

Abstract
AIM: To investigate the risk association of xeroderma pigmentosum group C (XPC) Lys939Gln polymorphism alone and in combination with cigarette smoking on colorectal cancer (CRC) predisposition.
METHODS: Peripheral blood samples of 510 study subjects (255 CRC patients, 255 controls)were collected. DNA was extracted and genotyping was performed using polymerase chain reaction-restriction fragment length polymorphism. The association between polymorphic genotype and CRC predisposition was determined using the OR and 95%CI.
RESULTS: The frequency of the homozygous variant (Gln/Gln) genotype was significantly higher in cases compared with controls (16.0% vs 10.2%, P = 0.049). The Gln/Gln genotype of XPC showed a significantly higher association with the risk of CRC (OR = 1.884; 95%CI: 1.082-3.277; P = 0.025). In the case of allele frequencies, variant allele C was associated with a significantly increased risk of CRC (OR = 1.375; 95%CI: 1.050-1.802; P = 0.020). Moreover, the risk was markedly higher for those who were carriers of the Gln/Gln variant genotype and were also cigarette smokers (OR = 3.409; 95%CI: 1.061-10.949; P = 0.032).
CONCLUSION: The XPC Gln/Gln genotype alone and in combination with smoking increases the risk of CRC among Malaysians.
Keywords: DNA repair, Xeroderma pigmentosum group C Lys939Gln polymorphism, Cigarette smoking, Colorectal cancer, Susceptibility risk
INTRODUCTION
Colorectal cancer (CRC) is a major public health problem worldwide, being the third most common cancer and the fourth most common cancer causing death[1]. In Malaysia, CRC has become the most common cancer among males and the second most common among females[2]. The development of CRC is a complex, multistep process involving interaction between environmental and genetic factors. Environmental factors such as dietary and lifestyle habits, smoking, alcohol consumption, and obesity interact with host’s genetic factors, especially genetic variations, and may modulate the risk of CRC[3]. Genetic variations, such as single nucleotide polymorphisms (SNPs), may increase the sensitivity to environmental carcinogens and may act as cancer predisposition factors. Environmentally sensitive genetic polymorphisms acting together with environmental factors are well documented candidates influencing cancer susceptibility.
DNA damage repair genes maintain the integrity of the genome against endogenous and exogenous factors. The xeroderma pigmentosum group C (XPC) gene is a DNA repair gene involved in the nucleotide excision repair (NER) mechanism which repairs bulky DNA lesions such as pyrimidine dimers, ultraviolet light-induced damage, photoproducts, intrastrand crosslinks, larger chemical adducts and other genotoxic agents[4]. Genetic variations in the XPC gene have been reported to modulate an individual’s susceptibility to developing cancer[5,6]. The XPC Lys939Gln polymorphism, which leads to an amino acid change from lysine to glutamine at codon 939, is the most common SNP studied in the XPC gene and has been shown to be associated with increased risk of several cancers such as skin[7], breast[8] and bladder cancers[9,10].
However, there are only limited reports on the association of this polymorphism with CRC susceptibility. A case-control study was undertaken in order to investigate the association of this polymorphism alone and also in conjunction with cigarette smoking on the risk of CRC. This polymorphism is believed to alter the gene expression and modulate the DNA repair function of the protein product, as it is located at the coding sequence of the XPC gene. Thus, we hypothesized that XPC Lys939Gln polymorphism may have an effect on modulating the susceptibility to CRC, and cigarette smoking may further enhance the effect on CRC risk.
MATERIALS AND METHODS
Study subjects
The study was approved by the Research Review Board and Ethics Committee of Universiti Sains Malaysia, Kelantan and the Ministry of Health, Malaysia. For this hospital-based case control study, subjects were recruited from various hospitals in Malaysia, including Hospital Universiti Sains Malaysia, Hospital Raja Perempuan Zainab II and Hospital Sultanah Bahiyah, Kedah, Malaysia. Genotyping was carried out at the Human Genome Center, Universiti Sains Malaysia. Two hundred and fifty five sporadic CRC patients and 255 healthy normal controls were recruited as study subjects. Cases were histopatologically confirmed sporadic CRC patients, aged > 25 years, who did not have previous colon/rectal or other cancers. Cases with known (as indicated in the pathology reports) familial adenomatous polyposis, ulcerative colitis or Crohn’s disease or any other previous malignancy were excluded. Controls were normal healthy individuals who were biologically unrelated to the study patients, aged > 25 years and had no history of cancer. Epidemiological data was collected from patients using a pre-structured questionnaire, which included sociodemographic status, physical status, dietary factors, occupation, tobacco/alcohol habits, previous illness, radiation exposure, etc.
Genotyping
Genotyping of XPC Lys939Gln polymorphism was carried out using polymerase chain reaction (PCR)-restriction fragment length polymorphism. Briefly, PCR primers for XPC Lys939Gln (F: 5’-GGCTTCCTGGTATCTGATTACT-3’R: 5’-CTCAGTTTGCCTTCTCAGCA-3’) were used to generate a 402 bp product containing the polymorphic site. The PCR reactions were carried out in a 25 µL volume of 1 × PCR Buffer, 2.0 mmol/L of MgCl2, 0.5 mmol/L dNTPs, 0.4 mmol/L of each primers and 1 U of AmpliTaq Gold Polymerase with a denaturation of 94 °C for 5 min, followed by 35 cycles at 94 °C for 30 s, 57 °C for 30 s, 72 °C for 30 s and finally 5 min at 72 °C. Following amplification, the PCR products were digested using PvuII restriction enzyme for 1 h at 37 °C and electrophoresed on 2% agarose gel. The homozygous wild-type genotype was identified by a single band at 402 bp level, the heterozygous genotype by 3 bands at 402, 276 and 126 bp levels and the homozygous variant by 2 bands at 276 and 126 bp levels (Figure 1).
Figure 1.
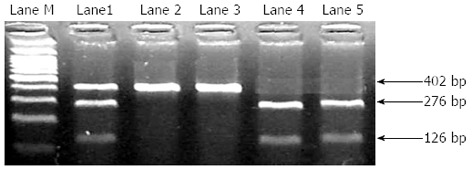
Gel picture showing the different categories of xeroderma pigmentosum group C Lys939Gln polymorphism genotype. Lane M: 100 bp DNA ladder; Lanes 2 and 3: Homozygous wildtype; Lane 1: Heterozygous; Lanes 4 and 5: Homozygous variant.
Statistical analysis
The sample size was calculated with power and sample size (PS) software version 2.1.31 using the uncorrected χ2 test with 80% power and 95%CI. The difference in distribution of genotypes, gender and age between cases and controls were assessed using the χ2 test. The ORs and 95%CI were calculated using binary logistic regression to evaluate the risk association. All statistical tests were two-sided, and statistical significance was determined as P < 0.05. SPSS v.18 (SPSS Inc., Chicago, IL, United States) was utilized for statistical analysis.
RESULTS
The demographic characteristics of the study subjects are shown in Table 1. This case-control study recruited 510 study subjects, including 255 histopathologically confirmed sporadic CRC patients and 255 healthy normal controls. Of the 255 CRC patients, 139 (54.5%) were male and 116 (45.5%) were female. Of the 255 normal controls, 115 (45.1%) were male and 140 (54.9%) were female. The ages of the sporadic CRC patients ranged from 27 to 93 years (mean age: 53.17 ± 7.07 years) and of the healthy normal controls ranged from 26 to 84 years (mean age: 46.47 ± 12.02 years).
Table 1.
Demographic characteristics of the study population n (%)
| Variable | Cases (n = 255 ) | Controls (n = 255) | P-value |
| Gender | |||
| Male | 139 (54.5) | 115 (45.1) | 0.0341 |
| Female | 116 (45.5) | 140 (54.9) | |
| Mean age (yr) | 53.17 ± 7.07 | 46.47 ± 12.02 | 0.086 |
| Cigarette smoking | |||
| Yes | 110 | 48 | < 0.0011 |
| No | 145 | 207 |
Statistically significant.
The frequencies of the homozygous wildtype (Lys/Lys), heterozygous (Lys/Gln) and homozygous variant (Gln/Gln) genotypes were 108 (42.4%), 106 (41.6%) and 41 (16.0%), respectively, among CRC cases. In controls, the frequencies were 129 (50.6%) for homozygous wildtype, 100 (39.2%) for heterozygous and 26 (10.2%) for homozygous variant genotypes. The frequency of the homozygous variant (CC) genotype was significantly higher in cases compared with the controls (P = 0.049). Table 2 shows the genotype and allele frequencies of XPC Lys939Gln polymorphism in cases and controls.
Table 2.
Genotype and allele frequencies of the xeroderma pigmentosum group CLys939Gln polymorphism in study subjects n (%)
| Cases (n = 255) | Controls (n = 255) | P-value | |
| Genotype | |||
| Homozygous wildtype (AA) | 108 (42.4) | 129 (50.6) | 0.062 |
| Heterozygous (AC) | 106 (41.6) | 100 (39.2) | 0.588 |
| Homozygous variant (CC) | 41 (16.0) | 26 (10.2) | 0.0491 |
| Allele | |||
| A | 322 (63.1) | 358 (70.0) | 0.294 |
| C | 188 (36.9) | 152 (30.0) |
Statistically significant.
Binary logistic regression analysis was performed in order to find the risk association. Table 3 shows the association between the XPC Lys939Gln polymorphism and risk of CRC. It can be clearly seen that the homozygous variant genotype was significantly associated with increased risk of CRC (OR = 1.884; 95%CI: 1.082-3.277; P = 0.025). The variant allele C was found to be significantly associated with increased risk of CRC (OR = 1.375; 95%CI: 1.050-1.802; P = 0.020). Furthermore, the study subjects were stratified into smokers and non-smokers and the risk association was evaluated. Results showed that, for those with the homozygous variant genotype (CC) and who were smokers, the risk was significantly increased (OR = 3.409; 95%CI: 1.061-10.949; P = 0.032). In the non-smoking group, no significant association was observed between the XPC Lys939Gln polymorphism and the risk of CRC.
Table 3.
Xeroderma pigmentosum group C Lys939Gln polymorphism, smoking and colorectal cancer susceptibility risk
| Cases | Controls | OR (95%CI) | P-value | |
| Genotype | ||||
| Lys/Lys (AA) | 108 | 129 | 1 (Reference)1 | |
| Lys/Gln (AC) | 106 | 100 | 1.266 (0.871–1.841) | 0.216 |
| Gln/Gln (CC) | 41 | 26 | 1.884 (1.082–3.277) | 0.0252 |
| Allele | ||||
| A | 322 | 358 | 1 (Reference)1 | - |
| C | 188 | 152 | 1.375 (1.050–1.802) | 0.0202 |
| Non-smoker | ||||
| AA | 64 | 104 | 1 (Reference)1 | |
| AC | 64 | 81 | 1.284 (0.817–2.018) | 0.278 |
| CC | 17 | 22 | 1.256 (0.620–2.542) | 0.526 |
| Smoker | ||||
| AA | 44 | 25 | 1 (Reference)1 | |
| AC | 42 | 19 | 1.256 (0.605–2.609) | 0.541 |
| CC | 24 | 4 | 3.409 (1.061–10.949) | 0.0322 |
Homozygous wildtype genotype served as the reference category;
Statistically significant.
DISCUSSION
To date, a variety of chemical carcinogens have been identified to cause DNA damage in humans. DNA damage repair genes are responsible for maintaining the integrity of the human genome through different pathways, locations and types of damage by base excision repair, NER, double strand break and mismatch repair pathways[4]. Genes involved in the NER pathway, such as XPC, commonly repair lesions induced by numerous exogenous agents such as those derived from food and smoking, including 2-amino-1-methyl-6-phenylimidazo-[4,5-β]-pyridine and benzo-[α]-pyrenediol-epoxide[11].
The XPC gene, located on chromosome 3p25, contains 16 exons and 15 introns and encodes a 940 amino acid protein[12]. Several polymorphic variants in the XPC gene have been identified and XPC Lys939Gln is one of the three most common SNPs. This case-control study investigated the genotype and allele frequencies of the XPC Lys939Gln polymorphism, and the risk association with sporadic CRC susceptibility. Results from the present study showed that the frequency of the XPC homozygous variant genotype was significantly higher in cases compared with controls (16.0% vs 10.2%, P = 0.049). On evaluation of the risk association, the homozygous variant genotype (CC) was associated with significantly increased risk of CRC (OR = 3.409; P = 0.032). This result conforms with a study by Yasuda et al[13], which demonstrated that a single amino acid alteration could be sufficient to compromise XPC function, thereby enhancing the risk of CRC development. Our results are in agreement with a few other reports by Qiu et al[5], Zhang et al[6] and Gil et al[14] which revealed that XPC is one the most important genes modulating an individual’s risk of developing sporadic cancer.
Carcinogens consumed through dietary and lifestyle habits as well as from the environment play a major role in damaging DNA. Cigarette smoking is one of the lifestyle factors that play an important role in the exposure of an individual to the carcinogens present in cigarette smoke. Carcinogens contained in tobacco smoke, such as polycyclic aromatic hydrocarbons, heterocyclic amines, nitrosamines and aromatic amines, are harmful to the human colon and rectum. Cigarette smoking has been reported to lead to the formation of DNA adducts and cause damage to the DNA in the human colon[15,16]. All these carcinogens can reach the colorectal mucosa, through direct ingestion or via the circulatory system, and has been demonstrated to induce bulky adducts in crypt cells and contribute to the formation of mutations in the colon[17,18]. In the present study, a combination of smoking and a variant genotype of XPC enhanced the risk of CRC 3-fold, indicating that smoking further enhanced the risk of CRC. Although non-smokers showed an OR of 1.256, this was not statistically significant.
The XPC gene encodes an important protein involved in the recognition of DNA damage in the NER pathway. This protein binds to HR23B protein to form XPC-HR23B that recognizes a DNA lesion and repairs the DNA damage along with other DNA repair proteins[19,20]. Polymorphism in the coding sequence of the XPC gene has been demonstrated to alter the gene expression and thereby modulate the DNA repair function[21]. The XPC Lys939Gln polymorphism is located in the coding sequence of the XPC gene. The nucleotide change from A to C leads to an amino acid change from lysine to glutamine in the coding sequence of the XPC gene and has been reported to lead to reduced repair capacity. This genetic variation has also been reported to result in reduced specificity of this gene in recognition and repair of the DNA damage as well as in protein expression, thus allowing more somatic DNA mutations or alterations to occur[21,22].
The XPC protein plays a crucial role in repairing the DNA damage caused by tobacco smoke. Individuals with the XPC variant genotype may possess deficient DNA repair capability. Accordingly, the XPC protein product may be less efficient in repairing the DNA lesions induced by tobacco smoke, and thereby could enhance the susceptibility, favoring the development of CRC.
Previous studies focusing on the role of the XPC variant in the modulation of an individual’s risk of developing CRC are scarce, and reported results are inconsistent. Berndt et al[22], in an American population, found an OR of 1.19 for the variant genotype’s association with risk of CRC, but the result was statistically insignificant (P = 0.50). In contrast, a study conducted by Hansen et al[23], in a Polish population, observed no significant risk association between this polymorphism and CRC susceptibility. However, when the variant genotype in conjunction with red meat consumption was evaluated, the risk was reported to be significantly higher (OR = 3.7) in those who carried the homozygous variant (CC) genotype and consumed 100 g red meat per day.
A recent study by Wu et al[24] on 421 CRC patients and 845 controls, showed that the homozygous variant (CC) genotype was significantly associated with higher risk of CRC in a Chinese population (OR =1.5; 95%CI: 1.0-2.2; P = 0.035). Moreover, combination (AC + CC) genotypes in the homozygous wildtype model also showed a significant association with risk of CRC (OR = 1.3; 95%CI: 1.0-1.7; P = 0.039). When the study subjects were stratified into tea drinkers and non-tea drinkers, the results showed that individuals who were non-tea drinkers and carried combination (AC + CC) genotypes, had increased risk of CRC (OR = 3.0; 95%CI: 1.9-4.7; P < 0.001).
Apart from CRC, XPC polymorphism was reported to be associated with the risk of other cancers, such as head and neck[25], lung[10], breast[8] and bladder[26]. In addition, abnormal expression of the XPC gene has been shown in hepatocarcinogenesis[27]. The variant allele of the XPC gene is associated with lower DNA repair capacity (DRC). Several studies showed that the Lys939Gln polymorphism had an interaction with other SNPs located in the same gene, such as Ala499Val, PAT (-/+) and IVS11 C/A polymorphisms. According to Khan et al[28], the XPC Lys939Gln polymorphism is in linkage disequilibrium with the XPC PAT polymorphism as well as the C/A polymorphism at 25 positions of intron 11 of XPC. This causes exon 12 to be skipped and deleted, resulting in the loss of XPC cDNA function, thus reducing the DNA repair activity.
Researchers have shown that carriers of the C variant allele of the XPC Lys939Gln polymorphism exhibited significantly more DNA damage induced by BPDE[21,29,30]. Moreover, combinations of haplotype C + C (Lys939Gln, PAT +, Ala499Val) have been reported to cause poor DRC, thus increasing the risk of CRC. In contrast, haplotype T-A (Ala499Val-PAT-Lys939Gln) has been demonstrated to be associated with the least DNA damage[21,24]. Also, another study has shown that accumulation or the presence of alleles in the XPC gene associated with increased risk might reduce DRC and thus increase CRC susceptibility[31].
The difference in results on the risk association between the present study and other previous studies might be explained by differences in genetic background of study subjects groups or populations, and also differences in exposure to environmental and lifestyle factors. Small sample sizes and/or inadequate control for certain confounder factors such as gender and age might also have contributed to the differing results and lack of association.
From the present study, it is reasonable to suggest that the XPC gene, especially the XPC variant genotype, may modulate the DRC of the host cell and thus play an important role in sporadic colorectal carcinogenesis. The risk could be much higher in individuals who possess the variant genotype and who are also cigarette smokers. The results also highlight the potential role of the NER pathway (especially of XPC) in modulation of an individual’s risk of sporadic CRC. Further studies to explore the interaction of XPC Lys939Gln with other SNPs of the XPC gene and with other genes involved in DNA repair pathways, either singly or in combination, and to examine the correlation with environmental factors such as alcohol consumption and dietary habits, as well as clinicopathological characteristics, would be beneficial in deriving more accurate risk predictive markers.
ACKNOWLEDGMENTS
We would like to thank Dr. Muhammad Radzi Abu Hassan (Hospital Sultanah Bahiyah, Kedah), Prof Dr. BM Biswal, Dr. VMK Bhavaraju (Hospital Universiti Sains Malaysia), Dr. Ahmad Shanwani and Dr. Zaidi Zakaria (Hospital Raja Perempuan Zainab II, Kota Bharu) for their cooperation in patient recruitment.
COMMENTS
Background
Colorectal cancer (CRC) which is the most common cancer among males and the second most common among females in Malaysia, has become a major public problem. Environmental factors, such as dietary carcinogens and tobacco smoke, interacting with the host’s genetic factors may modulate the risk of CRC. However, the genetic predisposition factors associated with colorectal carcinogenesis remain largely undetermined. Identification of the host’s genetic predisposition factors may help in understanding the carcinogenic process. Thus, it was of interest to explore the contribution of single nucleotide polymorphisms (SNPs) in a DNA repair gene, alone and in combination with tobacco smoke, as a predisposition factor for CRC susceptibility.
Research frontiers
Exposure to mutagens and carcinogens through diet, tobacco smoke,etc can cause varying degrees of DNA damage and can cause mutations in humans if left unrepaired. The xeroderma pigmentosum group C (XPC) gene is a DNA repair gene involved in the nucleotide excision repair (NER) pathway. The SNP, Lys939Gln, located at the coding region of the XPC gene has been associated with lower DNA repair capacity and has been shown to be associated with increased risk of cancers of the skin, breast and bladder. This study was designed to determine the frequencies and influence of the XPC Lys939Gln polymorphic genotype, alone and in combination with smoking, on sporadic CRC susceptibility risk in a Malaysian population.
Innovations and breakthroughs
Only limited reports are available on the association of XPC Lys939Gln polymorphism with CRC susceptibility risk. To the best of our knowledge, this is the first report of an association between the genetic variant of XPC with CRC susceptibility risk in the Malaysian population. The observation that genetic variation of the XPC gene influences susceptibility to CRC implies the importance of the NER pathway in modulation of an individual’s risk of CRC. This study provides support for the hypothesis that genetic variation in the XPC gene, acting together with environmental factors, contributes to CRC development and may be considered as a candidate marker for CRC predisposition risk in the Malaysian population.
Applications
The present study observed that the genetic variation Lys939Gln of the XPC gene influences the risk of CRC in the Malaysian population. Further SNP mapping studies utilizing high throughout genotyping methods could facilitate the analysis of multiple polymorphisms within DNA repair genes. Genotyping of Malaysian CRC patients for polymorphism(s) in DNA repair genes will help in understanding the specific polymorphism(s), which act as predisposing genotype(s) in CRC susceptibility. In future, the study can be extended to a population level, to identify individuals with high risk predisposition genotypes. Appropriate surveillance programs could then be developed in order to reduce the morbidity and mortality of CRC at the population level.
Peer review
This case-control study investigated the association between SNPs and cigarette smoking on sporadic CRC susceptibility risk in Malaysian population. The genotype and allele frequencies of XPC Lys939Gln were investigated in 255 CRC patients and 255 healthy controls. Homozygous variant (CC) genotype frequency was found to be significantly higher in CRC cases compared to controls. When the risk association was evaluated singly, the homozygous variant XPC CC genotype was associated with an increased risk of CRC susceptibility. Moreover, when the risk was evaluated after stratifying the study subjects into smokers and non-smokers, the combination of smoking habits and variant genotype of XPC was found to enhance the CRC susceptibility risk to 3-fold times higher. These results prompt us to suggest that genetic variation Lys939Gln in XPC gene might modify the effect of smoking and contribute to sporadic colorectal cancer etiology.
Footnotes
Supported by Fundamental Research Grant Scheme (FRGS), No. 203/PPSP/6171112
P- Reviewer Gusella M S- Editor Wen LL L- Editor Cant MR E- Editor Zhang DN
References
- 1.Rim HJ, Kim SJ, Sun IJ, Lee JS. [Antigenic localities in the tissues of Paragonimus westermani by developmental stages using immunogold labeling method] Kisaengchunghak Chapchi. 1992;30:1–14. doi: 10.3347/kjp.1992.30.1.1. [DOI] [PubMed] [Google Scholar]
- 2.Malaysia Cancer Statistics-Data and Figure Peninsular Malaysia (2006) National Cancer Registry. Ministry of Health Malaysia. Available from: http://www.makna.org.my/PDF/MalaysiaCancerStatistics.pdf.
- 3.Naccarati A, Pardini B, Hemminki K, Vodicka P. Sporadic colorectal cancer and individual susceptibility: a review of the association studies investigating the role of DNA repair genetic polymorphisms. Mutat Res. 2007;635:118–145. doi: 10.1016/j.mrrev.2007.02.001. [DOI] [PubMed] [Google Scholar]
- 4.Christmann M, Tomicic MT, Roos WP, Kaina B. Mechanisms of human DNA repair: an update. Toxicology. 2003;193:3–34. doi: 10.1016/s0300-483x(03)00287-7. [DOI] [PubMed] [Google Scholar]
- 5.Qiu L, Wang Z, Shi X, Wang Z. Associations between XPC polymorphisms and risk of cancers: A meta-analysis. Eur J Cancer. 2008;44:2241–2253. doi: 10.1016/j.ejca.2008.06.024. [DOI] [PubMed] [Google Scholar]
- 6.Zhang D, Chen C, Fu X, Gu S, Mao Y, Xie Y, Huang Y, Li Y. A meta-analysis of DNA repair gene XPC polymorphisms and cancer risk. J Hum Genet. 2008;53:18–33. doi: 10.1007/s10038-007-0215-5. [DOI] [PubMed] [Google Scholar]
- 7.Blankenburg S, König IR, Moessner R, Laspe P, Thoms KM, Krueger U, Khan SG, Westphal G, Berking C, Volkenandt M, et al. Assessment of 3 xeroderma pigmentosum group C gene polymorphisms and risk of cutaneous melanoma: a case-control study. Carcinogenesis. 2005;26:1085–1090. doi: 10.1093/carcin/bgi055. [DOI] [PubMed] [Google Scholar]
- 8.Sugisawa H, Nakamura R, Nakano I, Sugisawa A. [Four-year follow-up study of self-rated health and life satisfaction among caregivers] Nihon Koshu Eisei Zasshi. 1992;39:22–32. [PubMed] [Google Scholar]
- 9.García-Closas M, Malats N, Real FX, Welch R, Kogevinas M, Chatterjee N, Pfeiffer R, Silverman D, Dosemeci M, Tardón A, et al. Genetic variation in the nucleotide excision repair pathway and bladder cancer risk. Cancer Epidemiol Biomarkers Prev. 2006;15:536–542. doi: 10.1158/1055-9965.EPI-05-0749. [DOI] [PubMed] [Google Scholar]
- 10.Sanyal S, Festa F, Sakano S, Zhang Z, Steineck G, Norming U, Wijkström H, Larsson P, Kumar R, Hemminki K. Polymorphisms in DNA repair and metabolic genes in bladder cancer. Carcinogenesis. 2004;25:729–734. doi: 10.1093/carcin/bgh058. [DOI] [PubMed] [Google Scholar]
- 11.Tyson J, Mathers JC. Dietary and genetic modulation of DNA repair in healthy human adults. Proc Nutr Soc. 2007;66:42–51. doi: 10.1017/S0029665107005289. [DOI] [PubMed] [Google Scholar]
- 12.Li L, Peterson C, Legerski R. Sequence of the mouse XPC cDNA and genomic structure of the human XPC gene. Nucleic Acids Res. 1996;24:1026–1028. doi: 10.1093/nar/24.6.1026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Yasuda G, Nishi R, Watanabe E, Mori T, Iwai S, Orioli D, Stefanini M, Hanaoka F, Sugasawa K. In vivo destabilization and functional defects of the xeroderma pigmentosum C protein caused by a pathogenic missense mutation. Mol Cell Biol. 2007;27:6606–6614. doi: 10.1128/MCB.02166-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Gil J, Ramsey D, Stembalska A, Karpinski P, Pesz KA, Laczmanska I, Leszczynski P, Grzebieniak Z, Sasiadek MM. The C/A polymorphism in intron 11 of the XPC gene plays a crucial role in the modulation of an individual’s susceptibility to sporadic colorectal cancer. Mol Biol Rep. 2012;39:527–534. doi: 10.1007/s11033-011-0767-5. [DOI] [PubMed] [Google Scholar]
- 15.Alexandrov K, Rojas M, Kadlubar FF, Lang NP, Bartsch H. Evidence of anti-benzo[a]pyrene diolepoxide-DNA adduct formation in human colon mucosa. Carcinogenesis. 1996;17:2081–2083. doi: 10.1093/carcin/17.9.2081. [DOI] [PubMed] [Google Scholar]
- 16.Giovannucci E. Alcohol, one-carbon metabolism, and colorectal cancer: recent insights from molecular studies. J Nutr. 2004;134:2475S–2481S. doi: 10.1093/jn/134.9.2475S. [DOI] [PubMed] [Google Scholar]
- 17.Stern MC, Conti DV, Siegmund KD, Corral R, Yuan JM, Koh WP, Yu MC. DNA repair single-nucleotide polymorphisms in colorectal cancer and their role as modifiers of the effect of cigarette smoking and alcohol in the Singapore Chinese Health Study. Cancer Epidemiol Biomarkers Prev. 2007;16:2363–2372. doi: 10.1158/1055-9965.EPI-07-0268. [DOI] [PubMed] [Google Scholar]
- 18.Wu X, Zhao H, Suk R, Christiani DC. Genetic susceptibility to tobacco-related cancer. Oncogene. 2004;23:6500–6523. doi: 10.1038/sj.onc.1207811. [DOI] [PubMed] [Google Scholar]
- 19.Sugasawa K. XPC: its product and biological roles. Adv Exp Med Biol. 2008;637:47–56. doi: 10.1007/978-0-387-09599-8_6. [DOI] [PubMed] [Google Scholar]
- 20.Volker M, Moné MJ, Karmakar P, van Hoffen A, Schul W, Vermeulen W, Hoeijmakers JH, van Driel R, van Zeeland AA, Mullenders LH. Sequential assembly of the nucleotide excision repair factors in vivo. Mol Cell. 2001;8:213–224. doi: 10.1016/s1097-2765(01)00281-7. [DOI] [PubMed] [Google Scholar]
- 21.Zhu Y, Yang H, Chen Q, Lin J, Grossman HB, Dinney CP, Wu X, Gu J. Modulation of DNA damage/DNA repair capacity by XPC polymorphisms. DNA Repair (Amst) 2008;7:141–148. doi: 10.1016/j.dnarep.2007.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Berndt SI, Platz EA, Fallin MD, Thuita LW, Hoffman SC, Helzlsouer KJ. Genetic variation in the nucleotide excision repair pathway and colorectal cancer risk. Cancer Epidemiol Biomarkers Prev. 2006;15:2263–2269. doi: 10.1158/1055-9965.EPI-06-0449. [DOI] [PubMed] [Google Scholar]
- 23.Hansen RD, Sørensen M, Tjønneland A, Overvad K, Wallin H, Raaschou-Nielsen O, Vogel U. XPA A23G, XPC Lys939Gln, XPD Lys751Gln and XPD Asp312Asn polymorphisms, interactions with smoking, alcohol and dietary factors, and risk of colorectal cancer. Mutat Res. 2007;619:68–80. doi: 10.1016/j.mrfmmm.2007.02.002. [DOI] [PubMed] [Google Scholar]
- 24.Wu Y, Jin M, Liu B, Liang X, Yu Y, Li Q, Ma X, Yao K, Chen K. The association of XPC polymorphisms and tea drinking with colorectal cancer risk in a Chinese population. Mol Carcinog. 2011;50:189–198. doi: 10.1002/mc.20704. [DOI] [PubMed] [Google Scholar]
- 25.Shen H, Sturgis EM, Khan SG, Qiao Y, Shahlavi T, Eicher SA, Xu Y, Wang X, Strom SS, Spitz MR, et al. An intronic poly (AT) polymorphism of the DNA repair gene XPC and risk of squamous cell carcinoma of the head and neck: a case-control study. Cancer Res. 2001;61:3321–3325. [PubMed] [Google Scholar]
- 26.Fontana L, Bosviel R, Delort L, Guy L, Chalabi N, Kwiatkowski F, Satih S, Rabiau N, Boiteux JP, Chamoux A, et al. DNA repair gene ERCC2, XPC, XRCC1, XRCC3 polymorphisms and associations with bladder cancer risk in a French cohort. Anticancer Res. 2008;28:1853–1856. [PubMed] [Google Scholar]
- 27.Fautrel A, Andrieux L, Musso O, Boudjema K, Guillouzo A, Langouët S. Overexpression of the two nucleotide excision repair genes ERCC1 and XPC in human hepatocellular carcinoma. J Hepatol. 2005;43:288–293. doi: 10.1016/j.jhep.2005.02.020. [DOI] [PubMed] [Google Scholar]
- 28.Khan SG, Muniz-Medina V, Shahlavi T, Baker CC, Inui H, Ueda T, Emmert S, Schneider TD, Kraemer KH. The human XPC DNA repair gene: arrangement, splice site information content and influence of a single nucleotide polymorphism in a splice acceptor site on alternative splicing and function. Nucleic Acids Res. 2002;30:3624–3631. doi: 10.1093/nar/gkf469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Rzeszowska-Wolny J, Polanska J, Pietrowska M, Palyvoda O, Jaworska J, Butkiewicz D, Hancock R. Influence of polymorphisms in DNA repair genes XPD, XRCC1 and MGMT on DNA damage induced by gamma radiation and its repair in lymphocytes in vitro. Radiat Res. 2005;164:132–140. doi: 10.1667/rr3400. [DOI] [PubMed] [Google Scholar]
- 30.Vodicka P, Kumar R, Stetina R, Sanyal S, Soucek P, Haufroid V, Dusinska M, Kuricova M, Zamecnikova M, Musak L, et al. Genetic polymorphisms in DNA repair genes and possible links with DNA repair rates, chromosomal aberrations and single-strand breaks in DNA. Carcinogenesis. 2004;25:757–763. doi: 10.1093/carcin/bgh064. [DOI] [PubMed] [Google Scholar]
- 31.Hu Z, Wang Y, Wang X, Liang G, Miao X, Xu Y, Tan W, Wei Q, Lin D, Shen H. DNA repair gene XPC genotypes/haplotypes and risk of lung cancer in a Chinese population. Int J Cancer. 2005;115:478–483. doi: 10.1002/ijc.20911. [DOI] [PubMed] [Google Scholar]