

Abstract
Objective
To conduct a meta-analysis of all published genetic association studies of 5-HTTLPR polymorphisms performed in PTSD cases
Methods Data Sources
Potential studies were identified through PubMed/MEDLINE, EMBASE, Web of Science databases (Web of Knowledge, WoK), PsychINFO, PsychArticles and HuGeNet (Human Genome Epidemiology Network) up until December 2011. Study Selection: Published observational studies reporting genotype or allele frequencies of this genetic factor in PTSD cases and in non-PTSD controls were all considered eligible for inclusion in this systematic review. Data Extraction: Two reviewers selected studies for possible inclusion and extracted data independently following a standardized protocol. Statistical analysis: A biallelic and a triallelic meta-analysis, including the total S and S' frequencies, the dominant (S+/LL and S'+/L'L') and the recessive model (SS/L+ and S'S'/L'+), was performed with a random-effect model to calculate the pooled OR and its corresponding 95% CI. Forest plots and Cochran's Q-Statistic and I2 index were calculated to check for heterogeneity. Subgroup analyses and meta-regression were carried out to analyze potential moderators. Publication bias and quality of reporting were also analyzed.
Results
13 studies met our inclusion criteria, providing a total sample of 1874 patients with PTSD and 7785 controls in the biallelic meta-analyses and 627 and 3524, respectively, in the triallelic. None of the meta-analyses showed evidence of an association between 5-HTTLPR and PTSD but several characteristics (exposure to the same principal stressor for PTSD cases and controls, adjustment for potential confounding variables, blind assessment, study design, type of PTSD, ethnic distribution and Total Quality Score) influenced the results in subgroup analyses and meta-regression. There was no evidence of potential publication bias.
Conclusions
Current evidence does not support a direct effect of 5-HTTLPR polymorphisms on PTSD. Further analyses of gene-environment interactions, epigenetic modulation and new studies with large samples and/or meta-analyses are required.
Background
Post-traumatic stress disorder (PTSD) is a mental disorder that occurs following exposure to a potentially traumatic life event (Criterion A) and is characterized by symptoms of re-experience, avoidance, dulling of the senses and hyperarousal. Those who develop PTSD are at substantially increased risk of unemployment, marital instability and health problems (major depression, substance dependence, impaired role functioning and reduced life course opportunities) [1]. Therefore, PTSD constitutes a potential major health care burden. However, while trauma exposure rates vary between 40–80% over the course of the life of individuals, only a percentage of those exposed to traumatic events (25% approximately) will develop PTSD [2].
Recent data indicate that both the risk of trauma exposure and PTSD may be influenced by genetic factors [3], [4]. The evidence for genetic influences on risk for PTSD comes from a diversity of family, twin and molecular genetic studies. An elevated risk of PTSD among relatives with PTSD and a heritability of almost 30% suggest that genes are important risk factors in the etiology of the disorder. Molecular genetic studies using case-control candidate gene-association designs have shown a variety of results. Several candidate genes related to the current understanding of the neurobiology of the disorder have been studied [3]. The most frequently investigated genetic variant recently studied is the role of the human serotonin transporter (5-hydroxytryptamine transporter, 5-HTT) gene (SLC6A4), through polymorphisms in its promoter region. The importance of this gene is primarily due to the increasing evidence of its role in modulating sensitivity to stress and vulnerability to psychopathology [5]. The promoter activity of the 5-HTT gene, located at 17q11.1-q12, is modified by sequential elements within the proximal 5 regulatory region, designated the 5-HTT gene-linked polymorphic region (5-HTTLPR) [6]. The less frequent short (S) allele in the 5-HTTLPR is associated with lower transcriptional efficiency of the promoter compared with the more frequent long (L) allele [7] and has been related to suicidal behavior [8], depression [9], neurotic personality trait [7], [10], alcoholism [11] and PTSD [12].
Other 5-HTTLPR variants have been described in a Japanese study [13] showing a third functional allele, LG, with an A>G polymorphism at position 6 of the first two 22-bp imperfect repeats that define the 16-repeat L allele (SNP rs25531). This LG is equivalent in expression to the S allele [11]. Thus, 5-HTTLPR is a triallelic locus with alleles designated as LG, LA, and S and the three of them appear to act codominantly [11]. Because the LG and S alleles have comparable levels of serotonin transporter expression, both of which are lower than that of LA, a novel approach has been used with a reclassification of the alleles on the basis of lower and higher levels of expression [14], [15]. This approach reclassified LG and S as S' and LA as L'.
Differences in ethnic distribution have been described with a higher frequency of the long variant allele (L) in Europeans (57%; 95% CI: 49.9–61.8%) than in Asians (27%; 95% CI: 23.9–32.9%) [8] and a different distribution of the triallelic classification between Afro-American adults (L'L': 34.8%; S'L': 47.8%; and S'S': 17.4%) and European-Americans (L'L': 25.1%; S'L': 54.9%; and S'S': 20.0%) [16]. The allelic frequency of LG is 0.09–0.14 in Caucasians and 0.24 in Afro-Americans [11].
Two studies have reported an association of PTSD with the SS genotype [17], [18] and a third [19] reported a significant association with the LALA but not with the SS genotype. Other studies [12], [16], [19]–[21] have not found a direct link between the 5-HTTLPR genotype and the risk of PTSD but have reported a gene-environment (GxE) interaction whereby the genetic effect of 5-HTTLPR is modified by the level of trauma exposure [4], [22]. Of the five studies that analyzed a potential GxE interaction, four found a significant interaction with the low-expression genotype [12], [16], [17], [19], [20] and another [19] found a significant interaction but with the LALA genotype. These inconsistencies in the published findings of genetic association studies of 5-HTTLPR and PTSD might be the result of several factors that need to be systematically assessed, including different designs, statistical power and quality characteristics of primary studies.
Although previous reviews of the genetic factors involved in PTSD have been published [3], [4], [22], [23], none of them can be considered a systematic review and no meta-analysis has been conducted [24]. The main aim of the current study was to perform a systematic review and a meta-analysis of all available genetic association studies of 5-HTTLPR polymorphisms in PTSD and to determine if the S allele or SS genotype increases the risk of PTSD in those exposed to a traumatic experience compared to the L allele or LL genotype. A secondary aim of this meta-analysis was to compare the association between the genotype using the biallelic and the triallelic models and to assess if the latter better captures the effect of the 5-HTTLPR variation on PTSD.
Methods
Search strategy
Potential published studies were identified through PubMed/MEDLINE, EMBASE, Web of Science databases (Web of Knowledge, WoK), PsychINFO, PsychArticles and HuGeNet (Human Genome Epidemiology Network) using the search terms: “[PTSD OR posttraumatic stress disorder OR trauma* stress] AND [ 5-HTTLPR OR 5-HTT OR SLC6A4 OR SERTPR OR Serotonin Transporter Gene]” up until December 2011. The reference lists of original studies and review articles included were manually searched to identify other potentially eligible studies. In addition, the MEDLINE option `Related Articles` was used for the same purpose. To minimize potential publication bias, no restrictions were placed on time period, sample size, population and language of publication or type of report.
Inclusion and exclusion criteria
Published observational studies (criterion 1) reporting genotype or allele frequencies of the genetic factor (criterion 2) in PTSD cases and in non-PTSD controls (criterion 3) were all considered eligible for inclusion in this systematic review. Case status was defined as having a current or one-year DSM-III, DSM-IIIR or DSM-IV (Diagnostic and Statistical Manual of Mental Disorders) diagnosis of PTSD assessed by established psychiatric interviews. Studies of all ethnic groups were considered eligible. Reviews, case-only studies, family-based designs and population studies with only healthy subjects were excluded, as well as other studies describing genetic effects on other anxiety or depression-related phenotypes such as anxiety, depression or on different personality traits.
Two review authors (MTE and FNM) independently selected studies for possible inclusion in the study. Firstly, titles and abstracts identified from the search were independently reviewed. Secondly, each review author independently examined the full text of all studies that they considered to be of possible relevance. Each review author compiled a list of studies that they believed met the inclusion criteria. The content of each review author's list was compared and any discrepancies discussed. Any disagreement was resolved by discussion and a consensus was reached by all authors.
Data extraction
Two investigators (FNM and MTE) independently extracted data using a standardized data extraction form and they were entered into separate databases by each of the reviewers independently following standardized procedures. In case of disagreement, a consensus was reached by the reviewers. Where more than one psychiatric disorder was described in a particular study, only data from the PTSD sample were extracted. Therefore, in the case of multiple papers from a single study, only the results of the publication with the highest number of participants were included, since the unit of analysis was the study rather than the reports to avoid duplicity. When essential data were missing from the study report, the corresponding author was contacted and asked to provide the required data.
Information on the data extraction form included: author(s), journal and year of publication; methodology details (study design, sample size for both cases and controls, diagnostic tools for determination of case status and definition of case status); and sample characteristics (gender ratio, mean age, ethnic background, Hardy-Weinberg Equilibrium (HWE) in controls, genotype and allele frequencies when appropriate, type of trauma event, time from trauma exposure, severity of PTSD, other mental health disorders, including comorbid substance abuse and suicide-related behaviors).
Quality of the studies
As poor reporting quality has been associated with a biased estimation of effects in clinical intervention studies [25], the quality of each study selected for inclusion was assessed by applying a 12-item quality checklist, derived from the STREGA statement (Strengthening the Reporting of Genetic Association Studies) [26] and other criteria [24], [27], [28]. Specifically, the quality criteria were: (1) representativeness of cases; (2) representativeness of controls; (3) same PTSD diagnostic instrument used for cases and controls; (4) trauma-exposed controls; (5) identical trauma exposure for cases and controls; (6) assessment of ethnicity; (7) blind assessment of genotyping and phenotyping; (8) quality control procedures for genotyping methods; (9) HWE testing; (10) adjustment for potential confounding variables; (11) control for multiple comparisons and (12) assessment of psychiatric comorbidity. A quality score of one was assigned if the criteria were correctly assessed and the Total Quality Score (TQS) of each study was calculated by adding all the corresponding quality item scores (range: 0–12 with a higher score indicating a higher overall quality). Discrepancies in the quality evaluation of each study were resolved by consensus. Consistent with current guidelines, we did not weight studies by TQS or exclude studies with low-quality scores.
Statistical analysis
Inter-rater agreement was measured by Cohen's kappa coefficient for inclusion and exclusion criteria. The association between the 5-HTLLPR allelic frequency and PTSD was examined by statistical analyses, firstly using the biallelic model (Biallelic Frequency Model, BFM) and then a triallelic one (Triallelic Frequency Model, TFM) by calculating the respective Odds Ratio (OR) and its corresponding 95% confidence interval (95% CI) as the effect size measurement for these analyses. The triallelic model was used whenever the original researchers provided the frequencies of S' (as the sum of the frequencies of LG and S) and the frequency of L' (LA). However, as the type of inheritance of the 5-HTTLPR polymorphism was not yet known, two more ORs were calculated per study and model, the first one reflecting the risk of PTSD associated with possessing at least one S or S' allele (Biallelic or Triallelic Dominant Model, BDM or TDM, respectively) and the second one reflecting the risk of association with two S or S' alleles (Biallelic or Triallelic Recessive Model, BRM or TRM respectively). ORs were calculated for each study so that an OR of greater than one reflected a higher risk for PTSD. To assess the possible differences between a biallelic or a triallelic model and due to the non-independent nature of the studies compared, a qualitative comparison of the results was performed on those studies which had data for both models in the same sample.
Random-effects models were applied in the statistical analyses because heterogeneity among the studies was expected. This assumes a genuine diversity in the results of the various studies and it incorporates a between-studies variance into the calculations. In each meta-analysis, a pooled OR and its corresponding 95% CI were calculated. In addition, the statistical significance of the pooled OR was assessed using the Z test [29]. A sensitivity analysis was performed to assess whether our results were substantially influenced by the presence of any individual study by systematically removing each study and recalculating the significance of the result.
Forest plots were constructed to represent the individual and pooled effect estimates, with their 95% CIs, and to allow visual inspection for study heterogeneity. To check for this among the studies, both the Cochran's Q-statistic and I2 index were calculated [30]. When the ORs are homogeneous, Q-statistic follows a chi-squared distribution with k – 1 degrees of freedom (d.f.) (k being the number of studies). The degree of heterogeneity was estimated with the I2 index (I2 = 100×(Q–d.f.)/Q), which can be interpreted as the percentage of total variation across the studies due to their different characteristics. I2 values around 25%, 50%, and 75% denote low, moderate and large heterogeneity, respectively.
To explore heterogeneity, different subgroup analyses, which allow chi-squared tests for between group differences, were performed taking as potential moderators the quality items, ethnicity and study design. In addition, meta-regression analyses were carried out with continuous moderators, such as the mean age of participants, the TQS, percentage of males, year of publication, percentage of Caucasian or Euro-Americans and percentage of Afro-Americans or Africans to test potential explanatory variables leading to heterogeneity.
Chi-squared tests were conducted to test for HWE in the reported genotype frequencies among the controls in case-control and cross-sectional studies and among the whole population in the cohort studies. Deviations from HWE (P-value <0.05) might indicate genotyping errors, limited population size, population substructure or newly occurring mutations. No deviation from HWE is considered a quality measure of a genetic association study and a subgroup analysis was performed to compare those studies with and without HWE.
To assess whether publication bias may be a threat to the validity of the pooled ORs, funnel plots with the Duval and Tweedie's trim-and-fill method [31] were applied, as well as the Egger test [32]. Where funnel plot asymmetry was observed, effect estimates corrected for small study effects, such as publication bias, were generated by the trim-and-fill method. This uses available data to estimate the number and outcome of missing (unreported) studies and recalculates the overall effect that would be observed with their inclusion. The Egger test is an unweighted regression consisting of taking the precision of each study as the independent variable (precision being defined as the inverse of the standard error of each effect size) and the effect size divided by its standard error as the dependent variable. A non-statistically significant result of the t-test for the hypothesis of an intercept equal to zero allows discounting of publication bias as a threat to the validity of the pooled effect [32].
All statistical tests were interpreted assuming a significance level of 5% (α = 0.05) and all were two-tailed. The main statistical analyses were carried out using the software package RevMan 5.029. Meta-regressions, funnel plots with the trim-and-fill method and the Egger test were calculated with the program Comprehensive Meta-analysis 2.0 [33]. Methods of the analysis and inclusion-exclusion criteria were specified a priori and documented in a protocol. The published recommendations for systematic reviews of genetic association studies were followed [24], [27], [34]. Since we only used previously published data, we did not consider it necessary to seek ethical approval or written informed consent.
Results
Figure 1 details the search process flow and results. From a total of 25 potentially eligible studies, seven were excluded because they were a case or case-series design [35]–[37] or were focused on other phenotypes apart from PTSD [38]–[41]. Cohen's kappa inter-rater agreement coefficient for the three inclusion criteria ranged from 0.70 (criterion 3) to 1.0 (criteria 1 and 2) and 0.905 for the exclusion criteria. From the 18 studies eligible for inclusion, three were excluded as they were based on the same sample ([12], [42] and [43]) as the final included studies ([16] and [44]), respectively. Fifteen studies were included in the review and their data were extracted [16]–[21], [45]–[50]. An additional study was sent by the corresponding author who was contacted for further information about a published study. By the time this meta-analysis was performed, this study was accepted for publication and, therefore, included in this review [51]. Three studies were excluded as essential data on 5-HTTLPR genotypic frequencies could not be provided by the authors [44], [52], [53]. Finally, 13 studies were included in the meta-analysis.
Figure 1. Flow chart of the Meta-Analysis of 5-HTTLPR polymorphisms and Post-traumatic Stress Disorder (PTSD).

Adapted from Sagoo GS, Little J, Higgins JP, Systematic Reviews of Genetic Association Studies. Human Genome Epidemiology Network. PLOS Med 2009; 6(3): e28 doi: e28 10.1371/journal.pmed.1000028 and Moher D, Liberati A, Tetzlaff J, Altman DG, The PRISMA Group (2009). Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med 6(6): e1000097. doi:10.1371/journal.pmed1000097.
The characteristics of studies eligible for inclusion are described in Table 1, including year of publication, study design, number of PTSD cases, controls and total sample, number and percentage of males, mean age, diagnostic instrument used to assess PTSD, type of PTSD assessed, biallelic or triallelic genotype approach, whether a GxE interaction had been analyzed and a brief description of the environment studied and of the final decision to include or exclude. The pooled population was 1,874 patients with PTSD and 7,785 controls in the biallelic meta-analysis and 627 and 3,524, respectively, in the triallelic one.
Table 1. Characteristics of association studies eligible for inclusion in Meta-Analysis of 5-HTTLPR polymorphisms and Post-traumatic Stress Disorder (PTSD).
| First author | Year | Study design | PTSD N | Controls N | Total sample (N) | Males N (%) | MeanAge | Diagnostic instrument | PTSD assessed | Biallelic/ Triallelic Genotype | GxE study | Interaction description | Remarks | |
| Lee [18] | 2005 | Case-Control | 100 | 197 | 297 | 119 (40.1) | 34.94 | SCID-I Korean Version | Current | Biallelic | No | - | Included | |
| Koenen [16] | 2009 | Cross-Sectional | 19 | 571 | 590 | 215 (36.4) | 65.1 | National Women's Study PTSD Module | Last 6 months | Triallelic | Yes | High crime & High unemployment rate | Included | |
| Mellman [21] | 2009 | Case-Control | 55 | 63 | 118 | 40 (33.9) | 39.97 | CAPS | Lifetime | Triallelic | No | - | Included | |
| Xie [20] | 2009 | Cross-Sectional | 229 | 1023 | 1252 | 656 (52.4) | 38.97 | SSADDA | Lifetime | Triallelic | Yes | Childhood adversity & Adult traumatic events | Included | |
| Grabe [19] | 2009 | Cross-Sectional | 67 | 1596 | 1663 | 857 (51.5) | 57.61 | PTSD Module of SCID | Lifetime | Triallelic | Yes | Frequent Trauma event | Included | |
| Thakur [47] | 2009 | Cohort study | 24 | 17 | 41 | 19 (46.3) | 31.96 | CAPS | Last year | Biallelic | No | – | Included | |
| King [52] | 2009 | Cohort study | – | – | 378 | – | – | Computer-aided telephone interviews. | Lifetime | Triallelic | Yes | Childhood adversity | Not included | |
| Kolassa [17] | 2010 | Cross-Sectional | 331 | 77 | 408 | 218 (53.4) | 34.72 | Post-traumatic Diagnostic Scale | Lifetime | Biallelic | Yes | Number of traumatic events experienced | Included | |
| Handwerger [53] | 2010 | Case-Control | 18 | 18 | 36 | – | – | – | – | – | No | – | Not included | |
| Saying [48] | 2010 | Cohort study | 29 | 48 | 77 | 47 (61) | – | CAPS | Lifetime | Biallelic | No | – | Included | |
| Koenen [50] | 2011 | Cross-Sectional | 23 | 77 | 100 | 40 (40) | 45.32 | Modified PTSD Checklist | Lifetime | Triallelic | Yes | Number of traumatic events | Included | |
| Morey [49] | 2011 | Case-Control | 22 | 20 | 42 | 22 (52.4) | 34.04 | DTS. Cutoff score = 32 | Current | Triallelic | No | – | Included | |
| Valente [46] | 2011 | Case-Control | 65 | 34 | 99 | 28 (28.3) | 39.99 | SCID-I & CAPS | Current | Biallelic | No | – | Included | |
| Wang [45] | 2011 | Case-Control | 212 | 176 | 388 | 200 (51.55) | 49.06 | SCID-I, MINI, CES y CAPS | Current | Triallelic | No | – | Included | |
| King [44] | 2011 | Case-Control | 16 | 14 | 30 | – | – | – | – | Biallelic | No | – | Not included | |
| Xie [51] | 2012 | Cross- Sectional | AA | 321 | 2078 | 2399 | 1343 (56) | 42.2 | SSADDA | Lifetime | Biallelic | Yes | Childhood adversity | Included |
| EA | 398 | 2381 | 2779 | 1559 (56.1) | 39 |
GxE study: Gene-Environmental interaction study; SCID-1: Structured Clincal Interview for DSM-IV; CAPS: Clinical Assessed PTSD Scale; SSADDA: Semi-Structured Assessment for Drug Dependence and Alcoholism; MINI: Mini-International Neuropsychiatric Interview; CES: Combat Exposure Scale; DTS: Davidson Trauma Scale; AA: Afro-Americans; EA: Euro-Americans.
Originally, six studies [17], [18], [46]–[48], [51] described a biallelic analysis and seven [16], [19]–[21], [45], [49], [50] a triallelic. Nevertheless, five of the latter group [19], [21], [45], [49], [50] provided the frequencies of the biallelic genotype so that they could also be included in the meta-analysis of the biallelic approach (Table 2).
Table 2. Frequencies of the 5-HTTLPR alleles and polymorphisms of the included studies&.
| Biallelic genotype approach | HWE | Triallelic genotype approach | HWE | ||||||||||||||||||||
| PTSD | Controls | P-value | PTSD | Controls | P-value | ||||||||||||||||||
| S | L | SS | SL | LL | S | L | SS | SL | LL | S' | L' | S'S' | S'L' | L'L' | S’ | L' | SvS' | S'Lv | LvL' | ||||
| Lee [18] | 2005 | 175 | 25 | 77 | 21 | 2 | 319 | 75 | 129 | 61 | 7 | 0.81 | – | – | – | – | – | – | – | – | – | – | – |
| Koenen [16] | 2009 | – | – | – | – | – | – | – | – | – | – | – | 17 | 21 | 4 | 9 | 6 | 539 | 603 | 116 | 307 | 148 | 0.06 |
| Mellman [21 | 2009 | 42 | 70 | 5 | 32 | 19 | 35 | 93 | 7 | 21 | 36 | 0.16 | 66 | 46 | 20 | 26 | 10 | 67 | 61 | 18 | 31 | 15 | 0.81 |
| Xie [20] | 2009 | – | – | – | – | – | – | – | – | – | – | – | 234 | 224 | 58 | 118 | 53 | 957 | 1089 | 234 | 489 | 300 | 0.20 |
| Grabe [19] | 2009 | 40 | 94 | 6 | 28 | 33 | 1294 | 1898 | 264 | 766 | 566 | 0.86 | 48 | 86 | 8 | 32 | 27 | 1512 | 1680 | 364 | 784 | 448 | 0.55 |
| Thakur [47] | 2009 | 23 | 25 | 8 | 7 | 9 | 22 | 12 | 7 | 8 | 2 | 0.09 | – | – | – | – | – | – | – | – | – | – | – |
| Kolassa [17] | 2010 | 112 | 550 | 15 | 82 | 234 | 29 | 125 | 1 | 27 | 49 | 0.20 | – | – | – | – | – | – | – | – | – | – | – |
| Saying [48] | 2010 | 30 | 28 | 6 | 18 | 5 | 41 | 55 | 6 | 29 | 13 | 0.045 | – | – | – | – | – | – | – | – | – | – | – |
| Koenen [50] | 2011 | 10 | 34 | 1 | 8 | 13 | 45 | 103 | 12 | 21 | 41 | 0.005 | 21 | 23 | 4 | 13 | 5 | 67 | 81 | 17 | 33 | 24 | 0.39 |
| Morey [49] | 2011 | 22 | 22 | 7 | 8 | 7 | 18 | 22 | 3 | 12 | 5 | 0.34 | 26 | 18 | 10 | 6 | 6 | 22 | 18 | 5 | 12 | 3 | 0.34 |
| Valente [46] | 2011 | 55 | 71 | 13 | 29 | 21 | 38 | 30 | 9 | 20 | 5 | 0.26 | – | – | – | – | – | – | – | – | – | – | – |
| Wang [45] | 2011 | 207 | 217 | 56 | 95 | 61 | 131 | 221 | 21 | 89 | 66 | 0.28 | 261 | 163 | 94 | 73 | 45 | 164 | 188 | 29 | 106 | 41 | 0.005 |
| Xie (AA) [51] | 2012 | 222 | 650 | 26 | 170 | 240 | 1269 | 3997 | 170 | 929 | 1534 | 0.07 | – | – | – | – | – | – | – | – | – | – | – |
| Xie (EA) [51] | 2012 | 460 | 564 | 96 | 268 | 148 | 2511 | 3187 | 566 | 1379 | 904 | 0.34 | – | – | – | – | – | – | – | – | – | – | – |
S, short allele; L, long allele; S' as S + LG; and L' as LA.
Dominant model (S+ vs LL or S'+vs L'L'): S+ (or S'+) genotype frequencies are calculated as the sum of SS (or S'S') and SL (or S'L') frequencies. Recessive model (SS vs L+ or S'S' vs L'+): L+ (or L'+) genotype frequencies are calculated by the sum of LL (or L'L') and SL (or SvL') genotype frequencies.
Meta-analysis of the allelic association with PTSD
Firstly, a meta-analysis of the biallelic approach was performed for: i) allele frequency (S vs L) (Fig 2); ii) the dominant model, BDM (S+vs LL) (Fig 3); and iii) the recessive model, BRM (SS vs L+) (fig 4). The overall association between genotypes and the risk of PTSD was not significant using all three different approaches: (i) OR = 1.05; 95% CI = 0.87, 1.26; p = 0.63; I2 = 63; ii) OR = 1.01; 95% CI = 0.78, 1.30; p = 0.95; I2 = 57; and iii) OR = 1.15; 95% CI = 0.82, 1.60; p = 0.41; I2 = 55, respectively) (Figs 2, 3 and 4).
Figure 2. Forest plot of the 5-HTTLPR biallelic frequency model (S vs L) and Post-traumatic Stress Disorder (PTSD).

Figure 3. Forest plot of 5-HTTLPR biallelic dominant model (S+ vs LL) and Post-traumatic Stress Disorder (PTSD).

Figure 4. Forest plot of 5-HTTLPR biallelic recessive model (SS vs L+) and Post-traumatic Stress Disorder (PTSD).

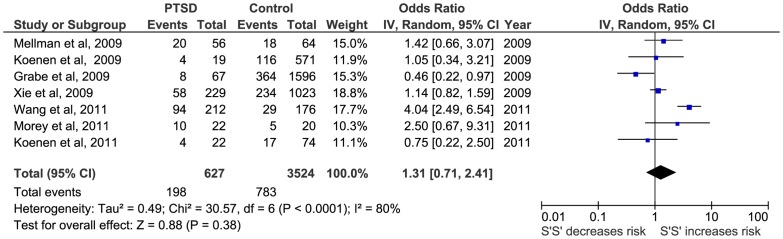
Secondly, a meta-analysis of the triallelic approach offered similar results for the TFM model (OR = 1.15; 95%CI = 0.82, 1.60; p = 0.41; I2 = 74), for TDM model (OR = 1.02; 95%CI = 0.73, 1.48; p = 0.93; I2 = 43) and for the TRM (OR = 1.31; 95% CI = 71, 2.41; p = 0.38; I2 = 80) (Figs 5, 6 and 7, respectively).
Figure 5. Forest plot of 5-HTTLPR triallelic frequency model (S' vs L') and Post-traumatic Stress Disorder (PTSD).
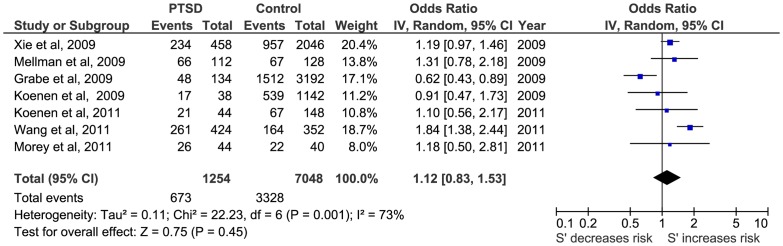
Figure 6. Forest plot of 5-HTTLPR triallelic dominant model (S'+ vs L'L') and Post-traumatic Stress Disorder (PTSD).

Figure 7. Forest plot of 5-HTTLPR triallelic recessive model (S'S' vs L'+) and Post-traumatic Stress Disorder (PTSD).
Comparison of the biallelic or triallelic models of the 5-HTTLPR polymorphisms
Of the seven studies describing a triallelic approach, it was not possible to calculate the biallelic frequencies of one of the studies and we could not obtain further data from the authors on request [16]. The biallelic frequencies of the second study [20] were included in a different study when requested [51] and the genotype frequencies from PTSD and controls could not be calculated from the text as they only provided the total biallelic frequencies. Finally, five studies [19], [21], [45], [49], [50] reported allele and 5-HTTLPR polymorphism frequencies of the same population allowing comparison of both approaches. The result were very similar in the allele frequency model (BFM, OR = 1.07; 95% CI: 0.66, 1.74; I2 = 78% and TFM, OR = 1.14; 95% CI: 0.70, 1.87; I2 = 81%), in the dominant model (BDM, OR = 1.09; 95% CI: 0.61, 1.96; I2 = 72%, TDM, OR = 0.93; 95% CI: 0.60, 1.44; I2 = 39%) and in the recessive model (BRM, OR = 1.05; 95% CI: 0.41, 2.67; I2 = 73%, TRM, OR = 1.40; 95% CI: 0.54, 3.57; I2 = 85%).
Sensitivity analysis
Only one study [19] was found to alter the results when individually removed from the meta-analysis during sensitivity analysis and it affected the TFM model (OR = 1.31; 95% CI = 1.06, 1.63; p = 0.01; I2 = 35). The remaining results were robust to sensitivity analysis, with the overall P-values remaining non-significant when each study was individually removed from the analysis. Analysis of heterogeneity varied from moderate (I2 = 43 in TDM) to extreme (I2 = 80 in TRM) in the different models.
Publication bias
In order to assess whether publication bias might be a threat to the pooled ORs, several graphical and analytic techniques were applied. Firstly, funnel plots were constructed and the trim-and-fill method proposed by Duval and Tweedie was applied to them in order to achieve symmetry when they showed an asymmetric pattern. Of the six funnel plots constructed, the trim-and-fill method did not require the addition of any new effect estimate in five of them to achieve symmetry (Fig 8). The only exception was with the Biallelic Recessive model where the Duval and Tweedie's method added a new effect estimate. However, the adjusted pooled OR (OR = 1.10; 95% CI: 0.79, 1.54) showed a negligible difference from the original pooled OR (OR = 1.14; 95% CI: 0.81, 1.59). In addition, the Egger test was applied to each of the six meta-analytic databases, in all cases achieving a non-statistically significant result for the intercept of the regression model (p>0.05) (Table 3). Therefore, on the basis of this, we felt that we could reasonably discount publication bias as a threat to our meta-analytic findings.
Figure 8. Funnel plots of 5-HTTLPR polymorphisms and Post-traumatic Stress Disorder (PTSD) to assess publication bias.

White circles represent each of the included studies. Black circles represent the new effect estimated to achieve symmetry.
Table 3. Analyses of publication bias by the Egger test.
| Model | Intercept | SE | T | df | p-value |
| Biallelic | |||||
| BFM | −0.319 | 0.857 | −0.371 | 10 | 0.718 |
| BDM | −0.658 | 0.721 | −0.912 | 10 | 0.383 |
| BRM | 0.273 | 0.763 | 0.358 | 10 | 0.727 |
| Triallelic | |||||
| TFM | −0.842 | 1.738 | −0.484 | 5 | 0.648 |
| TDM | −0.775 | 1.160 | −0.668 | 5 | 0.534 |
| TRM | −0.625 | 2.010 | −0.311 | 5 | 0.768 |
SE: Standard error; T: T-test; df: Degrees of freedom. BFM: Biallelic Frequency Model; BDM: Biallelic Dominant Model; BRM: Biallelic Recessive Model; TFM: Triallelic Frequency Model; TDM: Triallelic Dominant Model; TRM: Triallelic Recessive Model.
Quality of the studies
Table 4 describes the quality characteristics of the studies analyzed. Included studies (N = 13) had a superior TQS (mean = 6.31, SD = 2.428) than those excluded studies (N = 3; mean = 1.33; SD = 1.528) (p = 0.005). The TQS of those studies with a biallelic approach (N = 6; mean = 4.17, SD = 1.329) was significantly lower than of those with a triallelic approach (N = 7; mean = 8.14; SD = 1.345) (p<0.001).
Table 4. Description of the quality characteristics of the included and excluded studies in the Meta-Analysis of 5-HTTLPR polymorphisms and Post-traumatic Stress Disorder (PTSD).
| First author | Year | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||||||||||
| Repre sentative cases$ | Repre sentative controls# | Same diagnostic instrument | Trauma Exposed controls | Identical trauma experience | Ethnicity& | Blind assessment | Quality control Genotyping | HWE† testing | Confounders adjustment | Control for multiple comparisons | Psychiatric Comorbidity assessed | TQS ‡ | |||||||||||
| INCLUDED STUDIES | |||||||||||||||||||||||
| Lee [18] | 2005 | No | No | No | No | No | AS (100) | No | No | Yes | No | No | No | 2 | |||||||||
| Koenen [16] | 2009 | No | Yes | Yes | Yes | Yes | EA (90.7) Others (9.5) | No | Yes | No | Yes | No | No | 7 | |||||||||
| Mellman [21] | 2009 | No | Yes | Yes | Yes | No | AA (100) | No | Yes | No | Yes | Yes | Yes | 8 | |||||||||
| Xie [20] | 2009 | No | Yes | Yes | Yes | No | AA (53.5) EA (46.5) | No | No | Yes | Yes | No | Yes | 7 | |||||||||
| Grabe [19] | 2009 | No | Yes | Yes | Yes | No | Eu (100) | No | Ye | Yes | Yes | No | Yes | 8 | |||||||||
| Thakur [47] | 2009 | No | Yes | Yes | Yes | No | Caucasian (95.1) Other (4.9) | Yes | No | Yes | No | No | No | 5 | |||||||||
| Kolassa [17] | 2010 | No | Yes | Yes | Yes | No | Af (100) | No | No | Yes | No | No | No | 5 | |||||||||
| Saying [48] | 2010 | No | Yes | Yes | Yes | No | No | No | No | No | Yes | No | Yes | 5 | |||||||||
| Koenen [50] | 2011 | Yes | Yes | Yes | Yes | No | AA (79) Others (21) | No | Yes | No | Yes | No | Yes | 8 | |||||||||
| Morey [49] | 2011 | No | Yes | Yes | Yes | Yes | Caucasian (47.6) Others (52.4) | No | No | Yes | No | Yes | Yes | 8 | |||||||||
| Valente [46] | 2011 | No | No | Yes | Mix | No | No | No | No | Yes | No | No | Yes | 3 | |||||||||
| Wang [45] | 2011 | No | Yes | Yes | Yes | Yes | Caucasian (71.5) AA (24) Others (4.5) | Yes | Yes | Yes | Yes | Yes | Yes | 11 | |||||||||
| Xie [51] | 2012 | No | No | Yes | No | No | AA (46.3) EA (53.7) | No | No | Yes | Yes | No | Yes | 5 | |||||||||
| EXCLUDED STUDIES | |||||||||||||||||||||||
| King [52] | 2009 | No | No | Yes | Mix | No | AA, EA | No | No | No | Yes | No | No | 3 | |||||||||
| Handwerger [53] | 2010 | No | No | – | Yes | – | – | No | No | No | No | No | No | 1 | |||||||||
| King [44] | 2011 | No | No | – | Mix | No | – | No | No | No | No | No | No | 0 | |||||||||
Representativeness of cases included all eligible cases with outcome of interest over a defined period of time, all cases in a defined catchment area, all cases in a defined hospital or clinic, group of hospitals, health maintenance organization, or an appropriate sample of those cases (e.g. random sample).
Representativeness of controls assesses whether the control series used in the study is derived from the same population as the cases and essentially would have been cases had the outcome been present and included community and clinical controls within the same community or hospitalized population as cases.
If Yes, the % of the different ethnics groups are provided in brackets.AS: Asiatic. EA: Euro-American; AA: Afro-Americans; Af: African; Eu: Europeans.
HWE: Hardy-Weinberg Equilibrium; ‡TQS: Total Quality Score.
The subgroup analysis of the different quality components (Tables 5 and 6) showed that there were some significant differences in estimated effect size between those studies that fulfilled the quality criteria and those that did not: the same principal trauma for PTSD and controls (BFM and BRM), reporting of HWE (BDM) and control for multiple comparisons (BFM and BDM) in the biallelic analysis. In the triallelic analysis, the significant differences were for the same principal trauma for PTSD and controls (TFM and TRM), the presence of blind assessment (TRM) and control for multiple comparisons (TFM and TRM). Moreover, when stratifying for the quality criteria, the pooled effect size of those that fulfilled them reached significance (same principal trauma for PTSD and controls – BFM, BRM and TRM – presence of blind assessment – TRM – and control for multiple comparisons – BFM, TFM and TRM -). On the other hand, there was one characteristic where not fulfilling the quality criteria significantly decreased the risk, i.e. those studies which did not use the same diagnostic instrument for PTSD and controls (BFM and BRM) (Tables 5 and 6).
Table 5. Subgroup analysis of study design, type of PTSD assessed and quality characteristics in the Meta-Analysis of 5-HTTLPR polymorphisms and Post-traumatic Stress Disorder (PTSD) in the biallelic approach.
| K‡ | Number of participants | Frequency Model # | Dominant Model | Recessive Model | |||||||||||
| OR | 95%CI | I2(%) | P-value & | OR | 95%CI | I2 (%) | P-value & | OR | 95%CI | I2 (%) | P-value & | ||||
| Case representativeness | |||||||||||||||
| Yes | 1 | 96 | 0.67 | 0.31, 1.48 | NA | 0.86 | 0.33, 2.26 | NA | 0.25 | 0.03, 2.01 | NA | ||||
| No | 11 | 9563 | 1.07 | 0.88, 1.26 | 63 | 0.27 | 1.02 | 0.78, 1.32 | 61 | 0.75 | 1.18 | 0.81, 1.65 | 56 | 0.15 | |
| Control representativeness | |||||||||||||||
| Yes | 8 | 2835 | 1.01 | 0.72, 1.43 | 70 | 0.98 | 0.86, 1.38 | 65 | 1.20 | 0.62, 2.30 | 50 | ||||
| No | 4 | 6824 | 1.06 | 0.88, 1.28 | 54 | 0.82 | 1.09 | 0.86, 1.38 | 39 | 0.66 | 1.03 | 0.77, 1.39 | 40 | 0.68 | |
| Same diagnostic instrument | |||||||||||||||
| Yes | 11 | 9362 | 1.01 | 0.84, 1.22 | 63 | 0.99 | 0.77, 1.29 | 61 | 1.07 | 0.74, 1.53 | 54 | ||||
| No | 1 | 297 | 1.65 | 1.01, 2.68 | NA | 0.07 | 1.81 | 0.37, 8.85 | NA | 0.47 | 1.76 | 1.02, 3.06 | NA | 0.13 | |
| Trauma exposed control | |||||||||||||||
| Yes | 9 | 2932 | 0.96 | 0.69, 1.33 | 70 | 0.89 | 0.57, 1.39 | 66 | 1.12 | 0.62, 2.02 | 57 | ||||
| No | 3 | 6727 | 1.09 | 0.94, 1.26 | 38 | 0.47 | 1.15 | 0.99, 1.32 | 0 | 0.28 | 1.08 | 0.76, 1.51 | 56 | 0.90 | |
| Same principal trauma for PTSD and controls | |||||||||||||||
| Yes | 2 | 430 | 1.57 | 1.19, 2.06 | 0 | 1.38 | 0.90, 2.12 | 2 | 2.65 | 1.58, 4.44 | 0 | ||||
| No | 10 | 9229 | 0.98 | 0.81, 1.26 | 63 | 0.005 | 0.96 | 0.72, 1.27 | 61 | 0.17 | 0.97 | 0.74, 1.26 | 20 | 0.0007 | |
| Assessment of ethnicity | |||||||||||||||
| Yes | 10 | 9485 | 1.07 | 0.88, 1.29 | 65 | 1.04 | 0.81, 1.33 | 57 | 1.15 | 0.79, 1.67 | 62 | ||||
| No | 2 | 174 | 0.93 | 0.40, 2.14 | 72 | 0.75 | 0.77 | 0.15, 3.87 | 76 | 0.72 | 1.06 | 0.43, 2.59 | 25 | 0.87 | |
| Presence of blind assessment | |||||||||||||||
| Yes | 2 | 429 | 0.98 | 0.31, 3.02 | 83 | 0.69 | 0.11, 4.29 | 78 | 1.57 | 0.45, 5.53 | 70 | ||||
| No | 10 | 9230 | 1.01 | 0.85, 1.20 | 52 | 0.95 | 0.99 | 0.76, 1.28 | 56 | 0.70 | 1.02 | 0.76, 1.36 | 29 | 0.51 | |
| Reporting of quality control procedures in genotyping methods | |||||||||||||||
| Yes | 4 | 2267 | 1.05 | 0.60, 1.83 | 84 | 1.15 | 0.60, 2.22 | 78 | 0.89 | 0.31, 2.56 | 77 | ||||
| No | 8 | 7392 | 1.04 | 0.89, 1.22 | 35 | 0.99 | 0.98 | 0.76, 1.27 | 41 | 0.66 | 1.09 | 0.83, 1.43 | 24 | 0.71 | |
| Reporting HWE† | |||||||||||||||
| Yes | 9 | 9366 | 1.01 | 0.82, 1.24 | 69 | 0.92 | 0.71, 1.20 | 60 | 1.18 | 0.81, 1.71 | 64 | ||||
| No | 3 | 293 | 1.23 | 0.76, 2.00 | 39 | 0.46 | 1.64 | 0.85, 3.17 | 33 | 0.11 | 0.90 | 0.34, 2.37 | 27 | 0.61 | |
| Adjustment for potential confounders | |||||||||||||||
| Yes | 7 | 8774 | 1.09 | 0.88, 1.35 | 70 | 1.16 | 0.38, 1.03 | 59 | 1.04 | 0.67, 1.60 | 67 | ||||
| No | 5 | 885 | 0.92 | 0.60, 1.41 | 57 | 0.48 | 0.63 | 0.38, 1.03 | 14 | 0.03 | 1.39 | 0.82, 2.35 | 20 | 0.40 | |
| study design. In addition, meta regres | |||||||||||||||
| Yes | 3 | 550 | 1.57 | 1.23, 2.01 | 0 | 1.60 | 0.97, 2.63 | 31 | 1.95 | 0.93, 4.07 | 38 | ||||
| No | 9 | 9109 | 0.94 | 0.87, 1.14 | 56 | 0.001 | 0.89 | 0.67, 1.16 | 55 | 0.04 | 0.98 | 0.73, 1.31 | 33 | 0.09 | |
| Psychiatric comorbidity assessed | |||||||||||||||
| Yes | 9 | 8913 | 1.05 | 0.86, 1.29 | 66 | 1.08 | 0.83, 1.40 | 59 | 1.07 | 0.73, 1.56 | 58 | ||||
| No | 3 | 746 | 0.97 | 0.53, 1.78 | 69 | 0.81 | 0.70 | 0.30, 1.62 | 36 | 0.34 | 1.57 | 0.87, 2.83 | 11 | 0.26 | |
| Study design | |||||||||||||||
| Cross-Sectional | 5 | 8597 | 0.94 | 0.78, 1.12 | 52 | 0.94 | 0.73, 1.21 | 58 | 0.87 | 0.64, 1.17 | 23 | ||||
| Case-Control | 5 | 944 | 1.32 | 0.93, 1.86 | 56 | 1.20 | 0.63, 2.28 | 60 | 1.61 | 0.97, 2.66 | 45 | ||||
| Cohort | 2 | 118 | 0.89 | 0.32, 2.49 | 71 | 0.22 | 0.69 | 0.09, 5.26 | 75 | 0.75 | 1.16 | 0.46, 2.91 | 6 | 0.12 | |
| Type of PTSD assessed | |||||||||||||||
| Lifetime PTSD | 7 | 8794 | 0.99 | 0.83, 1.19 | 50 | 1.04 | 0.74, 1.36 | 61 | 0.90 | 0.71, 1.14 | 8 | ||||
| Current PTSD | 4 | 824 | 1.24 | 0.80, 1.93 | 66 | 0.93 | 0.43, 2.02 | 56 | 1.79 | 1.06, 3.01 | 45 | ||||
| Last year | 1 | 41 | 0.50 | 0.20, 1.24 | NA | 0.21 | 0.22 | 0.04, 1.21 | NA | 0.21 | 0.71 | 0.20, 2.58 | NA | 0.06 | |
K: Number of studies; #Number of alleles S or L duplicates the number of participants as each participant has two alleles (S, L or one of each one); OR: Odds Ratio; 95% CI: 95% Confident Interval; & Chi-squared test for subgroup differences; NA: Not applicable; †HWE: Hardy-Weinberg Equilibrium.
Table 6. Subgroup analyses of study design, type of PTSD assessed and quality characteristics in the Meta-Analysis of 5-HTTLPR polymorphisms and Post-traumatic Stress Disorder (PTSD) in the triallelic approach.
| K‡ | Number of participants | Frequency Model# | Dominant Model | Recessive Model | ||||||||||||||||||||||
| OR | 95%CI | I2 (%) | P-value & | OR | 95%CI | I2 (%) | P-value & | OR | 95%CI | I2 (%) | P-val ue & | |||||||||||||||
| Case representativeness | ||||||||||||||||||||||||||
| Yes | 1 | 96 | 1.10 | 0.56, 2.17 | NA | 1.63 | 0.54, 4.95 | NA | 0.75 | 0.22, 2.50 | NA | |||||||||||||||
| No | 6 | 4055 | 1.13 | 0.80, 1.58 | 77 | 0.96 | 0.97 | 0.68, 1.40 | 50 | 0.39 | 1.41 | 0.73, 2.73 | 83 | 0.37 | ||||||||||||
| Control representativeness | ||||||||||||||||||||||||||
| Yes | 7 | 4151 | 1.09 | 0.80, 1.49 | 73 | 1.02 | 0.73, 1.42 | 43 | 1.31 | 0.71, 2.41 | 80 | |||||||||||||||
| No | 0 | 0 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | ||||||||||||
| Same diagnostic instrument | ||||||||||||||||||||||||||
| Yes | 7 | 4151 | 1.12 | 0.83, 1.53 | 73 | 1.02 | 0.73, 1.42 | 43 | 1.31 | 0.71, 2.41 | 80 | |||||||||||||||
| No | 0 | 0 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | ||||||||||||
| Trauma exposed control | ||||||||||||||||||||||||||
| Yes | 7 | 4151 | 1.12 | 0.83, 1.53 | 73 | 1.02 | 0.73, 1.42 | 43 | 1.31 | 0.71, 2.41 | 80 | |||||||||||||||
| No | 0 | 0 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | ||||||||||||
| Same principal trauma for PTSD and controls | ||||||||||||||||||||||||||
| Yes | 3 | 1020 | 1.76 | 0.83, 2.31 | 0 | 0.99 | 0.65, 1.49 | 0 | 2.45 | 1.05, 5.72 | 59 | |||||||||||||||
| No | 4 | 3131 | 0.99 | 0.72, 1.35 | 62 | 0.006 | 1.09 | 0.64, 1.86 | 66 | 0.75 | 0.92 | 0.57, 1.49 | 48 | 0.05 | ||||||||||||
| Assessment of ethnicity | ||||||||||||||||||||||||||
| Yes | 7 | 4151 | 1.12 | 0.83, 1.53 | 73 | 1.02 | 0.73, 1.42 | 43 | 1.31 | 0.71, 2.41 | 80 | |||||||||||||||
| No | 0 | 0 | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | ||||||||||||
| Presence of blind assessment | ||||||||||||||||||||||||||
| Yes | 1 | 388 | 1.09 | 0.67, 1.77 | 82 | 1.13 | 0.70, 1.82 | NA | 4.04 | 2.49, 6.54 | NA | |||||||||||||||
| No | 6 | 3763 | 1.19 | 0.98, 1.45 | 0 | 0.75 | 0.98 | 0.63, 1.52 | 52 | 0.67 | 1.02 | 0.69, 1.51 | 34 | <0.0001 | ||||||||||||
| Reporting of quality control procedures in genotyping methods | ||||||||||||||||||||||||||
| Yes | 5 | 2857 | 1.09 | 0.67, 1.77 | 82 | 0.94 | 0.63, 1.39 | 34 | 1.20 | 0.48, 3.05 | 85 | |||||||||||||||
| No | 2 | 1294 | 1.19 | 0.98, 1.45 | 0 | 0.75 | 1.06 | 0.43, 2.62 | 44 | 0.81 | 1.29 | 0.74, 2.25 | 22 | 0.90 | ||||||||||||
| Reporting HWE† | ||||||||||||||||||||||||||
| Yes | 4 | 3345 | 1.13 | 0.71, 1.79 | 86 | 0.93 | 0.58, 1.50 | 67 | 1.49 | 0.59, 3.76 | 90 | |||||||||||||||
| No | 3 | 806 | 1.13 | 0.80, 1.59 | 0 | 0.99 | 1.19 | 0.67, 2.10 | 0 | 0.52 | 1.14 | 0.65, 2.01 | 0 | 0.63 | ||||||||||||
| Adjustment for potential confounders | ||||||||||||||||||||||||||
| Yes | 6 | 4109 | 1.12 | 0.80, 1.56 | 78 | 1.05 | 0.74, 1.48 | 47 | 1.22 | 0.63, 2.35 | 83 | |||||||||||||||
| No | 1 | 42 | 1.18 | 0.50, 2.81 | NA | 0.91 | 0.47 | 0.10, 2.21 | NA | 0.32 | 2.50 | 0.67, 9.31 | NA | 0.34 | ||||||||||||
| Control for multiple comparisons | ||||||||||||||||||||||||||
| Yes | 3 | 550 | 1.65 | 1.29, 2.09 | 0 | 1.11 | 0.74, 1.67 | 0 | 2.55 | 1.22, 5.32 | 61 | |||||||||||||||
| No | 4 | 3601 | 0.92 | 0.63, 1.35 | 69 | 0.01 | 0.97 | 0.56, 1.69 | 67 | 0.70 | 0.85 | 0.53, 1.38 | 40 | 0.01 | ||||||||||||
| Psychiatric comorbidity assessed | ||||||||||||||||||||||||||
| Yes | 6 | 3561 | 1.15 | 0.82, 1.62 | 77 | 1.04 | 0.72, 1.51 | 50 | 1.35 | 0.69, 2.66 | 83 | |||||||||||||||
| No | 1 | 590 | 0.91 | 0.47, 1.73 | NA | 0.52 | 0.76 | 0.28, 2.03 | NA | 0.55 | 1.05 | 0.34, 3.21 | NA | 0.70 | ||||||||||||
| Study design | ||||||||||||||||||||||||||
| Case-Control | 4 | 3601 | 0.92 | 0.63, 1.35 | 69 | 0.97 | 0.56, 1.69 | 67 | 0.85 | 0.53, 1.38 | 40 | |||||||||||||||
| Cohort | 3 | 550 | 1.65 | 1.29, 2.09 | 0 | 0.01 | 1.11 | 0.74, 1.67 | 0 | 0.70 | 2.55 | 1.22, 5.32 | 61 | 0.01 | ||||||||||||
| Type of PTSD assessed | ||||||||||||||||||||||||||
| Lifetime PTSD | 4 | 3131 | 1.00 | 0.69, 1.45 | 71 | 1.09 | 0.64, 1.86 | 66 | 0.92 | 0.57, 1.49 | 48 | |||||||||||||||
| Current PTSD | 2 | 430 | 1.76 | 1.34, 2.31 | 0 | 1.00 | 0.56, 1.79 | 11 | 3.82 | 2.43, 6.00 | 0 | |||||||||||||||
| Last 6 months | 1 | 590 | 0.91 | 0.47, 1.73 | NA | 0.02 | 0.76 | 0.28, 2.03 | NA | 0.82 | 1.05 | 0.34, 3.21 | 80 | <0.0001 | ||||||||||||
K: Number of studies; #Number of alleles S or L duplicates the number of participants as each participant has two alleles (S, L or one of each one); OR: Odds Ratio; 95% CI: 95% Confident Interval; &Chi-squared test for subgroup differences; NA: Not applicable; †HWE: Hardy-Weinberg Equilibrium.
The subgroup analysis of study design and the type of PTSD assessed revealed significant differences between study design in TFM and TRM and between types of PTSD assessed (TFM and TRM). The pooled risk effect of cohort studies and restriction of the analysis to current PTSD significantly increased the risk in those carrying S' (TFM) or S'S' (TRM) (Table 6).
Four studies did not calculate HWE (Table 4) but, when we calculated it (Table 2), two of them were in HWE [16], [21] and the rest were not [48], [50]. Another study was only in HWE in the biallelic approach but not in the triallelic one [45]. Given these discrepancies, we performed a subgroup analysis with our calculated HWE and there were significant differences in the triallelic analysis between those studies in HWE when compared to those without in the TFM (OR = 1.00; 95% CI = 0.76, 1.32; I2 = 53 and OR = 1.84; 95% CI = 1.38, 2.44; I2 = Not applicable, respectively, p-value of the Chi-squared test for subgroup difference = 0.003) and in TRM (OR = 1.02; 95% CI = 0.69, 1.51; I2 = 34 and OR = 4.04; 95% CI = 2.49, 6.54; I2 = Not applicable, respectively, p-value of the Chi-squared test for subgroup difference <0.0001).
Meta-regression analysis (Table 7) showed modifying effects of four variables, two of them in the direction of decreasing the risk of the S'+ on PTSD (the median age of participants –TDM- and the percentage of European or Caucasian -TDM-) and the other two increasing the effect of S'+ on PTSD (the percentage of Afro-Americans or Africans – TDM -) and that of S'S' (TQS – TRM -). No significant association with other pre-specified moderator variables was identified.
Table 7. Meta-regression analysis of potential modifying variables in the Meta-Analyses of 5-HTTLPR polymorphisms and Post-traumatic Stress Disorder (PTSD).
| Frequency Model | Dominant Model | Recessive Model | |||||||
| bj | 95%CI | p-value | bj | 95%CI | p-value | bj | 95%CI | p-value | |
| Year of publication | |||||||||
| Biallelic approach | −0.0521 | −0.1764, 0.0723 | 0.41 | −0.0728 | −0.3205, 0.1749 | 0.56 | −0.0132 | −0.2193, 0.1928 | 0.90 |
| Triallelic approach | 0.2077 | −0.0769, 0.4924 | 0.15 | 0.0550 | −0.3532, 0.4631 | 0.79 | 0.4648 | −0.0054, 0.9351 | 0.50 |
| Males (%) | |||||||||
| Biallelic approach | 0.0094 | −0.0227, 0.4140 | 0.57 | 0.0042 | −0.0440, 0.0524 | 0.86 | 0.0346 | −0.0175, 0.0868 | 0.19 |
| Triallelic approach | 0.0007 | −0.0431, 0.0445 | 0.97 | −0.0124 | −0.0639, 0.0390 | 0.64 | 0.0167 | −0.0696, 0.1030 | 0.70 |
| Mean Age | |||||||||
| Biallelic approach | −0.0110 | −0.0431, 0.0212 | 0.50 | 0.0046 | −0.0561, 0.0652 | 0.88 | −0.0315 | −0.1027, 0.0397 | 0.39 |
| Triallelic approach | −0.0177 | −0.0516, 0.0162 | 0.31 | −0.0324 | −0.0582, −0.0065 | 0.01 | −0.0259 | −0.0964, 0.0446 | 0.47 |
| European or Caucasian (%) | |||||||||
| Biallelic approach | −0.0048 | −0.0126, 0.0030 | 0.23 | −0.0072 | −0.0188, 0.0044 | 0.23 | −0.0045 | −0.0203, 0.0113 | 0.57 |
| Triallelic approach | −0.0039 | −0.0126, 0.0048 | 0.38 | −0.0102 | −0.0178, −0.0025 | <0.01 | −0.0018 | −0.0199, 0.0163 | 0.85 |
| Afro-Americans and Africans (%) | |||||||||
| Biallelic approach | 0.0015 | −0.0066, 0.0096 | 0.72 | 0.0053 | −0.0060, 0.0166 | 0.36 | 0.0002 | −0.0158, 0.0162 | 0.98 |
| Triallelic approach | 0.0035 | −0.0052, 0.0123 | 0.43 | 0.0110 | 0.0033, 0.0187 | 0.005 | −0.0004 | −0.0180, 0.0172 | 0.96 |
| Total Quality Score (TQS) | |||||||||
| Biallelic approach | 0.0120 | −0.0483, 0.0723 | 0.70 | 0.0175 | −0.0666, 0.1017 | 0.69 | 0.0418 | −0.0605, 0.1440 | 0.42 |
| Triallelic approach | 0.1484 | −0.0670, 0.3638 | 0.18 | 0.0113 | −0.2636, 0.2863 | 0.94 | 0.3518 | 0.4017, 0.6619 | 0.03 |
bj: regression coefficient for the moderator variable; 95% CI: 95% Confident Interval.
Gene-Environmental (GxE) Interactions
Only six included studies analyzed GxE interactions [16], [17], [19], [20], [50], [51] (Table 1). The environments studied were those of high crime and high unemployment rates [16], adult traumatic events [20], childhood adversities [20], [51] and number of traumatic events experienced [17], [19], [50]. Four studies detected a significant GxE interaction, three of them with the S or S' allele interacting with the environment [16], [20], [51] and the fourth described an additive GxE interaction with the L' (LA) allele [19]. Interestingly, the GxE interaction effect, with childhood adversity as the modifying variable, was only described in European Americans but not in Afro-Americans [20], [51]. Two studies did not find a significant GxE interaction for 5-HTTLPR genotype [17], [50]. The low number of studies of each different environment did not permit a meta-analysis.
Discussion
To our knowledge, this is the first published meta-analysis of the relationship between 5-HTTLPR polymorphisms and PTSD. We found no significant relationship between biallelic or triallelic polymorphism and PTSD, using the three possible approaches (allele frequency, dominant and recessive model). Contrary to expectation, the triallelic approach (S' and L') did not appear to alter the results of the meta-analysis obtained by the biallelic approach (S and L). There was no apparent publication bias but there was great variability in TQS and in some individual quality characteristics of the included studies. For example, there was a significant main effect when the analysis was restricted to those studies measuring current PTSD (BRM, TFM and TRM) and a significant effect in cohort studies was apparent in TFM and TRM, the study design which is less prone to bias than case-control and cross-sectional ones. Finally, several potential moderators were detected in the meta-regression analyses (mean age of participants, ethnic distribution of the population studied and the TQS) and these significantly modified the results. These results suggest that when more homogeneous groups within studies are considered, thus fulfilling these criteria, potential evidence that PTSD is associated with 5-HTTLPR may indeed emerge and this warrants further research.
There has recently been increasing interest in developing guiding principles for reporting results of different study designs in order to improve the quality of research reports. The STrengthening the REporting of Genetic Association studies (STREGA) Statement was published in 2009 [26] as an extension of the Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) [54] and was specifically designed to enhance the transparency of the reports of genetic association studies. Interestingly, none of the seven studies published since then [17], [45], [46], [48]–[51] has followed these recommendations.
One possible consequence of our results might be that PTSD is not directly associated with 5-HTTLPR. However, several other reasons need to be considered to explain the absence of a significant effect of the 5-HTTLPR on PTSD, given the increasing evidence suggesting the implication of the 5-HTTLPR polymorphisms and different psychopathological conditions related to stress sensitivity [5]. Firstly, published work in this research area has only considered a third functional allele in the 5-HTTLPR polymorphisms. Our findings do not support significant differences between the biallelic and the triallelic analyses. Nevertheless, at least ten allelic variants have been described in 5-HTTLPR polymorphisms in humans [13] and it is not clear whether the magnitude of any association may be affected or moderated by this variability.
Secondly, the lack of a significant association may be due to limited statistical power. Emerging evidence from large genetic studies of other mental disorders suggest the individual effect size of specific alleles may be very small with disorders which are highly polygenic [55]. Consistent with this, findings of other meta-analyses of the same genetic factor with different phenotypes have found a small association. For example, the pooled OR for the S allele and unipolar depression is 1.08 (95% CI = 1.03, 1.12) [56], 1.12 (95% CI = 1.03, 1.21) for bipolar disorder [57] and 1.18 (95% CI = 1.02, 1.33) for alcohol dependence [58]. In our meta-analysis, twelve studies were included in the biallelic model with 1,874 PTSD cases but only seven were included in the triallelic with 627 PTSD cases. Therefore, if we consider the different study designs and types of PTSD assessed, it is reasonable to consider the possibility of a lack a statistical power as necessary to detect a true relationship between 5-HTTLPR and PTSD. If this is the case, new association studies with larger samples are necessary and further meta-analyses should be performed in future to detect a more accurate estimation of a true effect. Another example is related to population stratification. Interestingly, the first published study [18] was performed in an Asian population and has not been replicated and all studies published since have been performed in Caucasian or African or Afro-American populations. As differences in ethnic distribution have been described [8], [11], [13], [16], population stratification should be controlled or stratified in future studies.
Thirdly, 5-HTTLPR might not have a primary direct effect on PTSD. As PTSD diagnosis requires exposure to an environmental stressor, it is an ideal condition for the investigation of GxE interactions, where the effects of environmental exposure are moderated by genotype [4], [59]. However, only six studies have formally tested GxE interactions in this field [16], [17], [19], [20], [50], [51]. So far, the low number of studies and the high variability of stressors analyzed do not allow formal meta-analyses of these interactions. On the other hand, there may be other interactions underlying the potential role of specific genes in the etiology of PTSD. For example, a gene-gene interaction might also explain the absence of a main genetic effect in PTSD as has been described in neuroticism, where BDNF Val66Met interacts with 5-HTTLPR to influence neuroticism [60]. It is also possible that the relationship between the 5-HTTLPR genotype and PTSD could be mediated by other personality traits as has been suggested in the case of neuroticism as a mediator of the association of the 5-HTTLPR polymorphism with lifetime major depression [61].
Finally, epigenetic modification offers a promising research area that may clarify the variability of the results obtained in PTSD research as well as in other psychiatric illnesses [62]. There has been an increasing interest in epigenetic factors in psychiatric disorders and this involves the study of inheritable changes in gene expression that occur without changes in the DNA sequence. The level of methylation of 5-HTTLPR modified the effect of the number of traumatic experiences on the risk of PTSD regardless of the 5-HTTLPR polymorphisms [50] and it may explain how the environment can modify gene expression regardless of the primary genetic sequence by changing the accessibility of information printed on the DNA [63].
We acknowledge some limitations of this meta-analysis in relation to interpretation of the findings. Some studies could not be included in the final analysis because of incomplete data. However, it is unlikely that their inclusion would have affected our main results due to the small sample size and their lower quality characteristics. Errors or bias in the design of or in the statistical tests in the primary observational studies could potentially affect the results of our meta-analysis. Although 61% of the included studies performed a controlled analysis for potential confounding variables, our meta-analyses were performed using the crude OR as the diversity of confounding variables did not allow the use of the adjusted OR. Although this is a general limitation of meta-analysis in observational studies, careful analyses of the individual quality aspects and the different study designs of the primary studies were performed to explore the moderate/large levels of heterogeneity detected.
Of particular concern is the heterogeneity resulting from the varying ethnic distribution within studies and its potential confounding effect on the pooled effect size. To analyze this particular aspect, different approaches were considered including a subgroup analysis of the assessment of ethnicity and specific meta-regression analyses of the effect of the percentage of different ethnic groups (European/Caucasian or Afro-Americans/Africans). Our results suggest that this ethnic distribution should be considered as an important modifying variable in future research.
On the other hand, several strengths of our meta-analysis deserve recognition. Firstly, although a TQS was calculated and used in the meta-regression analyses, the relevant methodological aspects were assessed individually and the influence on the magnitude of the effect was explored as the TQS (as a summation of points) giving equal weight to all characteristics [28]. Secondly, we analyzed three inheritance models, as it was not yet clear which one better represents the association between the 5-HTTLPR poylymorphisms and PTSD as well as performing two allelic studies (biallelic and triallelic). Given that there was little variation according to the model or the approach studied, this supports our view that our findings are indeed robust. Finally, we have followed current guidelines for design and reporting of systematic reviews of genetic association studies [24], [27], [34]. Nevertheless, these guidelines should be adapted to the specific characteristics of psychiatric genetics as has recently been suggested for the adaptation of the Meta-analysis of Observational Studies in Epidemiology (MOOSE) guidelines [64] to be used in psychiatric epidemiology [65].
Implications
Although current evidence does not support a direct main effect of the 5-HTTLPR polymorphisms on PTSD, GxE interactions and epigenetic modulation offer promising research areas in the near future. Further studies of possible genetic associations between 5-HTTLPR and PTSD are needed to clarify this relationship and the role of the potential moderators detected. Special attention should be paid to some characteristics specifically in relation to PTSD research, including exposure of controls to the same stressor as PTSD cases, type of PTSD assessed, psychiatric comorbidity and population stratification. These new studies, in addition to those already published on the effect of GxE interactions and on new potential environmental factor candidates to interact with the 5-HTTLPR polymorphisms, may help to clarify the role of this and other genetic factors in PTSD.
Acknowledgments
We thank Carmen Navarro Sánchez, Juan Antonio Sánchez Sánchez and Salvador Martínez Pérez for their collaboration in the initial phases of this project and Mike Tobin for his helpful discussions and contribution during the English translation of the document. The contribution of data from the investigators of the original studies included in this meta-analysis is gratefully acknowledged.
Funding Statement
This work has been financed by the Regional Health Authorities of Murcia (Consejería de Sanidad y Política Social and Servicio Murciano de Salud) and the Fundación para la Formación e Investigación Sanitaria (FFIS) of the Murcia Region. The funding agencies had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript. Jim van Os
References
- 1. Kessler RC (2000) Posttraumatic stress disorder: the burden to the individual and to society. J Clin Psychiatry 61 Suppl 54–12. [PubMed] [Google Scholar]
- 2. Green BL (1994) Psychosocial research in traumatic stress: an update. J Trauma Stress 7: 341–362. [DOI] [PubMed] [Google Scholar]
- 3. Koenen KC (2007) Genetics of posttraumatic stress disorder: Review and recommendations for future studies. J Trauma Stress 20: 737–750 10.1002/jts.20205 [doi] [DOI] [PubMed] [Google Scholar]
- 4. Koenen KC, Amstadter AB, Nugent NR (2009) Gene-environment interaction in posttraumatic stress disorder: an update. J Trauma Stress 22: 416–426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Caspi A, Hariri AR, Holmes A, Uher R, Moffitt TE (2010) Genetic sensitivity to the environment: the case of the serotonin transporter gene and its implications for studying complex diseases and traits. Am J Psychiatry 167: 509–527 appi.ajp.2010.09101452 [pii];10.1176/appi.ajp.2010.09101452 [doi] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Caspi A, Sugden K, Moffitt TE, Taylor A, Craig IW, et al. (2003) Influence of life stress on depression: moderation by a polymorphism in the 5-HTT gene. Science 301: 386–389. [DOI] [PubMed] [Google Scholar]
- 7. Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, et al. (1996) Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274: 1527–1531. [DOI] [PubMed] [Google Scholar]
- 8. Li D, He L (2007) Meta-analysis supports association between serotonin transporter (5-HTT) and suicidal behavior. Mol Psychiatry 12: 47–54. [DOI] [PubMed] [Google Scholar]
- 9. Lotrich F, Pollock BG (2004) Meta-analysis of serotonin transporter polymorphisms and affective disorders. Psychiatr Genet 14: 121–129. [DOI] [PubMed] [Google Scholar]
- 10. Takano A, Arakawa R, Hayashi M, Takahashi H, Ito H, et al. (2007) Relationship between neuroticism personality trait and serotonin transporter binding. Biol Psychiatry 62: 588–592. [DOI] [PubMed] [Google Scholar]
- 11.Hu X, Oroszi G, Chun J, Smith TL, Goldman D, et al. (2005) An expanded evaluation of the relationship of four alleles to the level of response to alcohol and the alcoholism risk. Alcohol Clin Exp Res 29: 8–16. 00000374–200501000–00002 [pii]. [DOI] [PubMed]
- 12. Kilpatrick DG, Koenen KC, Ruggiero KJ, Acierno R, Galea S, et al. (2007) The serotonin transporter genotype and social support and moderation of posttraumatic stress disorder and depression in hurricane-exposed adults. Am J Psychiatry 164: 1693–1699 164/11/1693 [pii];10.1176/appi.ajp.2007.06122007 [doi] [DOI] [PubMed] [Google Scholar]
- 13. Nakamura M, Ueno S, Sano A, Tanabe H (2000) The human serotonin transporter gene linked polymorphism (5-HTTLPR) shows ten novel allelic variants. Mol Psychiatry 5: 32–38. [DOI] [PubMed] [Google Scholar]
- 14. Zalsman G, Huang YY, Oquendo MA, Burke AK, Hu XZ, et al. (2006) Association of a triallelic serotonin transporter gene promoter region (5-HTTLPR) polymorphism with stressful life events and severity of depression. Am J Psychiatry 163: 1588–1593. [DOI] [PubMed] [Google Scholar]
- 15. Parsey RV, Hastings RS, Oquendo MA, Hu X, Goldman D, et al. (2006) Effect of a Triallelic Functional Polymorphism of the Serotonin-Transporter-Linked Promoter Region on Expression of Serotonin Transporter in the Human Brain. American Journal of Psychiatry 163: 48–51. [DOI] [PubMed] [Google Scholar]
- 16. Koenen KC, Aiello AE, Bakshis E, Amstadter AB, Ruggiero KJ, et al. (2009) Modification of the association between serotonin transporter genotype and risk of posttraumatic stress disorder in adults by county-level social environment. Am J Epidemiol 169: 704–711 kwn397 [pii];10.1093/aje/kwn397 [doi] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Kolassa IT, Ertl V, Eckart C, Glockner F, Kolassa S, et al. (2010) Association study of trauma load and SLC6A4 promoter polymorphism in posttraumatic stress disorder: evidence from survivors of the Rwandan genocide. J Clin Psychiatry 71: 543–547 10.4088/JCP.08m04787blu [doi] [DOI] [PubMed] [Google Scholar]
- 18. Lee HJ, Lee MS, Kang RH, Kim H, Kim SD, et al. (2005) Influence of the serotonin transporter promoter gene polymorphism on susceptibility to posttraumatic stress disorder. Depress Anxiety 21: 135–139 10.1002/da.20064 [doi] [DOI] [PubMed] [Google Scholar]
- 19. Grabe HJ, Spitzer C, Schwahn C, Marcinek A, Frahnow A, et al. (2009) Serotonin transporter gene (SLC6A4) promoter polymorphisms and the susceptibility to posttraumatic stress disorder in the general population. Am J Psychiatry 166: 926–933 appi.ajp.2009.08101542 [pii];10.1176/appi.ajp.2009.08101542 [doi] [DOI] [PubMed] [Google Scholar]
- 20. Xie P, Kranzler HR, Poling J, Stein MB, Anton RF, et al. (2009) Interactive effect of stressful life events and the serotonin transporter 5-HTTLPR genotype on posttraumatic stress disorder diagnosis in 2 independent populations. Arch Gen Psychiatry 66: 1201–1209 66/11/1201 [pii];10.1001/archgenpsychiatry.2009.153 [doi] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Mellman TA, Brown DD, Jenifer ES, Hipolito MM, Randall OS (2009) Posttraumatic stress disorder and nocturnal blood pressure dipping in young adult African Americans. Psychosom Med 71: 627–630 PSY.0b013e3181a54341 [pii];10.1097/PSY.0b013e3181a54341 [doi]. [DOI] [PubMed] [Google Scholar]
- 22. Koenen KC, Nugent NR, Amstadter AB (2008) Gene-environment interaction in posttraumatic stress disorder: review, strategy and new directions for future research. Eur Arch Psychiatry Clin Neurosci 258: 82–96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Nugent NR, Amstadter AB, Koenen KC (2008) Genetics of post-traumatic stress disorder: informing clinical conceptualizations and promoting future research. Am J Med Genet C Semin Med Genet 148C: 127–132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Sagoo GS, Little J, Higgins JP (2009) Systematic Reviews of Genetic Association Studies. Human Genome Epidemiology Network. PLoS Med 6: e28 10.1371/journal.pmed.1000028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Gluud LL (2006) Bias in clinical intervention research. Am J Epidemiol 163: 493–501 kwj069 [pii];10.1093/aje/kwj069 [doi] [DOI] [PubMed] [Google Scholar]
- 26.Little J, Higgins JP, Ioannidis JP, Moher D, Gagnon F, et al.. (2009) STrengthening the REporting of Genetic Association studies (STREGA): an extension of the STROBE Statement. Ann Intern Med 150: 206–215. 150/3/206 [pii]. [DOI] [PubMed]
- 27. Hirschhorn JN, Lohmueller K, Byrne E, Hirschhorn K (2002) A comprehensive review of genetic association studies. Genet Med 4: 45–61. [DOI] [PubMed] [Google Scholar]
- 28. Little J, Bradley L, Bray MS, Clyne M, Dorman J, et al. (2002) Reporting, appraising, and integrating data on genotype prevalence and gene-disease associations. Am J Epidemiol 156: 300–310. [DOI] [PubMed] [Google Scholar]
- 29. Sanchez-Meca J, Marin-Martinez F (2008) Confidence intervals for the overall effect size in random-effects meta-analysis. Psychol Methods 13: 31–48 2008–02525–003 [pii];–10.1037/1082–989X.13.1.31 [doi] [DOI] [PubMed] [Google Scholar]
- 30.Huedo-Medina T, Sánchez-Meca J, Marín-Martínez F, Botella J (2006) Assessing heterogeneity in meta-analysis: Q statistic or I2 index? CHIP Documents Paper 19. [DOI] [PubMed]
- 31. Duval S, Tweedie R (2000) Trim and fill: A simple funnel-plot-based method of testing and adjusting for publication bias in meta-analysis. Biometrics 56: 455–463. [DOI] [PubMed] [Google Scholar]
- 32.Sterne JAC, Egger M (2005) Regression methods to detect publication and other bias in meta-analysis. In: Rothstein HR, Sutton AJ, Borestein M, editors. Publication bias in meta-analysis: Prevention, assessment and adjustments. Chichester, UK: Wiley. 99–100.
- 33.Borenstein M, Hedges L, Higgins J, Rothstein H (2005) Comprehensive Meta-Analysis, Version 2, version Englewood, NJ: Biostat.
- 34.Bray M, Higgins J, Ioannidis J, Khoury M, Little J, et al. (2006) The HuGENetTM HUGE Review Handbook, version 1.0.
- 35. Mushtaq D, Ali A, Margoob MA, Murtaza I, Andrade C (2011) Association between serotonin transporter gene promoter-region polymorphism and 4- and 12-week treatment response to sertraline in posttraumatic stress disorder. J Affect Disord 136: 955–62 10.1016/j.jad.2011.08.033. [DOI] [PubMed] [Google Scholar]
- 36. Bryant RA, Felmingham KL, Falconer EM, Pe BL, Dobson-Stone C, et al. (2010) Preliminary evidence of the short allele of the serotonin transporter gene predicting poor response to cognitive behavior therapy in posttraumatic stress disorder. Biol Psychiatry 67: 1217–1219. [DOI] [PubMed] [Google Scholar]
- 37. Wang ZW, Harrer J, Tuerk P, Acierno R, Hamner M, et al. (2009) 5-HTTLPR Influence PTSD Treatment Outcome. Biol Psychiatry 65: 119S. [Google Scholar]
- 38. Beach SRH, Brody GH, Todorov AA, Gunter TD, Philibert RA (2010) Methylation at SLC6A4 is linked to family history of child abuse: An examination of the Iowa adoptee sample. Am J Med Genet Part B Neuropsychiatr Genet 153: 710–713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Holman EA, Lucas-Thompson RG, Lu T (2011) Social constraints, genetic vulnerability, and mental health following collective stress. J Trauma Stress 24: 497–505. [DOI] [PubMed] [Google Scholar]
- 40. Kaufman J (2009) Genes, environment, and development of affect processing and brain reward circuitry. Biol Psychiatry 65: 106S. [Google Scholar]
- 41. Mercer KB, Orcutt HK, Quinn JF, Fitzgerald CA, Conneely KN, et al. (2011) Acute and Posttraumatic Stress Symptoms in a Prospective Gene x Environment Study of a University Campus Shooting. Arch Gen Psychiatry 69: 89–97 10.1001/archgenpsychiatry.2011.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Koenen K, Kilpatrick D, Acierno R, Ruggiero K, Galea S, et al. (2006) Serotonin transporter gene and social support moderate post-traumatic stress disorder in hurricane exposed adults. American Journal of Medical Genetics Part B-Neuropsychiatric Genetics 141B: 742–743. [Google Scholar]
- 43. King AP, Garfinkel SN, Wang X, Kaufman R, Sripada C, et al. (2010) Neurocircuitry of emotional regulation in OEF/ OIF veterans with PTSD: Effects of diagnosis and 5-HTTLPR genotype. Biol Psychiatry 67: 209S. [Google Scholar]
- 44. King AP, Garfinkel SN, Sripada R, Wang X, Taylor AB, et al. (2011) Neurocircuitry of emotional regulation in returning OEF/OIF veterans with PTSD: Effects of diagnosis and genotype. Biol Psychiatry 69: 74S. [Google Scholar]
- 45. Wang Z, Baker DG, Harrer J, Hamner M, Price M, et al. (2011) The relationship between combat-related posttraumatic stress disorder and the 5-HTTLPR/rs25531 polymorphism. Depress Anxiety 28: 1067–1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Valente NL, Vallada H, Cordeiro Q, Miguita K, Bressan RA, et al. (2011) Candidate-gene approach in posttraumatic stress disorder after urban violence: association analysis of the genes encoding serotonin transporter, dopamine transporter, and BDNF. J Mol Neurosci 44: 59–67. [DOI] [PubMed] [Google Scholar]
- 47. Thakur GA, Joober R, Brunet A (2009) Development and persistence of posttraumatic stress disorder and the 5-HTTLPR polymorphism. J Trauma Stress 22: 240–243. [DOI] [PubMed] [Google Scholar]
- 48. Sayin A, Kucukyildirim S, Akar T, Bakkaloglu Z, Demircan A, et al. (2010) A prospective study of serotonin transporter gene promoter (5-HTT gene linked polymorphic region) and intron 2 (variable number of tandem repeats) polymorphisms as predictors of trauma response to mild physical injury. DNA Cell Biol 29: 71–77. [DOI] [PubMed] [Google Scholar]
- 49. Morey RA, Hariri AR, Gold AL, Hauser MA, Munger HJ, et al. (2011) Serotonin transporter gene polymorphisms and brain function during emotional distraction from cognitive processing in posttraumatic stress disorder. BMC Psychiatry 11: 76 10.1186/1471–244X–11–76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Koenen KC, Uddin M, Chang SC, Aiello AE, Wildman DE, et al. (2011) SLC6A4 methylation modifies the effect of the number of traumatic events on risk for posttraumatic stress disorder. Depress Anxiety 28: 639–647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Xie P, Kranzler HR, Farrer L, Gelernter J (2012) Serotonin transporter 5-HTTLPR genotype moderates the effects of childhood adversity on posttraumatic stress disorder risk: a replication study. Am J Med Genet B Neuropsychiatr Genet 159B: 644–652 10.1002/ajmg.b.32068 [doi] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. King AP, Seng J, Taylor A, Palabindala V, Sperlich M, et al. (2009) Effects of Childhood Adversity, Socioeconomic Risk, and Triallelic 5-HTTLPR Genotype on PTSD, Neuroticism, and Cortisol in a Cohort of Detroit-Area Trauma-Exposed Mothers. Biol Psychiatry 65: 6S. [Google Scholar]
- 53.Handwerger, K L. (2010) The neural correlates of emotional memory in posttraumatic stress disorder and the mediating role of genetic influences [dissertation]. US: ProQuest Information & Learning.
- 54. von EE, Altman DG, Egger M, Pocock SJ, Gotzsche PC, et al. (2007) The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: guidelines for reporting observational studies. PLoS Med 4: e296 07-PLME-RA-0728 [pii];10.1371/journal.pmed.0040296 [doi]. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Purcell SM, Wray NR, Stone JL, Visscher PM, O'Donovan MC, et al. (2009) Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 460: 748–752 nature08185 [pii];10.1038/nature08185 [doi] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Clarke H, Flint J, Attwood AS, Munafo MR (2010) Association of the 5- HTTLPR genotype and unipolar depression: a meta-analysis. Psychol Med 40: 1767–1778 S0033291710000516 [pii];10.1017/S0033291710000516 [doi] [DOI] [PubMed] [Google Scholar]
- 57. Cho HJ, Meira-Lima I, Cordeiro Q, Michelon L, Sham P, et al. (2005) Population-based and family-based studies on the serotonin transporter gene polymorphisms and bipolar disorder: a systematic review and meta-analysis. Mol Psychiatry 10: 771–781. [DOI] [PubMed] [Google Scholar]
- 58. Feinn R, Nellissery M, Kranzler HR (2005) Meta-analysis of the association of a functional serotonin transporter promoter polymorphism with alcohol dependence. Am J Med Genet B Neuropsychiatr Genet 133B: 79–84. [DOI] [PubMed] [Google Scholar]
- 59. Uher R, McGuffin P (2010) The moderation by the serotonin transporter gene of environmental adversity in the etiology of depression: 2009 update. Mol Psychiatry 15: 18–22. [DOI] [PubMed] [Google Scholar]
- 60. Terracciano A, Tanaka T, Sutin AR, Deiana B, Balaci L, et al. (2010) BDNF Val66Met is associated with introversion and interacts with 5-HTTLPR to influence neuroticism. Neuropsychopharmacology 35: 1083–1089 npp2009213 [pii];10.1038/npp.2009.213 [doi] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Munafo MR, Clark TG, Roberts KH, Johnstone EC (2006) Neuroticism mediates the association of the serotonin transporter gene with lifetime major depression. Neuropsychobiology 53: 1–8. [DOI] [PubMed] [Google Scholar]
- 62. Toyokawa S, Uddin M, Koenen KC, Galea S (2012) How does the social environment 'get into the mind'? Epigenetics at the intersection of social and psychiatric epidemiology. Soc Sci Med 74: 67–74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63. Esteller M (2006) The necessity of a human epigenome project. Carcinogenesis 27: 1121–1125 bgl033 [pii];10.1093/carcin/bgl033 [doi]. [DOI] [PubMed] [Google Scholar]
- 64.Stroup DF, Berlin JA, Morton SC, Olkin I, Williamson GD, et al.. (2000) Meta-analysis of observational studies in epidemiology: a proposal for reporting. Meta-analysis Of Observational Studies in Epidemiology (MOOSE) group. JAMA 283: 2008–2012. jst00003 [pii]. [DOI] [PubMed]
- 65. Brugha TS, Matthews R, Morgan Z, Hill T, Alonso J, et al. (2012) Methodology and reporting of systematic reviews and meta-analyses of observational studies in psychiatric epidemiology: systematic review. Br J Psychiatry 200: 446–453 200/6/446 [pii];10.1192/bjp.bp.111.098103 [doi]. [DOI] [PubMed] [Google Scholar]