

Abstract
Society values landscapes that reliably provide many ecosystem functions. As the study of ecosystem functioning expands to include more locations, time spans, and functions, the functional importance of individual species is becoming more apparent. However, the functional importance of individual species does not necessarily translate to the functional importance of biodiversity measured in whole communities of interacting species. Furthermore, ecological diversity at scales larger than neighborhood species richness could also influence the provision of multiple functions over extended time scales. We created experimental landscapes based on whole communities from the world’s longest running biodiversity-functioning field experiment to investigate how local species richness (α diversity), distinctness among communities (β diversity), and larger scale species richness (γ diversity) affected eight ecosystem functions over 10 y. Using both threshold-based and unique multifunctionality metrics, we found that α diversity had strong positive effects on most individual functions and multifunctionality, and that positive effects of β and γ diversity emerged only when multiple functions were considered simultaneously. Higher β diversity also reduced the variability in multifunctionality. Thus, in addition to conserving important species, maintaining ecosystem multifunctionality will require diverse landscape mosaics of diverse communities.
Keywords: alpha diversity, gamma diversity, ecosystem service, Cedar Creek, beta diversity
As global biodiversity losses accelerate, a growing body of research has documented the consequences of species losses to ecosystem functions and services. Syntheses of experiments in this field show that local species richness (α diversity) positively affects the provision and maintenance of many ecosystem functions (1, 2). The effects of individual species (3–5) and α diversity (6) also become stronger as more functions are considered simultaneously (ecosystem multifunctionality). However, the small scale and limited number of functions measured in biodiversity experiments do not match the scales at which society usually manages biodiversity or ecosystem functions and services. Furthermore, knowledge of how biodiversity contributes to ecosystem functioning at multiple scales is critical to conserving, managing, and restoring multifunctional landscapes. Larger scale observational studies on biodiversity–ecosystem service relationships are increasing (7–19), but their reliance on correlation and remote-sensing introduces uncertainties that might underlie these studies’ inconsistent findings about the concordance between biodiversity and landscape-level ecosystem multifunctionality.
In addition to species richness within communities (α diversity), it is plausible that the number of distinct communities within a landscape (β diversity) and the total number of unique species among all communities in a landscape (γ diversity) might also contribute to ecosystem multifunctionality (6). No field experiments have directly explored the effects of these higher scales of diversity. Here we analyze the multifunctionality of over 7,500 experimental landscapes, each consisting of a different combination of 24 of the 168 experimental perennial grassland communities of the Biodiversity II experiment at the Cedar Creek Ecosystem Science Reserve in Minnesota (20). We used data from experimental communities in each experimental landscape to determine each landscape’s diversity at three levels (α, β, and γ), and each landscape’s aggregate functioning for eight ecosystem functions (listed in Fig. 1). We did this for 5 different years between 1997 and 2006, and on average across this full time period. In contrast to previous work relying on species-based models (3–5), we measured multifunctionality within 9 m × 9 m experimental communities of interacting species (6). Measuring functions at the level of each experimental community allowed us to assess the effects of species diversity (rather than of the sum of species identities) on ecosystem functioning. This approach incorporates potential tradeoffs that can occur between species and between functions (6), and thus more accurately reflects ecosystem functioning in natural communities.
Fig. 1.

Biodiversity effect sizes. Standardized regression coefficients for 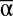 , β, and γ diversities in nine multiple regressions explaining cross-year averages of MF and each single function (soil carbon data from 2006 only). R2s reflect the proportion of variance explained in each regression (P < 0.001 for all coefficients). β diversity is calculated as the number of experimental communities in an experimental landscape that do not share exactly the same species composition. Results with β calculated as 1 – Sorensen’s Index are qualitatively similar, as are analyses of partial r2s (Fig. S1 and Table S2).
, β, and γ diversities in nine multiple regressions explaining cross-year averages of MF and each single function (soil carbon data from 2006 only). R2s reflect the proportion of variance explained in each regression (P < 0.001 for all coefficients). β diversity is calculated as the number of experimental communities in an experimental landscape that do not share exactly the same species composition. Results with β calculated as 1 – Sorensen’s Index are qualitatively similar, as are analyses of partial r2s (Fig. S1 and Table S2).
Results and Discussion
We found that 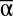 diversity was the strongest contributor to landscape-level multifunctionality, and that β and γ diversity also made significant contributions. Across all years and functions, all three scales of diversity had significant positive effects on multifunctionality measured as the mean of all functions minus its standard deviation in each experimental landscape (MF): MF = 0.46
diversity was the strongest contributor to landscape-level multifunctionality, and that β and γ diversity also made significant contributions. Across all years and functions, all three scales of diversity had significant positive effects on multifunctionality measured as the mean of all functions minus its standard deviation in each experimental landscape (MF): MF = 0.46 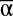 + 0.11β + 0.16γ (P < 0.001, R2 = 0.32, n = 7,512, Fig.1, Table S1). The nature of these effects and their interactions became more apparent after exploring the individual impact of each scale of diversity on MF (Fig. 2). Whereas the main effects of increased
+ 0.11β + 0.16γ (P < 0.001, R2 = 0.32, n = 7,512, Fig.1, Table S1). The nature of these effects and their interactions became more apparent after exploring the individual impact of each scale of diversity on MF (Fig. 2). Whereas the main effects of increased 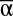 and γ diversity were saturating increases in MF, higher β diversity mainly decreased landscape-to-landscape variance in MF (analysis of residual variance, P < 0.001, r2 = 0.24, n = 7,512).
and γ diversity were saturating increases in MF, higher β diversity mainly decreased landscape-to-landscape variance in MF (analysis of residual variance, P < 0.001, r2 = 0.24, n = 7,512).
Fig. 2.

Effects of 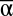 , β, and γ diversity on MF. Relationships between MF and
, β, and γ diversity on MF. Relationships between MF and 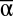 (A), β (B), and γ (C) diversity in 7,512 experimental landscapes.
(A), β (B), and γ (C) diversity in 7,512 experimental landscapes. 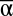 and γ increase multifunctionality, while β mainly decreases its variance. Flexible, best-fit saturating curves are not forced through 0 (P < 0.001 for all). β ranges from 1 (blue) to 24 (red) in A and
and γ increase multifunctionality, while β mainly decreases its variance. Flexible, best-fit saturating curves are not forced through 0 (P < 0.001 for all). β ranges from 1 (blue) to 24 (red) in A and 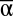 ranges from 1 (blue) to 16 (red) in B and C.
ranges from 1 (blue) to 16 (red) in B and C.
While 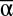 diversity consistently influenced each individual function, all three scales of diversity affected MF (Fig. 1). Likewise, while
diversity consistently influenced each individual function, all three scales of diversity affected MF (Fig. 1). Likewise, while 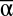 consistently accounted for most of the explainable variation (R2) of single function regressions, the explanatory power of β and γ diversity became much more pronounced when multiple functions were considered simultaneously (Fig. S1). Thus, understanding the drivers of multifunctionality requires examination of multiple scales of biodiversity. Furthermore, the full effect of biodiversity at each scale of diversity is not apparent unless multiple functions are considered together. While consideration of multiple scales of diversity clearly improves our ability to explain the magnitude of and variation in multifunctionality, much variation remains unexplained. Other components of diversity not examined here (e.g., genetic diversity, species evenness, etc.) likely contribute to differences in multifunctionality, as do environmental conditions and broad drivers of global change (21).
consistently accounted for most of the explainable variation (R2) of single function regressions, the explanatory power of β and γ diversity became much more pronounced when multiple functions were considered simultaneously (Fig. S1). Thus, understanding the drivers of multifunctionality requires examination of multiple scales of biodiversity. Furthermore, the full effect of biodiversity at each scale of diversity is not apparent unless multiple functions are considered together. While consideration of multiple scales of diversity clearly improves our ability to explain the magnitude of and variation in multifunctionality, much variation remains unexplained. Other components of diversity not examined here (e.g., genetic diversity, species evenness, etc.) likely contribute to differences in multifunctionality, as do environmental conditions and broad drivers of global change (21).
All three scales of diversity also affected multifunctionality measured as the number of functions that exceeded discrete functional thresholds (T) (6). Functional thresholds were set at six quantiles (T = 0.2, T = 0.3, T = 0.4, T = 0.5, T = 0.6, and T = 0.7) based on the minimum and maximum observed functioning for all functions across all experimental landscapes, and each experimental landscape was assessed for the number of functions it could achieve at each threshold. Unsurprisingly, fewer experimental landscapes could achieve higher numbers of functions as functional thresholds increased. However, experimental landscapes with high mean 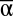 diversity consistently achieved more functions at every threshold than experimental landscapes with lower
diversity consistently achieved more functions at every threshold than experimental landscapes with lower 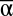 diversity (Table S1 and S2). In contrast, higher β diversity increased the number of functions achieved at lower thresholds but reduced the number achieved at higher thresholds, with the transition occurring between the 50th and 60th quantile-based thresholds (Fig. 3).
diversity (Table S1 and S2). In contrast, higher β diversity increased the number of functions achieved at lower thresholds but reduced the number achieved at higher thresholds, with the transition occurring between the 50th and 60th quantile-based thresholds (Fig. 3).
Fig. 3.

β diversity effects across functional thresholds. Effect of β diversity on the average number of functions achieved by each experimental landscape above thresholds (T) across all years, where T is the quantile-based ranking of each function across all experimental landscapes. Results where T is based on the percent of maximum functioning are qualitatively similar (Fig. S2) (n = 7,512 experimental landscapes, P < 0.001 for all).
The switch in the directional effect of β diversity occurred because only experimental landscapes with low β diversity were capable of achieving very high multifunctionality, whereas high β-diverse experimental landscapes more consistently achieved moderate multifunctionality (Fig. 2B), a phenomenon that became more pronounced as functional thresholds increased (Fig. 3). Thus, increasing β diversity leads to higher multifunctionality only if we require a low functional threshold for each function. As functional thresholds increased, only a few low β-diverse experimental landscapes composed of highly multifunctional communities repeated many times were capable of producing high numbers of functions at high thresholds (Fig. 3). While the high multifunctionality of these particular low β diversity experimental landscapes drives this particular relationship, the aforementioned high functional variability of low β diversity experimental landscapes relative to high β diversity experimental landscapes is once again apparent in our analysis of multifunctionality thresholds here (Fig. 3).
This variability suggests that high β diversity gives rise to a multifunctional version of the portfolio effect (22) that eliminates the possibility of extremely high and low functioning experimental landscapes, but ensures a moderate level of multifunctionality that low β diversity experimental landscapes cannot guarantee. In the same way that species-rich communities are generally more successful at providing multiple functions due to functional complementarity, so too are β-diverse landscapes more reliable at providing moderate levels of multiple functions at larger scales. While ecosystem functioning is largely driven by species identity and richness at the local and landscape scale, β-diversity provides another level of insurance against loss of ecosystem functioning.
Since each plot in this study consists of a randomly assembled experimental community, our results may also be influenced by the assembly process of the original biodiversity experiment. Some experimental communities may have low functionality simply because they have species that are not well-suited to the general locale, a situation that is less likely to occur in natural communities structured by nonneutral assembly processes. As such, the effects of β diversity reported here are likely conservative relative to communities filtered through natural assembly processes and containing complementary suites of species better adapted to local conditions. While connectivity, dispersal, and succession may also affect landscape-level functionality in some ecosystems (23–25), we expect that most landscapes will possess few intercommunity species interactions relevant to the types of functions measured in this study, with the possible exception of those mediated by consumers that can move between communities (e.g., insect richness and abundance).
This analysis takes advantage of experimental precision, control, and replication not possible in observational studies of large landscapes. Nevertheless, the differences in species composition and functioning between communities in some landscapes, particularly at ecological boundaries, likely exceed those in this study. Thus, our results are also probably conservative because the original experimental design limited the range of α and γ diversities in these experimental landscapes. We expect that studies incorporating more realistic community structure with larger gradients in community composition will confirm and increase estimates of the distinct contributions of all three scales of biological diversity to ecosystem functioning.
This study highlights the importance of unique ecological and conservation considerations regarding the relationship between biodiversity and ecosystem functioning. Most importantly, analyses of individual functions and single scales of diversity underestimate the effects of biodiversity on ecosystem functioning. Our results confirm the importance of local species richness to ecosystem multifunctionality, and emphasize the neglected but important contributions of β and γ diversity as well.
Second, high multifunctionality is associated with high 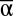 and high γ diversity, suggesting that both local species richness and larger scale diversity contribute to ecosystem functioning. Put differently, the experimental landscapes with the highest multifunctionality are those that are mainly composed of a suite of high α diversity communities. However, since landscapes with both high α and γ diversity are not always possible, it is important to consider the valuable role of β diversity to reduce functional variability in landscapes lacking high species diversity.
and high γ diversity, suggesting that both local species richness and larger scale diversity contribute to ecosystem functioning. Put differently, the experimental landscapes with the highest multifunctionality are those that are mainly composed of a suite of high α diversity communities. However, since landscapes with both high α and γ diversity are not always possible, it is important to consider the valuable role of β diversity to reduce functional variability in landscapes lacking high species diversity.
Our analyses reinforce earlier studies demonstrating the increased importance of species diversity for ecosystem functioning as more functional, temporal, and environmental contexts are considered (3–5). Moreover, they show that both local and regional diversity can simultaneously contribute to ecosystem multifunctionality at larger scales. While multifunctionality at larger scales was influenced by landscape-scale species richness, the effects of local species richness were even stronger, suggesting that the way species are organized into communities on landscapes heavily influences the effects of diversity on ecosystem functioning. As such, the processes that control the assembly of natural communities may be of central importance in relating the results of experimental studies of randomly assembled communities to nature.
Methods
We measured eight ecosystem functions (Fig. 1) using 1997, 1998, 1999, 2002, and 2006 data from the Cedar Creek Biodiversity II experiment in which 168 9 m × 9 m field plots were planted with randomized combinations of 1, 2, 4, 8, and 16 perennial grassland species (20). We created 7,512 simulated experimental landscapes, each composed of a constrained random selection of 24 plots, to create a broad and even distribution of α, β, and γ diversity (see SI Methods for details). Since there is no spatial component to these experimental landscapes, plots were selected independently of their spatial location in the original biodiversity experiment.
We assessed the ability of experimental landscapes to achieve several quantile-based thresholds of each function across all experimental landscapes and functions (6). We also calculated a unique multifunctionality metric (MF) as the mean of all functions minus its SD in each experimental landscape, where each function was scaled to the maximum observed among all experimental landscapes, and individual functions at the landscape scale were the sums of each function among its 24 experimental communities.
Within each experimental landscape, γ diversity was calculated as the number of unique species, and 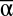 diversity as the average α of all 24 experimental communities. We calculated β using two separate metrics with contrasting response sensitivities. Results using the less sensitive metric reported in the text (β, the number of experimental communities in an experimental landscape that do not share exactly the same species composition) suggest similar but usually more conservative effects than results using our more sensitive metric (β, 1-Sorensen’s Index, Table S1 and S2).
diversity as the average α of all 24 experimental communities. We calculated β using two separate metrics with contrasting response sensitivities. Results using the less sensitive metric reported in the text (β, the number of experimental communities in an experimental landscape that do not share exactly the same species composition) suggest similar but usually more conservative effects than results using our more sensitive metric (β, 1-Sorensen’s Index, Table S1 and S2).
We used ordinary least squares regression to determine how the independent variables of 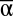 , β, and γ diversity influenced the dependent variables of each individual function, the number of functions achieved above each threshold, and MF. To compare the strength of effects and explanatory power of each independent variable, we calculated standardized regression weights and partial coefficients of determination (partial r2) for each diversity metric in multivariate models of each dependent variable.
, β, and γ diversity influenced the dependent variables of each individual function, the number of functions achieved above each threshold, and MF. To compare the strength of effects and explanatory power of each independent variable, we calculated standardized regression weights and partial coefficients of determination (partial r2) for each diversity metric in multivariate models of each dependent variable.
Standard transformations of independent variables did not improve regression diagnostics, or significantly improve model fit. Multicollinearity tolerances always exceeded 0.2, indicating that multicollinearity did not bias interpretations (26, 27). All three biodiversity metrics were included in all regressions reported and displayed in the main text and are the best models according to Akaike Information Criterion (AIC). We tested for landscape independence and correlation structure by regressing model residuals against each diversity metric. No correlation structure was detected for any regression and P values approached 1. Including diversity interaction terms in multiple regressions on multifunctionality increased overall model R2s by less than 10% relative to regressions with no interaction terms. Given the difficulties of interpreting interactions between continuous variables (28) and their limited explanatory power in this case, we do not report them. We performed all analyses using the R software package (29).
Supplementary Material
Acknowledgments
This work was supported by National Science Foundation Graduate Research Fellowships (to J.R.P. and T.L.).
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1220333110/-/DCSupplemental.
References
- 1.Balvanera P, et al. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol Lett. 2006;9(10):1146–1156. doi: 10.1111/j.1461-0248.2006.00963.x. [DOI] [PubMed] [Google Scholar]
- 2.Hooper DU, et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol Monogr. 2005;75(1):3–35. [Google Scholar]
- 3.Isbell F, et al. High plant diversity is needed to maintain ecosystem services. Nature. 2011;477(7363):199–202. doi: 10.1038/nature10282. [DOI] [PubMed] [Google Scholar]
- 4.Hector A, Bagchi R. Biodiversity and ecosystem multifunctionality. Nature. 2007;448(7150):188–190. doi: 10.1038/nature05947. [DOI] [PubMed] [Google Scholar]
- 5.Gamfeldt L, Hillebrand H, Jonsson PR. Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology. 2008;89(5):1223–1231. doi: 10.1890/06-2091.1. [DOI] [PubMed] [Google Scholar]
- 6.Zavaleta ES, Pasari JR, Hulvey KB, Tilman GD. Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proc Natl Acad Sci USA. 2010;107(4):1443–1446. doi: 10.1073/pnas.0906829107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Harrison S, Davies KF, Safford HD, Viers JH. Beta diversity and the scale-dependence of the productivity-diversity relationship: A test in the Californian serpentine flora. J Ecol. 2006;94(1):110–117. [Google Scholar]
- 8.Maestre FT, et al. Plant species richness and ecosystem multifunctionality in global drylands. Science. 2012;335(6065):214–218. doi: 10.1126/science.1215442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Anderson BJ, et al. Spatial covariance between biodiversity and other ecosystem service priorities. J Appl Ecol. 2009;46(4):888–896. [Google Scholar]
- 10.Chan KMA, Shaw MR, Cameron DR, Underwood EC, Daily GC. Conservation planning for ecosystem services. PLoS Biol. 2006;4(11):e379. doi: 10.1371/journal.pbio.0040379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Naidoo R, et al. Global mapping of ecosystem services and conservation priorities. Proc Natl Acad Sci USA. 2008;105(28):9495–9500. doi: 10.1073/pnas.0707823105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lavorel S, et al. Using plant functional traits to understand the landscape distribution of multiple ecosystem services. J Ecol. 2011;99(1):135–147. [Google Scholar]
- 13.Egoh B, Reyers B, Rouget M, Bode M, Richardson DM. Spatial congruence between biodiversity and ecosystem services in South Africa. Biol Conserv. 2009;142(3):553–562. doi: 10.1111/j.1523-1739.2009.01442.x. [DOI] [PubMed] [Google Scholar]
- 14.Nelson E, et al. Modeling multiple ecosystem services, biodiversity conservation, commodity production, and tradeoffs at landscape scales. Front Ecol Environ. 2009;7(1):4–11. [Google Scholar]
- 15.Millennium Ecosystem Assessment . Ecosystems and Human Well Being: Synthesis. Washington, DC: Island Press; 2005. [Google Scholar]
- 16.Gimona A, van der Horst D. Mapping hotspots of multiple landscape functions: A case study on farmland afforestation in Scotland. Landscape Ecol. 2007;22(8):1255–1264. [Google Scholar]
- 17.Naidoo R, Ricketts TH. Mapping the economic costs and benefits of conservation. PLoS Biol. 2006;4(11):e360. doi: 10.1371/journal.pbio.0040360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Aragon R, Oesterheld M, Irisarri G, Texeira M. Stability of ecosystem functioning and diversity of grasslands at the landscape scale. Landscape Ecol. 2011;26(7):1011–1022. [Google Scholar]
- 19.Holland RA, et al. Spatial covariation between freshwater and terrestrial ecosystem services. Ecol Appl. 2011;21(6):2034–2048. doi: 10.1890/09-2195.1. [DOI] [PubMed] [Google Scholar]
- 20.Tilman D, Reich PB, Knops JMH. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature. 2006;441(7093):629–632. doi: 10.1038/nature04742. [DOI] [PubMed] [Google Scholar]
- 21.Hooper DU, et al. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature. 2012;486:105–108. doi: 10.1038/nature11118. [DOI] [PubMed] [Google Scholar]
- 22.Tilman D, Lehman CL, Bristow CE. Diversity-stability relationships: Statistical inevitability or ecological consequence? Am Nat. 1998;151(3):277–282. doi: 10.1086/286118. [DOI] [PubMed] [Google Scholar]
- 23.Loreau M, Mouquet N, Gonzalez A. Biodiversity as spatial insurance in heterogeneous landscapes. Proc Natl Acad Sci USA. 2003;100(22):12765–12770. doi: 10.1073/pnas.2235465100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Cardinale BJ, Ives AR, Inchausti P. Effects of species diversity on the primary productivity of ecosystems: Extending our spatial and temporal scales of inference. Oikos. 2004;104(3):437–450. [Google Scholar]
- 25.Staddon P, Lindo Z, Crittenden PD, Gilbert F, Gonzalez A. Connectivity, non-random extinction and ecosystem function in experimental metacommunities. Ecol Lett. 2010;13(5):543–552. doi: 10.1111/j.1461-0248.2010.01450.x. [DOI] [PubMed] [Google Scholar]
- 26.Zar JH. Biostatistical Analysis. Upper Saddle River, NJ: Pearson; 2004. [Google Scholar]
- 27.Belsley DA, Kuh E, Welsch RE. Regression Diagnostics: Identifying Influential Data and Sources of Collinearity. New York: Wiley; 1980. [Google Scholar]
- 28.Aiken LS, West SG, Reno RR. Multiple Regression: Testing and Interpreting Interactions. Newbury Park, CA: Sage; 1991. [Google Scholar]
- 29.R Development Core Team . R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing; 2011. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.