

Abstract
Rexoid, curly hair mutations have been selected to develop new domestic cat breeds. The Selkirk Rex is the most recently established curly-coated cat breed originating from a spontaneous mutation that was discovered in the United States in 1987. Unlike the earlier and well-established Cornish and Devon Rex breeds with curly-coat mutations, the Selkirk Rex mutation is suggested as autosomal dominant and has a different curl phenotype. This study provides a genetic analysis of the Selkirk Rex breed. An informal segregation analysis of genetically proven matings supported an autosomal, incomplete dominant expression of the curly trait in the Selkirk Rex. Homozygous curl cats can be distinguished from heterozygous cats by head and body type, as well as the presentation of the hair curl. Bayesian clustering of short tandem repeat (STR) genotypes from 31 cats that represent the future breeding stock supported the close relationship of the Selkirk Rex to the British Shorthair, Scottish Fold, Persian, and Exotic Shorthair, suggesting the Selkirk as part of the Persian breed family. The high heterozygosity of 0.630 and the low mean inbreeding coefficient of 0.057 suggest that Selkirk Rex has a diverse genetic foundation. A new locus for Selkirk autosomal dominant Rex, SADRE, is suggested for the curly trait.
Key words: curly, dominant, feline, hair
Rexoid or curly pelage types have been identified in many mammalian species including; mice (Dunn 1937; Taylor et al. 2000; Runkel et al. 2006), rats (Robinson 1981; Kuramoto et al. 2010), horses (Scott 2004), rabbits (Diribarne et al. 2011), and dogs (Cadieu et al. 2009), often defining their domesticated breeds and varieties. Most feline breeds were developed for strictly aesthetic purposes and often one simple genetic variety, such as long hair or curly hair, is all that is required to demarcate a new cat breed. As a result, several rexoid—curly pelage mutations have been documented in the domestic cat, including Cornish Rex (Searle and Jude, 1956), Devon Rex (Robinson 1969), German Rex (Robinson 1968), and Oregon Rex (Robinson 1972). The causative mutations in the three rexoid mutants that have been successfully developed into competitive breeds, the Cornish, Devon, and German Rexes, have been identified (Gandolfi et al. 2010 and Gandolfi B et al. unpublished data), and Selkirk Rex is proven to be non-allelic in the respective causative genes, P2RY5 and KRT71, as no Selkirk Rex–specific mutations were identified. Two additional recent rexoid types, the American Wirehair and LaPerm, have been developed into breeds but neither has been scientifically documented. As proven by sequence analysis, the American Wirehair and LaPerm rexoid mutants are also non-allelic to Cornish, Devon, and German Rex but no association with Selkirk Rex is yet proven. These three mutants could be allelic or variants at different genes, Selkirk Rex and LaPerm being the most phenotypically similar and both considered as autosomal dominant mutations.
The Selkirk Rex curly coat is the most recent mutation of any type in the domestic cat fancy that has successfully been developed into an internationally recognized breed. According to lay literature, the breed originated in 1987 from a domestic shorthair cat that was rescued from a shelter in Sheridan, MT, USA (Bass D, personal communication). The rescued cat produced a curly-coated female kitten that was mated with a Persian male, resulting in curly- and normal-haired kittens. This, as well as additional matings, suggested an autosomal dominant mode of inheritance. The Selkirk Rex breed was first recognized by major cat associations in the USA in the early 1990s (Morris 1999; TICA 2010; Weaver 2010). Today, registered Selkirk Rex breeders and their cats are found around the globe, including Europe, Australia, Russia, the Far and Middle East, and South Africa.
Previous studies have used microsatellites, short tandem repeats (STRs), and single-nucleotide polymorphisms (SNPs) to genetically demarcate breeds and their relationships to random-bred populations (Lipinski et al. 2008; Menotti-Raymond et al. 2008; Kusushima 2011). Despite the Selkirk Rex’s worldwide recognition, the genetic distinctness of this newly formed breed of cat has not been established, currently perhaps a variety of Persian or British Shorthair. The state of the genetic diversity of the breed will help Selkirk Rex breeders to better plan continued breed management programs. The objectives of this study are to: 1) characterize the mode of inheritance of the curly trait, and 2) access the breed’s genetic diversity, distinction, and population structure. An informal segregation analysis and STR data were generated and compared with existing data from various cat breeds (Lipinski et al. 2008) to establish the genetic baseline of the Selkirk Rex breed.
Materials and Methods
Studied Populations
Buccal samples from pedigreed Selkirk Rex were solicited from breeders, which were collected using sterile cotton swabs. Breeders were encouraged to submit pedigrees and samples from complete litters, including both parents and all kittens. The curly-coat phenotypes, including straight, curly homozygous, or curly heterozygous, were determined by visual inspection by the owners. Cats were assumed to be homozygous for their curl in accordance with their breeders’ impression of having tighter, softer hair, an episode of massive hair loss in early age, and a more slender body composition with bigger ears and a generally less rounded appearance to the head.
Genotyping
Genomic DNA was isolated from buccal swabs using Nexttec (Nexttec GmbH Biotechnologie, Larezhausen, Germany) DNA isolation system, following the manufacturer’s instructions except with 200 µL lysis buffer and incubating at 60 °C for 5h. Thirty-eight microsatellites were genotyped and analyzed as in previous studies (Lipinski et al. 2007, 2008). However, forward primers were fluorescently labeled with 6-FAM, HEX, and VIC dyes, exchanging VIC/PET with HEX and HEX with VIC labels. Genotyping polymerase chain reaction was conducted on MWG Primus 96 thermal cyclers (Eurofins MWG Operon, Ebersberg, Germany) following the previously published thermocycling conditions (Lipinski et al. 2008).
Population Statistics
A subset of 31 cats unrelated to the grand-parents level was used for a population assessment study. All cats were either intact or altered siblings of the actively breeding individuals. The population summary statistics of Hardy–Weinberg equilibrium (HWE), inbreeding coefficient (F IS), and observed and expected heterozygosities were calculated using GenAlEx v.6.3 (Peakall and Smouse 2006). Previously published STR data (Lipinski et al. 2008; Kurushima 2011) from other breeds were used for comparison.
Population Structure Analysis
Data sets were analyzed with the Bayesian clustering program STRUCTURE (Pritchard et al. 2000) under the admixture model with correlated allele frequencies and a burn-in of 10 000 with 100 000 additional iterations. For worldwide breed assessment, 30 breeds (N = 786) were used and values of K were calculated from K = 1 to K = 6, each run was replicated five times. For Western breed analysis, 19 breeds (n = 500) were included and values of K were calculated from K = 1 to K = 19, each run was replicated five times. Posterior log likelihoods were used in the calculation of ∆K to best estimate the number of ancestral populations through the program Harvester (Evanno et al. 2005). All five iterations were then combined through the program CLUMP (Jakobsson and Rosenberg 2007) to create a consensus clustering. The structure images were generated using the software DISTRUCT (Rosenberg 2004).
Principal components analyses (PCA) were conducted through the calculation of Nei’s genetic distance using the software GenAlEx v.6.3 (Peakall and Smouse 2006). For the PCA plots, both the data in the present manuscript and data from previously studied cat breeds and the worldwide random-bred populations were considered to show the relationship among cat breeds and their random-bred population origins (Lipinski et al. 2008; Kurushima 2011).
Results and Discussion
Selkirk Rex Breed, Curly Phenotype, and Morphology
The Selkirk Rex is a medium-build cat breed with a round, sturdy body type, wide-set eyes and ears, and a moderately brachycephalic muzzle (Figure 1a). Recent registry census suggest Selkirk Rex has a small but growing breed population with 1581 cats being registered with The International Cat Registry (TICA Executive Office, personal communication), 1371 cats registered with Cat Fanciers’ Association (CFA 2011), and 1291 cats registered with the Governing Council of the Cat Fancy (GCCF 2011). All genetically possible coat colors, patterns, and lengths are allowable in the breed. Selkirk Rex cats are curly at birth (see Supplementary Figure 1b online) and can easily be differentiated from their straight-haired littermates by their curled whiskers (see Supplementary Figure 1a online). The whiskers tend to break easily and generally stay short into adulthood (Figure 1a,b). The desired heterozygous cats exhibit a full randomly curled coat (Figure 1a,c) whereas homozygous Selkirk Rex cats have a tight, soft, and wavy curl, similar to the Cornish Rex coat (Figure 1b,d). The Selkirk Rex has allowable outcrosses with Persians, Exotic Shorthairs, British Shorthairs, and British Longhair cats, depending on the specific cat registry regulations. The length, density, and rigidity of the coat reflect the recent outcross to other breeds. Additionally, homozygous cats for the Selkirk Rex curl tend to lose their coat as kittens but usually neither develop nor exhibit bald areas of skin later in their lives. Cats assumed to be homozygous for the curly trait appear to share a more slender body composition and bigger ears (Figure 1b). Generally, breeders prefer the heterozygous Selkirk Rex as they maintain the random thick curls and the round head morphology, which is in accordance with the written standards. Stiff hair types tend to exhibit less curls than soft types, which are more preferred by breeders. As in sheep and other wool and fleece animals, a measure of hair crimp and fiber diameter may provide an objective measure for Selkirk Rex hair types and subsequent selection.
Figure 1.

Phenotypes of the Selkirk Rex. (a) Adult heterozygous Selkirk Rex male, short-haired variety. The head morphology and ear type of the heterozygous cat are more desired. (b) Adult homozygous Selkirk Rex. The head morphology and bigger ears of a homozygous curled cat are not desired by the breed standard. (c) Desired curly coat of a heterozygous curled cat. (d) Not desired curly coat of a homozygous cat is more similar to Cornish Rex marcel wave.
Segregation Analysis
A total of 129 DNA-sampled cats and 24 non-sampled cats formed 30 litters, including parents, to evaluate the inheritance of the curly-coated trait (Table 1). Eleven cats were suspected to be homozygous curly, 90 to be heterozygous curly, and 52 to be straight coated. Homozygous curly to homozygous curly type matings, litters sired by homozygous curly males, and registered straight hair (wild-type) litters were not available. All sampled kittens were parentage confirmed by genotyping. Average litter size was four kittens, ranging from two to seven kittens. Interestingly, the average litter for the outcross litters (n = 15), including straight to heterozygous or homozygous curly, was 4.53 kittens, whereas litters that could produce homozygous curly cats (n = 15) had an average of 3.13 kittens. No significant sex-bias was observed in the outcrosses, 35 male and 33 females (P < 0.05), or its gender distribution, 20 males, 18 females. Fifteen matings of curly to wild-type straight-haired cats produced 38 curly- and 30 straight-coated offspring with none of the kittens matching the morphology of homozygous Selkirk Rex. Curly males bred to straight females produced curly females, excluding a Y-linked inheritance, and these matings produced both curly- and straight-coated females, excluding an X-linked mode of inheritance. Thirteen matings of heterozygous to heterozygous cats as defined by their phenotypes produced 26.83% (n = 11) straight-coated and 73.17% (n = 30) curly-coated kittens, as expected for a dominant trait. However, of the curly kittens, 23.33% (n = 7) had morphology indicative of cats homozygous for the curly trait, suggesting either a bias towards heterozygous cats or a misrepresentation of curl type, particularly in males since male heterozygote cats seemed to be overproduced. No straight to straight-coated mating produced curly-coated cats (data not shown). Three matings with one parent being homozygous for curl produced 100% curly kittens.
Table 1 .
Segregation analysis of curly trait in the Selkirk Rex breed
| Curly | ||||||||||
| Mating type | ho | ht | Straight | Total | ||||||
| Sire | Queen | Matings | M | F | M | F | M | F | Male | Female |
| Curly ht | Straight | 8 | 0 | 0 | 13 | 10 | 6 | 10 | 19 | 20 |
| Straight | Curly ht | 6 | 0 | 0 | 4 | 7 | 9 | 5 | 13 | 12 |
| Straight | Curly ho | 1 | 0 | 0 | 3 | 1 | 0 | 0 | 3 | 1 |
| Total | 15 | 0 | 0 | 20 | 18 | 15 | 15 | 35 | 33 | |
| Curly ht | Curly ht | 13 | 1 | 6 | 16 | 7 | 6 | 5 | 23 | 18 |
| Curly ht | Curly ho | 2 | 1 | 0 | 3 | 2 | 0 | 0 | 4 | 2 |
| Total | 15 | 2 | 6 | 19 | 9 | 6 | 5 | 27 | 20 | |
| All total | 30 | 2 | 6 | 39 | 27 | 21 | 20 | 62 | 53 | |
M, male; F, female; Curly cats were considered to be either heterozygous (ht) or homozygous (ho) for their hair curl by phenotype. Unavailable mating types are not shown.
An extended pedigree of 55 cats within the 220 sampled could be constructed (Figure 2). All curly-coated cats in the pedigree have at least one curly-coated parent. Both male and female parents produce male and female curly-coated offspring. Nine curly male to curly female matings (three shown in pedigree) produced straight-haired kittens, excluding a recessive mode of inheritance. Curly females and curly males produced curly offspring, male and female, when mated to a straight-haired cat, excluding a sex-linked mode of inheritance (see Supplementary Figure 2 online). Overall, the data support an autosomal incomplete dominant mode of inheritance. As no formal nomenclature committee exists for the cat, mouse and human conversions were adopted and the locus name, SADRE, is suggested to represent the Selkirk autosomal dominant rex mutation. Based on 20 available pedigrees, the mutation age is estimated to be an average of 8.4 generations from the original cat with the curly mutation (data not shown), which is consistent with the 1987 mutation origin, suggesting that the breed has 2.8 years per generation. Although cats can be sexually active and conceive litters as early as 6 months, show cats can be presented as kittens up to 1 year of age, therefore not active breeders. Also, considering that not all cats contribute to the breeding pool and some lines are abandoned, the estimated mutation age and generation time seems a reasonable estimate.
Figure 2.
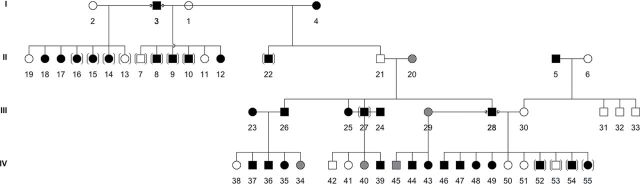
Pedigree of Selkirk Rex cats segregating for curled hair type. Cats with filled symbols are heterozygous curled; cats with stippled symbols are homozygous curled. Cats in parentheses did not have their DNA sampled and have unconfirmed parentage, however completed the represented litters.
Population Statistics
The phenotype for the 31 unrelated Selkirk Rex cats and genotypes for the 38 STRs are presented in Supplementary Table 2 online. Three markers significantly deviated from HWE (P < 0.001) but were considered in analysis. At the threshold frequency of 0.01, one unique allele to the Selkirk Rex breed was identified (FCA223, allele 213). The average number of alleles per loci was 5.935, ranging from 2 to 9. Average allelic richness was 3.04. The Selkirk Rex has an observed heterozygosity (Ho) of 0.630, an expected heterozygosity (He) of 0.654, and a mean inbreeding coefficient (F IS) of 0.057. Locus-specific values are shown in Supplementary Table 3 online. In comparison to published diversity data, the Selkirk Rex breed is a diverse breed. The observed heterozygosity and number of alleles per locus of the Selkirk Rex were similar to those of the Norwegian Forest Cat, Siberian, and Japanese Bobtail breeds, as well as most random-bred populations. Additionally, the inbreeding coefficient is low, suggesting that the current breeding population is genetically diverse and rich.
The Bayesian clustering and PCA are also in agreement with the Selkirk Rex breeding history, grouping the Selkirk Rex with the lineage of Western breeds (Figure 3a). The Selkirk Rex formed a tight grouping with British Shorthairs, Scottish Fold, Persian, and Exotic Shorthairs when only Western breeds were analyzed (Figure 3b). A direct examination of the Persian family group indicated that Selkirk Rex cats have strong influences from both Persian and British Shorthair breeds, the current population having a slightly stronger influence from British Shorthair (Figure 3b). The association with the British Shorthair is concordant with the subjective observation that the Selkirk Rex cats have an overall round body shape and more closely resembles the British Shorthair than the Persian breed. Principal coordinate analysis was concordant with Bayesian clustering, not differentiating the Selkirk Rex from the British Shorthair, Persian, Exotic Shorthair, and Scottish fold breeds (Figure 3c). PCA was also performed within the Selkirk Rex breed, showing a division between European and US outcrossed cats. Selkirk Rex cats from the UK clustered with Persian cats from the USA, whereas cats from Russia were closer to British Shorthairs bred on continental Europe (Figure 3d).
Figure 3.

Population genetics assessment of the Selkirk Rex breed. (a) Bayesian clustering of 30 recognized cat breeds at K = 3. The Selkirk Rex clusters within the Western I group. Sokoke and Turkish Van, denoted by * and ** respectively, are breeds that group within Western II at K = 3 but form their distinct groups at higher K values. (b) Bayesian clustering of 19 Western breeds at K = 15. Selkirk Rex clusters with breeds that belong to the Persian family. (c) Principal component analysis of the Persian family breeds. (d) Principal component analysis of the Selkirk Rex breed, which shows the geographic distribution of the breed (BSH: British Shorthair, PER: Persian, ESH: Exotic Short Hair, S. Fold: Scottish Fold, S. Rex: Selkirk Rex).
Spontaneous mutations that cause novel morphological traits often become the selective focus for new domesticated breed development. Many mutations causing hair abnormalities are documented in humans, dogs, and other species (Cadieu et al. 2009; Gandolfi et al. 2010; Kuramoto et al. 2010; Diribarne et al. 2011). Rexoid mutations cause hair abnormalities through a major change in the hair follicle, which leads to hair shaft anomalies. These abnormalities cause a variety of phenotypes such as sparse hair (hypotrichosis), excessive hair (hypertrichosis), and absence of hair (atrichia) (see review Shimomura 2010). Some hair mutants have pleiotrophic effects and are more severe ectodermal dysplasias, such as teeth loss in the Chinese Crested dog breed (Drögemüller et al. 2008). In the cat, other than undesired periodic patches of baldness, occasional seborrhoeic dermatitis, and excessive yeast carriage (Bond et al. 2008; Ahman and Bergström 2009), the cat has no other direct deleterious effects in its curly-coated varieties.
Cat breeds like the Selkirk Rex are valuable populations for genetic studies. Elucidation of the gene controlling the curly hair trait will contribute to research into hair development, follicle formation pathways, and potentially baldness. The past 24 years of breed development that allowed substantial outcrossing with assorted breeds encouraged all color varieties and both long and short hair, and has established a breed with a wide genetic foundation. Thus, the breed history could be used as a model for new cat breed development, especially when the breed defining mutation is dominant. This defined genetic foundation will also allow the breeders to monitor and assess the genetic diversity in the future, supporting a breed management plan that would prevent any loss of variation, which may potentially lead to health concerns. These analyses indicate that the Selkirk Rex has a sufficiently diverse genetic base that may support some limitation to outcrossing, which may be beneficial for the breed in developing a unique body and head morphology. An outcrossing limitation can aid further differentiation between the Selkirk Rex, the Persian, and British Shorthair breeds. Because the Selkirk Rex is so closely related to the Persian breed family, diseases, such as hypertrophic cardiomyopathy (Meurs et al. 2009), polycystic kidney disease (Biller et al. 1990; Eaton et al. 1997; Lyons et al. 2004), and problems associated with a more than moderate degree of brachycephalic, head shape (Gunn-Moore and Thrusfield 1995; Schlueter et al. 2009; Hartley 2010) should be monitored in the Selkirk Rex. The Selkirk Rex breed can potentially serve as a model for new breed development, and an example for efficient and effective breed population management plans through an outcross allowance system.
Supplementary Material
Supplementary material can be found at http:// www.jhered.oxfordjournals.org/.
Funding
This work was supported by Agrobiogen GmbH, Hilgertshausen, Germany; the National Center for Research Resources (R24 RR016094), and is currently supported by the Office of Research Infrastructure Programs (OD R24OD010928) to L.A.L.; the Winn Feline Foundation (grant 10-015); the University of Veterinary Medicine Vienna; the Center for Companion Animal Health; and the Veterinary Genetics Laboratory, School of Veterinary Medicine, University of California–Davis.
Supplementary Material
Acknowledgments
We would like to thank the many Selkirk Rex owners and breeders who so generously contributed samples and time to this study, especially Geneviève Alidarso, Donna Bass, Mary Jones, Dee Marsland-Round, Elena Nikonova, and Claire Winman.
References
- Ahman SE,, Bergström KE. Cutaneous carriage of Malassezia species in healthy and seborrhoeic Sphynx cats and a comparison to carriage in Devon Rex cats. J Feline Med Surg. 2009.;11((12):):970–976. doi: 10.1016/j.jfms.2009.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biller DS,, Chew DJ,, DiBartola SP. Polycystic kidney disease in a family of Persian cats. J Am Vet Med Assoc. 1990.;196::1288–1290. [PubMed] [Google Scholar]
- Bond R,, Stevens K,, Perrins N,, Ahman S. Carriage of Malassezia spp. Yeasts in Cornish Rex, Devon Rex and Domestic short-haired cats: a cross-sectional survey. Vet Dermatol. 2008.;19((5):):299–304. doi: 10.1111/j.1365-3164.2008.00695.x. [DOI] [PubMed] [Google Scholar]
- Cadieu E,, Neff MW,, Quignon P,, Walsh K,, Chase K,, Parker HG,, Vonholdt BM,, Rhue A,, Boyko A,, Byers A, et al. Coat variation in the domestic dog is governed by variants in three genes. Science. 2009.;326::150–153. doi: 10.1126/science.1177808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castle WE. The rex rabbit. J Hered. 1929.;20:(193) [Google Scholar]
- CFA. 2011.. http://secure.cfa.org/cfaAlmanac/Arch-SS/RS/2010_RegistrationStatistics.pdf http://secure.cfa.org/cfaAlmanac/Arch-SS/RS/2010_RegistrationStatistics.pdf Cat Fanciers' Association Registration Statistics 2010. [cited 2012 March 14]. Available from:
- Diribarne M,, Mata X,, Chantry-Darmon C,, Vaiman A,, Auvinet G,, Bouet S,, Deretz S,, Cribiu EP,, de Rochambeau H,, Allain D, et al. A deletion in exon 9 of the LIPH gene is responsible for the rex hair coat phenotype in rabbits (Oryctolagus cuniculus). PLoS One. 2011.;6((4):):e19281. doi: 10.1371/journal.pone.0019281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drögemüller C,, Karlsson EK,, Hytönen MK,, Perloski M,, Dolf G,, Sainio K,, Lohi H,, Lindblad-Toh K,, Leeb T. A mutation in hairless dogs implicates FOXI3 in ectodermal development. Science. 2008.;321((5895):) doi: 10.1126/science.1162525. [DOI] [PubMed] [Google Scholar]
- Dunn LC. Caracul, a dominant mutation. J. Hered. 1937.;28: [Google Scholar]
- Eaton KA,, Biller DS,, DiBartola SP,, Radin MJ,, Wellman ML. Autosomal dominant polycystic kidney disease in Persian and Persian-cross cats. Vet Pathol. 1997.;34::117–126. doi: 10.1177/030098589703400204. [DOI] [PubMed] [Google Scholar]
- Evanno G,, Regnaut S,, Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol. 2005.;14::2611–2620. doi: 10.1111/j.1365-294X.2005.02553.x. [DOI] [PubMed] [Google Scholar]
- Gandolfi B,, Outerbridge CA,, Beresford LG,, Myers JA,, Pimentel M,, Alhaddad H,, Grahn JC,, Grahn RA,, Lyons LA. The naked truth: Sphynx and Devon Rex cat breed mutations in KRT71. . Mamm Genome. 2010.;21::509–515. doi: 10.1007/s00335-010-9290-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GCCF. 2011.. http://www.gccfcats.org/breeds.html http://www.gccfcats.org/breeds.html Recognised breeds and registration policies. [cited 2011 Dec 27]. Available from:
- Gough A,, Thomas A. 2004.. Breed predispositions to disease in dogs and cats. Oxford, UK: Wiley-Blackwell; p. 165 172 [Google Scholar]
- Gunn-Moore DA,, Thrusfield MV. Feline dystocia: prevalence, and association with cranial conformation and breed. Vet Rec. 1995.;136::350–353. doi: 10.1136/vr.136.14.350. [DOI] [PubMed] [Google Scholar]
- Hartley C. Aetiology of corneal ulcers. J Feline Med Surg. 2010.;12::24–35. doi: 10.1016/j.jfms.2009.12.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakobsson M,, Rosenberg NA. CLUMPP: a cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics. 2007.;23::1801–1806. doi: 10.1093/bioinformatics/btm233. [DOI] [PubMed] [Google Scholar]
- Jude AC. The rex—mutant in the cat. Nature. 1953.;172::81–82. doi: 10.1038/172081b0. [DOI] [PubMed] [Google Scholar]
- Kuramoto T,, Hirano R,, Kuwamura M,, Serikawa T. Identification of the Rat rex mutation as a 7-bp deletion at splicing acceptor site of the Krt71 gene. J Vet Med Sci. 2010.;72::909–912. doi: 10.1292/jvms.09-0554. [DOI] [PubMed] [Google Scholar]
- Kurushima J. 2011.. Genetic analysis of domestication patterns in the cat (Felis catus) [Worldwide population structure, and human-mediated breeding patterns both modern and ancient]. Davis (CA): University of California–Davis; p. 21 93 [Google Scholar]
- Lipinski MJ,, Amigues Y,, Blasi M,, Broad TE,, Cherbonnel C,, Cho GJ,, Corley S,, Daftari P,, Delattre DR,, Dileanis S, et al. An international parentage and identification panel for the domestic cat (Felis catus). Anim Genet. 2007.;38::371–377. doi: 10.1111/j.1365-2052.2007.01632.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipinski MJ,, Froenicke L,, Baysac KC,, Billings NC,, Leutenegger CM,, Levy AM,, Longeri M,, Niini T,, Ozpinar H,, Slater MR, et al. The ascent of cat breeds: genetic evaluations of breeds and worldwide random-bred populations. Genomics. 2008.;91::12–21. doi: 10.1016/j.ygeno.2007.10.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyons LA,, Biller DS,, Erdman CA,, Lipinski MJ,, Young AE,, Roe BA,, Qin B,, Grahn RA. Feline polycystic kidney disease mutation identified in PKD1. . J Am Soc Nephrol. 2004.;15::2548–2555. doi: 10.1097/01.ASN.0000141776.38527.BB. [DOI] [PubMed] [Google Scholar]
- Maas SA,, Suzuki T,, Fallon JF. Identification of spontaneous mutations within the long-range limb-specific Sonic hedgehog enhancer (ZRS) that alter Sonic hedgehog expression in the chicken limb mutants oligozeugodactyly and silkie breed. Dev Dyn. 2011.;240::1212–1222. doi: 10.1002/dvdy.22634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Menotti-Raymond M,, David VA,, Pflueger SM,, Lindblad-Toh K,, Wade CM,, O'Brien SJ,, Johnson WE. Patterns of molecular genetic variation among cat breeds. Genomics. 2008.;91::1–11. doi: 10.1016/j.ygeno.2007.08.008. [DOI] [PubMed] [Google Scholar]
- Meurs KM,, Norgard MM,, Kuan M,, Haggstrom J,, Kittleson M. Analysis of 8 sarcomeric candidate genes for feline hypertrophic cardiomyopathy mutations in cats with hypertrophic cardiomyopathy. J Vet Intern Med. 2009.;23::840–843. doi: 10.1111/j.1939-1676.2009.0341.x. [DOI] [PubMed] [Google Scholar]
- Morris D. 1999.. Cat breeds of the world: a complete illustrated encyclopedia. New York: Viking Penquin; p. 217 118 [Google Scholar]
- Olsson M,, Meadows JR,, Truvé K,, Rosengren Pielberg G,, Puppo F,, Mauceli E,, Quilez J,, Tonomura N,, Zanna G,, Docampo MJ, et al. A novel unstable duplication upstream of HAS2 predisposes to a breed-defining skin phenotype and a periodic fever syndrome in Chinese Shar-Pei dogs. PLoS Genet. 2011.;7::e1001332. doi: 10.1371/journal.pgen.1001332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peakall R,, Smouse PE. GenAlEx 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol. Notes. 2006.;6::288–295. doi: 10.1093/bioinformatics/bts460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pritchard JK,, Stephens M,, Donnelly P. Inference of population structure using multilocus genotype data. Genetics. 2000.;155::945–959. doi: 10.1093/genetics/155.2.945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson R. German Rex: a rexoid coat mutant in the cat. Genetica. 1968.;39::351–352. doi: 10.1007/BF02324474. [DOI] [PubMed] [Google Scholar]
- Robinson R. Devon Rex—a third rexoid coat mutant in cat. Genetica. 1969.;40::597. [Google Scholar]
- Robinson R. Oregon Rex—a fourth rexoid coat mutant in the cat. Genetica. 1972.;43::236–238. doi: 10.1007/BF00123630. [DOI] [PubMed] [Google Scholar]
- Robinson R. Rex mutant in the Norway rat. J Hered. 1981.;72::131–132. doi: 10.1093/oxfordjournals.jhered.a109445. [DOI] [PubMed] [Google Scholar]
- Rosenberg NA. DISTRUCT: a program for the graphical display of population structure. Mol Ecol Notes. 2004.;4::137–138. [Google Scholar]
- Runkel F,, Klaften M,, Koch K,, Bohnert V,, Bussow H,, Fuchs H,, Franz T,, Hrabe de Angelis M. Morphologic and molecular characterization of two novel Krt71 (Krt2-6g) mutations: Krt71rco12 and Krt71rco13. Mamm Genome. 2006.;17::1172–1182. doi: 10.1007/s00335-006-0084-9. [DOI] [PubMed] [Google Scholar]
- Schlueter C,, Budras KD,, Ludewig E,, Mayrhofer E,, Koenig HE,, Walter A,, Oechtering GU. Brachycephalic feline noses. CT and anatomical study of the relationship between head conformation and the nasolacrimal drainage system. J Feline Med Surg. 2009.;11::891–900. doi: 10.1016/j.jfms.2009.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott DW. Skin of the neck, mane and tail of the curly horse. Equine Vet Educ. 2004.;4::201–206. [Google Scholar]
- Searle AG, Jude AC. The rex type of coat in the domestic cat. J genet. (1956);54:506–512. [Google Scholar]
- SelkirkRexCatClub. 2011.. http://www.selkirkrexcatclub.co.uk/Arrival%20to%20UK.htm http://www.selkirkrexcatclub.co.uk/Arrival%20to%20UK.htm Arrival to UK. [cited 2012 March 14]. Available from:
- Shimomura Y. Congenital hair loss disorders: rare, but not too rare. J Dermatol. 2012.;39((1):):3–10. doi: 10.1111/j.1346-8138.2011.01395.x. [DOI] [PubMed] [Google Scholar]
- Taylor LA,, Harris MJ,, Juriloff DM. Whiskers amiss, a new vibrissae and hair mutation near the Krt1 cluster on mouse chromosome 11. Mamm Genome. 2000.;11::255–259. doi: 10.1007/s003350010050. [DOI] [PubMed] [Google Scholar]
- TICA. 2010.. http://www.tica.org/public/breeds/sr/intro.php http://www.tica.org/public/breeds/sr/intro.php TICA Selkirk Rex Introduction. [cited 2012 March 14]. Available from:
- Weaver F. 2010.. http://www.cfa.org/client/breedSelkirkRex.aspx http://www.cfa.org/client/breedSelkirkRex.aspx Selkirk Rex Breed Profile. [cited 2012 March 14]. Available from:
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.