

Abstract
Leishmaniasis is a disease that affects millions of people worldwide. The drugs that are available for the treatment of this infection exhibit high toxicity and various side effects. Several studies have focused on the development of new chemotherapeutic agents that are less toxic and more effective against trypanosomatids. We investigated the effects of N-butyl-1-(4-dimethylamino)phenyl-1,2,3,4-tetrahydro-β-carboline-3-carboxamide (C4) and its possible targets against L. amazonensis. The results showed morphological and ultrastructural alterations, depolarization of the mitochondrial membrane, the loss of cell membrane integrity, and an increase in the formation of mitochondrial superoxide anions in L. amazonensis treated with C4. Our results indicate that C4 is a selective antileishmanial agent, and its effects appear to be mediated by mitochondrial dysfunction.
1. Introduction
Leishmaniasis is a disease caused by the protozoa of the genus Leishmania, which belongs to the order Kinetoplastida, family Trypanosomatidae. An estimated 12 million people are affected by this disease [1], with 2 million new cases worldwide [2] reported in 88 countries and four continents [3]. This disease has two clinical forms, visceral and cutaneous leishmaniasis.
The currently available first-line treatments are pentavalent antimonials [4]. These drugs have high toxicity and adverse side effects [5]. Amphotericin B and pentamidine are second-line therapies but are associated with long-term treatment, limited effectiveness, significant side effects, and toxicity [6–8]. Consequently, an urgent need exists to discover new drugs that are effective against leishmaniasis.
Several studies are being conducted to find new antileishmanial compounds. Carbolines comprise a class of compounds that have an alkaloid indole nucleus and hydrogenated six-member pyridine ring [9]. They are distributed throughout nature in many living beings, including vegetables, fungi, animals, and even human fluids [10]. Carbolines can be divided into three groups: fully aromatic, dihydrocarbolinic, and tetrahydro-β-carboline [11]. Interestingly, natural and synthetic β-carbolines are well known to possess several biological properties. For example, our group recently reported the effective trypanocidal activity of the synthetic compound N-butyl-1-(4-dimethylamino)phenyl-1,2,3,4-tetrahydro-β-carboline-3-carboxamide (C4; Figure 1) against Trypanosoma cruzi [12, 13].
Figure 1.
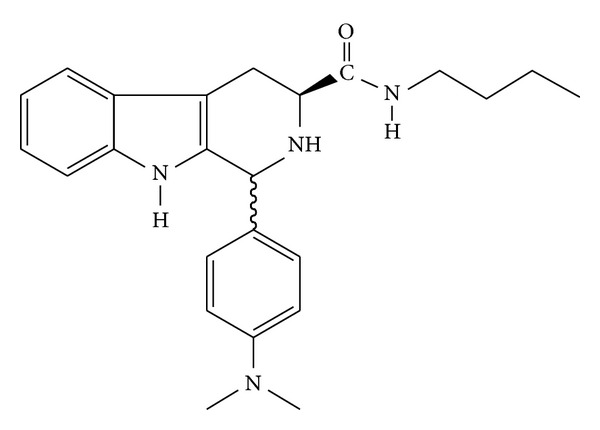
Chemical structure of N-butyl-1-(4-dimethylamino)phenyl-1,2,3,4-tetrahydro-β-carboline-3-carboxamide (C4).
The mechanism of action of β-carboline compounds involves changes in DNA and inhibition of the respiratory chain in epimastigote forms of T. cruzi [10]. Although the mode of biological action of this compound in different cell types is not fully understood, previous studies have shown that the β-carboline ring in the molecule might be deposited in base pairs of DNA, thus contributing to its biological activity [14, 15] or inducing apoptosis [16].
Considering the low efficacy of drugs against leishmaniasis and previous studies of the effects of β-carboline compounds on T. cruzi, we investigated the potential effect of the synthetic compound N-butyl-1-(4-dimethylamino)phenyl-1,2,3,4-tetrahydro-β-carboline-3-carboxamide (C4) against L. amazonensis and its possible targets in this protozoan.
2. Materials and Methods
2.1. Chemicals
Antimycin A, carbonyl cyanide m-chlorophenylhydrazone (CCCP), digitonin, dimethylsulfoxide (DMSO), and rhodamine 123 (Rh123) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Fetal bovine serum (FBS) and RPMI-1640 were obtained from Invitrogen (Grand Island, NY, USA). 3,8-Phenanthridinediamine-5-(6-triphenylphosphoniumhexyl)-5,6-dihydro-6-phenyl (MitoSOX) and propidium iodide (PI) were obtained from Invitrogen (Eugene, OR, USA). All of the other reagents were of analytical grade.
2.2. Synthesis of C4 Compound
C4 was synthesized as previously described by Valdez et al. [12].
2.3. Preparation of Drugs
The C4 compound was prepared in DMSO. All of the groups, including controls, were tested at final concentrations of less than 1% DMSO, a concentration that was found to not affect parasite or mammalian cells (data not shown).
2.4. Parasites and Cell Culture
L. amazonensis (strain WHOM/BR/75/JOSEFA) was originally isolated from a patient with diffuse cutaneous leishmaniasis by C.A. Cuba-Cuba (Universidade de Brasília, Brazil). Promastigotes were cultured in Warren's medium (brain heart infusion, hemin, and folic acid; pH 7.0) supplemented with 10% inactivated FBS at 25°C. Axenic amastigotes were cultured in Schneider's medium (Sigma, St. Louis, MO, USA; pH 4.6) supplemented with 20% FBS at 32°C.
Macrophages (J774G8) were maintained in tissue flasks with RPMI-1640 (pH 7.2), added with sodium bicarbonate and L-glutamine (As annex), and supplemented with 10% FBS at 37°C in a 5% CO2 atmosphere.
2.5. Antiproliferative Assay
Promastigote and axenic amastigote forms in the logarithmic phase (1 × 106 cells/mL) were cultured on a 24-well plate in Warren's and Schneider's media, respectively, supplemented with FBS in the presence or absence of 2.56, 12.8, 25.6, 128.0, and 256.0 μM of C4. The activity against promastigotes and axenic amastigotes was evaluated after 72 h of incubation. The results are expressed as a percentage, and the IC50 (i.e., 50% inhibitory concentration) was determined after-incubation [17].
2.6. Cytotoxicity Assay in Macrophage Cells
Cytotoxicity was evaluated in J774G8 macrophage cells. A suspension of 5 × 105 cells/mL was cultured in RPMI-1640 medium supplemented with 10% FBS and added to each well in 96-well microplates. The plates were incubated at 37°C in a 5% CO2-air mixture to obtain the confluent growth of the cells. After 24 h, the compound was added to each well at increasing concentrations (160.0, 320.0, 640.0, and 1,280.0 μM), and the plates were incubated for 48 h in a 5% CO2-air mixture at 37°C. After treatment, the medium was removed and washed with phosphate-buffered saline (PBS), and 50 μL of MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide formazan; 2 mg/mL) was added to each well for 4 h in a 5% CO2-air mixture at 37°C. DMSO (50 μL) was then added, and the plates were homogenized. Absorbance was read in a 96-well plate reader (BIO-TEK Power Wave XS spectrophotometer) at 492 nm. The percentage of viable cells was calculated relative to controls, consisting of cells cultured in medium alone, according to CC50 values (i.e., 50% cytotoxicity concentration). The CC50 was determined by logarithm regression analysis.
2.7. Scanning Electron Microscopy
The promastigote forms of the parasite in the logarithmic phase (1 × 106 cells/mL) in the presence or absence of 16.0 and 103.0 μM of C4 for 48 h were washed with 0.01 M PBS and fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer for 2 h at room temperature. The parasites were adhered on poly-l-lysine-coated coverslips, dehydrated in different concentrations of ethanol, critical-point dried with CO2, sputter coated with gold, and observed in a Shimadzu SS-550 scanning electron microscope [18].
2.8. Transmission Electron Microscopy
The promastigote forms of the parasite in the logarithmic-phase (1 × 106 cells/mL) in the presence or absence of 16.0 and 103.0 μM of C4 for 48 h were harvested by centrifugation and fixed in 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer. The cells were then post-fixed in a solution that contained 1% osmium tetroxide and 0.8% potassium ferrocyanide at room temperature for 60 min, dehydrated in different concentrations of acetone, and embedded in Epon resin. Thin sections were stained with uranyl acetate and lead citrate and examined in a JM 1400 JEOL transmission electron microscope [19].
2.9. Mitochondrial Membrane Potential and Cell Membrane Integrity Assay
The promastigote forms of the parasite in the logarithmic phase (5 × 106 cells/mL) in the presence or absence of 16.0 and 103.0 μM of C4 for 24 h were harvested and washed with PBS. The parasites were then washed and incubated at 37°C with Rh 123 (5 μg/mL for 15 min) to evaluate mitochondrial membrane potential (ΔΨm) and PI (0.2 μg/mL for 10 min) to verify possible alterations in cell membrane integrity. CCCP (100 μM) and digitonin (40 μM) were used as positive controls for mitochondria membrane potential alterations and cell membrane alterations, respectively. The material was kept on ice until analysis. The mean fluorescence intensity of the cells was analyzed by flow cytometry using FACSCalibur and CellQuest software. A total of 10,000 events were acquired in the region that was previously established as the one that corresponded to the parasites [20].
2.10. Fluorimetric Detection of Mitochondrial-Derived O2 ∙−
Promastigote forms of the parasite in the logarithmic phase (2 × 107 cells/mL) were harvested and washed with Krebs-Henseleit (KH) solution buffer that contained 15 mM NaHCO3, 5 mM KCl, 120 mM NaCl, 0.7 mM Na2HPO4, and 1.5 mM NaH2PO4 (pH 7.3). The cells were loaded with 5 μM MitoSOX reagent. The parasites were incubated for 10 min at room temperature (25°C) and protected from light. After incubation with MitoSOX reagent, the parasites were washed twice with KH buffer and treated or not with 16.0, 103.0, and 205.0 μM of C4. Antimycin A (10 μM), a stimulus that is known to induce superoxide anion (O2 ∙−) production by mitochondria, was used as a positive control. MitoSOX detection was performed using black 96-well plates for 3 h. Fluorescence was measured using a fluorescence microplate reader (Victor X3; PerkinElmer) at 194 λex = 510 nm and λem = 580 nm [21].
2.11. Statistical Analysis
The data shown in the graphs are expressed as the mean ± standard error of at least three independent experiments. The data were analyzed using analysis of variance (ANOVA). Significant differences among means were identified using Tukey post hoc test. Values of P ≤ 0.05 were considered statistically significant. The statistical analyses were performed using Statistica software.
3. Results
3.1. Antileishmanial Activity
The treatment of the parasites with C4 dose dependently inhibited the growth of the promastigote and axenic amastigote forms of L. amazonensis. The inhibition percentages of the parasites and concentrations that corresponded to 50% (IC50) and 90% (IC90) of growth inhibition of the promastigotes were calculated by plotting the concentration versus percentage growth inhibition using linear regression after directly counting free-living parasites in a Neubauer chamber. The IC50 and IC90 in promastigotes were 16.0 ± 2.28 μM and 103.0 ± 20.17 μM, respectively. In axenic amastigotes, the IC50 and IC90 were 16.3 ± 2.38 μM and 118.0 ± 45 μM, respectively (Table 1).
Table 1.
Effect of C4 against promastigote and axenic amastigote forms of Leishmania amazonensis, cytotoxicity in macrophages cells, and selectivity index.
| Cells | C4 (µM) IC50 a | C4 (µM) IC90 b | SIc |
|---|---|---|---|
| Promastigotes | 16.0 ± 2.28 | 103.0 ± 20.17 | 45.1 |
| Axenic amastigotes | 16.3 ± 2.38 | 118.0 ± 45 | 44.2 |
| Macrophages (CC50)d | 722.0 ± 45 | — | — |
aConcentration that inhibited 50% of growth.
bConcentration that inhibited 90% of growth.
cSelectivity index (SI; CC50 macrophages/IC50).
d50% cytotoxicity concentration in macrophages.
3.2. Cytotoxicity Assay
The cytotoxic effect (50% cytotoxic concentration [CC50]) of C4 in J774G8 macrophages after 48 h of treatment was 722.0 ± 45 μM (Table 1). The toxicity in macrophages was compared with activity against the promastigote and axenic amastigote forms, yielding the selectivity index (SI). C4 was more selective against the promastigote and axenic amastigote forms than against mammalian cells, with SIs of 45.1 and 44.2, respectively (Table 1).
3.3. Scanning and Transmission Electron Microscopy
Morphological alterations in promastigotes treated with C4 were observed using scanning electron microscopy. The control showed normal characteristics of the parasite, such as an elongated body and free flagellum (Figure 2(a)). Parasites treated with the IC50 of C4 exhibited distortions in the cell body (Figures 2(b) and 2(c)), and the group treated with the IC90 showed rounding of the cell body (Figure 2(d)).
Figure 2.
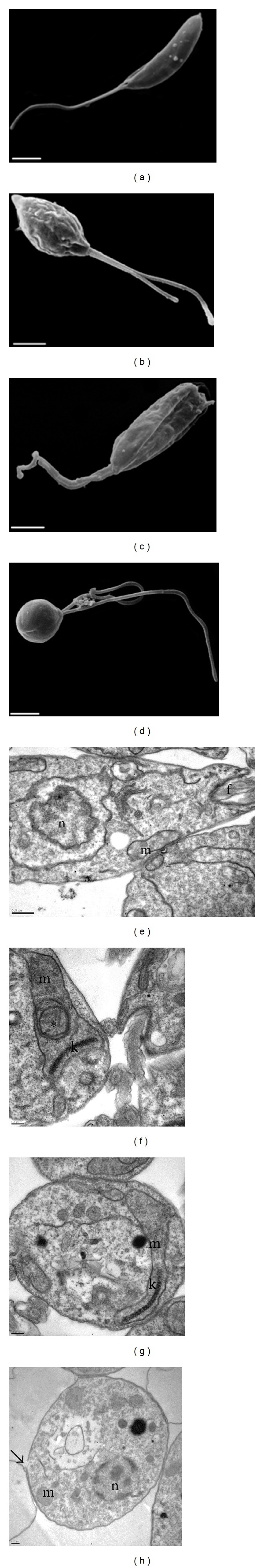
Scanning electron microscopy (SEM) and transmission electron microscopy (TEM) of Leishmania amazonensis promastigotes after 48 h of treatment with C4. (a) SEM image of an untreated promastigote, showing typical elongated morphology. (b) and (c) SEM images of promastigotes after treatment with the IC50 (16.0 μM), showing distortion of the cell body. (d) SEM image of promastigote after treatment with the IC90 (103.0 μM), showing rounding of the cell body. Scale bar: 2 μm. (e) TEM image of untreated promastigote, showing normal mitochondria (m), nucleus (n), and flagellum (f). (f) and (g) TEM images of promastigotes treated with C4 at 16.0 μM, showing mitochondrial swelling and the presence of concentric membranes inside mitochondria (black asterisk). (h) Promastigotes treated with C4 at 103.0 μM, showing alteration of the plasma membrane (black arrow). Scale bar: 0.5 μm in (e) and 0.2 μm in (f)–(h).
Ultrastructural changes in promastigotes treated with C4 at the IC50 and IC90 are illustrated in Figure 2, showing significant alterations. The mitochondria showed intense swelling (Figures 2(f) and 2(g)), the presence of concentric membrane structures inside the organelle (Figure 2(f)), and alterations of the plasma membrane (Figure 2(h)). These ultrastructural changes were not observed in untreated parasites (Figure 2(e)).
3.4. Mitochondrial Membrane Potential
The effect of C4 on mitochondrial membrane potential (ΔΨm) in promastigote forms was assessed by flow cytometry using Rh 123, a fluorescent marker that indicates mitochondrial membrane potential. The IC50 and IC90 induced 73.3% and 82.5% decreases in total Rh 123 fluorescence intensity, respectively, compared with the control group, indicating depolarization of mitochondrial membrane potential (Figure 3(b)). Promastigotes treated with CCCP showed a 94.8% decrease in membrane potential (Figure 3(a)).
Figure 3.
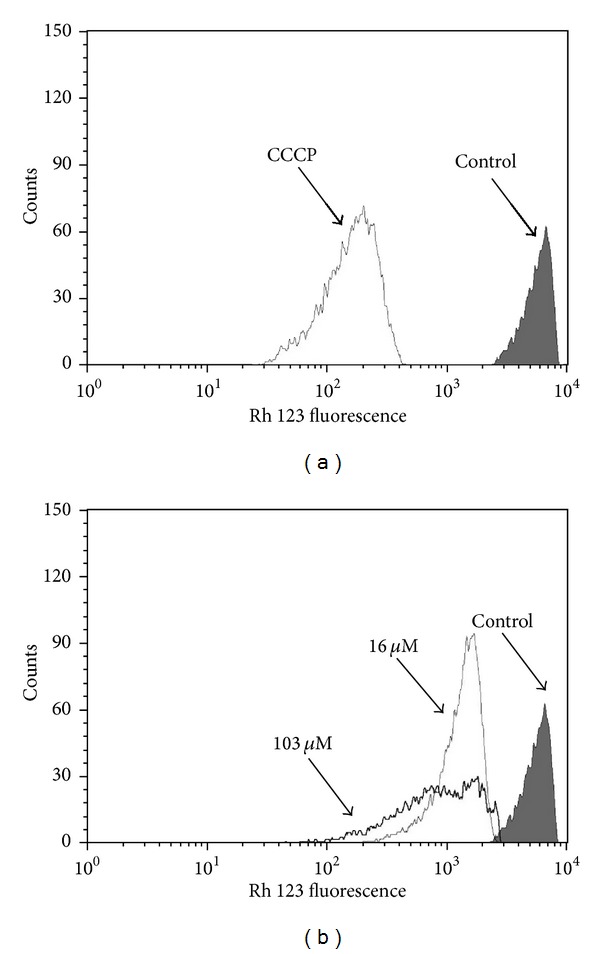
Flow cytometry analysis of promastigotes of Leishmania amazonensis treated with C4 for 24 h and stained with Rh 123. (a) Promastigotes treated with 100 μM CCCP (positive control). (b) Promastigotes treated with 16.0 and 103.0 μM of C4. The control group (i.e., untreated parasites) is also shown.
3.5. Plasma Membrane Integrity
Cell membrane integrity in promastigotes treated with C4 was assessed by flow cytometry using PI, which diffuses across disrupted plasma membranes of cells and binds to nucleic acids. C4 at the IC90 increased total PI fluorescence intensity by 22.2% (Figure 4(c)), compared with the control group (5.7%; Figure 4(a)), indicating the alteration of cell membrane integrity. However, C4 at the IC50 did not induce any membrane alterations (Figure 4(b)). The positive control (i.e., digitonin) also showed an increase in fluorescence (data not shown).
Figure 4.
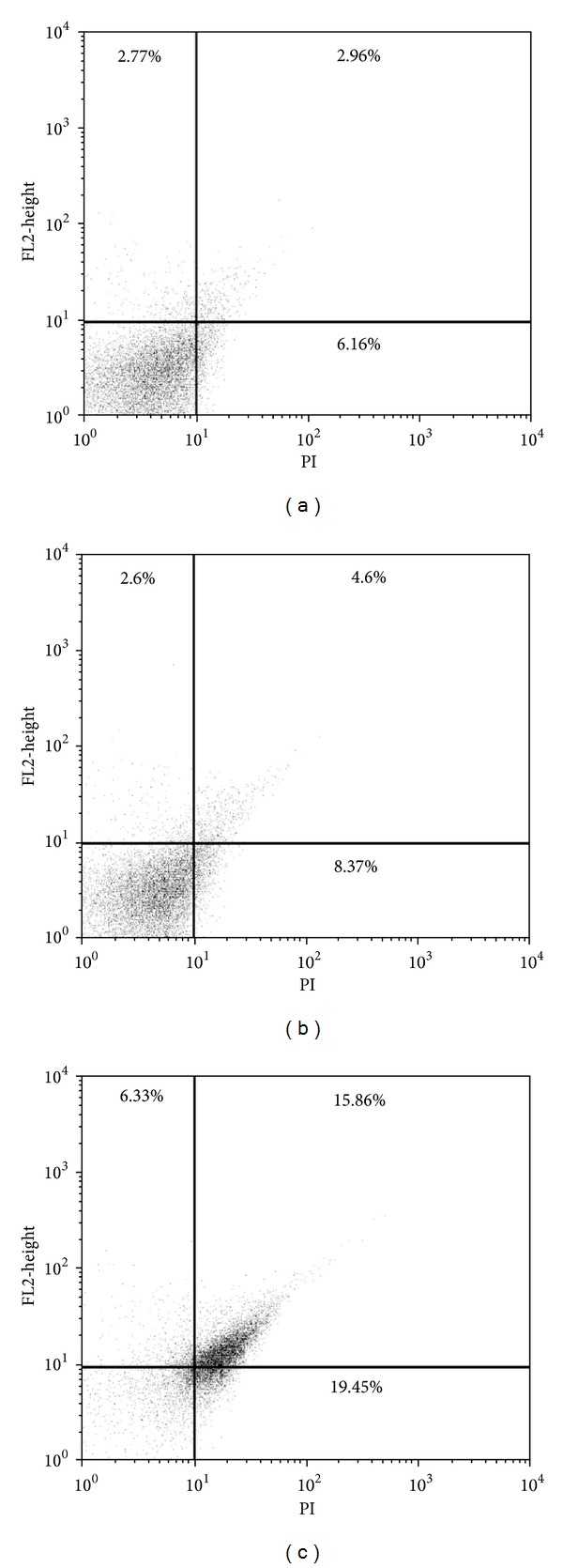
Flow cytometry analysis of promastigotes of Leishmania amazonensis treated with C4 for 24 h and stained with propidium iodide (PI). (a) Control group (i.e., untreated cells). (b) Promastigotes treated with 16.0 μM of C4. (c) Promastigotes treated with 103.0 μM of C4. The percentages of PI-stained positive cells are shown in the upper right and left quadrants.
3.6. Detection of Mitochondria-Derived O2 ∙−
The production of O2 ∙− was evaluated in promastigotes treated with C4 using MitoSOX reagent, which measures the accumulation of mitochondrial superoxide. Figure 5 shows that C4 increased the production of mitochondrial O2 ∙− at all concentrations and times tested compared with the control group. However, C4 induced a significant increase in O2 ∙− production only after 2 and 3 h of treatment at the higher concentration (205 μM). The positive control (i.e., antimycin A) also showed an increase in fluorescence (data not shown).
Figure 5.
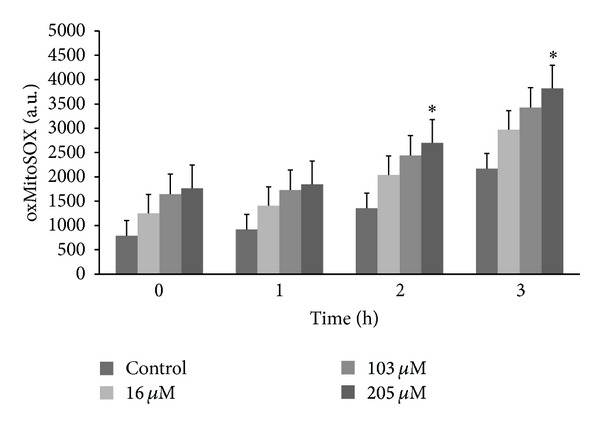
Mitochondrial O2 ∙− production in promastigote forms of Leishmania amazonensis treated with C4 for up to 3 h. Mitochondrial O2 ∙− production was evaluated using the fluorescent probe MitoSOX. At the indicated times, promastigotes were used to measure oxidized MitoSOX (oxMitoSOX). The results are expressed in arbitrary units (mean ± SE of at least three independent experiments). *P ≤ 0.05, significant difference compared with the control group (i.e., untreated cells; two-way analysis of variance followed by Tukey post hoc test).
4. Discussion
Several natural and synthetic compounds have been studied for the treatment of leishmaniasis [22–29]. Nevertheless, the treatment of this infection remains a problem because of the high toxicity and adverse side effects. β-Carbolines have been previously reported to have various biological properties, including antioxidant, antimicrobial [30], antiparasitic [12, 13], antitumoral [31], and antiviral effects [32]. These compounds also exhibited activity against protozoa, such as T. cruzi, especially against bloodstream trypomastigotes. Our previous studies demonstrated the effective and selective action of C4 against T. cruzi [12, 13]. The present study evaluated the antileishmanial activity of C4 against L. amazonensis and its possible targets in this parasite.
C4 compound inhibited the growth of the promastigote forms of the parasite and caused morphological and ultrastructural alterations, especially in mitochondria. The Rh 123 assay confirmed the mitochondrial action of C4, reflected by a decrease in Rh 123 fluorescence intensity. The mitochondria of trypanosomatids are attractive chemotherapeutic targets because they have structural and functional characteristics that are distinct from mammalian mitochondria [33]. In fact, increasing reports have described compounds that target parasite mitochondrial destabilization and disorganization [24, 26].
Mitochondrial alterations may be a consequence of many potentially harmful effects induced by both exogenous and endogenous toxic compounds. We currently consider C4 as an exogenous toxic compound. However, C4 may also induce the production of endogenous toxic compounds, including reactive oxygen species (ROS), that may be responsible for mitochondrial dysfunction and induce oxidative damage in lipids and proteins, the main macromolecules of biological membranes. This indirect effect of C4 was demonstrated in the MitoSOX and PI assays in the present study, similar to the effects of compound reported by Desoti et al. [34].
In conclusion, our data indicate that C4 is a leishmanicidal compound that is able to induce parasite disorders that are mainly mediated by mitochondrial dysfunction. Further in vitro and in vivo studies are necessary to increase our understanding of the mode of action of this compound and determine whether it can be exploited alone or in combination with other drugs for the treatment of antileishmaniasis.
Conflict of Interests
There is no conflict of interests declared by the authors.
Acknowledgments
The authors thank the staff of the microbiology laboratory for their collaboration. This study was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Capacitação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Financiadora de Estudos e Projetos (FINEP), Programa de Pós-Graduação em Ciências Biológicas da Universidade Estadual de Maringá, and Complexo de Centrais de Apoio a Pesquisa (COMCAP-UEM).
References
- 1.Desjeux P. Leishmaniasis: current situation and new perspectives. Comparative Immunology, Microbiology and Infectious Diseases. 2004;27(5):305–318. doi: 10.1016/j.cimid.2004.03.004. [DOI] [PubMed] [Google Scholar]
- 2.Alvar J, Yactayo S, Bern C. Leishmaniasis and poverty. Trends in Parasitology. 2006;22(12):552–557. doi: 10.1016/j.pt.2006.09.004. [DOI] [PubMed] [Google Scholar]
- 3.World Health Organization. First WHO Report on Neglected Tropical Diseases: Working to Overcome the Global Impact of Neglected Tropical Diseases. Geneva, Switzerland: 2010. http://whqlibdoc.who.int/publications/2010/9789241564090_eng.pdf. [Google Scholar]
- 4.Croft SL, Coombs GH. Leishmaniasis—current chemotherapy and recent advances in the search for novel drugs. Trends in Parasitology. 2003;19(11):502–508. doi: 10.1016/j.pt.2003.09.008. [DOI] [PubMed] [Google Scholar]
- 5.Croft SL, Seifert K, Yardley V. Current scenario of drug development for leishmaniasis. Indian Journal of Medical Research. 2006;123(3):399–410. [PubMed] [Google Scholar]
- 6.Guerin PJ, Olliaro P, Sundar S, et al. Visceral leishmaniasis: current status of control, diagnosis, and treatment, and a proposed research and development agenda. The Lancet Infectious Diseases. 2002;2(8):494–501. doi: 10.1016/s1473-3099(02)00347-x. [DOI] [PubMed] [Google Scholar]
- 7.Rodrigues JCF, Urbina JA, de Souza W. Antiproliferative and ultrastructural effects of BPQ-OH, a specific inhibitor of squalene synthase, on Leishmania amazonensis . Experimental Parasitology. 2005;111(4):230–238. doi: 10.1016/j.exppara.2005.08.006. [DOI] [PubMed] [Google Scholar]
- 8.Sereno D, Cordeiro da Silva A, Mathieu-Daude F, Ouaissi A. Advances and perspectives in Leishmania cell based drug-screening procedures. Parasitology International. 2007;56(1):3–7. doi: 10.1016/j.parint.2006.09.001. [DOI] [PubMed] [Google Scholar]
- 9.Henriques AT, Kerber VA, Moreno PRH. Alcaloides: generalidades e aspectos básicos. In: Simões CMO, Ros Petrovick P, editors. Farmacognosia—Da Planta Ao Medicamento. Porto Alegre/Florianópolis, Brazil: Universidade/UFRS. Ed. UFSC; 1999. [Google Scholar]
- 10.Rivas P, Cassels BK, Morello A, Repetto Y. Effects of some β-carboline alkaloids on intact Trypanosoma cruzi epimastigotes. Comparative Biochemistry and Physiology—C Pharmacology Toxicology and Endocrinology. 1999;122(1):27–31. doi: 10.1016/s0742-8413(98)10069-5. [DOI] [PubMed] [Google Scholar]
- 11.Glennon RA, Dukat M, Grella B, et al. Binding of β-carbolines and related agents at serotonin (5-HT2 and 5-HT1A), dopamine (D2) and benzodiazepine receptors. Drug and Alcohol Dependence. 2000;60(2):121–132. doi: 10.1016/s0376-8716(99)00148-9. [DOI] [PubMed] [Google Scholar]
- 12.Valdez RH, Tonin LTD, Ueda-Nakamura T, et al. Biological activity of 1,2,3,4-tetrahydro-β-carboline-3-carboxamides against Trypanosoma cruzi . Acta Tropica. 2009;110(1):7–14. doi: 10.1016/j.actatropica.2008.11.008. [DOI] [PubMed] [Google Scholar]
- 13.Valdez RH, Tonin LTD, Ueda-Nakamura T, et al. In vitro and in vivo trypanocidal synergistic activity of N-butyl-1-(4-dimethylamino)phenyl-1,2,3,4-tetrahydro-β-carboline-3-carboxamide associated with benznidazole. Antimicrobial Agents and Chemotherapy. 2012;56(1):507–512. doi: 10.1128/AAC.05575-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Boursereau Y, Coldham I. Synthesis and biological studies of 1-amino β-carbolines. Bioorganic and Medicinal Chemistry Letters. 2004;14(23):5841–5844. doi: 10.1016/j.bmcl.2004.09.036. [DOI] [PubMed] [Google Scholar]
- 15.Zhao M, Bi L, Wang W, et al. Synthesis and cytotoxic activities of β-carboline amino acid ester conjugates. Bioorganic and Medicinal Chemistry. 2006;14(20):6998–7010. doi: 10.1016/j.bmc.2006.06.021. [DOI] [PubMed] [Google Scholar]
- 16.Hans G, Malgrange B, Lallemend F, et al. β-carbolines induce apoptosis in cultured cerebellar granule neurons via the mitochondrial pathway. Neuropharmacology. 2005;48(1):105–117. doi: 10.1016/j.neuropharm.2004.09.001. [DOI] [PubMed] [Google Scholar]
- 17.Vendrametto MC, Santos AOD, Nakamura CV, Filho BPD, Cortez DAG, Ueda-Nakamura T. Evaluation of antileishmanial activity of eupomatenoid-5, a compound isolated from leaves of Piper regnellii var. pallescens . Parasitology International. 2010;59(2):154–158. doi: 10.1016/j.parint.2009.12.009. [DOI] [PubMed] [Google Scholar]
- 18.Pelizzaro-Rocha KJ, Veiga-Santos P, Lazarin-Bidóia D, et al. Trypanocidal action of eupomatenoid-5 is related to mitochondrion dysfunction and oxidative damage in Trypanosoma cruzi . Microbes and Infection. 2011;13(12-13):1018–1024. doi: 10.1016/j.micinf.2011.05.011. [DOI] [PubMed] [Google Scholar]
- 19.dos Santos AO, Britta EA, Bianco EM, et al. 4-acetoxydolastane diterpene from the Brazilian brown alga Canistrocarpus cervicornis as antileishmanial agent. Marine Drugs. 2011;9(11):2369–2383. doi: 10.3390/md9112369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Menna-Barreto RFS, Henriques-Pons A, Pinto AV, Morgado-Diaz JA, Soares MJ, de Castro SL. Effect of a β-lapachone-derived naphthoimidazole on Trypanosoma cruzi: identification of target organelles. Journal of Antimicrobial Chemotherapy. 2005;56(6):1034–1041. doi: 10.1093/jac/dki403. [DOI] [PubMed] [Google Scholar]
- 21.Piacenza L, Irigoín F, Alvarez MN, et al. Mitochondrial superoxide radicals mediate programmed cell death in Trypanosoma cruzi: cytoprotective action of mitochondrial iron superoxide dismutase overexpression. Biochemical Journal. 2007;403(2):323–334. doi: 10.1042/BJ20061281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Das R, Roy A, Dutta N, Majumder HK. Reactive oxygen species and imbalance of calcium homeostasis contributes to curcumin induced programmed cell death in Leishmania donovani . Apoptosis. 2008;13(7):867–882. doi: 10.1007/s10495-008-0224-7. [DOI] [PubMed] [Google Scholar]
- 23.Inacio JD, Canto-Cavalheiro MM, Menna-Barreto RF, et al. Mitochondrial damage contribute to epigallocatechin-3-gallate induced death in Leishmania amazonensis . Experimental Parasitology. 2012;132(2):151–155. doi: 10.1016/j.exppara.2012.06.008. [DOI] [PubMed] [Google Scholar]
- 24.Lopes MV, Desoti VC, Caleare AO, et al. Mitochondria superoxide anion production contributes to geranylgeraniol induced death in Leishmania amazonensis . Evidence-Based Complementary and Alternative Medicine. 2012;2012:9 pages. doi: 10.1155/2012/298320.298320 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.dos Santos AO, Veiga-Santos P, Ueda-Nakamura T, et al. Effect of elatol, isolated from red seaweed Laurencia dendroidea, on Leishmania amazonensis . Marine Drugs. 2010;8(11):2733–2743. doi: 10.3390/md8112733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.dos Santos AO, Ueda-Nakamura T, Dias Filho BP, et al. Copaiba oil: an alternative to development of new drugs against leishmaniasis. Evidence-Based Complementary and Alternative Medicine. 2012;2012:7 pages. doi: 10.1155/2012/898419.898419 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shukla AK, Patra S, Dubey VK. Iridoid glucosides from Nyctanthes arbortristis result in increased reactive oxygen species and cellular redox homeostasis imbalance in Leishmania parasite. European Journal of Medicinal Chemistry. 2012;54:49–58. doi: 10.1016/j.ejmech.2012.04.034. [DOI] [PubMed] [Google Scholar]
- 28.Paris C, Loiseau PM, Bories C, Bréard J. Miltefosine induces apoptosis-like death in Leishmania donovani promastigotes. Antimicrobial Agents and Chemotherapy. 2004;48(3):852–859. doi: 10.1128/AAC.48.3.852-859.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Britta EA, Silva APB, Ueda-Nakamura T, et al. Benzaldehyde thiosemicarbazone derived from limonene complexed with copper induced mitochondrial dysfunction in Leishmania amazonensis . Public Library of Science One. 2012;10:1631–1646. doi: 10.1371/journal.pone.0041440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cao R, Chen H, Peng W, et al. Design, synthesis and in vitro and in vivo antitumor activities of novel β-carboline derivatives. European Journal of Medicinal Chemistry. 2005;40(10):991–1001. doi: 10.1016/j.ejmech.2005.04.008. [DOI] [PubMed] [Google Scholar]
- 31.Formagio ASN, Tonin LTD, Foglio MA, et al. Synthesis and antitumoral activity of novel 3-(2-substituted-1,3,4-oxadiazol-5-yl) and 3-(5-substituted-1,2,4-triazol-3-yl) β-carboline derivatives. Bioorganic & Medicinal Chemistry. 2008;16(22):9660–9667. doi: 10.1016/j.bmc.2008.10.008. [DOI] [PubMed] [Google Scholar]
- 32.Formagio ASN, Santos PR, Zanoli K, et al. Synthesis and antiviral activity of β-carboline derivatives bearing a substituted carbohydrazide at C-3 against poliovirus and herpes simplex virus (HSV-1) European Journal of Medicinal Chemistry. 2009;44(11):4695–4701. doi: 10.1016/j.ejmech.2009.07.005. [DOI] [PubMed] [Google Scholar]
- 33.Menna-Barreto RFS, Goncalves RLS, Costa EM, et al. The effects on Trypanosoma cruzi of novel synthetic naphthoquinones are mediated by mitochondrial dysfunction. Free Radical Biology and Medicine. 2009;47(5):644–653. doi: 10.1016/j.freeradbiomed.2009.06.004. [DOI] [PubMed] [Google Scholar]
- 34.Desoti VC, Lazarin-Bidóia D, Sudatti DB, et al. Trypanocidal action of (−)-elatol involves an oxidative stress triggered by mitochondria dysfunction. Marine Drugs. 2012;10:1631–1646. doi: 10.3390/md10081631. [DOI] [PMC free article] [PubMed] [Google Scholar]