

Abstract
Reading speed in normal peripheral vision is slow but can be increased through training on a letter-recognition task. The aim of the present study is to investigate the sensory and cognitive factors responsible for this improvement. The visual span is hypothesized to be a sensory bottleneck limiting reading speed. Three sensory factors—letter acuity, crowding, and mislocations (errors in the spatial order of letters)—may limit the size of the visual span. Reading speed is also influenced by cognitive factors including the utilization of information from sentence context. We conducted a perceptual training experiment to investigate the roles of these factors. Training consisted of four daily sessions of trigram letter-recognition trials at 10° in the lower visual field. Subjects' visual-span profiles and reading speeds were measured in pre- and posttests. Effects of the three sensory factors were isolated through a decomposition analysis of the visual span profiles. The impact of sentence context was indexed by context gain, the ratio of reading speeds for ordered and unordered text. Following training, visual spans increased in size by 5.4 bits of information transmitted, and reading speeds increased by 45%. Training induced a substantial reduction in the magnitude of crowding (4.8 bits) and a smaller reduction for mislocations (0.7 bits), but no change in letter acuity or context gain. These results indicate that the basis of the training-related improvement in reading speed is a large reduction in the interfering effect of crowding and a small reduction of mislocation errors.
Keywords: reading, peripheral vision, visual span, crowding, context gain
Introduction
Previous research has shown that reading speed in peripheral vision increases following practice on a letter-recognition task (Chung, Legge, & Cheung, 2004; Yu, Legge, Park, Gage, & Chung, 2010). The primary focus of this paper is to evaluate the sensory and cognitive bases for this improvement in reading speed.
Reading is crucial to full participation in modern society. For most people with low vision, reading is often slow and difficult. This is especially true for people with central-field loss from macular degeneration who must rely on their peripheral vision to read. Research with normally-sighted subjects has shown that even when the text is appropriately scaled to compensate for decreasing peripheral acuity, reading speed decreases by a factor of six from fovea to 20° in the lower visual field (Chung, Mansfield, & Legge, 1998). These facts motivated our interest in peripheral reading.
Reading speed can be influenced by sensory, oculomotor, and cognitive/linguistic factors (Legge et al., 2007). In previous research (Chung et al., 2004; Yu et al., 2010), the influence of oculomotor factors was minimized by utilizing a Rapid Serial Visual Presentation (RSVP) paradigm (Forster, 1970; Rubin & Turano, 1992). In the present study, the influence of sensory factors was explored by investigating the visual-span hypothesis. According to this hypothesis, an important factor limiting reading speed is the size of the visual span—the number of text letters that can be recognized accurately without eye movements (Legge, Mansfield, & Chung, 2001). Legge and colleagues (2001) measured visual span size and reading speed at different retinal eccentricities and found a correlation between reading speed and the size of the visual span. Accordingly, slow reading in peripheral vision maybe due, at least in part, to a reduced visual span.
Some simple forms of visual discrimination improve in peripheral vision with practice, a phenomenon termed visual “perceptual learning” (for a review, see Fine & Jacobs, 2002). Chung et al., (2004) examined the possibility that peripheral reading speed could be improved by enlarging the visual span through practice. If so, perceptual learning might be useful as a form of reading rehabilitation for people with macular degeneration. They showed that four daily practice sessions on a letter-recognition task with trigrams (three horizontally-adjacent letters) in normal peripheral vision resulted in an expanded visual span, accompanied by a mean increase of 41% in reading speed in the trained field. Follow-up studies have confirmed that trigram training can enhance the peripheral reading speed of normally sighted subjects from 40% to 83.5% (Lee, Kwon, Legge, & Gefroh, 2010, computed from their table C1; Yu et al., 2010).
What sensory limitations determine the size of the visual span? Three factors have been proposed (Legge, 2007) —letter acuity, crowding, and mislocations (errors in the spatial order of letters). It remains unclear which of these factors change due to training and is responsible for the enlargement of the visual span. Pelli et al. (2007) have argued that crowding is the dominant factor determining the size of the visual span (which they called the “uncrowded span”) and that reading rate is proportional to the uncrowded span. If they are right, it seems likely that the enlargement of the visual span (and corresponding improvement in reading speed) is due to a training-related reduction in crowding. To test this possibility, we conducted a training experiment, using methods similar to those of Chung et al. (2004), followed by a decomposition analysis to isolate the contributions from acuity, crowding, and mislocations (see Method). Although our main focus here is on the factors affecting the spatial extent of the visual span, we note the influence of temporal factors, an issue to be considered in the Discussion below.
Cognitive and linguistic factors also influence reading speed. In particular, the effect of sentence context has been studied by comparing reading speed for meaningful sentences with reading speed for “sentences” with scrambled word order, termed “unordered text” (Sass, Legge, & Lee, 2006; Pelli et al., 2007; Morton, 2008). Pelli et al. (2007) showed that the context gain (defined as the ratio of reading speeds for ordered and unordered text) was much larger in peripheral vision than in central vision. The authors claimed that since crowding impairs the access to word content, the readers would rely more on word order in peripheral vision. Pelli and Tillman (2007) found a triple dissociation between the influence of letter recognition, word shape and context on reading speed, with each of the three processes functioning independently of the other two and contributing a constant number of words per minute to the total reading speed. Therefore, it is possible that part of the training-related improvement of reading speed resulted from better utilization of context information. To examine how the training with trigrams would affect the access to word content and word order, both ordered and unordered text were used in our study to measure reading speed.
To summarize, this study aims to evaluate the sensory and cognitive bases for the improvement of reading speed after perceptual training on a letter-recognition task. The goal of this study is to better understand the relationship between visual span, context and reading speed, and furthermore to develop a more effective training paradigm to improve reading in peripheral vision.
Method
Subjects
Twelve native English-speaking students with normal vision were recruited from the University of Minnesota with six assigned at random to each of two groups: a training group and a control group. Before the main experiment, binocular acuity (Lighthouse Near Acuity Chart, Lighthouse Low Vision Products, Long Island City, NY) and reading performance (MNRead, Lighthouse Low Vision Products) were measured for each subject. Group characteristics are shown in Table 1. All of the subjects were naïve to the tests in this study and signed an Institutional Review Board-approved consent form prior to the experiments.
Table 1.
Subject groups (mean ± STD).
|
|
Training |
Control |
| Gender ratio (M:F) | 4:2 | 3:3 |
| Age | 19.6 ± 2.1 | 22.4 ± 2.1 |
| Visual acuity (LogMAR) | −0.07 ± 0.05 | −0.07 ± 0.03 |
| MNREAD | ||
| Reading acuity (LogMAR) | −0.45 ± 0.06 | −0.28 ± 0.02 |
| Critical print size (LogMAR) | 0.17 ± 0.08 | 0.22 ± 0.14 |
| Maximum reading speed (wpm) | 251 ± 28 | 230 ± 4 |
Apparatus and stimuli
The stimuli were generated and presented using MATLAB R2010a with Psychophysics Toolbox (Brainard, 1997; Pelli, 1997). The stimuli were black letters, trigrams or words on a white background (luminance of 84 cd/m2, Weber contrast >99%), presented in the Courier font. The viewing distance was 40 cm. A letter size (defined as the height of a lowercase “x”) of 3° was used because it exceeds the critical print size at 10° in peripheral vision (Chung et al., 1998). A chinrest was used to stabilize the subject's head to ensure a constant viewing distance, and a webcam was used to monitor subjects' fixation on every trial. A previous study has shown that saccades of 2° can be reliably detected by a similar method (Cheong, Legge, Lawrence, Cheung, & Ruff, 2007). Trials were cancelled when fixation was not maintained. The stimuli were displayed on an NEC MultiSync CRT monitor (model: FP2141SB-BK; refresh rate: 100 Hz; resolution: 1280 × 1024, with 71 pixels per x-height; NEC-Mitsubishi Electric Visual System Cooperation, Tokyo, Japan), controlled by a Mac Pro Quad-Core computer (model No.: A1186; Apple, Inc., Cupertino, CA).
Experimental design
The experiment contained three parts: pretest, training, and posttest, as illustrated in Figure 1. In pre- and posttests, subjects' visual span profiles were measured with a trigram letter-recognition task, and their reading speeds were measured with an RSVP reading task. In addition, we measured the visual span profiles with single letters in order to conduct the decomposition analysis (see below). The sequence of these tests was single-letter visual span measurement (four blocks), trigram visual span measurement (eight blocks), and RSVP reading task with ordered text (four blocks) and with unordered text (four blocks). Half of the blocks were tested at 10° in the upper visual field and half at 10° in the lower visual field, and the upper and lower blocks were interleaved. The order of the two visual fields, i.e., upper-lower or lower-upper, was kept the same in pre- and posttests for each subject but counterbalanced between subjects. Each daily training session contained sixteen blocks of trigram visual span measurements at 10° in the lower visual field. Only the training group participated in the training sessions while the control group had no training during the corresponding time.
Figure 1.
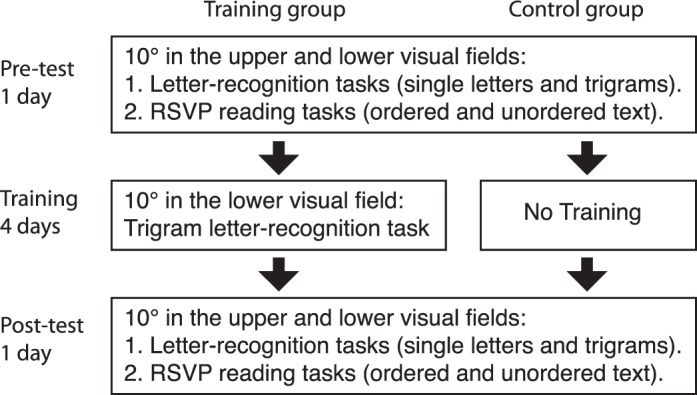
Schematic diagram of the experiment sequence.
Visual span measurement
Figure 2 illustrates the visual span measurement. Subjects were asked to fixate on a green dot in the center of the screen and recognize single letters or trigrams on a horizontal line 10° in the upper or lower visual field. The stimuli (trigrams or single letters) were located at eleven different slots along this horizontal line, determined by normal letter spacing in the Courier font (center-to-center spacing = 3.48°). The slot at the midline is labeled position 0. The left slots are labeled with negative numbers (−5 to −1) and the right slots with positive numbers (1 to 5). In each trial, a letter or a trigram was presented for 200 ms. Subjects reported the letter or the trigram and the experimenter recorded their answer. Each block consisted of 55 trials, with stimuli centered on each of the 11 slots five times.
Figure 2.
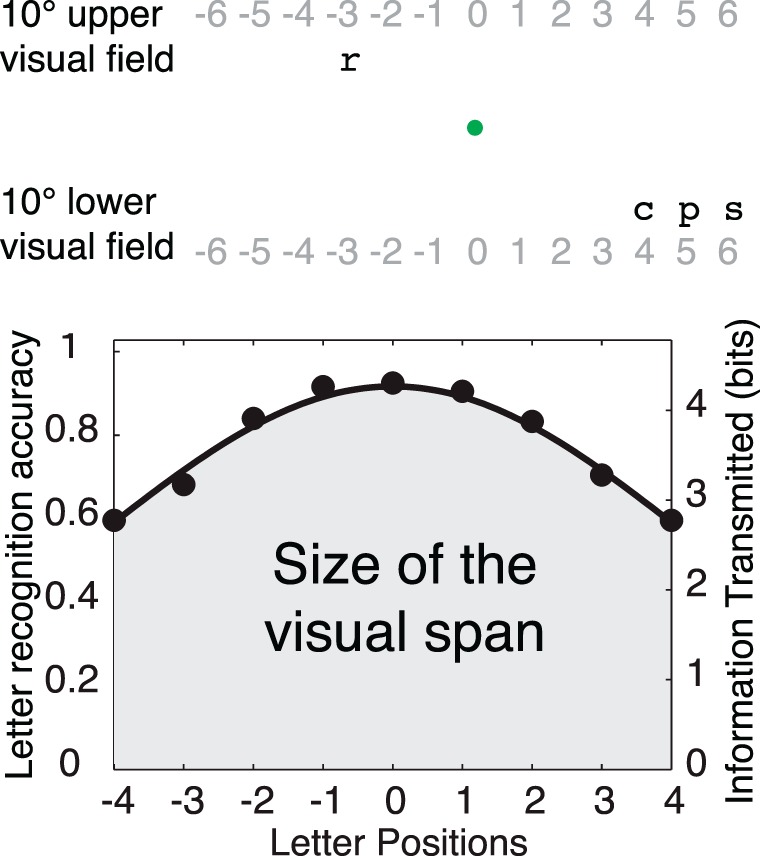
Upper: visual span measurement. The letter “r” appears at position −3 in the upper visual field, and the trigram “cps” is centered at position 5 in the lower visual field. The gray numbers indicate the position of each slot, which do not appear during the test. Lower: diagram of the visual span. A visual span profile is plotted as letter-recognition accuracy versus letter position and then converted to information transmitted (bits). The area under the curve is the size of the visual span. (Adapted with permission from Yu et al., 2010.)
The accuracy data (percent correct recognition) from the visual span measurements were fitted with a split Gaussian curve (Legge et al., 2001), as shown in Figure 2. The fitting has three parameters: height (the peak of the curve, assumed to occur at letter position 0), SDleft (the standard deviation of the left half Gaussian curve) and SDright. Only data from slots −4 to 4 were used, because there were no inner letters at slot ±5 and no inner or middle letters at slot ±6, which would lead to imbalanced sample size in further analysis. In the visual-span profile, the accuracy of letter recognition in each slot was converted into information transmitted in bits1, ranging from 0 bits for chance performance to 4.7 bits for 100% accuracy in recognizing one of 26 characters. The size of the visual span was defined as the sum of these values (in bits) for performance in slots −4 to 4, similar to computing the area under the visual-span profile.
Decomposition analysis
Our analysis focused on determining the factors that account for the difference between perfect performance (100% accuracy at all letter positions) in the trigram task, and the actual performance represented by the visual-span profile. To isolate the impact of acuity, mislocations, and crowding, we used a decomposition analysis, involving comparison between three data profiles (Figure 3). We assessed the impact of letter acuity by comparing perfect performance (100% accuracy at all positions) and an isolated-letter profile based on recognition accuracy for single letters. Mislocations in the trigram task occur when a stimulus letter is reported out of order. To estimate the rate of mislocations, letter-recognition accuracy was calculated in two ways: exact accuracy (requiring both correct identity and spatial order) and mislocational accuracy (correct identity regardless of spatial-order errors). The impact of mislocations was the difference between the mislocational and exact trigram profiles. Finally, the impact of crowding was calculated as the difference between the isolated-letter and mislocational trigram profiles. In our analysis we adopted a linear model to isolate the impact of each factor, but we acknowledge that there may also be interactions between factors, which are not captured by our current analysis.
Figure 3.
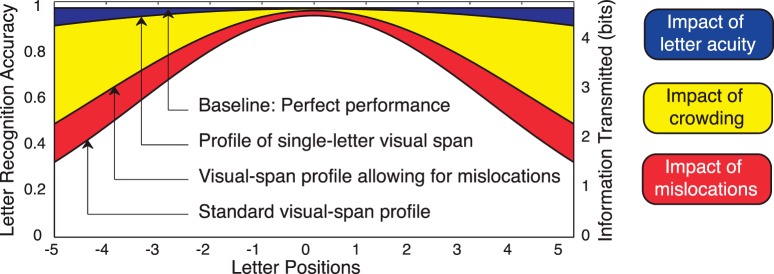
Diagram of decomposition analysis. The decomposition analysis can be expressed in equation form in bits as follows:
Perfect Performance – Exact Profile = Acuity Effect + Crowding Effect + Mislocation Effect
Where:
Acuity Effect = Perfect Profile – Single-Letter Profile (Blue Area)
Crowding Effect = Single-Letter Profile – Mislocational Profile (Yellow Area)
Mislocation Effect = Mislocational Profile – Exact Profile (Red Area)
Reading speed measurement
An RSVP paradigm (Chung et al., 2004) was used to measure the reading speed for both ordered and unordered text. A random sample was drawn from a sentence pool for each trial. No sentence was tested on one subject more than once. The sentence length was between 7 and 15 words and averaged 11 words per sentence. The word length ranged from 1 to 12 letters, with an average of four letters per word. In an RSVP reading trial, subjects fixated on a horizontal line in the middle of the screen, and the sentence appeared word by word, left justified, in the upper or lower visual field. The position of the sixth letter fell on the midline. The subject read the words out loud, and the experimenter recorded the number of correctly read words in each sentence. Horizontal eye movements along the fixation line were permitted, but when vertical eye movements were observed during the presentation of the text, the trial was cancelled. In the ordered text condition, the words in the sentence were displayed in their original meaningful order. In the unordered text condition, the order of the words from the randomly selected sentence was scrambled. A row of “x” 's was used as a premask and a postmask.
Depending on the performance of each subject in practice trials, the word-exposure time could be one of two duration sets: 30, 53, 93, 164, 290, and 511 ms, or 53, 93, 164, 290, 511, and 900 ms. The word-exposure time was fixed during one trial, but varied between different trials. Each block consisted of 36 trials, with each of the six durations tested for six sentences. Psychometric functions (accuracy vs. duration) were fitted with cumulative Gaussian curves on linear-log axes, and the reading speed in words per minute (wpm) was then calculated from the exposure duration yielding 80% word accuracy.
Results
Effect of training on the size of the visual span
Figure 4A shows the average visual span profiles for the training and control groups (for profiles of individual subjects, see Appendix A). As expected, there was an upward shift of the visual span profiles for the training group in the posttest, but no statistically significant difference in visual span size was found between pre- and posttests in the control group. An examination of the three estimated parameters from the curve fitting (height, SDleft and SDright) showed a significant increase of curve height in both the trained and untrained field for the training group, as well as a significant increase of SDleft and SDright in the trained field. These changes indicated that training produced both elevation and broadening of visual-span profiles.
Figure 4.

Training effect on the visual span. (A) Visual span profiles for each group average. Open and filled circles are original data points from pre- and posttests, respectively. Dashed and solid lines are fitted split Gaussian curves. (B) Changes in the size of the visual span. White bars, pretest; black bars, posttest. In all figures, error bars represent ±1 SEM; **, p < 0.01; ***, p < 0.001.
In order to make a quantitative comparison, we computed the size of the visual span in terms of the information transmitted in bits (see Method). The results are shown in Figure 4B.
A paired-samples one-tailed t-test was conducted to compare the size of the visual span in pre- and posttests (Table 2). For the training group, there was a significant increase of visual span size from pre- to posttest in the trained lower visual field from 29.93 bits to 35.36 bits (p < 0.001). In the untrained upper visual field, there was also a significant increase from 27.85 to 32.74 bits, t(5) = −5.48, p = 0.0014. The transfer of training from lower to upper visual field was 90%, defined as the ratio of the improvement (bits) in the untrained to the trained field. For the control group, no significant change was found in either field, lower field: t(5) = −.75, p = 0.49; upper field: t(5) = −1.05, p = 0.34, nor was there a significant difference between the pretest values of the training group and the control group, lower field: t(10) = −1.14, p = 0.28; upper field: t(10) = −1.39, p = 0.20.
Table 2.
Summary of statistical analysis comparing pre- and posttests.
| Item |
Visual field |
Pretest |
Posttest |
t(5) |
p |
| Training group | |||||
| Visual span size (bits) | Trained | M = 29.93, SD = 2.00 | M = 35.36, SD = 2.00 | −6.66 | <0.001 |
| Untrained | M = 27.85, SD = 2.18 | M = 32.74, SD = 2.18 | −5.48 | 0.0014 | |
| Reading speed, ordered (wpm) | Trained | M = 219, SD = 62.7 | M = 308, SD = 62.7 | −3.51 | 0.0086 |
| Untrained | M = 184, SD = 46.3 | M = 248, SD = 46.3 | −3.41 | 0.0095 | |
| Reading speed, unordered (wpm) | Trained | M = 186, SD = 30.7 | M = 218, SD = 30.7 | −2.61 | 0.024 |
| Untrained | M = 171, SD = 31.6 | M = 186, SD = 31.6 | −1.21 | 0.14 | |
| Control group | |||||
| Visual span size (bits) | Lower | M = 32.56, SD = 1.57 | M = 33.04, SD = 1.57 | −.75 | 0.49 |
| Upper | M = 30.53, SD = 1.09 | M = 31.00, SD = 1.09 | −1.05 | 0.34 | |
| Reading speed, ordered (wpm) | Lower | M = 228, SD = 41.1 | M = 237, SD = 41.1 | −.51 | 0.32 |
| Upper | M = 174, SD = 61.3 | M = 216, SD = 61.3 | −1.66 | 0.079 | |
| Reading speed, unordered (wpm) | Lower | M = 158, SD = 26.5 | M = 171, SD = 26.5 | −1.19 | 0.14 |
| Upper | M = 143, SD = 29.2 | M = 156, SD = 29.2 | −1.10 | 0.16 | |
| Decomposition (training group) | |||||
| Impact of acuity (bits) | Trained | M = .23, SD = .75 | M = .31, SD = .75 | −.25 | 0.6 |
| Untrained | M = .55, SD = .51 | M = .55, SD = .51 | ≈0 | 0.5 | |
| Impact of crowding (bits) | Trained | M = 10.29, SD = 2.34 | M = 5.46, SD = 2.34 | 5.05 | 0.002 |
| Untrained | M = 11.65, SD = 2.32 | M = 7.39, SD = 2.32 | 4.50 | 0.0032 | |
| Impact of mislocations (bits) | Trained | M = 1.30, SD = .37 | M = .62, SD = .37 | 4.50 | 0.0032 |
| Untrained | M = 1.70, SD = .14 | M = 1.08, SD = .14 | 10.95 | <0.001 | |
Decomposition analysis
The decomposition analysis, described in Method, was applied to the data from the training group to assess the contributions of acuity, crowding and mislocations. The results are shown in Figure 5.
Figure 5.

Decomposition analysis of the visual span. Upper panel: comparison of decomposition profiles in pre- and posttests. Areas with different colors represent the effect size of individual factors. Blue area, letter acuity; yellow area, crowding; red area, mislocations. Lower panel: comparison of effect size for each factor in pre- and posttests. White bars, pretest; black bars, posttest.
In the decomposition profiles, the area representing the crowding effect (yellow) is larger than the areas representing the acuity (red) and mislocation (blue) effects. This means that crowding contributes more than acuity loss or mislocation errors to the reduction from 100% in accuracy in the trigram letter-recognition task. We observed a notable shrinkage of the area representing crowding (yellow) from pretest to posttest. There was also slight shrinkage of the area representing mislocation errors (red). In contrast, there was no significant change in the very small area representing the impact of acuity (blue).
A 3 × 2 [influencing factors (acuity, crowding, mislocations) × test time (pre-/ posttest)] two-way repeated measures ANOVA was conducted to examine the training-related changes in the three factors. In the trained field, there was a significant main effect of the influencing factors, F(2, 30) = 7.43, p = 0.011, indicating a difference among the magnitudes (in bits) of the three factors. Pairwise comparison between the three factors indicated that in both pre- and posttests, the impact of crowding (pretest: 10.29 bits; posttest: 5.46 bits) was significantly larger than that of acuity (pretest: 0.23 bits; posttest: 0.31 bits) or mislocations (pretest: 1.3 bits; posttest: 0.62 bits), but the impact of acuity and mislocations did not differ. Also, there was a significant main effect of the test time, F(1, 30) = 53.41, p < 0.001, in the trained field, revealing a change in the size of the factors from pretest to posttest. For the untrained field, there were significant main effects of the influencing factors, F(2, 30) = 7.85, p = 0.0088, and test time, F(1, 30) = 97.05, p < 0.001, with similar results from the pairwise comparison between the size of the three factors (pretest: 11.65, 1.7 and 0.23 bits for the impact of crowding, mislocations and acuity; posttest: 7.39, 1.08 and 0.55 bits).
Results from paired-samples t-tests showed that after training, in both the trained and untrained field, there was a significant reduction in the impact of crowding (trained: 4.83 bits; untrained: 4.26 bits) and mislocations (trained: 0.68 bits; untrained: 0.62 bits), but the impact of acuity remained the same (Table 2). Further comparison revealed that in both visual fields, the reduction of crowding in bits was significantly larger than that of mislocations. Despite the fact that our training reduced the impact of both crowding and mislocations by approximately the same percentage, the larger magnitude of the reduction in crowding is more significant in guiding future designs of training protocols.
In sum, these results show that: 1) Crowding has a greater influence on the size of the visual span than letter acuity and mislocations; 2) After training, there was a substantial reduction in the magnitude of crowding and a smaller reduction for mislocations while the impact of letter acuity was unchanged; 3) The training effects largely transferred to an untrained retinal location.
Effects of training on reading speed, and context gain
Figure 6 shows mean RSVP reading speeds for the trained and untrained groups computed for pre- and posttests (for individual subjects' reading curves, see Appendix B). The training group showed a significant increase of reading speed for ordered text in both the trained and untrained visual fields, trained: 89 wpm, t(5) = −3.51, p = 0.0086; untrained: 64 wpm, t(5) = −3.41, p = 0.0095 (Table 2). For unordered text, however, a significant increase of reading speed was only observed in the trained field, 32 wpm, t(5) = −2.61, p = 0.024, but not in the untrained field, t(5) = −1.21, p = 0.14. The control group did not show any significant pre/post changes in the reading tasks, nor was there a significant difference between the pretest values of the training group and the control group, ordered text – lower field: t(10) = −0.27, p = 0.79; upper field: t(10) = 0.28, p = 0.79; unordered text – lower field: t(10) = 1.08, p = 0.30; upper field: t(10) = 1.45, p = 0.18).
Figure 6.

Training effect on reading speed. White bars, pretest; black bars, posttest.
These results show that our training with the trigram letter-recognition task produced an improvement in RSVP reading speed for both ordered and unordered text, and this effect was partly transferable to an untrained retinal location.
As an aside, we note that the RSVP reading speed for low-vision subjects who read with a retinal locus at similar eccentricity (10° or larger) is roughly comparable to what we found in normally sighted subjects here (Cheong et al., 2007), although the variance is larger for low-vision subjects. This makes the potential generalization of our results to low-vision subjects more promising.
To evaluate the effects of training on the use of context, we defined context gain to be the ratio of reading speeds for ordered and unordered text (Pelli et al., 2007). We compared context gain in the pre- and posttests for both the trained and control groups. The only significant change was an increase in context gain in the untrained field for the training group, from 1.06 to 1.35, t(5) = −5.96, p = 0.0019. Variability in the context gain measure may hide a training-related change in the trained field: Two subjects in the group showed decreases of context gain by 0.39 and 0.26, while the others showed increases ranging from 0.3 to 0.5. Nevertheless, we have insufficient evidence to conclude that there is a training-related effect on context gain.
According to Pelli and Tillman (2007), context contributes additively to reading speed, independent of letter recognition and word shape. Therefore, it is also reasonable to compute context gain as the difference in reading speed between ordered and unordered text, rather than as a ratio. Computed as a difference, the effect of training on context gain showed the same pattern described above: the only significant increase was observed in the untrained field of the training group, 49 wpm, t(5) = −5.13, p = 0.0037. In the trained field of the training group, the same two subjects showed reduction of context gain by 24 and 38 wpm while others showed increases between 77 and 107 wpm.
Discussion
We observed an enlargement of the visual span by 5.4 bits and an average increase in reading speed by 45% after training. The increase in visual span is roughly comparable to those from previous studies using a similar training procedure—6 bits (Chung et al., 2004) and 4.7 bits (Yu et al., 2010), but smaller than the 8.8 bits from Lee et al. (2010). The training-related improvement in reading speed we observed is slightly larger than the 41% improvement found by Chung et al. (2004) but smaller than the 54% found by Yu et al. (2010) and 83.5% found by Lee et al. (2010, computed from their table C1). We observed a strong transfer of the training effect from the trained to untrained field (90% for visual span and 109% for reading speed), whereas previous studies observed weaker transfer effects ranging from 50% to 80% (Chung et al., 2004; Lee et al., 2010; Yu et al., 2010).
Our decomposition analysis revealed that the crowding effect has the largest influence on the size of the visual span, followed by a small impact of mislocations and a negligible impact of letter acuity. The mechanism underlying the training-related expansion of the visual span is a large reduction of the crowding effect and a small reduction of mislocation errors.
Our finding that crowding is the major factor determining the size of the visual span is consistent with the findings of Pelli et al. (2007). These authors established a proportional relationship between the size of the “uncrowded span” and reading speed. However, the findings of Chung (2007) pose a problem for the hypothesized link between crowding, the size of the visual span and reading speed. Chung trained her normally sighted subjects in peripheral vision on identifying target letters flanked by other letters. The target-flanker separation was less than the standard spacing of letters in text (center-to-center separation of 0.8 × letter size). This training reduced the impact of crowding; the spatial extent of crowding (defined as the letter spacing yielding 50% letter-recognition accuracy) was reduced by 38%. But despite this training-related reduction in crowding, there was no statistically significant improvement in reading speed. Chung's training was confined to target recognition on the midline, corresponding to our letter position 0, and used target-flanker separation smaller than letter spacing in text. For our stimuli, letters on the midline were at or near the peak of the visual-span profile where the effects of crowding were modest for the letter spacing in our study (Figure 5). It is possible that the reduction of crowding for the conditions in Chung's experiment would not generalize to the conditions of our experiment.
In another study, Chung and Mansfield (2009) showed that the crowding effect was reduced by using opposite contrast polarity for target and flanking letters (white-on-gray and black-on-gray). But despite this reduction in crowding, they found no change in the size of visual-span profiles or reading speeds for strings of letters with alternating text polarity. This result is also difficult to reconcile with the proposed linkage between reading, visual span and crowding. Our decomposition analysis would imply that if polarity alternation does not affect the shape of the visual span, then some aspect of crowding between letters in text occurs at a site in the visual pathway at a stage where letter representations no longer code for contrast polarity. This stage may also be nonretinotopic, accounting for the strong degree of transfer of training we observed from the lower to upper visual field.
If a reduction in crowding results in faster reading, we might ask whether increased letter or line spacing in text would yield faster reading, particularly in peripheral vision. Chung (2002) found that reading speed in normal central and peripheral vision was improved by increasing letter spacing, until the spacing reaches and exceeds a critical letter spacing close to the standard letter separation. Consistent with this, Yu, Cheung, Legge, and Chung (2007) found that both reading speed and the size of visual-span profiles decreased with extra-wide spacing in central vision. Similarly, subjects with central-field loss from age-related macular degeneration (AMD) do not read faster with extra-wide spacing (Chung, 2012). Chung (2004) found that increasing vertical spacing between words in RSVP can increase reading speed in normal peripheral vision, but increasing line spacing in text has little or no effect on the maximum reading speed for AMD subjects (Chung, Jarvis, Woo, Hanson, & Jose, 2008; Calabrèse et al., 2010).
What underlying process accounts for the training-related reduction in crowding we observed? According to theories of crowding, our training procedure might reduce the inappropriate integration of features between adjacent letters in a bottom-up manner (Pelli, Palomares, & Majaj, 2004; Freeman & Pelli, 2007), or improve the top-down attentional resolution (He, Cavanagh, & Intriligator, 1996). Alternatively, the sensory mechanism of crowding may be unaffected by training, but subjects may learn to better recognize letters in the presence of the interfering effects of crowding. Consistent with this possibility, Chung, Levi, and Tjan (2005) observed that subjects' contrast thresholds for letter recognition in noise, measured in peripheral vision, decreased over several days of practice. They attributed the training effect to the improvement of the corresponding perceptual template for the letters, leading to better extraction of information from the noisy signal. Given the substantial transfer of training effect we observed from lower to upper visual field, nonretinotopic mechanisms may be implicated in the training effect, suggesting a modification of the top-down influence of crowding, or improved templates to recognize letters. Our experiments do not distinguish between these two explanations for the training-related improvement. Nevertheless, both explanations are consistent with the visual-span hypothesis, i.e., that training results in a release from the impact of crowding, which in turn yields a larger visual span and faster reading.
Mislocation errors are sometimes regarded as part of crowding, but we isolated the effect of mislocations as an independent factor. Chung and Legge (2009) measured recognition of adjacent pairs of letters (presented sequentially) at varying distances from fixation and attributed the increasing number of mislocations in the periphery to decreased precision of position coding of letters. They proposed a model in which the position of each letter is encoded with a Gaussian distribution. The reduction of mislocation errors after training in our study is possibly due to more precise coding of position for individual letters, i.e., reduced positional uncertainty.
In addition to the spatial factors influencing the size of the visual span, it is likely that temporal factors also play a role in determining reading speed in peripheral vision. In the trigram letter-recognition task, performance depends on stimulus exposure time. Empirical findings have shown that the temporal threshold (defined as the exposure time yielding 80% letter-recognition accuracy of the middle letter in a trigram) is much longer for macular degeneration subjects than controls (Cheong et al., 2007), and longer in normal peripheral vision than in normal central vision (Legge et al., 2001). Therefore, slow visual processing, in addition to reduced extent of the visual span, likely contributes to slow reading speed in peripheral vision. This leaves open the possibility that perceptual training yields faster visual processing in peripheral vision. To evaluate this possibility, future research could measure temporal thresholds for letter recognition before and after training.
We also considered whether changes in the utilization of context information might account for the benefit of training in peripheral vision. In our study, “context” refers to the enhancement of reading speed due to the syntactic and semantic properties of single sentences. We compared reading speed for meaningful sentences (ordered text) and scrambled “sentences” in random word order (unordered text). Context gain may be defined as the ratio of reading speeds for ordered and unordered text. Previous empirical findings are indecisive about whether better vision is associated with greater context gain. Two opposing views of context gain have been used to explain the data. According to the compensatory model (Stanovich, 1980), when the sensory access to word content is deficient, the brain will rely more on context information. The increased context gain from central to peripheral vision (Pelli et al., 2007) is consistent with this model. Another view is that more cognitive resources are required to process deficient visual input (e.g., in peripheral vision or low vision) leaving fewer cognitive resources to take advantage of contextual information. In support of this view, researchers have found greater context gain for fast readers than slow readers (Morton, 2008) and for normally-sighted subjects than low-vision subjects (Sass et al., 2006). Other researchers have found a lack of difference in context gain between central-field-loss subjects and normally sighted subjects (Fine & Peli, 1996), or between central and peripheral vision (Fine, Hazel, Petre, & Rubin, 1999), inconsistent with both views. Similarly, our study did not reveal a conclusive change in context gain following training. Assuming that the training did serve to enhance sensory processing of text in peripheral vision, we might have expected to see an increase or decrease in context gain, supporting one of the two opposed views.
Our investigation of the training-related enlargement of the visual span revealed that reduction of crowding effect and reduction of mislocations play major and minor roles respectively. These may be the sensory factors explaining the improved reading speed following training, if there is a causal link between the size of the visual span and reading speed. In sum, our results support and supplement the visual-span hypothesis, providing useful information about the relationship between crowding, visual span, reading speed, and context gain. Exploring the potential of peripheral vision will allow us to better understand its perceptual limitations for reading and its plasticity after training. To develop better rehabilitation paradigms for low-vision people, the next step is to evaluate the training effect using a more targeted paradigm for reducing the effects of crowding.
Acknowledgments
A preliminary report was given at the VSS conference (He & Legge, 2012). The authors would like to thank Susana T.L. Chung for helpful discussions. This research was supported by NIH grant EY002934.
Commercial relationships: none.
Corresponding author: Yingchen He.
Email: hexxx340@umn.edu.
Address: Department of Psychology, University of Minnesota, Twin Cities, Minneapolis, MN, USA.
Appendix A
Individual visual-span profiles
Figure A1.

Visual span profiles for individual subjects. Subjects 1 to 6 were assigned to the training group and subjects 7 to 12 to the control group.
Appendix B
Individual reading curves

Figure B1. Reading curves for ordered text. Open and filled circles are original data from pre- and posttests, respectively. Dashed lines and solid lines are fitted cumulative Gaussian curves.

Figure B2. Reading curves for unordered text.
Footnotes
The conversion is done using the following formulas, which was computed from confusion matrices of letter recognition measured by Beckman (1998):
Information transmitted in bits = −0.036996 + 4.6761 × letter recognition accuracy.
Contributor Information
Yingchen He, Email: hexxx340@umn.edu.
Gordon E. Legge, Email: legge@umn.edu.
Deyue Yu, Email: yu.858@osu.edu.
References
- Beckmann P. J. (1998). Preneural factors limiting letter identification in central and peripheral vision (Unpublished doctoral dissertation). University of Minnesota, Twin Cities, Minneapolis, MN: [Google Scholar]
- Brainard D. H. (1997). The Psychophysics Toolbox. Spatial Vision, 10 (4), 433–436 Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9176952 [PubMed] [Google Scholar]
- Calabrèse A., Bernard J.-B., Hoffart L., Faure G., Barouch F., Conrath J., et al. (2010). Small effect of interline spacing on maximal reading speed in low-vision patients with central field loss irrespective of scotoma size. Investigative Ophthalmology & Visual Science, 51 (2), 1247–1254, http://www.iovs.org/content/51/2/1247, doi:10.1167/iovs.09-3682. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Cheong A. M. Y., Legge G. E., Lawrence M. G., Cheung S.-H., Ruff M. A. (2007). Relationship between slow visual processing and reading speed in people with macular degeneration. Vision Research, 47 (23), 2943–2955, doi:10.1016/j.visres.2007.07.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L. (2002). The effect of letter spacing on reading speed in central and peripheral vision. Investigative Ophthalmology & Visual Science, 43 (4), 1270–1276, http://www.iovs.org/content/43/4/1270. [PubMed] [Article] [PubMed] [Google Scholar]
- Chung S. T. L. (2004). Reading speed benefits from increased vertical word spacing in normal peripheral vision. Optometry and Vision Science, 81 (7), 525–535 Retrieved from http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2734885/ [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L. (2007). Learning to identify crowded letters: Does it improve reading speed? Vision Research, 47 (25), 3150–3159, doi:10.1016/j.visres.2007.08.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L. (2012). Dependence of reading speed on letter spacing in central vision loss. Optometry and Vision Science, 89 (9), 1288–1298 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L., Jarvis S. H., Woo S. Y., Hanson K., Jose R. T. (2008). Reading speed does not benefit from increased line spacing in AMD patients. Optometry and Vision Science, 85 (9), 827–833, doi:10.1097/OPX.0b013e31818527ea.Reading [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L., Legge G. E. (2009). Precision of position signals for letters. Vision Research, 49 (15), 1948–1960, doi:10.1016/j.visres.2009.05.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L., Legge G. E., Cheung S.-H. (2004). Letter-recognition and reading speed in peripheral vision benefit from perceptual learning. Vision Research, 44 (7), 695–709, doi:10.1016/j.visres.2003.09.028 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L., Levi D. M., Tjan B. S. (2005). Learning letter identification in peripheral vision. Vision Research, 45 (11), 1399–1412, doi:10.1016/j.visres.2004.11.021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L., Mansfield J. S. (2009). Contrast polarity differences reduce crowding but do not benefit reading performance in peripheral vision. Vision Research, 49 (23), 2782–2789, doi:10.1016/j.visres.2009.08.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S. T. L., Mansfield J. S., Legge G. E. (1998). Psychophysics of reading. XVIII. The effect of print size on reading speed in normal peripheral vision. Vision Research, 38 (19), 2949–2962 Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9797990 [DOI] [PubMed] [Google Scholar]
- Fine E., Hazel C., Petre K., Rubin G. (1999). Are the benefits of sentence context different in central and peripheral vision? Optometry and Vision Science, 76 (11), 764–769 Retrieved from http://ukpmc.ac.uk/abstract/MED/10566861 [DOI] [PubMed] [Google Scholar]
- Fine E. M., Peli E. (1996). The role of context in reading with central field loss. Optometry and Vision Science, 73 (8), 533–539 Retrieved from http://ukpmc.ac.uk/abstract/MED/8869984 [DOI] [PubMed] [Google Scholar]
- Fine I., Jacobs R. A. (2002). Comparing perceptual learning tasks: A review. Journal of Vision, 2 (2): 5, 190–203, http://www.journalofvision.org/content/2/2/5, doi:10.1167/2.2.5. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Forster K. I. (1970). Visual perception of rapidly presented word sequences of varying complexity. Perception & Psychophysics, 8 (4), 215–221, doi:10.3758/BF03210208 [Google Scholar]
- Freeman J., Pelli D. G. (2007). An escape from crowding. Journal of Vision, 7 (2): 22, 1–14, http://www.journalofvision.org/content/7/2/22/, doi:10.1167/7.2.22. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- He S., Cavanagh P., Intriligator J. (1996). Attentional resolution and the locus of visual awareness. Nature, 383, 334–337 [DOI] [PubMed] [Google Scholar]
- He Y., Legge G. E. (2012). Reduced crowding accounts for enlargement of the visual span after training. Journal of Vision, 12 (9): 535, http://www.journalofvision.org/content/12/9/535, doi:10.1167/12.9.535. [Abstract] [Google Scholar]
- Lee H.-W., Kwon M., Legge G. E., Gefroh J. J. (2010). Training improves reading speed in peripheral vision: Is it due to attention? Journal of Vision, 10 (6): 18, 1–15, http://www.journalofvision.org/content/10/6/18, doi:10.1167/10.6.18. [PubMed] [Article] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Legge G. E. (2007). Psychophysics of reading in normal and low vision. Mahwah, NJ: Lawrence Erlbaum Associates, Inc; [Google Scholar]
- Legge G. E., Cheung S.-H., Yu D., Chung S. T. L., Lee H.-W., Owens D. P. (2007). The case for the visual span as a sensory bottleneck in reading. Journal of Vision, 7 (2): 9, 1–15, http://www.journalofvision.org/content/7/2/9, doi:10.1167/7.2.9. [PubMed] [Article] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Legge G. E., Mansfield J. S., Chung S. T. L. (2001). Psychophysics of reading. XX. Linking letter recognition to reading speed in central and peripheral vision. Vision Research, 41 (6), 725–743 Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/11248262. [DOI] [PubMed] [Google Scholar]
- Morton J. (2008). The effects of context upon speed of reading, eye movements and eye-voice span. Quarterly Journal of Experimental Psychology, 16 (4), 340–354 [Google Scholar]
- Pelli D. G. (1997). The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spatial Vision, 10 (4), 437–442, doi:10.1163/156856897X00366 [PubMed] [Google Scholar]
- Pelli D. G., Palomares M., Majaj N. J. (2004). Crowding is unlike ordinary masking: Distinguishing feature integration from detection. Journal of Vision, 4 (12): 12, 1136–1169, http://www.journalofvision.org/content/4/12/12, doi:10.1167/4.12.12. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Pelli D. G., Tillman K. A. (2007). Parts, wholes, and context in reading: A triple dissociation. PLoS ONE, 2( 8), e680, doi:10.1371/journal.pone.0000680 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelli D. G., Tillman K. A., Freeman J., Su M., Berger T. D., Majaj N. J. (2007). Crowding and eccentricity determine reading rate. Journal of Vision, 7 (2): 20, 1–36, http://www.journalofvision.org/content/7/2/20, doi:10.1167/7.2.20. [PubMed] [Article] [DOI] [PubMed] [Google Scholar]
- Rubin G. S., Turano K. (1992). Reading without saccadic eye movements. Vision Research, 32 (5), 895–902 Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/16005930 [DOI] [PubMed] [Google Scholar]
- Sass S. M., Legge G. E., Lee H. (2006). Low-vision reading speed: Influences of linguistic inference and aging. Optometry and Vision Science, 83 (3), 166–177 [DOI] [PubMed] [Google Scholar]
- Stanovich K. E. (1980). Toward an interactive-compensatory model of individual differences in the development of reading fluency. Reading Research Quarterly, 16 (1), 32–71 [Google Scholar]
- Yu D., Cheung S.-H., Legge G. E., Chung S. T. L. (2007). Effect of letter spacing on visual span and reading speed. Journal of Vision, 7 (2): 2, 1–10, http://www.journalofvision.org/content/7/2/2, doi:10.1167/7.2.2. [PubMed] [Article] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yu D., Legge G. E., Park H., Gage E., Chung S. T. L. (2010). Development of a training protocol to improve reading performance in peripheral vision. Vision Research, 50 (1), 36–45, doi:10.1016/j.visres.2009.10.005 [DOI] [PMC free article] [PubMed] [Google Scholar]