

SUMMARY
Parasitic helminths are ubiquitous in most host, including human, populations. Helminths often alter the likelihood of infection and disease progression of coinfecting microparasitic pathogens (viruses, bacteria, protozoa), and there is great interest in incorporating deworming into control programmes for many major diseases (e.g. HIV, tuberculosis, malaria). However, such calls are controversial; studies show the consequences of deworming for the severity and spread of pathogens to be highly variable. Hence, the benefits of deworming, although clear for reducing the morbidity due to helminth infection per se, are unclear regarding the outcome of coinfections and comorbidities. I develop a theoretical framework to explore how helminth coinfection with other pathogens affects host mortality and pathogen spread and evolution under different interspecific parasite interactions. In all cases the outcomes of coinfection are highly context-dependent, depending on the mechanism of helminth-pathogen interaction and the quantitative level of helminth infection, with the effects of deworming potentially switching from beneficial to detrimental depending on helminth burden. Such context-dependency may explain some of the variation in the benefits of deworming seen between studies, and highlights the need for obtaining a quantitative understanding of parasite interactions across realistic helminth infection ranges. However, despite this complexity, this framework reveals predictable patterns in the effects of helminths that may aid the development of more effective, integrated management strategies to combat pathogens in this coinfected world.
Key words: Parasite interactions, basic reproduction number, treatment, trade-off, Th1–Th2, concurrent infection, virulence, mass drug administration, anthelmintic
INTRODUCTION
Individual hosts, including humans in many communities around the globe, typically harbour chronic helminth infections (Petney and Andrews, 1998; http://www.thiswormyworld.org). These helminths have great potential to alter a multitude of aspects of the within-host environment (e.g. the host's immune response, energetic reserves and utilization strategies and, ultimately, survival), with important implications for the host's ability to fight potentially harmful, coinfecting microparasitic pathogens (e.g. virus, bacteria or protozoa) (Cox, 2001; Graham et al. 2007; Pedersen and Fenton, 2007; Fenton et al. 2008; Graham, 2008; Griffiths et al. 2011). Given this, various papers have suggested that on-going helminth infections may increase susceptibility to pathogen infection and exacerbate the escalation of disease, leading to a number of calls to incorporate deworming into control programmes targeting some of the most serious human pathogens such as HIV, tuberculosis and malaria (Bentwich et al. 1999; Harms and Feldmeier, 2002; Druilhe et al. 2005; Harris et al. 2009). However, such calls have proven controversial, with suggestions that such deworming may increase disease severity in some circumstances (e.g. Nacher, 2006).
Recently a Cochrane review (Taylor-Robinson et al. 2012) reviewed available evidence on the success of deworming programmes, and found great variability in their benefits across and within studies. Again, however these findings were not without controversy; see, for example, Hawkes (2012) for a supportive commentary on this review, and Bundy et al. (2013) for an alternative view. While this review did not specifically examine the use of deworming in the context of coinfection, it raises the possibility that there may be similar levels of variability in the effects of incorporating deworming into pathogen control programmes. What is not clear at present is whether such variability is unpredictable ‘noise’, meaning there would be little hope for generating clear guidelines for coinfection management between different systems or even different locations of the same system, or whether the variability is predictable, arising from (possibly subtle) differences in quantifiable processes between studies. If the latter is the case then there may be hope for understanding the nature of those processes and developing coherent and effective guidelines for the control of pathogens under helminth coinfection.
A major obstacle for understanding how deworming may help the management of pathogenic diseases is the multitude of ways in which helminths can interact with other (non-helminth) microparasitic pathogens. Within-host interactions between helminths and pathogens can alter the likelihood of pathogen establishment, subsequent growth and replication rates, the rate of clearance from the host, disease severity and, ultimately, transmission potential at the population level (e.g. see Petney and Andrews, 1998; Cox, 2001; Griffiths et al. 2011 for reviews). Therefore, understanding the outcome of coinfection is far from straightforward, and a variety of empirical (Lello et al. 2004; Behnke et al. 2005; Jolles et al. 2008; Ezenwa et al. 2010; Telfer et al. 2010) and theoretical (Fenton, 2008; Fenton and Perkins, 2010; Fenton et al. 2010; Ezenwa and Jolles, 2011) approaches have been used to assess the occurrence of parasite interactions in nature, and their possible implications for parasite dynamics. However, previous mathematical models have either not examined how different mechanisms of interspecific interaction alter the outcome of coinfection, or have not considered their effects at both the individual level (host health) and the population level (disease transmission) or have not examined the quantitative effects of helminth burdens on coinfection dynamics. This last point is important because it is quite likely that the magnitude of coinfection effects will vary with helminth burden, meaning that it may not be sufficient to simply consider a host to be coinfected or not, but that it is necessary to consider how heavily that host is infected with helminths. Finally, deworming is rarely 100% effective (Basáñez et al. 2012) so the outcome of a given coinfection, and the benefits (or not) of deworming, could be highly context-dependent, with potentially subtle variations depending on the mechanisms and magnitude of any interspecific interaction between helminths and pathogens, and the pre- and post-treatment helminth burdens. To date these factors have not been accounted for in any theoretical or empirical study of helminth-pathogen coinfection.
Here I use a general theoretical framework to explore the quantitative effects of helminth infection and deworming (the reduction in, but not necessarily elimination of, mean worm burden across a host population) on three aspects of helminth-pathogen coinfection: (1) the expected lifespan of the host following pathogen infection, an individual-level measure, (2) the pathogen's basic reproduction number, a population-level measure, and (3) the evolution of pathogen virulence, all under different forms of within-host interspecific parasite interaction. I show that the effect of the helminth on the pathogen under each scenario can be highly context-dependent, depending not only on the form of interaction between the parasites but also on the quantitative level of helminth infection. However, a better understanding of these processes can lead to insight into the circumstances under which deworming may be either beneficial or detrimental in the context of pathogen coinfection.
MODELLING FRAMEWORK
Baseline epidemiological model
In all that follows I assume a simple host–microparasite–macroparasite system, where ‘microparasite’ refers to a pathogen (e.g. virus, bacteria or protozoa) which causes acute, and potentially severe, infection, replicating within the host before being cleared. The term ‘macroparasite’ refers to a parasitic helminth, which may cause chronic infections and, in what follows, is assumed to be present within the host prior to infection by the microparasite. For clarity I assume the relatively simple scenario of just a single helminth species coinfecting with a single microparasitic pathogen species, with no other species or pathogen strains occurring within the host population. The epidemiological dynamics of this system are described using a hybrid model (Fenton 2008; Fenton et al. 2008), previously developed by amalgamating standard host–microparasite and host–macroparasite frameworks (see Fig. 1 for a schematic representation of the model, Table 1 for definitions of all parameters, and Appendix 1 for model equations). The model tracks changes in the number of hosts that are either susceptible to (S), infected by (I) or recovered from (R) the pathogen (the total host population, H, is given by S+I+R). It is assumed that the helminth infection is unaffected by the presence of the pathogen, and the burden of helminth infection is assumed constant throughout the duration of pathogen infection (Fenton (2008) considers more complex scenarios where the helminth population is dynamic and may be affected by the presence of the pathogen). It should be emphasized that the intention of these analyses is to derive general insight into how different helminth interactions can affect coinfecting pathogens, and so this framework is intentionally generic and is not assumed to match any disease system in particular; while these general predictions and guidelines are highly informative, the model would have to be tailored and parameterized in order to make accurate predictions about any specific system of interest.
Fig. 1.
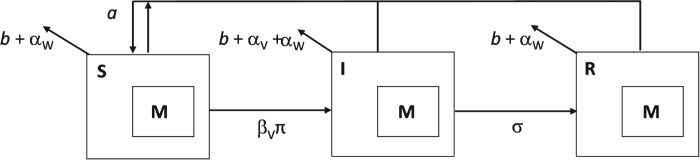
Schematic diagram of the helminth–pathogen co-infection model (see Table 1 for parameter definitions).
Table 1.
Definitions of baseline parameters and state variables
| State variable or parameter | Definition |
|---|---|
| S | Number of hosts susceptibility to the pathogen |
| I | Number of hosts infected by the pathogen |
| R | Number of hosts recovered from pathogen infection |
| M | Mean helminth burden |
| a (time−1) | Host reproduction rate |
| b (time−1) | Background host mortality rate |
| σV (time−1) | Recovery rate from pathogen infection |
| βV (time−1) | Pathogen infection rate (contact component) |
| ∥ | Probability of pathogen infection (host susceptibility component) |
| αV (time−1) | Excess mortality due to pathogen infection (‘virulence’) |
| αW (worm−1 time−1) | Per capita excess mortality due to helminth infection |
Pathogen transmission is determined by the product of three processes: (1) the per capita contact rate between individuals; (2) the level of infectiousness of those infected hosts (e.g. the level of shedding of infectious particles), which together are subsumed into a single parameter βV (where the subscript ‘V’ is used throughout for ‘virus’ (although the same framework applies for other microparasites such as bacteria or protozoa) to denote parameters specific to the pathogen); and (3) the probability of transmission given contact between a susceptible and infected individual (π; a measure of host susceptibility to infection). Hence βV determines the number of infectious contacts per unit time, and π describes the proportion of those contacts that result in infection; the need for this distinction is explained below. Infected hosts recover from the pathogen at rate σV and achieve lifelong immunity. All hosts die at background mortality rate, b, or through the detrimental impact of the pathogen, which increases host mortality by rate αV. Finally, although helminths are typically thought to cause host morbidity, mortality can occur at high worm burdens. For that reason I make the simple assumption that helminths can increase host mortality at a burden-dependent (but low) per capita rate αW (the subscript ‘W’ is used throughout for ‘worms’, to denote parameters specific to the helminths) multiplied by the mean helminth burden M (thereby assuming, for simplicity, that the overall impact of the helminth on its host increases linearly with burden).
The expected host lifespan following pathogen infection under helminth coinfection
Given the above framework, it is possible to calculate the mean expected lifespan of the host following pathogen infection (Appendix 2):
| (1) |
where θ1=b+αv+αw and θ2=b+αwM. Clearly, a coinfecting helminth may alter the impact of the pathogen on its host, either directly by increasing host mortality, or indirectly by affecting the development of the pathogen or the host's ability to clear it. I therefore use equation (1) to explore how changes in mean helminth burden (M), and different forms of within-host interaction between the helminth and the pathogen species (via functional links between M and the pathogen-specific parameters in the model; see below), influence L.
Pathogen's basic reproduction number under helminth coinfection
Equation 1 describes how a coinfecting helminth may affect the impact of a pathogen at the individual host level. However, helminths are also likely to affect the spread of the pathogen at the population level. Given the epidemiological framework above, the basic reproduction number of the pathogen (R0,V) is:
| (2) |
(Fenton, 2008), which describes the initial potential for spread of the pathogen through a population of susceptible hosts. Using this equation I explore how mean worm burden (M) affects R0,V under the various interaction scenarios described below.
The evolution of pathogen virulence (αV) under helminth coinfection
The evolution of pathogen virulence has been, and continues to be, a major focus of evolutionary research. Typically the associated theoretical work assumes that virulence is an unavoidable consequence of the pathogen's exploitation of the host, necessary for the pathogen to fuel its replication and subsequent transmission (Levin and Pimentel, 1981; Anderson and May, 1982; Bremermann and Pickering, 1983; Ewald, 1983; Frank, 1996). Although these initial models have been greatly expanded upon (Antia et al. 1994; Bull, 1994; Lenski and May, 1994; Bonhoeffer et al. 1996; Ganusov et al. 2002; Restif and Koella, 2003; Boots et al. 2004; Fenton et al. 2006; Day et al. 2007; Kamo et al. 2007; Alizon, 2008a, 2008b; Alizon and van Baalen, 2008b; Frank and Schmid-Hempel, 2008; Mideo et al. 2008; Carval and Ferriere, 2010), they have so far ignored one ubiquitous component of virtually every pathogen's environment in natural populations: the presence of coinfecting helminths.
To model the evolution of pathogen virulence under helminth coinfection, I follow the standard (although not uncontroversial: Lipsitch and Moxon, 1997; Ebert and Bull, 2003; Alizon et al. 2009) assumption of a trade-off between increased pathogen infectivity (captured within the parameter βV) and the damage caused to the host (αV); that is βV increases with αV (e.g. Ewald, 1983; Frank, 1992; Bull, 1994; Lenski and May, 1994; May and Nowak, 1995; Bonhoeffer et al. 1996; Ebert and Herre, 1996; Day, 2003; Bolker et al. 2010). I follow this assumption, recognizing on-going debate regarding its applicability, to ensure a clear connection between the present work and the large body of theory that has been developed around this basic framework. As is well known for such models, if there is no relationship between βV and αV then evolution will act to maximize infectiousness (βV will increase indefinitely) and virulence (αV) will diminish to zero (Anderson and May, 1982). In addition, if there is a linear or accelerating relationship between βV and αV (such that large increases in βV can be achieved with relatively small increase in αV) then evolution will select for ever-increasing values of both. Similar results hold for all scenarios explored in this paper. I therefore focus attention on the more interesting case where there is a saturating relationship between βV and αV, such that progressive increases in transmission (βV) come at the cost of rapid increases in host mortality (αV). Specifically, I assume the functional relationship:
| (3) |
where βV,MAX is the maximal value of βV (at high αV) and k is the half-saturation constant, determining the rate of approach to the maximum (βV initially increases at rate βV,MAX/k as αV increases from small).
Invasion analysis (Appendix 4) shows that, under the simple assumption of no coinfection by multiple strains of the pathogen, evolution will act to maximize the pathogen's basic reproduction number (equation 2). Hence, in what follows I ignore potential complexities that may arise from coinfection by multiple pathogen strains, and consider the optimal level of pathogen virulence (αV*) as the value of αV that maximizes R0,V, found by inserting the assumed βV–αV trade-off relationship (equation 3) into equation (2), differentiating with respect to αV, setting equal to 0 and solving for αV (Appendix 4).
Incorporating within-host interactions between the pathogen and coinfecting helminths
Helminths may interact in a variety of ways with coinfecting pathogens. Here I modify the baseline framework described above to examine a range of possible mechanisms of interaction, either positive (synergistic) or negative (antagonistic), affecting different aspects of pathogen life-history. Due to a lack of quantified empirical information on how the strength of these various potential interactions vary with worm burden, in each case I assume the simplest possible relationships, typically linear where possible or constrained to prevent biologically impossible scenarios from occurring (e.g. probabilities exceeding 1, or rates becoming negative). As will be seen from the results, the quantitative outcome of each scenario can depend quite sensitively on the relationship between worm burden and the interaction strength; hence there is a clear need to obtain more precise, quantified measures of these relationships from empirical studies. Nevertheless, the simple scenarios presented here offer clear general insight into when and how such interactions may affect pathogen spread, impact and evolution.
Here I consider three scenarios:
-
(1)Helminths alter the ability of hosts to clear coinfecting pathogens (interaction via recovery, σV). Helminths frequently have the potential to immune-modulate their hosts, damping a wide range of specific and non-specific immune effectors (Maizels et al. 2004; van Riet et al. 2007). Furthermore, an established (albeit over-simplified) paradigm of coinfection immunology is that hosts may face a trade-off under coinfection, such that they are not able to mount maximal immune responses against both helminth and pathogen infections; helminths typically stimulate one arm of the host's immune response (the Th2 arm), which may inhibit the host from mounting an effective Th1 response against coinfecting viruses or bacteria (Abbas et al. 1996). Although highly simplified, both these concepts suggest that on-going infection by helminths may reduce the host's ability to clear pathogen infections (i.e. a positive, synergistic, effect of helminth infection on the pathogen). I modelled this scenario by assuming an inverse relationship between mean worm burden (M) and the host's recovery rate from the pathogen (σV):
where σMIN (⩾0) is the minimum recovery rate from pathogen infection and A and B are constants (units: worms time−1 and worms, respectively) that determine the rate of decline in recovery rate with increasing helminth burden (Fig. 2A).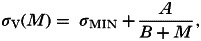
(4) Alternatively, helminths may increase the ability of hosts to clear coinfecting pathogens (e.g. Knowles et al. 2013), either via physical means (helminths interfere directly with pathogens or their sites of infection) or via the host's immune response (hosts are able to clear pathogen infections faster in the presence of helminths than in their absence). Hence, helminths may have a negative, antagonistic, effect on co-infecting pathogens via recovery. I modelled this scenario by assuming a positive relationship (assumed, for simplicity, to be linear) between mean worm burden and pathogen recovery rate:
where C (worm−1 time−1) determines the rate of increase in recovery rate with increasing worm burden (Fig. 2B).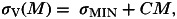
(5) -
(2)Helminths alter host susceptibility to pathogen infection (interaction via host susceptibility to the pathogen, π). As in the first part of scenario 1, hosts may struggle to mount effective immune responses against both helminths and pathogens but this may occur prior to pathogen infection, such that helminth infection reduces the ability of the host to prevent the pathogen from infecting in the first place (i.e. a positive effect of the helminth on the pathogen). I modelled this scenario by assuming a saturating relationship between worm burden and susceptibility to pathogen infection (π):
where πMIN (>0) is the minimum degree of host susceptibility, D determines the maximum degree of host susceptibility (such that the maximum πMIN+D⩽1) and E is a constant (units: worms) that, together with D, determines the rate at which susceptibility initially increases with worm burden (Fig. 2C).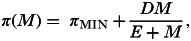
(6) In addition I explored the opposite scenario, whereby helminth infection decreases host susceptibility to pathogen infection (a negative effect on the pathogen via π). Here I assumed a functional relationship between mean worm burden and susceptibility to the pathogen similar to that shown in Fig. 2A:
where F and G are constants (units: worms).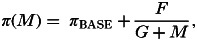
(7) -
(3)Helminths alter the detrimental effect to the host of the pathogen (interaction via pathogen virulence αV). So far I have assumed that worm and pathogen effects on host mortality (αW and αV) act additively. However, it is quite possible that the presence of helminths would either diminish or exacerbate the detrimental impact to the host of coinfecting pathogen infection (Griffiths et al. 2011). As an example of the former, the harmful effects of infection may arise from immunopathology (e.g. a major factor leading to severe cerebral malaria is the occurrence of a harmful inflammatory (Th1) response; Hartgers and Yazdanbakhsh, 2006). In this scenario the presence of helminths can reduce disease severity by stimulating counteractive Th2 or Treg responses. I model this positive interaction by assuming a decreasing relationship between worm burden (M) and pathogen virulence (αV):
where αV,BASE (units: time−1) determines the maximum pathogen virulence when there are no helminths present (M=0), and J (units: worms) influences the rate of reduction in virulence with increasing helminth burdens.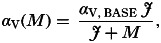
(8)
Alternatively, the presence of helminths may exacerbate pathogen damage, for example by reducing the host's somatic maintenance. I model this negative interaction by assuming an increasing (linear) relationship between helminth burden and pathogen virulence:
| (9) |
where αV,MIN (units: time−1) and K (units: worm−1) are constants. Note that, in both these cases, I assume these alterations of pathogen virulence arise through increased or decreased ability of the host to manage the damage caused by the presence of the pathogen, but do not alter the pathogen's replication or transmission rate. This distinction is especially important when considering how coinfection affects the evolution of pathogen virulence under these scenarios (Appendix 4).
Fig. 2.

Assumed interspecific interactions between helminth and pathogen, showing the relationship between mean worm burden (M) and (A) host recovery rate from the pathogen (σV) under a positive interaction, (B) host recovery rate from the pathogen under a negative interaction and (C) host susceptibility to pathogen infection (π).
In each of the above cases I inserted the relevant function (equations 4–9) into equations (1) and (2) and, for each scenario, explored the effect of varying the mean helminth burden (M) on the expected lifespan of the host (L), the pathogen's basic reproduction number (R0,V), and the evolution of pathogen virulence (αV*). Once again, it should be emphasized that these analyses are not intending to replicate any specific host-helminth–pathogen system, and so parameter values and the quantitative levels of predicted responses are purely arbitrary. However, by examining a range of interaction scenarios, this work reveals important context dependencies in the effects of helminth coinfection, and deworming, on coinfecting pathogen dynamics and host health.
RESULTS
Impact of within-host interactions on expected host lifespan following pathogen infection (L)
The different forms of within-host interaction between coinfecting helminths and pathogens have important qualitative and quantitative effects on the host's expected lifespan. Overall most forms of within-host interaction tend to result in host life expectancy following pathogen infection reducing with increasing helminth burdens (Fig. 3). Hence, reducing mean helminth burdens through deworming will tend to increase mean life expectancy under most scenarios considered. Clearly the magnitude of this relationship will depend, at least in part, on the damage caused by the helminths themselves; more benign helminths (low αW) will only cause a gradual reduction in host lifespan with increasing burdens. Furthermore, when coinfecting helminths either exacerbate the damage caused by the pathogen or reduce the ability of the host to clear the pathogen, the expected host lifespan can drop very rapidly with increasing worm burdens. However, when helminths either increase the host's ability to clear the pathogen or reduce the damage caused by the pathogen, low to intermediate worm burdens tend to be beneficial to the host, increasing life expectancy over that seen in helminth-free hosts (Fig. 3). It is only when worm burdens reach very high levels that host lifespan is reduced, as the helminth's own detrimental impact on the host at high burdens becomes detrimental to the host. Hence, in these cases, although deworming is likely to be beneficial to heavily infected individuals, it can result in a reduction in host survival in those with intermediate worm burdens under these scenarios.
Fig. 3.

Effect of mean worm burden (M) on the expected host lifespan following pathogen infection, relative to the baseline lifespan in the absence of helminth infection (Lrel), under the different interspecific interactions shown. Parameter values: H=20, βV=55 t−1, π=0·05, αV=8 t−1, b=0·1 t−1, αW=0·001 w−1t−1, σV=2 t−1, A=100 w t−1, B=50 w, C=0·05 w−1 t−1, D=0·1, E=200 w, F=100 w, G=2040 w, J=60 w, K=0·01 w−1, σMIN=2 t−1, αV,BASE=8 t−1, αV,MIN=8 t−1, π,MIN=0·05, π,BASE=0·001.
Impact of within-host interactions on the pathogen's basic reproduction number (R0,V)
Firstly it should be noted that, even in the absence of any explicit interaction between parasites, increasing helminth burden results in a reduction in R0,V, due to a decrease in duration of the pathogen's infectious period through helminth-induced host mortality (Fig. 4). Clearly, the less virulent the helminth is (low αW), the more gradual this reduction in R0,V will be. Hence, deworming treatments that reduce mean worm burdens will tend to increase the pathogen's rate of spread through the host population.
Fig. 4.

Effect of mean worm burden (M) on the pathogen's basic reproduction number (R0,V) under the different interspecific interactions shown. Parameter values are the same as used in Fig. 3.
The reduction in R0,V with increasing worm burden is exacerbated when there is an explicit negative (antagonistic) within-host interaction between helminths and pathogens, either via an increased ability of the host to clear the pathogen, reduced susceptibility of the hosts to pathogen infection, or increased host mortality due to the pathogen (Fig. 4, dashed lines). However these reductions in R0,V happen for different reasons, at different scales of organization. When helminths increase the host's ability to clear pathogen infection, or when helminths increase the rate of pathogen–host mortality, R0,V is reduced due to a decrease in the mean duration of pathogen infectiousness (a within-host effect). However, when helminths protect hosts from pathogen infection by reducing host susceptibility, R0,V is reduced due to a reduction in the availability of susceptible hosts (a between-host effect). Therefore, although similar relationships between helminth burdens and the pathogen's basic reproduction number are seen under all three antagonistic scenarios, they are happening for very different reasons.
When there is a positive (synergistic) interaction between helminths and coinfecting pathogens, under any of the mechanisms examined, the relationship between mean helminth burden and pathogen spread becomes more complex (Fig. 4, solid lines). In all cases, there is initially a net positive effect of coinfecting helminths on R0,V as mean helminth burdens increase from rare; low to intermediate worm burdens are beneficial to pathogens due to their effects on the host's ability to evade or remove or tolerate pathogen infection. In each case, however, there becomes a point at which the detrimental effect that the helminths themselves have on the host's survival, and hence the pathogen's infectious period, becomes sufficiently strong to override its direct synergistic effect on the pathogen, resulting in an overall negative effect of high worm burdens on R0,V. The point at which these curves turn over (i.e. the burden at which the net effect of helminths on R0,V changes from being positive to negative) is inversely related to the helminth's own virulence (αW; see Appendix 3). Hence, relatively benign helminths (low αW) are more likely to have a net beneficial effect on coinfecting pathogens, and will continue to do so at higher burdens, than more pathogenic helminths. Overall, under a positive interaction between helminth and pathogen, deworming could either increase or decrease the pathogen's R0, depending on the initial worm burden, the efficacy and coverage of treatment (the number or proportion of worms killed) and the pathogenicity of the helminth itself.
Impact of within-host interactions on the evolution of pathogen virulence (αV*)
In general, increasing helminth burdens tend to select for increased virulence in coinfecting pathogens, even in the absence of any explicit interaction between them (Appendix 4; Fig. 5). This is because the helminth's own detrimental impact to the host reduces the duration of infectiousness of the pathogen (the more worms, or the more pathogenic those worms are, the shorter the host's lifespan, and hence the shorter the duration of pathogen infectiousness), thereby reducing the pathogen's fitness. Effectively, since the duration of pathogen infectiousness is reduced by the presence of coinfecting helminths, the pathogen has little to lose by increasing its exploitation rate, and so it pays to increase its transmission rate (βV) at the expense of its virulence (αV). Similar results have previously been shown simply by increasing host's natural mortality rate (Anderson and May, 1982; Ebert and Bull, 2003; Alizon et al. 2009). Hence, in many ways, the impact of helminths in this scenario can simply be considered an environmental factor that increases the background rate of host mortality. However, as described below, if those helminths dynamically alter the pathogen's life-history via a within-host interspecific interaction, then alternative outcomes may be possible.
Fig. 5.

Effect of mean worm burden (M) on the pathogen's optimal virulence (αV*) under the different interspecific interactions shown. Parameter values are the same as used in Fig. 3, with the addition of: k=50 w, βV,MAX=400 t−1.
The different mechanisms of parasite interaction can alter the magnitude, and even direction, of pathogen virulence evolution. First, interactions that affect host susceptibility to pathogen infection (either positively or negatively) have no additional impact on pathogen evolution over that of helminths that do not interact with the pathogen (see Appendix 4); by only altering whether a pathogen infects, not how long it infects for, such interactions impose no additional selection on pathogen virulence. Second, helminth interactions that either increase the rate of host recovery from pathogen infection (a negative effect on the pathogen), or reduce the rate of host mortality due to the pathogen (a positive effect on the pathogen), can both strongly select for higher levels of pathogen virulence than in the absence of any interaction (Fig. 5), but for very different reasons. Under the former scenario, the increased host recovery rate means that the pathogen's duration of infectiousness is dramatically reduced, especially at high helminth burdens, selecting for faster host exploitation rates by the pathogen. Under the latter scenario, by reducing host mortality rate but allowing the pathogen to maintain a high transmission rate, the helminths effectively reduce the cost of high pathogen exploitation, allowing it to evolve high transmission (and hence virulence) levels. In all these cases deworming would have the added benefit of selecting for reduced virulence in the pathogen.
There are, however, two exceptions to this general pattern. First, when the helminth exacerbates pathogen-induced host mortality (a negative effect on the pathogen), this selects for reduced pathogen virulence with increasing helminth burdens (Fig. 5), as it now pays the pathogen to prolong host lifespan in order to maximize transmission potential. Deworming in this case would have the unfortunate side-effect of selecting for increased pathogen virulence. Second, when helminths reduce the host's ability to clear the pathogen (a positive effect on the pathogen), the direction of selection on pathogen virulence can vary depending on the level of helminth infection; low levels of infection can select for reduced pathogen virulence, whereas high levels of infection select for increased virulence (Fig. 5). When helminth infection levels are low, the interaction via recovery dominates, and a (small) increase in helminth burdens reduces the host's ability to clear the pathogen, prolonging its duration of infectiousness, leading to reduced host exploitation rates. However, at high helminth burdens, their detrimental impact on host mortality dominates, reducing the pathogen's infectious period, and selecting for increased virulence. In this case, deworming will either select for reduced or increased pathogen virulence, purely depending on the underlying mean worm burden.
DISCUSSION
There is considerable interest in the development of integrated control programmes that incorporate deworming components to improve the treatment of many important human pathogens (e.g. HIV, tuberculosis, malaria: Bentwich et al. 1999; Harms and Feldmeier, 2002; Druilhe et al. 2005; Harris et al. 2009). The present work shows that the outcome of such deworming programmes may be highly context-dependent, with counterintuitive, and potentially undesirable, consequences for the spread, persistence and severity of disease caused by a microparasitic pathogen. Recent papers (Taylor-Robinson et al. 2012; Bundy et al. 2013) have highlighted the variability between studies in the benefits of deworming, and inconsistency of evidence in support of it. The results presented here may help to shed some light on those results in the context of coinfection. In particular, the ecological and evolutionary impacts of deworming programmes would depend, possibly quite subtly, on the specific mechanism of interaction between helminths and other coinfecting pathogens, and on the precise pre- and post-treatment helminth burdens. Hence, without a quantitative understanding of how helminths affect the development, survival or susceptibility to coinfecting pathogens, it could be hard to interpret observed individual- or population-level responses to helminth infection or deworming strategies. However, these results are also encouraging, by showing that apparently highly variable or counterintuitive responses to infection or deworming may not simply be random noise, and hence totally unpredictable, but rather they arise from the balance of several interacting processes, such as the quantitative impact of coinfecting helminths on both the pathogen and the host, and the scaling relationships from within-host to population-level processes. Hence, by understanding the mechanism of within-host interaction and the quantitative levels of infection it may be possible to predict the likely impact of coinfection, and subsequent effects of deworming, on host health, pathogen spread and evolution.
At the individual level, high helminth burdens are likely to decrease host life expectancy (and, presumably, increase host morbidity, as is typical for most parasitic helminths). As such there would seem to be direct benefits of deworming, alleviating individual suffering and mortality (e.g. Bundy et al. 2013). However in terms of the indirect benefits, in the context of coinfection, deworming may prove detrimental to the individual. In particular if the individual has low to moderate levels of helminth infection, and those helminths either increase the host's ability to clear the pathogen or if the helminths reduce the pathogen's detrimental impact to the host (e.g. through immuno-modulatory effects that reduce immunopathology; Hartgers and Yazdanbakhsh, 2006), then removing those helminths will result in a reduction in host life expectancy (Fig. 3), or an otherwise detrimental effect on host health (e.g. Nacher, 2006). Clearly the quantitative benefits of deworming on host life expectancy are going to depend on the pathogenicity of the worms. Typically helminth infections are thought to primarily cause host morbidity (e.g. Little et al. 2004a), only causing significant mortality under very high infection levels (although increases in helminth burdens have been shown to cause increases in human mortality in some cases; Little et al. 2004b; Walker et al. 2012). Depending on the relationship between worm burden and excess host mortality, the results presented may be modified. For example, if helminths only affect mortality at very high burdens, a step function may be more appropriate, resulting in the net effect on host life expectancy resembling Fig. 3, but with a more pronounced benefit of helminth infection, and corresponding detriment of deworming, under the two interaction scenarios described above. Alternatively, if excess host mortality initially increases rapidly with helminth burden and then saturates (e.g. Little et al. 2004b; Walker et al. 2012) then the initial benefits of light helminth infections on host lifespan are less likely to occur.
At the population level, previous theory (Fenton, 2008; Jolles et al. 2008; Ezenwa and Jolles, 2011) has shown that the outcome of helminth coinfection on a pathogen's R0 can depend on the balance of various, potentially opposing, mechanisms of interaction. However, the present framework goes further than previous studies by explicitly incorporating the quantitative effect of varying helminth burden (M), rather than just a qualitative measure of being infected or not. In so doing, the present model shows that the magnitude and even qualitative direction of the net balance of opposing forces can vary depending on the mean helminth burden considered. If the helminth acts antagonistically to the pathogen then deworming will always benefit the pathogen, increasing its basic reproduction number (Fig. 4, dashed lines). However, if there is a synergistic effect of the helminth on the pathogen (Fig. 4, solid lines) the net effects of deworming may either be beneficial to the pathogen, increasing its basic reproduction number (R0,V), thereby reducing the community-wide benefits of control, or detrimental (decreasing R0,V), providing added community-wide value to treatment, depending purely on the quantitative level of helminth infection.
Importantly, comparing Figs 3 and 4 shows that the individual- and population-level benefits of deworming may conflict with each other, depending on the underlying mechanism of interaction. For example, if the helminth exacerbates the severity of disease caused by the pathogen (Figs 3 and 4, dashed blue lines) then deworming (reducing helminth burden) could dramatically increase host life expectancy (Fig. 3) but would also increase the pathogen's rate of spread throughout the host population (Fig. 4). Therefore, for certain within-host interactions between helminths and coinfecting pathogens, it may not be possible to have beneficial treatment effects at both the individual and population levels, such that reductions in population-level prevalence following a deworming programme may be counteracted by an increase in disease severity at the individual level.
Coinfection studies to date have typically focused on the ecological effects of interspecific parasite interactions. However, I also explore the evolutionary consequences of helminth–pathogen coinfection, showing that most deworming programmes would be predicted to select for reduced pathogen virulence under the scenarios considered here (Fig. 5). Even in the absence of an explicit interaction, the presence of coinfecting helminths may reduce host survival and hence the duration of the pathogen's mean infectious period, thereby selecting for increased virulence. However if the presence of the helminth exacerbates disease severity, or if helminths restrict the ability of the host to clear the pathogen, then deworming can select for increased pathogen virulence, particularly when pre-treatment helminth burdens are low (Fig. 5). Clearly, for such results to be relevant, any deworming effort would need to be maintained for sufficiently long for the pathogen to respond evolutionarily. Typically many (although by no means all) deworming programmes have been relatively short-lived, and follow-up studies typically show that worm burdens rapidly bounce back to pre-treatment levels once treatment has ceased (Bundy et al. 1985; Njongmeta et al. 2004). In such cases, we may expect a rapid return to selection for pre-treatment levels of pathogen virulence.
This evolutionary analysis builds on a large body of theory exploring the evolution of pathogen virulence in response to intraspecific coinfection (Frank, 1992, 1996; Bull, 1994; Levin and Bull, 1994; Nowak and May, 1994; May and Nowak, 1995; van Baalen and Sabelis, 1995; Mosquera and Adler, 1998; Read and Taylor, 2001; Alizon, 2008b; Alizon and van Baalen, 2008a). In those models a distinction is often made between coinfections involving related strains, which tend to select for reduced virulence through kin selection (Frank, 1992, 1996; Chao et al. 2000; Schjorring and Koella, 2003; Lively, 2005), and those involving unrelated strains which can select for increased virulence (e.g. due to competition between strains via cross-immunity, or replacement by more virulent strains, in the case of super-infection) (Bonhoeffer and Nowak, 1994; May and Nowak, 1994; van Baalen and Sabelis, 1995; Frank, 1996; Mosquera and Adler, 1998). Under interspecific coinfection, as explored here, the species involved may come from very different taxa (e.g. viruses or bacteria coinfecting with parasitic helminths), and so the analysis presented here connects closest with the previous theory on unrelated intraspecific strains. It is shown here that negative interactions between coinfecting helminths and pathogens, similar to those between competing strains in the intraspecific coinfection literature, can select for increased virulence. However, the present work extends that theory considerably by, firstly, considering the effect of quantitative levels of infection (i.e. varying helminth burdens, rather than simply the presence or absence of coinfection) and, secondly, by exploring a wider range of possible mechanisms of interaction than those typically assumed in the intraspecific coinfection literature. Hence, existing models of intraspecific coinfection are not sufficient to capture the full range of within-host interactions seen in natural multi-parasite disease systems.
Overall this paper shows there is considerable context dependence in the ecological, clinical and evolutionary impact of helminths on coinfecting pathogens. From an empirical point of view, this suggests that studies that sample from different areas or different time points of the same system, with the same underlying mechanism of interspecific interaction, may see very different effects of coinfection, purely depending on the infection burdens in each sample. Hence, studies that attempt to infer the direction of an interspecific parasite interaction purely from observed patterns of association between coinfecting parasites may result in conflicting, or erroneous, results. Furthermore, the work presented here shows that variability between studies in the effects of deworming treatments can be explained and understood through a quantitative understanding of the precise mechanisms of interspecific parasite interactions (i.e. not just whether they are antagonistic or synergistic, but whether they act on host susceptibility, pathogen replication, immune-mediated clearance etc.), ideally across a realistic range of helminth infection loads. Clearly to achieve this for a specific disease system is not a trivial task, and the present simple and highly generalized model would need to be explicitly tailored to capture the relevant aspects of that system. However, by incorporating such quantitative empirical measurements within such a theoretical framework, we may be able to better understand the occurrence and impacts of interspecific parasite interactions, and develop effective integrated control programmes appropriately.
ACKNOWLEDGEMENTS
The author would like to thank Nicole Mideo, Amy Pedersen, Sarah Knowles and Sarah Reece for invaluable discussions and comments throughout the development of this work.
Appendix 1: The theoretical framework for helminth-pathogen coinfection
The baseline epidemiological framework is described by the equations:
where the total host population, H=S+I+R.
Appendix 2: Calculating the expected host lifespan following pathogen infection
The expected lifespan of a host following pathogen infection, L, is given as:
 |
Given the framework outlined in Appendix 1, this is:
Where θ1=b+αv+αwM and θ2=b+αwM. From this, if the host never recovers from pathogen infection (σV=0), the mean host life expectancy following pathogen infection is 1/θ1. If the host recovers immediately from infection (σV→∞), the mean host life expectancy is 1/θ2.
Appendix 3: Analyses of the mean helminth burden at which the net effect of the helminth on the pathogen's basic reproduction number (R0,V) reverses, under different direct positive interaction scenarios
As shown in Fig. 4 there is a tendency for a humped relationship between mean helminth burden and R0,V under positive (synergistic) interspecific interactions. The helminth burden at which the relationship turns over in each case (M′) can be found by differentiating the appropriate equation for R0,V with respect to M, setting equal to zero and solving for M. Here I present those analyses for each of the three positive interaction scenarios shown in Fig. 4.
1. Positive interaction via reduced recovery from pathogen infection
Here the equation for R0,V is:
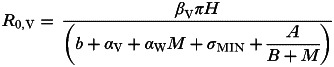 |
giving 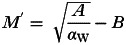 .
.
2. Positive interaction via increased susceptibility to pathogen infection
Here the equation for R0,V is:
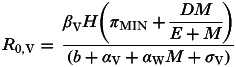 |
giving
3. Positive interaction via reduced damage to the host
Here the equation for R0,V is:
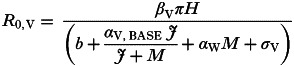 |
giving
In all cases the mean helminth burden at which the relationship with R0,V turns over is inversely related to the per capita helminth virulence (αW).
Appendix 4: Analyses of evolution of pathogen virulence under different interspecific interactions with coinfecting helminths
Invasion analysis
To explore the evolution of pathogen virulence under coinfection I use an invasion analysis approach. Here it is assumed there are two pathogen ‘strains’ (denoted by the subscripts 1 and 2 in what follows), which differ in their virulence and any relevant parameters via assumed functional links to virulence. For simplicity we assume these two pathogen strains cannot coexist within the same host individual, with the first strain to infect being assumed to prevent infection of that host by the other strain. By extension of the single-strain framework presented in Appendix 1, this leads to the following system of equations for the dynamics of the 2-strain system:
where 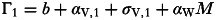 and
and 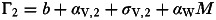 and
H=S+I1+I2+R.
This system has three steady states (where variables are listed in the order (S,
I1, I2, R), and
equilibria are denoted by ‘*’):
and
H=S+I1+I2+R.
This system has three steady states (where variables are listed in the order (S,
I1, I2, R), and
equilibria are denoted by ‘*’):
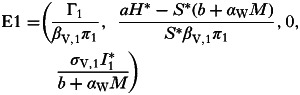 |
and
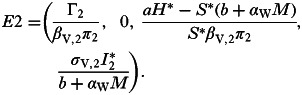 |
From this it is clear that the two pathogen strains cannot coexist; either strain 1 or strain 2 will win out and exclude the other. To determine the conditions under which either of these outcomes happens we calculate the Jacobian of the system:
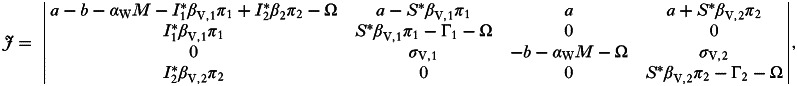 |
where the Ω's are the eigenvalues of the system. If we assume we are at stable state E1, where strain 1 (the ‘resident’) is at endemic equilibrium with the host, and strain 2 (the invading mutant) is initially vanishingly rare (I2 ≈ 0), the Jacobian simplifies to:
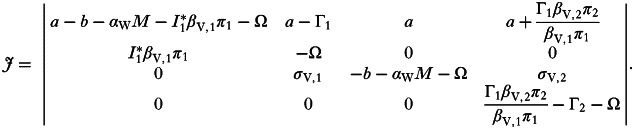 |
This matrix has four eigenvalues, the first three of which correspond to those of the 3×3 sub-matrix in the top left corner of J, representing the single-strain (I1) system; as stated above, we assume the resident strain 1 is at endemic equilibrium with the host and so the stability criteria for this sub-matrix are fulfilled (Ωi<0, i∈{1, 2, 3}). Therefore stability of J is determined solely by the positivity of the eigenvalue in the bottom right entry of J:
If this eigenvalue is negative then state E1 is stable and strain 2 cannot invade. However, if it is positive then strain 2 invades and replaces strain 1; this happens when:
In other words, the winner is the strain with the highest basic reproduction number. Hence, the optimal level of virulence is the one that maximizes R0,V, leading to the analyses presented in the main text.
Expressions for the optimal level of virulence under each interaction scenario
1. Baseline model (no explicit interaction between helminths and pathogen)
Under the assumption of a saturating trade-off between virulence and transmission (equation 3, main text), the basic reproduction number for the pathogen in the absence of any explicit interaction with the helminth is:
| (A1) |
To find the optimal virulence (the value of αV that maximizes the above expression), we differentiate equation (A1) with respect to αV, set equal to zero and solve for αV. In the absence of any explicit interaction between the helminth and pathogen, this results in:
showing, first, that evolution tends to select for intermediate levels of pathogen virulence (0⩽αV*⩽∞) and second that, in the absence of any interaction between helminths and pathogen, the presence of the helminth tends to select for increased pathogen virulence due to the reduced host life expectancy shortening the pathogen's infectious period at high worm burdens. The rate at which this happens depends on the magnitude of the helminth's per capita virulence; benign helminths (low αW) would only impact on pathogen virulence at very high burdens.
2. Positive effect of helminths on pathogen, via reduced host recovery from infection
Here we insert the relationship between mean worm burden (M) and host recovery (σV) from equation (4) in the main text into equation (A1) and solve to obtain the following expression for the optimal pathogen virulence under this positive interaction via recovery:
3. Negative effect of helminths on pathogen, via increased host recovery from infection
Inserting the relationship between mean worm burden (M) and host recovery (σV) from equation (5) into equation (A1) provides the following expression for the optimal pathogen virulence under this negative interaction via recovery:
which has the same functional form, in terms of the effect of helminth burden, as the relationship in the absence of any explicit interaction between helminths and the pathogen, although the absolute optimal virulence is increased in this case due to the parameter C, which determines the strength of the interspecific interaction.
4. Positive effect of helminths on pathogen, via increased host susceptibility to infection
Inserting the relationship from equation (6) into equation (A1) provides the following expression for the optimal pathogen virulence under this positive interaction via host susceptibility:
which is the same as the expression in the absence of any explicit interaction between the helminth and pathogen. Hence, although this form of interaction affects the pathogen's basic reproduction number, it does not affect selection on pathogen virulence.
5. Negative effect of helminths on pathogen, via reduced host susceptibility to infection
Inserting the relationship from equation (7) into equation (A1) provides the following expression for αV*:
which, again, is the same as the expression in the absence of any interaction between helminth and pathogen.
6. Positive effect of helminths on pathogen, via reduced host damage caused by pathogen
Inserting the relationship from equation (8) into equation (A1) provides the following expression for the optimal pathogen virulence under this positive interaction via host damage:
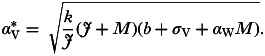 |
7. Negative effect of helminths on pathogen, via increased host damage caused by pathogen
Inserting the relationship from equation (9) into equation (A1) provides the following expression for the optimal pathogen virulence under this positive interaction via host damage:
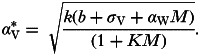 |
FINANCIAL SUPPORT
This work was funded by grants from the Natural Environment Research Council (NERC; NE/G007349 and NE/G006830).
REFERENCES
- Abbas A. K., Murphy K. M. and Sher A. (1996). Functional diversity of helper T lymphocytes. Nature 383, 787–793 [DOI] [PubMed] [Google Scholar]
- Alizon S. (2008a). Transmission-recovery trade-offs to study parasite evolution. American Naturalist 172, E113–E121. 10.1086/589892 [DOI] [PubMed] [Google Scholar]
- Alizon S. (2008b). Decreased overall virulence in coinfected hosts leads to the persistence of virulent parasites. American Naturalist 172, E67–E79. 10.1086/588077 [DOI] [PubMed] [Google Scholar]
- Alizon S. and van Baalen M. (2008a). Multiple infections, immune dynamics, and the evolution of virulence. American Naturalist 172, E150–E168. 10.1086/590958 [DOI] [PubMed] [Google Scholar]
- Alizon S. and van Baalen M. (2008b). Transmission-virulence trade-offs in vector-borne diseases. Theoretical Population Biology 74, 6–15. 10.1016/j.tpb.2008.04.003 [DOI] [PubMed] [Google Scholar]
- Alizon S., Hurford A., Mideo N. and Van Baalen M. (2009). Virulence evolution and the trade-off hypothesis: history, current state of affairs and the future. Journal of Evolutionary Biology 22, 245–259. 10.1111/j.1420-9101.2008.01658.x [DOI] [PubMed] [Google Scholar]
- Anderson R. M. and May R. M. (1982). Coevolution of hosts and parasites. Parasitology 85, 411–426 [DOI] [PubMed] [Google Scholar]
- Antia R., Levin B. R. and May R. M. (1994). Within-host population dynamics and the evolution and maintenance of microparasite virulence. American Naturalist 144, 457–472 [Google Scholar]
- Basáñez M.-G., French M. D., Walker M. and Churcher T. S. (2012). Paradigm lost: how parasite control may alter pattern and process in human helminthiases. Trends in Parasitology 28, 161–171. 10.1016/j.pt.2012.02.004 [DOI] [PubMed] [Google Scholar]
- Behnke J. M., Gilbert F. S., Abu-Madi M. A. and Lewis J. W. (2005). Do the helminth parasites of wood mice interact? Journal of Animal Ecology 74, 982–993 [Google Scholar]
- Bentwich Z., Kalinkovich A., Weisman Z., Borkow G., Beyers N. and Beyers A. D. (1999). Can eradication of helminthic infections change the face of AIDS and tuberculosis? Immunology Today 20, 485–487 [DOI] [PubMed] [Google Scholar]
- Bolker B. M., Nanda A. and Shah D. (2010). Transient virulence of emerging pathogens. Journal of the Royal Society Interface 7, 811–822. 10.1098/rsif.2009.0384 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonhoeffer S. and Nowak M. A. (1994). Mutation and the evolution of virulence. Proceedings of the Royal Society of London Series B 258, 133–140 [Google Scholar]
- Bonhoeffer S., Lenski R. E. and Ebert D. (1996). The curse of the pharaoh: the evolution of virulence in pathogens with long living propagules. Proceedings of the Royal Society of London Series B 263, 715–721 [DOI] [PubMed] [Google Scholar]
- Boots M., Hudson P. J. and Sasaki A. (2004). Large shifts in pathogen virulence relate to host population structure. Science 303, 842–844 [DOI] [PubMed] [Google Scholar]
- Bremermann H. J. and Pickering J. (1983). A game-theoretical model of parasite virulence. Journal of Theoretical Biology 100, 411–426 [DOI] [PubMed] [Google Scholar]
- Bull J. J. (1994). Perspective – Virulence. Evolution 48, 1423–1437 [DOI] [PubMed] [Google Scholar]
- Bundy D. A. P., Thompson D. E., Cooper E. S., Golden M. H. N. and Anderson R. M. (1985). Population dynamics and chemotherapeutic control of Trichuris trichuria infection of children in Jamaica and St Lucia. Transactions of the Royal Society of Tropical Medicine and Hygiene 79, 759–764. 10.1016/0035-9203(85)90110-5 [DOI] [PubMed] [Google Scholar]
- Bundy D. A. P., Walson J. L. and Watkins K. L. (2013). Worms, wisdom, and wealth: why deworming can make economic sense. Trends in Parasitology 29, 142–148 [DOI] [PubMed] [Google Scholar]
- Carval D. and Ferriere R. (2010). A unified model for the coevolution of resistance, tolerance and virulence. Evolution 64, 2988–3009. 10.1111/j.1558-5646.2010.01035.x [DOI] [PubMed] [Google Scholar]
- Chao L., Hanley K. A., Burch C. L., Dahlberg C. and Turner P. E. (2000). Kin selection and parasite evolution: higher and lower virulence with hard and soft selection. Quarterly Review of Biology 75, 261–275. 10.1086/393499 [DOI] [PubMed] [Google Scholar]
- Cox F. E. G. (2001). Concomitant infections, parasites and immune responses. Parasitology 122, S23–S38 [DOI] [PubMed] [Google Scholar]
- Day T. (2003). Virulence evolution and the timing of disease life-history events. Trends in Ecology and Evolution 18, 113–118. 10.1016/s0169-5347(02)00049-6 [DOI] [Google Scholar]
- Day T., Graham A. L. and Read A. F. (2007). Evolution of parasite virulence when host responses cause disease. Proceedings of the Royal Society B–Biological Sciences 274, 2685–2692. 10.1098/rspb.2007.0809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Druilhe P., Tall A. and Sokhna C. (2005). Worms can worsen malaria: towards a new means to roll back malaria? Trends in Parasitology 21, 359–362 [DOI] [PubMed] [Google Scholar]
- Ebert D. and Bull J. J. (2003). Challenging the trade-off model for the evolution of virulence: is virulence management feasible? Trends in Microbiology 11, 15–20. 10.1016/s0966-842x(02)00003-3 [DOI] [PubMed] [Google Scholar]
- Ebert D. and Herre E. A. (1996). The evolution of parasitic diseases. Parasitology Today 12, 96–101. 10.1016/0169-4758(96)80668-5 [DOI] [PubMed] [Google Scholar]
- Ewald P. W. (1983). Host–parasite relations, vectors, and the evolution of disease severity. Annual Review of Ecology and Systematics 14, 465–485. 10.1146/annurev.es.14.110183.002341 [DOI] [Google Scholar]
- Ezenwa V. O. and Jolles A. E. (2011). From host immunity to pathogen invasion: the effects of helminth coinfection on the dynamics of microparasites. Integrative and Comparative Biology 51, 540–551. 10.1093/icb/icr058 [DOI] [PubMed] [Google Scholar]
- Ezenwa V. O., Etienne R. S., Luikart G., Beja-Pereira A. and Jolles A. E. (2010). Hidden consequences of living in a wormy world: nematode-induced immune suppression facilitates tuberculosis invasion in African buffalo. American Naturalist 176, 613–624. 10.1086/656496 [DOI] [PubMed] [Google Scholar]
- Fenton A. (2008). Worms and germs: the population dynamic consequences of microparasite–macroparasite co-infection. Parasitology 135, 1545–1560 [DOI] [PubMed] [Google Scholar]
- Fenton A. and Perkins S. E. (2010). Applying predator-prey theory to modelling immune-mediated, within-host interspecific parasite interactions. Parasitology 137, 1027–1038. 10.1017/s0031182009991788 [DOI] [PubMed] [Google Scholar]
- Fenton A., Lello J. and Bonsall M. B. (2006). Pathogen responses to host immunity: the impact of time delays and memory on the evolution of virulence. Proceedings of the Royal Society B–Biological Sciences 273, 2083–2090 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fenton A., Lamb T. and Graham A. L. (2008). Optimality analysis of Th1/Th2 immune responses during microparasite–macroparasite co-infection, with epidemiological feedbacks. Parasitology 135, 841–853 [DOI] [PubMed] [Google Scholar]
- Fenton A., Viney M. E. and Lello J. (2010). Detecting interspecific macroparasite interactions from ecological data: patterns and process. Ecology Letters 13, 606–615. 10.1111/j.1461-0248.2010.01458.x [DOI] [PubMed] [Google Scholar]
- Frank S. A. (1992). A kin selection model for the evolution of virulence. Proceedings of the Royal Society of London Series B 250, 195–197 [DOI] [PubMed] [Google Scholar]
- Frank S. A. (1996). Models of parasite virulence. Quarterly Review of Biology 71, 37–78 [DOI] [PubMed] [Google Scholar]
- Frank S. A. and Schmid-Hempel P. (2008). Mechanisms of pathogenesis and the evolution of parasite virulence. Journal of Evolutionary Biology 21, 396–404. 10.1111/j.1420-9101.2007.01480.x [DOI] [PubMed] [Google Scholar]
- Ganusov V. V., Bergstrom C. T. and Antia R. (2002). Within-host population dynamics and the evolution of microparasites in a heterogeneous host population. Evolution 56, 213–223 [DOI] [PubMed] [Google Scholar]
- Graham A. L. (2008). Ecological rules governing helminth–microparasite coinfection. Proceedings of the National Academy of Sciences USA 105, 566–570 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham A. L., Cattadori I. M., Lloyd-Smith J. O., Ferrari M. J. and Bjornstad O. N. (2007). Transmission consequences of coinfection: cytokines writ large? Trends in Parasitology 23, 284–291 [DOI] [PubMed] [Google Scholar]
- Griffiths E. C., Pedersen A. B., Fenton A. and Petchey O. L. (2011). The nature and consequences of coinfection in humans. Journal of Infection 63, 200–206. 10.1016/j.jinf.2011.06.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harms G. and Feldmeier H. (2002). HIV infection and tropical parasitic diseases – deleterious interactions in both directions? Tropical Medicine and International Health 7, 479–488 [DOI] [PubMed] [Google Scholar]
- Harris J. B., Podolsky M. J., Bhuiyan T. R., Chowdhury F., Khan A. I., LaRocque R. C., Logvinenko T., Kendall J., Faruque A. S. G., Nagler C. R., Ryan E. T., Qadri F. and Calderwood S. B. (2009). Immunologic responses to Vibrio cholerae in patients co-infected with intestinal parasites in Bangladesh. PLoS Neglected Tropical Diseases 3, e403. 10.1371/journal.pntd.0000403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartgers F. C. and Yazdanbakhsh M. (2006). Co-infection of helminths and malaria: modulation of the immune responses to malaria. Parasite Immunology 28, 497–506. 10.1111/j.1365-3024.2006.00901.x [DOI] [PubMed] [Google Scholar]
- Hawkes N. (2012). Deworming debunked. Br Med J 345, e8558. 10.1136/bmj.e8558 [DOI] [PubMed] [Google Scholar]
- Jolles A. E., Ezenwa V. O., Etienne R. S., Turner W. C. and Olff H. (2008). Interactions between macroparasites and microparasites drive infection patterns in free-ranging African buffalo. Ecology 89, 2239–2250. 10.1890/07-0995.1 [DOI] [PubMed] [Google Scholar]
- Kamo M., Sasaki A. and Boots M. (2007). The role of trade-off shapes in the evolution of parasites in spatial host populations: an approximate analytical approach. Journal of Theoretical Biology 244, 588–596. 10.1016/j.jtbi.2006.08.013 [DOI] [PubMed] [Google Scholar]
- Knowles S. C. L., Fenton A., Petchey O. L., Jones R. C., Barber R. and Pedersen A. B. (2013). Stability of within-host parasite communities in a wild mammal system. Proceedings of the Royal Society of London Series B (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lello J., Boag B., Fenton A., Stevenson I. R. and Hudson P. J. (2004). Competition and mutualism among the gut helminths of a mammalian host. Nature 428, 840–844 [DOI] [PubMed] [Google Scholar]
- Lenski R. E. and May R. M. (1994). The evolution of virulence in parasites and pathogens: reconciliation between two competing hypotheses. Journal of Theoretical Biology 169, 253–265. 10.1006/jtbi.1994.1146 [DOI] [PubMed] [Google Scholar]
- Levin B. R. and Bull J. J. (1994). Short-sighted evolution and the virulence of pathogenic microorganisms. Trends in Microbiology 2, 76–81. 10.1016/0966-842x(94)90538-x [DOI] [PubMed] [Google Scholar]
- Levin S. and Pimentel D. (1981). Selection of intermediate rates of increase in parasite–host systems. American Naturalist 117, 308–315. 10.1086/283708 [DOI] [Google Scholar]
- Lipsitch M. and Moxon E. R. (1997). Virulence and transmissibility of pathogens: what is the relationship? Trends in Microbiology 5, 31–37. 10.1016/s0966-842x(97)81772-6 [DOI] [PubMed] [Google Scholar]
- Little M. P., Basanez M. G., Breitling L. P., Boatin B. A. and Alley E. S. (2004a). Incidence of blindness during the Onchocerciasis Control Programme in western Africa, 1971–2002. Journal of Infectious Diseases 189, 1932–1941. 10.1086/383326 [DOI] [PubMed] [Google Scholar]
- Little M. P., Breitling L. P., Basanez M. G., Alley E. S. and Boatin B. A. (2004b). Association between microfilarial load and excess mortality in onchocerciasis: an epidemiological study. Lancet 363, 1514–1521. 10.1016/s0140-6736(04)16151-5 [DOI] [PubMed] [Google Scholar]
- Lively C. M. (2005). Evolution of virulence: coinfection and propagule production in spore-producing parasites. BMC Evolutionary Biology 5, 64. 10.1186/1471-2148-5-64 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maizels R. M., Balic A., Gomez-Escobar N., Nair M., Taylor M. D. and Allen J. E. (2004). Helminth parasites – masters of regulation. Immunological Reviews 201, 89–116. 10.1111/j.0105-2896.2004.00191.x [DOI] [PubMed] [Google Scholar]
- May R. M. and Nowak M. A. (1994). Superinfection, metapopulation dynamics, and the evolution of diversity. Journal of Theoretical Biology 170, 95–114 [DOI] [PubMed] [Google Scholar]
- May R. M. and Nowak M. A. (1995). Coinfection and the evolution of parasite virulence. Proceedings of the Royal Society of London Series B 261, 209–215 [DOI] [PubMed] [Google Scholar]
- Mideo N., Alizon S. and Day T. (2008). Linking within- and between-host dynamics in the evolutionary epidemiology of infectious diseases. Trends in Ecology and Evolution 23, 511–517. 10.1016/j.tree.2008.05.009 [DOI] [PubMed] [Google Scholar]
- Mosquera J. and Adler F. R. (1998). Evolution of virulence: a unified framework for coinfection and superinfection. Journal of Theoretical Biology 195, 293–313 [DOI] [PubMed] [Google Scholar]
- Nacher M. (2006). Worms and malaria: resisting the temptation to generalize. Trends in Parasitology 22, 350–351. 10.1016/j.pt.2006.06.003 [DOI] [PubMed] [Google Scholar]
- Njongmeta L. M., Nfon C. K., Gilbert J., Makepeace B. L., Tanya V. N. and Trees A. J. (2004). Cattle protected from onchocerciasis by ivermectin are highly susceptible to infection after drug withdrawal. International Journal for Parasitology 34, 1069–1074. 10.1016/j.ijpara.2004.04.011 [DOI] [PubMed] [Google Scholar]
- Nowak M. A. and May R. M. (1994). Superinfection and the evolution of parasite virulence. Proceedings of the Royal Society of London Series B 255, 81–89 [DOI] [PubMed] [Google Scholar]
- Pedersen A. B. and Fenton A. (2007). Emphasising the ecology in parasite community ecology. Trends in Ecology and Evolution 22, 133–139 [DOI] [PubMed] [Google Scholar]
- Petney T. N. and Andrews R. H. (1998). Multiparasite communities in animals and humans: frequency, structure and pathogenic significance. International Journal for Parasitology 28, 377–393 [DOI] [PubMed] [Google Scholar]
- Read A. F. and Taylor L. H. (2001). The ecology of genetically diverse infections. Science 292, 1099–1102. 10.1126/science.1059410 [DOI] [PubMed] [Google Scholar]
- Restif O. and Koella J. C. (2003). Shared control of epidemiological traits in a coevolutionary model of host–parasite interactions. American Naturalist 161, 827–836 [DOI] [PubMed] [Google Scholar]
- Schjorring S. and Koella J. C. (2003). Sub-lethal effects of pathogens can lead to the evolution of lower virulence in multiple infections. Proceedings of the Royal Society B–Biological Sciences 270, 189–193. 10.1098/rspb.2002.2233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor-Robinson D. C., Maayan N., Soares-Weiser K., Donegan S. and Garner P. (2012). Deworming drugs for soil-transmitted intestinal worms in children: effects on nutritional indicators, haemoglobin and school performance. Cochrane Database of Systematic Reviews 7, CD000371. [DOI] [PubMed] [Google Scholar]
- Telfer S., Lambin X., Birtles R., Beldomenico P., Burthe S., Paterson S. and Begon M. (2010). Species interactions in a parasite community drive infection risk in a wildlife population. Science 330, 243–246 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Baalen M. and Sabelis M. W. (1995). The dynamics of multiple infection and the evolution of virulence. American Naturalist 146, 881–910 [Google Scholar]
- van Riet E., Hartgers F. C. and Yazdanbakhsh M. (2007). Chronic helminth infections induce immunomodulation: consequences and mechanisms. Immunobiology 212, 475–490. 10.1016/j.imbio.2007.03.009 [DOI] [PubMed] [Google Scholar]
- Walker M., Little M. P., Wagner K. S., Soumbey-Alley E. W., Boatin B. A. and Basanez M.-G. (2012). Density-dependent mortality of the human host in onchocerciasis: relationships between microfilarial load and excess mortality. PLoS Neglected Tropical Diseases 6. 10.1371/journal.pntd.0001578 [DOI] [PMC free article] [PubMed] [Google Scholar]