerstrup S.P. Stem cells for heart valve regeneration. Swiss Med Wkly. 2012;142:w13622. doi: 10.4414/smw.2012.13622. [DOI] [PubMed] [Google Scholar]
- 56.Weber B. Emmert M.Y. Behr L. Schoenauer R. Brokopp C. Drogemuller C., et al. Prenatally engineered autologous amniotic fluid stem cell-based heart valves in the fetal circulation. Biomaterials. 2012;33:4031. doi: 10.1016/j.biomaterials.2011.11.087. [DOI] [PubMed] [Google Scholar]
- 57.Shim W.S. Jiang S. Wong P. Tan J. Chua Y.L. Tan Y.S., et al. Ex vivo differentiation of human adult bone marrow stem cells into cardiomyocyte-like cells. Biochem Biophys Res Commun. 2004;324:481. doi: 10.1016/j.bbrc.2004.09.087. [DOI] [PubMed] [Google Scholar]
- 58.Chen W.Q. Siegel N. Li L. Pollak A. Hengstschlager M. Lubec G. Variations of protein levels in human amniotic fluid stem cells cd117/2 over passages 5–25. J Proteome Res. 2009;8:5285. doi: 10.1021/pr900630s. [DOI] [PubMed] [Google Scholar]
- 59.Noort W.A. Oerlemans M.I. Rozemuller H. Feyen D. Jaksani S. Stecher D., et al. Human versus porcine mesenchymal stromal cells: phenotype, differentiation potential, immunomodulation and cardiac improvement after transplantation. J Cell Mol Med. 2012;16:1827. doi: 10.1111/j.1582-4934.2011.01455.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Numasawa Y. Kimura T. Miyoshi S. Nishiyama N. Hida N. Tsuji H., et al. Treatment of human mesenchymal stem cells with angiotensin receptor blocker improved efficiency of cardiomyogenic transdifferentiation and improved cardiac function via angiogenesis. Stem Cells. 2011;29:1405. doi: 10.1002/stem.691. [DOI] [PubMed] [Google Scholar]
- 61.Peng C. Yang K. Xiang P. Zhang C. Zou L. Wu X., et al. Effect of transplantation with autologous bone marrow stem cells on acute myocardial infarction. Int J Cardiol. 2013;162:158. doi: 10.1016/j.ijcard.2011.05.077. [DOI] [PubMed] [Google Scholar]
- 62.Trachtenberg B. Velazquez D.L. Williams A.R. McNiece I. Fishman J. Nguyen K., et al. Rationale and design of the transendocardial injection of autologous human cells (bone marrow or mesenchymal) in chronic ischemic left ventricular dysfunction and heart failure secondary to myocardial infarction (tac-hft) trial: a randomized, double-blind, placebo-controlled study of safety and efficacy. Am Heart J. 2011;161:487. doi: 10.1016/j.ahj.2010.11.024. [DOI] [PubMed] [Google Scholar]
- 63.Bollini S. Cheung K.K. Riegler J. Dong X. Smart N. Ghionzoli M., et al. Amniotic fluid stem cells are cardioprotective following acute myocardial infarction. Stem Cells Dev. 2011;20:1985. doi: 10.1089/scd.2010.0424. [DOI] [PubMed] [Google Scholar]
- 64.Fishbein M.C. Meerbaum S. Rit J. Lando U. Kanmatsuse K. Mercier J.C., et al. Early phase acute myocardial infarct size quantification: validation of the triphenyl tetrazolium chloride tissue enzyme staining technique. Am Heart J. 1981;101:593. doi: 10.1016/0002-8703(81)90226-x. [DOI] [PubMed] [Google Scholar]
- 65.Delo D.M. Guan X. Wang Z. Groban L. Callahan M. Smith T., et al. Calcification after myocardial infarction is independent of amniotic fluid stem cell injection. Cardiovasc Pathol. 2011;20:e69. doi: 10.1016/j.carpath.2010.03.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Filova E. Straka F. Mirejovsky T. Masin J. Bacakova L. Tissue-engineered heart valves. Physiol Res. 2009;58:s141. doi: 10.33549/physiolres.931919. [DOI] [PubMed] [Google Scholar]
- 67.Schmidt D. Mol A. Breymann C. Achermann J. Odermatt B. Gossi M., et al. Living autologous heart valves engineered from human prenatally harvested progenitors. Circulation. 2006;114:I125. doi: 10.1161/CIRCULATIONAHA.105.001040. [DOI] [PubMed] [Google Scholar]
- 68.Weber B. Zeisberger S.M. Hoerstrup S.P. Prenatally harvested cells for cardiovascular tissue engineering: fabrication of autologous implants prior to birth. Placenta. 2011;32:S316. doi: 10.1016/j.placenta.2011.04.001. [DOI] [PubMed] [Google Scholar]
- 69.Sacks M.S. Schoen F.J. Mayer J.E. Bioengineering challenges for heart valve tissue engineering. Annu Rev Biomed Eng. 2009;11:289. doi: 10.1146/annurev-bioeng-061008-124903. [DOI] [PubMed] [Google Scholar]
- 70.Cannegieter S.C. Rosendaal F.R. Briet E. Thromboembolic and bleeding complications in patients with mechanical heart-valve prostheses. Circulation. 1994;89:635. doi: 10.1161/01.cir.89.2.635. [DOI] [PubMed] [Google Scholar]
- 71.Schoen F.J. Levy R.J. Pathology of substitute heart-valves-new concepts and developments. J Cardiac Surg. 1994;9:222. doi: 10.1111/j.1540-8191.1994.tb00932.x. [DOI] [PubMed] [Google Scholar]
- 72.Schmidt D. Achermann J. Odermatt B. Breymann C. Mol A. Genoni M., et al. Prenatally fabricated autologous human living heart valves based on amniotic fluid-derived progenitor cells as single cell source. Circulation. 2007;116:i64. doi: 10.1161/CIRCULATIONAHA.106.681494. [DOI] [PubMed] [Google Scholar]
- 73.Schmidt D. Achermann J. Odermatt B. Genoni M. Zund G. Hoerstrup S.P. Cryopreserved amniotic fluid-derived cells: a life-long autologous fetal stem cell source for heart valve tissue engineering. J Heart Valve Dis. 2008;17:446. [PubMed] [Google Scholar]
- 74.Weber B. Hoerstrup S.P. Regenerating heart valves. In: Cohen I.S., editor; Gaudette G.R., editor. Regenerating the Heart: Stem Cells and the Cardiovascular System. New York: Humana Press; 2011. pp. 403–442. [Google Scholar]
- 75.Ye Q. Zund G. Jockenhoevel S. Hoerstrup S.P. Schoeberlein A. Grunenfelder J., et al. Tissue engineering in cardiovascular surgery: new approach to develop completely human autologous tissue. Eur J Cardiothorac Surg. 2000;17:449. doi: 10.1016/s1010-7940(00)00371-7. [DOI] [PubMed] [Google Scholar]
- 76.Sutherland F.W.H. Perry T.E. Yu Y. Sherwood M.C. Rabkin E. Masuda Y., et al. From stem cells to viable autologous semilunar heart valve. Circulation. 2005;111:2783. doi: 10.1161/CIRCULATIONAHA.104.498378. [DOI] [PubMed] [Google Scholar]
- 77.Schmidt D. Breymann C. Weber A. Guenter C.I. Neuenschwander S. Zund G., et al. Umbilical cord blood derived endothelial progenitor cells for tissue engineering of vascular grafts. Ann Thorac Surg. 2004;78:2094. doi: 10.1016/j.athoracsur.2004.06.052. [DOI] [PubMed] [Google Scholar]
- 78.Erices A. Conget P. Minguell J.J. Mesenchymal progenitor cells in human umbilical cord blood. Br J Haematol. 2000;109:235. doi: 10.1046/j.1365-2141.2000.01986.x. [DOI] [PubMed] [Google Scholar]
- 79.Melero-Martin J.M. De Obaldia M.E. Kang S.Y. Khan Z.A. Yuan L. Oettgen P., et al. Engineering robust and functional vascular networks in vivo with human adult and cord blood-derived progenitor cells. Circ Res. 2008;103:194. doi: 10.1161/CIRCRESAHA.108.178590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Bieback K. Kern S. Kluter H. Eichler H. Critical parameters for the isolation of mesenchymal stem cells from umbilical cord blood. Stem Cells. 2004;22:625. doi: 10.1634/stemcells.22-4-625. [DOI] [PubMed] [Google Scholar]
- 81.Schmidt D. Stock U.A. Hoerstrup S.P. Tissue engineering of heart valves using decellularized xenogeneic or polymeric starter matrices. Philos Trans R Soc Lond B Biol Sci. 2007;362:1505. doi: 10.1098/rstb.2007.2131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Kim S. Von Recum H. Endothelial stem cells and precursors for tissue engineering: cell source, differentiation, selection, and application. Tissue Eng Part B Rev. 2008;14:133. doi: 10.1089/teb.2007.0304. [DOI] [PubMed] [Google Scholar]
- 83.Perry T.E. Kaushal S. Sutherland F.W.H. Guleserian K.J. Bischoff J. Sacks M., et al. Bone marrow as a cell source for tissue engineering heart valves. Annals of Thorac Surg. 2003;75:761. doi: 10.1016/s0003-4975(02)03776-1. [DOI] [PubMed] [Google Scholar]
- 84.Colazzo F. Sarathchandra P. Smolenski R.T. Chester A.H. Tseng Y.T. Czernuszka J.T., et al. Extracellular matrix production by adipose-derived stem cells: implications for heart valve tissue engineering. Biomaterials. 2011;32:119. doi: 10.1016/j.biomaterials.2010.09.003. [DOI] [PubMed] [Google Scholar]
- 85.Zhang J.H. Wilson G.F. Soerens A.G. Koonce C.H. Yu J.Y. Palecek S.P., et al. Functional cardiomyocytes derived from human induced pluripotent stem cells. Circ Res. 2009;104:E30. doi: 10.1161/CIRCRESAHA.108.192237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Schmidt D. Dijkman P.E. Driessen-Mol A. Stenger R. Mariani C. Puolakka A., et al. Minimally-invasive implantation of living tissue engineered heart valves: a comprehensive approach from autologous vascular cells to stem cells. J Am Coll Cardiol. 2010;56:510. doi: 10.1016/j.jacc.2010.04.024. [DOI] [PubMed] [Google Scholar]
- 87.Badylak S.F. Taylor D. Uygun K. Whole-organ tissue engineering: decellularization and recellularization of three-dimensional matrix scaffolds. In: Yarmush M.L., editor; Duncan J.S., editor; Gray M.L., editor. Annual Review of Biomedical Engineering. Palo Alto, CA: Annual Reviews; 2011. pp. 27–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Camci-Unal G. Nichol J.W. Bae H. Tekin H. Bischoff J. Khademhosseini A. Hydrogel surfaces to promote attachment and spreading of endothelial progenitor cells. J Tissue Eng Regen Med. 2012 doi: 10.1002/term.517. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.de Mel A. Jell G. Stevens M.M. Seifalian A.M. Biofunctionalization of biomaterials for accelerated in situ endothelialization: a review. Biomacromolecules. 2008;9:2969. doi: 10.1021/bm800681k. [DOI] [PubMed] [Google Scholar]
- 90.Camci-Unal G. Aubin H. Ahari A.F. Bae H. Nichol J.W. Khademhosseini A. Surface-modified hyaluronic acid hydrogels to capture endothelial progenitor cells. Soft Matter. 2010;6:5120. doi: 10.1039/c0sm00508h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Avci-Adali M. Paul A. Ziemer G. Wendel H.P. New strategies for in vivo tissue engineering by mimicry of homing factors for self-endothelialisation of blood contacting materials. Biomaterials. 2008;29:3936. doi: 10.1016/j.biomaterials.2008.07.002. [DOI] [PubMed] [Google Scholar]
- 92.Wang H. Riha G.M. Yan S. Li M. Chai H. Yang H., et al. Shear stress induces endothelial differentiation from a murine embryonic mesenchymal progenitor cell line. Arterioscler Thromb Vasc Biol. 2005;25:1817. doi: 10.1161/01.ATV.0000175840.90510.a8. [DOI] [PubMed] [Google Scholar]
- 93.Liu J.W. Dunoyer-Geindre S. Serre-Beinier V. Mai G. Lambert J.F. Fish R.J., et al. Characterization of endothelial-like cells derived from human mesenchymal stem cells. J Thromb Haemost. 2007;5:826. doi: 10.1111/j.1538-7836.2007.02381.x. [DOI] [PubMed] [Google Scholar]
- 94.Cao Y. Sun Z. Liao L.M. Meng Y. Han Q. Zhao R.C.H. Human adipose tissue-derived stem cells differentiate into endothelial cells in vitro and improve postnatal neovascularization in vivo. Biochem Biophys Res Commun. 2005;332:370. doi: 10.1016/j.bbrc.2005.04.135. [DOI] [PubMed] [Google Scholar]
- 95.Oswald J. Boxberger S. Jorgensen B. Feldmann S. Ehninger G. Bornhauser M., et al. Mesenchymal stem cells can be differentiated into endothelial cells in vitro. Stem Cells. 2004;22:377. doi: 10.1634/stemcells.22-3-377. [DOI] [PubMed] [Google Scholar]
- 96.Li Z. Wu J.C. Sheikh A.Y. Kraft D. Cao F. Xie X.Y., et al. Differentiation, survival, and function of embryonic stem cell-derived endothelial cells for ischemic heart disease. Circulation. 2007;116:I46. doi: 10.1161/CIRCULATIONAHA.106.680561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kunisaki S.M. Armant M. Kao G.S. Stevenson K. Kim H. Fauza D.O. Tissue engineering from human mesenchymal amniocytes: a prelude to clinical trials. J Pediatr Surg. 2007;42:974. doi: 10.1016/j.jpedsurg.2007.01.031. [DOI] [PubMed] [Google Scholar]
- 98.Moorefield E.C. Delo D.M. De Coppi P. Atala A. Pluripotent Stem Cells. New York: Nova Science; 2010. Amniotic Fluid and Placental Stem Cells; pp. 113–135. [Google Scholar]
- 99.Kim J. Lee Y. Kim H. Hwang K.J. Kwon H.C. Kim S.K., et al. Human amniotic fluid-derived stem cells have characteristics of multipotent stem cells. Cell Prolif. 2007;40:75. doi: 10.1111/j.1365-2184.2007.00414.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Siegel N. Rosner M. Hanneder M. Freilinger A. Hengstschlager M. Human amniotic fluid stem cells: a new perspective. Amino Acids. 2008;35:291. doi: 10.1007/s00726-007-0593-1. [DOI] [PubMed] [Google Scholar]
- 101.Prusa A.R. Marton E. Rosner M. Bettelheim D. Lubec G. Pollack A., et al. Neurogenic cells in human amniotic fluid. Am J Obstet Gynecol. 2004;191:309. doi: 10.1016/j.ajog.2003.12.014. [DOI] [PubMed] [Google Scholar]
- 102.Cipriani S. Bonini D. Marchina E. Balgkouranidou I. Caimi L. Zucconi G.G., et al. Mesenchymal cells from human amniotic fluid survive and migrate after transplantation into adult rat brain. Cell Biol Int. 2007;31:845. doi: 10.1016/j.cellbi.2007.01.037. [DOI] [PubMed] [Google Scholar]
- 103.Hilfiker A. Kasper C. Hass R. Haverich A. Mesenchymal stem cells and progenitor cells in connective tissue engineering and regenerative medicine: Is there a future for transplantation? Langenbecks Arch Surg. 2011;396:489. doi: 10.1007/s00423-011-0762-2. [DOI] [PubMed] [Google Scholar]
- 104.Leri A. Kajstura J. Anversa P. Cardiac stem cells and mechanisms of myocardial regeneration. Physiol Rev. 2005;85:1373. doi: 10.1152/physrev.00013.2005. [DOI] [PubMed] [Google Scholar]