

Abstract
Mutants created by deleting the ddrA, ddrB, ddrC, ddrD, and pprA loci of Deinococcus radiodurans R1alone and in all possible combinations of pairs revealed that the encoded gene products contribute to this species’ resistance to UV light and/or mitomycin C. Deleting pprA from an otherwise wild type cell sensitizes the resulting strain to UV irradiation, reducing viability by as much as eight fold relative to R1. If this deletion is introduced into a ΔddrA or ΔddrD background, the resulting strains become profoundly sensitive to the lethal effects of UV light. At a fluence of 1000 Jm-2, the ΔddrA ΔpprA and ΔddrD ΔpprA strains are 100- and 1000-fold more sensitive to UV relative to the strain that has only lost pprA. Deletion of ddrA results in a 100 fold increase in strain sensitivity to mitomycin C, but in backgrounds that combine a deletion of ddrA with deletions of either ddrC or ddrD, mitomycin resistance is restored to wild type levels. Inactivation of ddrB also increases D. radiodurans sensitivity to mitomycin, but unlike the ddrA mutant deleting ddrC or ddrD from a ΔddrB background further increases that sensitivity. Despite the effect that loss of these gene products has on DNA damage resistance, none appear to directly affect either excision repair or homologous recombination suggesting that they participate in novel processes that facilitate tolerance to UV light and interstrand crosslinks in this species.
Introduction
Deinococcus radiodurans R1 is the type species for the Deinococcaceae, a family of bacteria [1,2] exhibiting extraordinary resistant to the lethal effects of many DNA damaging agents, including ionizing radiation (IR), ultraviolet (UV) light, and interstrand cross-linking agents (ICLs) [3]. For a vegetative cell, D. radiodurans R1 is particularly resistant to UV radiation, surviving doses as high as 750 Jm-2 with minimal loss of viability [4]. In comparison, the D37 dose (the dose that on average is necessary to inactivate a cell) for E. coli B/r is approximately 30 Jm-2 [5]. UV exposures as high as those that D. radiodurans tolerates introduce enormous amounts of DNA damage. Exposure to a fluence of 500 Jm-2 UV should generate 5000 thymine-containing pyrimidine dimers per genome in this species [6,7], an average of one lesion of this type for every 640 base pairs. Clearly, D. radiodurans expresses efficient mechanisms for dealing with UV-induced DNA damage, and it is not unreasonable to assume that UV resistance in this species might rely on processes not found in more UV sensitive microorganisms.
D. radiodurans R1 encodes all of the components of the UvrABC-dependent nucleotide excision repair (NER) system characterized in many species including E. coli [8]. In contrast to E. coli, inactivating NER does not reduce D. radiodurans resistance to UV light. NER-defective strains of D. radiodurans only become sensitive to UV when a second locus, designated uvs (DR1819) [8], is also inactivated [9,10]. The uvs gene encodes a UV DNA damage endonuclease similar to that reported in Schizosacchromyces pombe [11,12] that is capable of completely compensating for the loss of NER. Likewise, if the uvs gene is inactivated in an otherwise wild type background, the resulting strain exhibits near wild-type levels of UV resistance, because of the overlapping action of NER. Thus, while the presence of two excision repair systems is a distinctive characteristic of D. radiodurans, the cell does not require this redundancy for UV resistance, making it difficult to argue that the presence of two repair systems contributes significantly to the species’ extraordinary tolerance of UV light. Mutational inactivation of the polA locus of R1 results in strains that are sensitive to all forms of DNA damage [13,14], indicating that DNA polymerase I of D. radiodurans (like its counterpart in E. coli) plays a central role in repair of UV-induced damage, presumably through gap filling after excision repair or in strand extension following recombinational repair. Deletion of recA also results in a strain of D. radiodurans that is extremely sensitive to DNA damage [15,16] presumably due to the cell’s inability to carry out homologous recombination.
From the above discussion, it seems apparent that although D. radiodurans expresses several proteins analogous to those known to counteract the lethal effects of UV light [9] in other species, there is nothing about these proteins that suggests an explanation for the extraordinary UV resistance of D. radiodurans.
Five novel gene products have been linked to the IR resistance of D. radiodurans R1 [16]. The transcripts of these genes – designated ddrA (DR0423), ddrB (DR0070), ddrC (DR0003), ddrD (DR0326), and pprA (DRA0346) – are the five most highly induced in response to IR and desiccation (a stress that like IR induces DNA double strand breaks). Mutants created by deleting these five loci alone and in all possible pairs revealed that each gene product partially contributed to IR resistance, but the functions of these proteins has remained obscure.
In the present study, the same collection of mutants is analyzed to determine if these gene products also affect D. radiodurans’ capacity to survive exposure to UV light and mitomycin C (MC). We report that the PprA, DdrA, and DdrD gene products are components of overlapping processes that make a major contribution to UV resistance. Deletion of PprA from R1 sensitizes R1 to UV light, whereas inactivation of DdrA and DdrD in a pprA + background does not alter UV resistance. When deletions of ddrA or ddrD are combined with a pprA deletion, cultures are as much as 1000-fold more sensitive to the lethal effects of UV light relative to their wild type parent. The functions of these proteins do not appear related to either excision repair or homologous recombination, suggesting they mediate previously undefined mechanisms that facilitate tolerance of UV-induced DNA damage in D. radiodurans.
Materials and Methods
All strains used in this study are described in Table 1. The D. radiodurans strains are derived from the R1 type strain (ATCC13939), and were grown at 30oC in TGY broth (0.8% tryptone, 0.1% glucose, 0.4% yeast extract) or on TGY agar (1.5% agar) as described previously [17,18]. D. radiodurans survival following exposure to UV radiation and mitomycin C was assessed in cultures during exponential phase growth (at a density between 2 x 106 and 4.5 x 107 CFU/ml). Prior to exposure to UV light, cells were harvested by centrifugation and re-suspended in an equivalent volume of sterile saline (0.9% NaCl). One ml aliquots were placed in a sterile Petri dish and irradiated uncovered at 25°C using a germicidal lamp emitting at a calibrated dose rate of 25 Jm-2s-1 UV–C. Irradiated cultures were diluted in saline, prior to plating on TGY agar. Cultures to be exposed mitomycin C were grown to the appropriate density, harvested by centrifugation, and re-suspended in an equivalent volume of TGY broth containing 20 µg/ml mitomycin C. Cultures were incubated at 30oC and aliquots of the MC-treated culture were removed at ten minute intervals, washed twice in an equal volume of TGY broth, diluted, and plated on TGY agar. With the exception of TNK113 ΔddrC ΔddrD, survival was scored using colony counts three days after plating. TNK113 is very slow growing and colony formation was not evident until seven days after plating.
Table 1. Strain List.
| Strain | Description | Reference |
|---|---|---|
| LSU2000 | As R1 but uvrA1, uvs::TnDrCat | [9] |
| TNK101 | As R1 but ΔddrC::pkat-aadA | [16] |
| TNK102 | As R1 but ΔddrB::pkat-cat | [16] |
| TNK103 | As R1 but ΔddrD::pkat-kan | [16] |
| TNK104 | As R1 but ΔddrA::pkat-hyg | [16] |
| TNK105 | As R1 but ΔpprA::pkat-aadA | [16] |
| TNK106 | As R1 but ΔrecA::pkat-cat | [16] |
| TNK112 | ΔddrB::pkat-cat, ΔddrC::pkat-aadA | [16] |
| TNK113 | ΔddrC::pkat-aadA, ΔddrD::pkat-kan | [16] |
| TNK114 | ΔddrA::pkat-hyg, ΔddrC::pkat-aadA | [16] |
| TNK115 | ΔddrC::pkat-aadA, ΔpprA::pkat-aadA | [16] |
| TNK116 | ΔddrB::pkat-cat, ΔddrD::pkat-kan | [16] |
| TNK117 | ΔddrA::pkat-hyg, ΔddrB::pkat-cat | [16] |
| TNK118 | ΔddrB::pkat-cat, ΔpprA::pkat-aadA | [16] |
| TNK119 | ΔddrA::pkat-hyg, ΔddrD::pkat-kan | [16] |
| TNK120 | ΔddrD::pkat-kan, ΔpprA::pkat-aadA | [16] |
| TNK121 | ΔddrA::pkat-hyg, ΔpprA::pkat-aadA | [16] |
All strains are derived from Deinococcus radiodurans R1 ATCC13939.
Results
Deletion of pprA (DRA0346) sensitizes D. radiodurans R1 to UV light
Strains TNK101 ΔddrC, TNK102 ΔddrB, TNK103 ΔddrD, and TNK104 ΔddrA, and TNK105 ΔpprA were evaluated for their ability to tolerate exposure to UV light (Figure 1) relative to their parent. Single deletions of ddrA, ddrB, ddrC, or ddrD have no effect on the UV resistance of D. radiodurans R1. Only deletion of pprA had a demonstrable effect on UV resistance. The reduction in the mean survival of TNK105 ΔpprA becomes statistically significant (unpaired t test, p=0.01, degrees of freedom (df) = 16) relative to R1 at 500 J/m2, at 1000J/m2 TNK105 is approximately eight-fold more sensitive to UV light when compared to R1.
Figure 1. Survival curves for ΔddrA, ΔddrB , ΔddrC, ΔddrD, and ΔpprA derivatives of D. radiodurans R1 strains exposed to UV light.
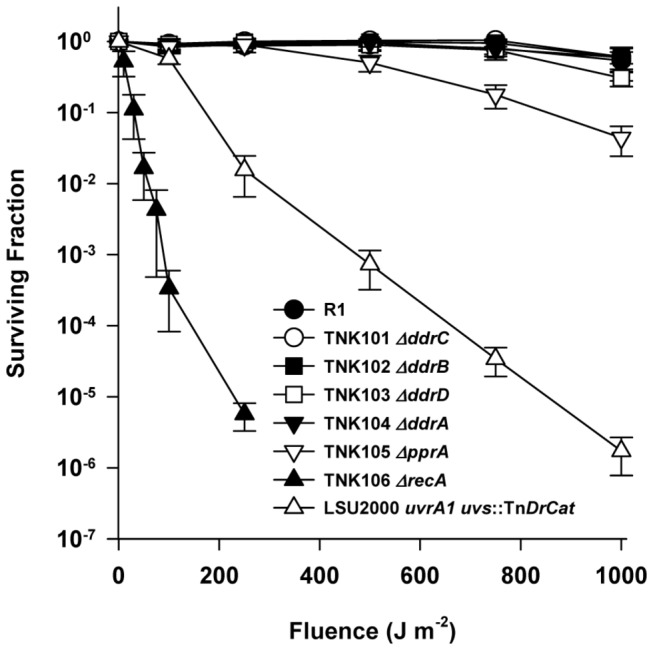
Exponential phase cultures of D. radiodurans R1, TNK101 ΔddrC, TNK102 ΔddrB, TNK103 ΔddrD, TNK104 ΔddrA, TNK105 ΔpprA, TNK106 ΔrecA, and LSU2000 uvrA1 uvs::TnDrCat were exposed to UV light at 25 J m-2s-1. Values are the means ± standard deviations of three independent experiments (n = 9).
For comparison, the survival curves for the UV sensitive strains LSU2000 [9] and TNK106 [16] are also included in Figure 1. LSU2000 carries a deletion in the uvrA1 (DR1771) coding sequence and an insertion into the uvs (DR1819) gene [9], eliminating excision repair by the UvrABC complex and the species’ UV damage endonuclease (UVDE). TNK106 is a ΔrecA strain incapable of RecA-dependent homologous recombination [16].
Deletion of ddrA (DR0423) in a ΔpprA background sensitizes D. radiodurans R1 to UV light
Deleting ddrA has no effect on the UV resistance of R1 (Figure 1), and strains TNK114 ΔddrA ΔddrC, TNK117 ΔddrA ΔddrB, and TNK119 ΔddrA ΔddrD are as resistant TNK104 ΔddrA (Figure 2). However, TNK121 ΔddrA ΔpprA demonstrates a rapid drop in UV resistance; there is a statistically significant (unpaired t test, p<0.0001, df = 22) two-fold reduction in the mean survival of TNK121 relative to TNK105 ΔpprA after exposure to a fluence of 250 Jm-2. TNK121 is 100-fold more UV sensitive than TNK105 ΔpprA (Figure 2) at 1000 Jm-2, indicating that DdrA functions in UV resistance in D. radiodurans, but its effect on survival is only apparent in the absence of a functional PprA.
Figure 2. Survival curves for ΔddrA derivatives of D. radiodurans R1 exposed to UV light.
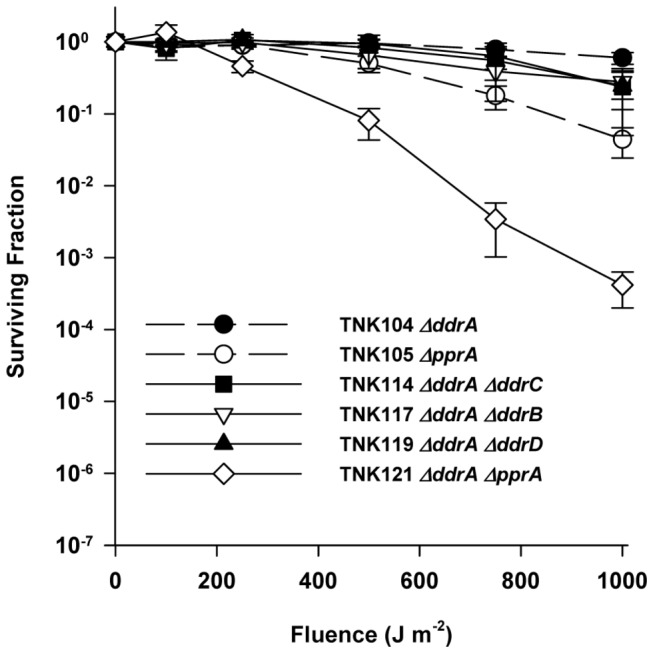
The UV resistances of TNK114 ΔddrA ΔddrC, TNK117 ΔddrA ΔddrB, TNK119 ΔddrA ΔddrD, TNK121 ΔddrA ΔpprA are compared with TNK104 ΔddrA, TNK105 ΔpprA, and D. radiodurans R1. Values are the means ± standard deviations of four independent experiments (n = 12).
The combined deletion of ddrB (DR0070) and ddrC (DR0003) decreases the UV resistance of D. radiodurans R1
We find little evidence that DdrB or DdrC participate in UV resistance in D. radiodurans. TNK101 ΔddrC and TNK102 ΔddrB are as resistant to UV light as R1 (Figure 1). TNK113 ΔddrC ΔddrD and TNK116 ΔddrD ΔddrB do not significantly alter survival relative to TNK101, TNK102, or TNK103 ΔddrD (Figure 3). TNK115 ΔddrC ΔpprA is more sensitive to UV light relative to TNK101 ΔddrC, but its survival curve was identical to that of the UV sensitive strain TNK105 ΔpprA (Figure 1), indicating that loss of PprA alone was responsible for the sensitization observed.
Figure 3. Survival curves for ΔddrB and ΔddrC derivatives of D. radiodurans R1 exposed to UV light.
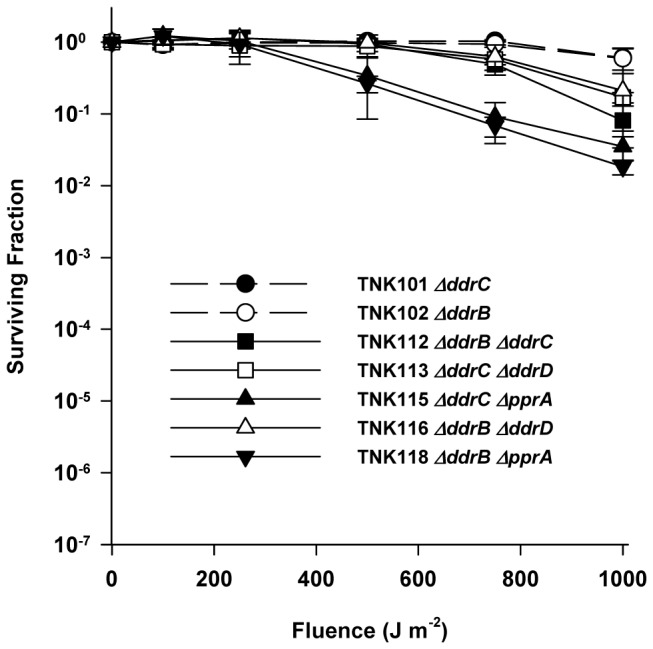
The UV resistances of TNK112 ΔddrB ΔddrC, TNK113 ΔddrC ΔddrD, TNK115 ΔddrC ΔpprA, TNK116 ΔddrB ΔddrD, and TNK118 ΔddrB ΔpprA are compared with TNK101 ΔddrC, and TNK102 ΔddrB. Values are the means ± standard deviations of six independent experiments (n = 18).
The double mutant TNK112 ΔddrB ΔddrC is approximately ten-fold more sensitive than TNK101 or TNK102 (Figure 3) after exposure at 1000 Jm-2. Although this increased sensitivity is reproducible (n=18, six independent trials, three replicates per trial), it is the only evidence that DdrB or DdrC affect UV resistance at the exposures examined.
Deletion of ddrD (DR0326) decreases the UV resistance of the ΔpprA strain TNK105
Deletions of ddrD alone (Figure 1) or in combination with deletions of ddrA (Figure 2), ddrB (Figure 3), or ddrC (Figure 3) do not demonstrate sensitivity to UV light. However, the combination of ΔddrD and ΔpprA in TNK120 dramatically increases UV sensitivity relative to TNK105 ΔpprA (Figure 4). At 250 Jm-2, there is a significant (unpaired t test, p<0.0001, df = 25) ten-fold difference in viability and at 1000 J/m-2 TNK120 is three orders of magnitude more sensitive than TNK105.
Figure 4. The survival curve of TNK120 exposed to UV light.
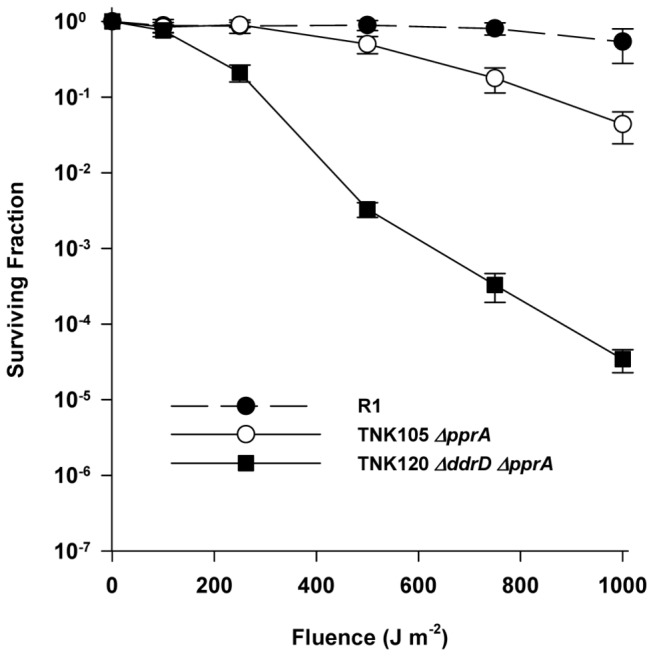
The UV resistances of TNK120 ΔddrD ΔpprA compared with the survival curves of TNK103 ΔddrD, TNK104 ΔddrA, and TNK105 ΔpprA. Values are the means ± standard deviations of four independent experiments (n = 12).
Deletion of ddrA (DR0423) sensitizes D. radiodurans R1 to mitomycin C
To further assess whether DdrA, DdrD, and PprA participate in excision repair or homologous recombination, strains carrying deletions of loci encoding these proteins were evaluated for resistance to the cross-linking agent mitomycin C (MC). Mitomycin C is highly toxic to prokaryotic cells [19]. After enzymatic reduction within the cell, MC generates reactive species that form DNA interstrand cross-links (ICLs) and a variety of guanine monoadducts in the minor groove [20,21]. There is an obligatory requirement for homologous recombination and NER during the repair of MC-induced DNA damage by D. radiodurans [22,23]. At their D37 dose, uvrA and recA defective strains are 60 and 300 times more sensitive to MC than R1, respectively.
TNK103 ΔddrD and TNK105 ΔpprA are as resistant to MC as R1 (Figure 5). In contrast, TNK104 ΔddrA cultures are very sensitive to mitomycin C (MC) relative to R1 (Figure 5) [24]; at each time interval sampled, R1 is between 150- and 400-fold more resistant to this reagent. Combining ΔddrA with ΔpprA does not change MC resistance; the sensitivity of TNK121 ΔddrA ΔpprA is indistinguishable from that of TNK104, indicating that loss of the DdrA is solely responsible for the phenotype. TNK120 ΔddrD ΔpprA is as resistant to MC as TNK103 ΔddrD, TNK105 ΔpprA, and R1 (Figure 5), suggesting that the sensitivity observed when ddrD and pprA are simultaneously deleted (Figure 4) is specific to UV-induced damage.
Figure 5. Survival curves for ΔpprA derivatives D. radiodurans R1 exposed to mitomycin C.
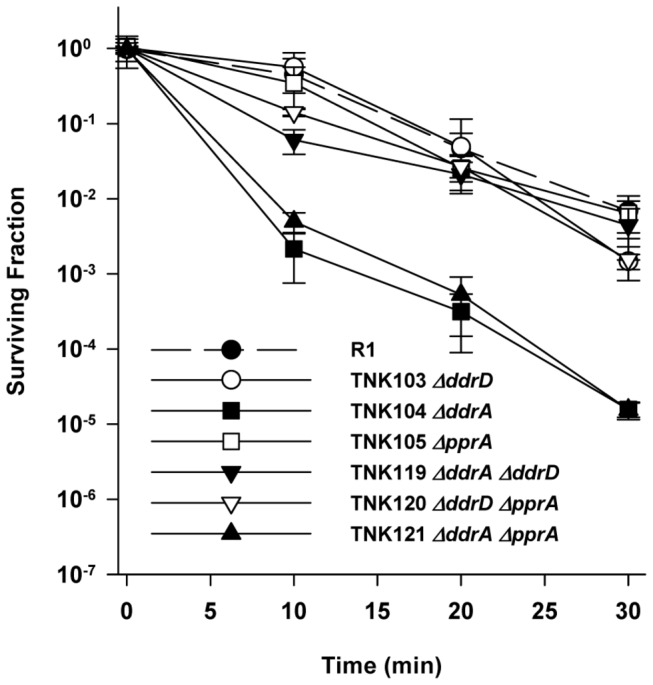
TNK120 ΔddrD ΔpprA and TNK121 ΔddrA ΔpprA are compared with D. radiodurans R1, TNK103 ΔddrD, and TNK105 ΔpprA. Values are the means ± standard deviations of three independent experiments (n = 9).
Deletion of ddrC (DR0003) or ddrD (DR0326) restores mitomycin C resistance to a ΔddrA (DR0423) strain of D. radiodurans R1
Combining the ddrA deletion with deletions of either ddrC or ddrD restored wild type MC resistance to the resulting strains. TNK114 ΔddrA ΔddrC and TNK119 ΔddrA ΔddrD tolerate MC as well as R1 (Figure 6). TNK117 ΔddrA ΔddrB is as MC sensitive as TNK104 ΔddrA. TNK101 and TNK103 are no more sensitive to MC than is R1 (Figure 7). However, the double deletion TNK113ΔddrC ΔddrD exhibits slightly increased sensitivity to MC; there is a significant (unpaired t test, p<0.001, df = 45) 2.5-fold reduction in viability in TNK113 cultures relative to TNK101 and TNK103 at all time points evaluated.
Figure 6. Survival curves for ΔddrC and ΔddrD derivatives of TNK104 ΔddrA exposed to mitomycin C.
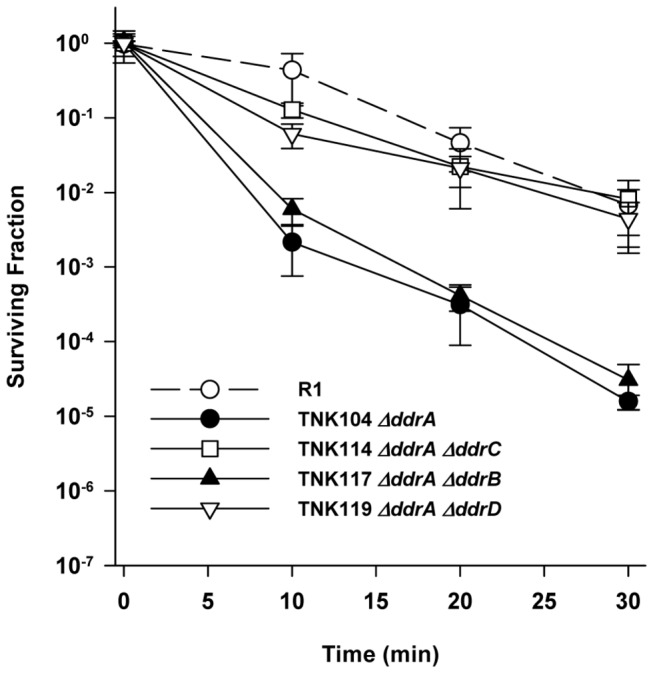
TNK114 ΔddrA ΔddrC, TNK117 ΔddrA ΔddrB, and TNK119 ΔddrA ΔddrD are compared with D. radiodurans R1 and TNK104 ΔddrA. Values are the means ± standard deviations of three independent experiments (n = 9).
Figure 7. Survival curves for ΔddrC and ΔddrD, and ΔpprA derivatives D. radiodurans R1 exposed to mitomycin C.
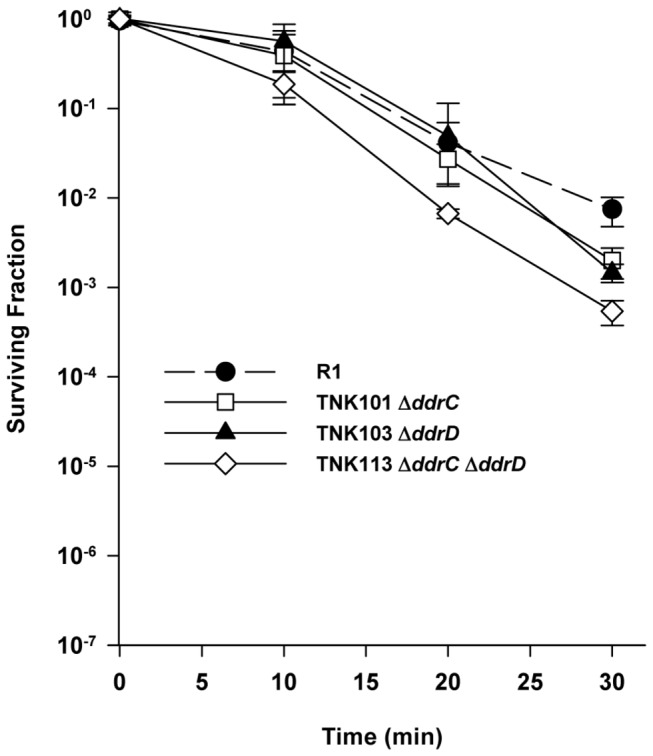
TNK101 ΔddrC, TNK103 ΔddrD, and TNK113 ΔddrC ΔddrD are compared with D. radiodurans R1. Values are the means ± standard deviations of eight independent experiments (n = 24).
Deletion of ddrC (DR0003) or ddrD (DR0326) in a ddrB (DR0070) background significantly reduces the mitomycin resistance of D. radiodurans R1
As illustrated in Figure 8, TNK102 ΔddrB is approximately ten-fold more sensitive to mitomycin C relative to R1 at exposures of 20 minutes or longer. Including the ΔddrC or ΔddrD alleles in a ΔddrB background further reduces cell viability on exposure to this DNA damaging agent. TNK112 ΔddrB ΔddrC and TNK116 ΔddrB ΔddrD are between 100- and 150-fold more sensitive to MC than R1 at all exposure times.
Figure 8. Survival curves for ΔddrC and ΔddrD derivatives of TNK104 ΔddrB exposed to mitomycin C.
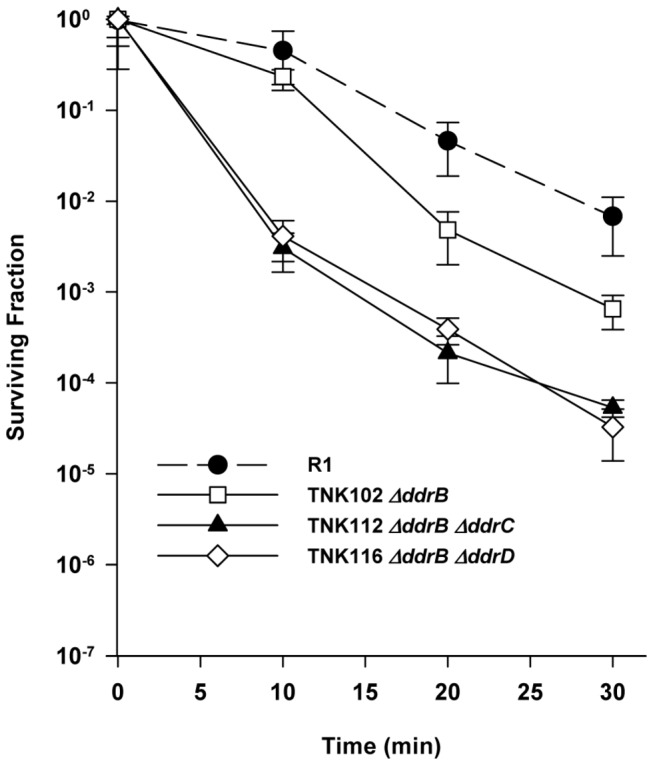
TNK102 ΔddrB, TNK112 ΔddrB ΔddrC, and TNK116 ΔddrB ΔddrD are compared with D. radiodurans R1. Values are the means ± standard deviations of eight independent experiments (n = 24).
Discussion
Deinococcus radiodurans R1 is the type species for a family of bacteria characterized by its exceptional capacity to tolerate DNA damage [2,25,26]. Notably, exponential phase cultures of this species can withstand exposure to 5000Gy ionizing radiation and 500Jm-2 UV light; doses that will all but eradicate many bacterial cultures. Wild-type D. radiodurans cultures are also more resistant to ICLs than are most vegetative bacteria [27,28,29]. This species survives incubation in the presence of 20µg ml-1 MC for 10 min at 30oC with minimal loss of viability. Kitayama [28] reported that under these conditions greater than 90% of isolated genomic DNA exists as non-denaturable double stranded DNA with an average molecular weight of 2 x 107 Daltons, and estimated that this level of exposure corresponds to approximately 100 MC-induced ICLs per D. radiodurans genome [28,29]. Approximately 20 ICLs are sufficient to inactivate repair proficient strains of Escherichia coli B/r [30].
In this study, we have demonstrated that the proteins DdrA, DdrD, and PprA play critical but previously unidentified roles in the UV resistance of D. radiodurans R1. The deletion of pprA in a wild type background sensitizes the resulting strain to UV light (Figure 1) and that sensitivity dramatically increases when ddrA or ddrD is also deleted from this background (Figures 2 and 4). The level of sensitivity we report for TNK120 ΔddrD ΔpprA is comparable to that observed in an uvrA uvs background that cannot carry out NER- and UVDE-mediated excision repair (Figures 1 and 4). TNK121 ΔddrA ΔpprA is approximately ten-fold more UV resistant than TNK120, but the strain remains significantly more sensitive than D. radiodurans R1 (Figure 2). The enhanced UV sensitivity of TNK120 and TNK121 suggest that the activities of PprA overlap with those of DdrA and DdrD; either they catalyze similar functions or they are required for separate processes that have an equivalent effect on UV resistance. The central role of homologous recombination and excision repair in reversing UV and mitomycin C-induced DNA damage in other species has been recognized for many years [31,32] forcing us to consider the possibility that DdrA, DdrD, and PprA contribute to these processes in D. radiodurans.
TNK103 ΔddrD, TNK104 ΔddrA, TNK105 ΔpprA, TNK119 ΔddrA ΔddrD, TNK120 ΔddrD ΔpprA and TNK121 ΔddrA ΔpprA are considered recombination-proficient [16]. These strains undergo natural transformation, a process that requires RecA-dependent recombination, with efficiencies identical to that of R1. In addition, the double mutants TNK119ΔddrA ΔddrD, TNK120 ΔddrD ΔpprA and TNK121 ΔddrA ΔpprA display varied responses to DNA damaging agents. TNK119 is modestly sensitive to IR [16], but as resistant as R1 to UV (Figure 2) and mitomycin C (Figure 6). TNK120 is sensitive to UV (Figure 4), but resistant to IR [16] and mitomycin (Figure 5). TNK121 is sensitive to IR [16], mitomycin (Figure 5), and UV (Figure 2). Since homologous recombination is required to survive exposure to IR, UV, and ICLs in D. radiodurans [22], the different patterns of resistance among these strains suggests that if DdrA, DdrD, or PprA contribute to DNA repair involving homologous recombination, their role is constrained in a manner specific to the DNA damaging agent or the lesions generated by that agent.
For similar reasons, it also seems unlikely that DdrA, DdrD or PprA are affecting excision repair. TNK120 is sensitive to UV and resistant to mitomycin. If one postulates that the inactivation of DdrD and PprA resulted in an excision repair defective strain, these opposing phenotypes cannot be reconciled. In addition, the UV sensitive double mutants TNK120 and TNK121 are wild type with respect to the uvrA, uvrB, uvrC, uvs, and polA genes that encode for excision repair systems capable of dealing with all major forms of UV-induced DNA damage [9]. It is difficult to comprehend how eliminating PprA and DdrA or PprA and DdrD could stop the contributions of NER and UVDE to UV resistance unless combined loss of these proteins influence UV resistance by affecting the function of both excision repair pathways simultaneously – in effect, recreating the situation that arises when the NER and UVDE systems are both inactivated. This scenario could occur if loss of DdrA and PprA or DdrD and PprA affected the stability of an intermediate or the activity of a component shared by the two pathways. At present, there is no evidence to support this idea.
The function of PprA is obscure; the protein has not been extensively characterized. Although homologues of PprA are encoded by all sequenced Deinococcus species [33,34,35], this protein shares no similarity to any other protein, amino acid sequence motif, or conserved domain described in the protein databases. Purified PprA is reported to stabilize the free ends formed at DNA double strand breaks and to recruit DNA ligase to the site of these breaks, improving the efficiency of ligation in vitro [36]. This report led to speculation that PprA performs the same function in vivo, and a suggestion that PprA participates in a form of non-homologous end joining (NHEJ) [37,38]. However, attempts to demonstrate NHEJ activity following D. radiodurans exposure to IR have failed to provide convincing evidence that this process is taking place [39,40]. While the results presented here do not directly support or refute the notion that D. radiodurans expresses the proteins needed for NHEJ, it is difficult to reconcile PprA’s central role in UV resistance with its possible involvement in NHEJ. Precedent suggests that NHEJ can play a minor role in repair of UV-induced DNA damage in bacteria [41]. The combined deletion of ykoU and ykoV of Bacillus subtilis, proteins that mediate NHEJ in this species, increases the strain’s sensitivity to UV light, but the effect is small, between one and a half and two-fold relative to the wild type at doses to 500 Jm-2. In contrast, the combined loss of PprA and DdrD results in a strain that is as much as 1000-fold more UV sensitive relative to the wild type (Figure 4). Whatever its activity, PprA makes a substantive contribution to UV resistance, and it seems doubtful that any contribution PprA may make to NHEJ fully defines its function following exposure to UV.
The ability of PprA to bind to the free ends created by DNA double strand breaks may explain why TNK121 ΔddrA ΔpprA is more sensitive to UV than TNK105 ΔpprA (Figure 2). DdrA is part of the Rad52 family of proteins [42]. In eukaryotes Rad52 promotes recombination and DNA double strand break repair through interaction with the Rad51 recombinase [43]. While DdrA contributes to the ionizing radiation resistance of D. radiodurans [16,24], there is no evidence that it behaves like the eukaryotic Rad52; the protein does not display DNA strand annealing activity [24]. In D. radiodurans, DdrA appears to function in vitro and in vivo by binding to the 3’ ends of single-stranded DNA and preventing DNA digestion by endogenous exonucleases following the cell’s exposure to high dose ionizing radiation [24]. It has been proposed that DdrA is part of a DNA end-protection system that helps to preserve genome integrity following DNA damage. In this capacity, it is not difficult to envision a situation where DdrA and PprA protect an intermediate created during repair of UV damage, and evidence of this protection is only obvious when both proteins are inactive.
The apparent overlap in DdrD and PprA activity is not as easily explained. Each protein appears to be a component of an independent process that facilitates UV resistance. DdrD function is unknown and this protein, which is found only in the Deinococci , shares no amino acid sequence similarity with any other characterized protein or sequence motif [33,34,35]. Inactivation of both proteins reveals their roles in UV resistance, but it is difficult to argue the DdrD acts to protect DNA as described for PprA and DdrA. PprA function is necessary if cells are to survive IR-induced damage, but DdrD seems to have only a minor role in IR resistance [16].
Figure 6 may provide a clue to DdrD function. TNK104 ΔddrA is two orders of magnitude more sensitive to mitomycin C than D. radiodurans R1. If deletions of ddrC or ddrD are inserted in this background, mitomycin resistance is restored to wild type levels. This result suggests that DdrC and DdrD are at least partially responsible for the increased sensitivity to mitomycin C observed in a ddrA strain. In other words, it appears that DdrA prevents a lethal event caused, directly or indirectly, by the wild type DdrC and DdrD proteins, and when either protein is inactivated that event is avoided. Like DdrD, DdrC shares no similarity with other proteins and is only known to be encoded by members of the genus Deinococcus .
Since inactivating DdrC and DdrD has the same effect in a ΔddrA background (Figure 6) or ΔddrB background (Figure 8), it is reasonable to ask if these proteins are part of the same repair complex. This possibility seems unlikely given the differences in UV sensitivity associated with TNK115 ΔddrC ΔpprA (Figure 3) and TNK120 ΔddrD ΔpprA (Figure 4); TNK120 is 1000 fold more UV sensitive when compared with TNK115. In addition, the simultaneous inactivation of ddrC and ddrD results in a strain with slightly decreased mitomycin C resistance (Figure 7). If DdrC and DdrD were different parts of a protein complex that carried out a single function, survival of the double mutant should not be different than TNK101 ΔddrC or TNK103 ΔddrD. Pending further investigation, we argue that the DdrC and DdrD proteins are components of different complementary processes involved in DNA damage tolerance.
At present, the phenotypes reported here are perhaps best explained by assuming that DdrA, DdrD, and PprA affect the efficiency of UV- and MC-induced lesion repair without directly catalyzing the removal of those lesions. We posit that these proteins suppress potentially lethal events that arise as repair proceeds following massive genetic insult, thereby facilitating survival. If this occurs, it may provide a context in which to better understand the extreme resistance of D. radiodurans to DNA damaging agents. We contend that these proteins assist the cell by allowing the cell enough time to achieve necessary repairs. Repair of damage after insult should be more effective if the time available for repair – the time between the appearance of damage and the lethal consequences of that damage – is extended. Assuming that a bacterial cell has the necessary repair proteins and sufficient time to carry out repairs, extreme tolerance to multiple DNA damaging agents could be conveniently explained.
Acknowledgments
The authors wish to thank L. Alice Simmons, Danielle R. Fontenot, and Blythe E. Johnson for their technical assistance in determining the UV survival curves.
Funding Statement
This work was supported by Grant DEFG0201ER63151 from the United States Department of Energy. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Battista JR, Rainey FA (2001) Phylum BIV. "Deinococcus-Thermus" Family 1. Deinococcaceae Brooks and Murray 1981, 356,vp emend. Rainey, Nobre, Schumann, Stackebrandt and da Costa 1997, 513. In: Boone DR, Castenholz RW. Bergey's Manual of Systematic Bacteriology. 2nd ed. New York: Springer Verlag; pp. 395-414. [Google Scholar]
- 2. Cox MM, Battista JR (2005) Deinococcus radiodurans - the consummate survivor. Nat Rev Microbiol 3: 882-892. doi:10.1038/nrmicro1264. PubMed: 16261171. [DOI] [PubMed] [Google Scholar]
- 3. Moseley BEB (1983) Photobiology and radiobiology of Micrococcus (Deinococcus) radiodurans . Photochem Photobiol Rev 7: 223-275. [Google Scholar]
- 4. Moseley BE, Mattingly A (1971) Repair of irradiation transforming deoxyribonucleic acid in wild type and a radiation-sensitive mutant of Micrococcus radiodurans . J Bacteriol 105: 976-983. PubMed: 4929286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Sweet DM, Moseley BE (1974) Accurate repair of ultraviolet-induced damage in Micrococcus radiodurans . Mutat Res 23: 311-318. doi:10.1016/0027-5107(74)90104-3. PubMed: 4407662. [DOI] [PubMed] [Google Scholar]
- 6. Boling ME, Setlow JK (1966) The resistance of Micrococcus radiodurans to ultraviolet radiation. 3. A repair mechanism. Biochim Biophys Acta 123: 26-33. doi:10.1016/0005-2787(66)90155-9. PubMed: 5964047. [DOI] [PubMed] [Google Scholar]
- 7. Varghese AJ, Day RS 3rd (1970) Excision of cytosine-thymine adduct from the DNA of ultraviolet-irradiated Micrococcus radiodurans . Photochem Photobiol 11: 511-517. doi:10.1111/j.1751-1097.1970.tb06022.x. PubMed: 5456276. [DOI] [PubMed] [Google Scholar]
- 8. White O, Eisen JA, Heidelberg JF, Hickey EK, Peterson JD et al. (1999) Genome sequence of the radioresistant bacterium Deinococcus radiodurans R1. Science 286: 1571-1577. doi:10.1126/science.286.5444.1571. PubMed: 10567266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Earl AM, Rankin SK, Kim KP, Lamendola ON, Battista JR (2002) Genetic evidence that the uvsE gene product of Deinococcus radiodurans R1 is a UV damage endonuclease. J Bacteriol 184: 1003-1009. doi:10.1128/jb.184.4.1003-1009.2002. PubMed: 11807060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Moseley BE, Evans DM (1983) Isolation and properties of strains of Micrococcus (Deinococcus) radiodurans unable to excise ultraviolet light-induced pyrimidine dimers from DNA: evidence for two excision pathways. J Gen Microbiol 129(8): 2437-2445. PubMed: 6415229. [DOI] [PubMed] [Google Scholar]
- 11. Bowman KK, Sidik K, Smith CA, Taylor JS, Doetsch PW et al. (1994) A new ATP-independent DNA endonuclease from Schizosaccharomyces pombe that recognizes cyclobutane pyrimidine dimers and 6-4 photoproducts. Nucleic Acids Res 22: 3026-3032. doi:10.1093/nar/22.15.3026. PubMed: 8065916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Takao M, Yonemasu R, Yamamoto K, Yasui A (1996) Characterization of a UV endonuclease gene from the fission yeast Schizosaccharomyces pombe and its bacterial homolog. Nucleic Acids Res 24: 1267-1271. doi:10.1093/nar/24.7.1267. PubMed: 8614629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Gutman PD, Fuchs P, Minton KW (1994) Restoration of the DNA damage resistance of Deinococcus radiodurans DNA polymerase mutants by Escherichia coli DNA polymerase I and Klenow fragment. Mutat Res 314: 87-97. doi:10.1016/0921-8777(94)90064-7. PubMed: 7504195. [DOI] [PubMed] [Google Scholar]
- 14. Gutman PD, Fuchs P, Ouyang L, Minton KW (1993) Identification, sequencing, and targeted mutagenesis of a DNA polymerase gene required for the extreme radioresistance of Deinococcus radiodurans . J Bacteriol 175: 3581-3590. PubMed: 8501062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Gutman PD, Carroll JD, Masters CI, Minton KW (1994) Sequencing, targeted mutagenesis and expression of a recA gene required for the extreme radioresistance of Deinococcus radiodurans . Gene 141: 31-37. doi:10.1016/0378-1119(94)90124-4. PubMed: 8163172. [DOI] [PubMed] [Google Scholar]
- 16. Tanaka M, Earl AM, Howell HA, Park MJ, Eisen JA et al. (2004) Analysis of Deinococcus radiodurans’ transcriptional response to ionizing radiation and desiccation reveals novel proteins that contribute to extreme radioresistance. Genetics 168: 21-33. doi:10.1534/genetics.104.029249. PubMed: 15454524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Earl AM, Mohundro MM, Mian IS, Battista JR (2002) The IrrE protein of Deinococcus radiodurans R1 is a novel regulator of recA expression. J Bacteriol 184: 6216-6224. doi:10.1128/JB.184.22.6216-6224.2002. PubMed: 12399492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Mattimore V, Udupa KS, Berne GA, Battista JR (1995) Genetic characterization of forty ionizing radiation-sensitive strains of Deinococcus radiodurans: linkage information from transformation. J Bacteriol 177: 5232-5237. PubMed: 7665511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Dronkert ML, Kanaar R (2001) Repair of DNA interstrand cross-links. Mutat Res 486: 217-247. doi:10.1016/S0921-8777(01)00092-1. PubMed: 11516927. [DOI] [PubMed] [Google Scholar]
- 20. Tomasz M, Chowdary D, Lipman R, Shimotakahara S, Veiro D et al. (1986) Reaction of DNA with chemically or enzymatically activated mitomycin C: isolation and structure of the major covalent adduct. Proc Natl Acad Sci U S A 83: 6702-6706. doi:10.1073/pnas.83.18.6702. PubMed: 3018744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Tomasz M, Lipman R, Lee MS, Verdine GL, Nakanishi K (1987) Reaction of acid-activated mitomycin C with calf thymus DNA and model guanines: elucidation of the base-catalyzed degradation of N7-alkylguanine nucleosides. Biochemistry 26: 2010-2027. doi:10.1021/bi00381a034. PubMed: 3109476. [DOI] [PubMed] [Google Scholar]
- 22. Moseley BE, Copland HJ (1975) Isolation and properties of a recombination-deficient mutant of Micrococcus radiodurans . J Bacteriol 121: 422-428. PubMed: 1112771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Moseley BE, Copland HJR (1978) Four mutants of Micrococcus radiodurans defective in the ability to repair DNA damaged by mitomycin-C, two of which have wild-type resistance to ultraviolet radiation. Mol Gen Genet 160: 331-337. doi:10.1007/BF00332977. PubMed: 672894. [DOI] [PubMed] [Google Scholar]
- 24. Harris DR, Tanaka M, Saveliev SV, Jolivet E, Earl AM et al. (2004) Preserving Genome Integrity: the DdrA protein of Deinococcus radiodurans R1. PLOS Biol 10: e304 PubMed: 15361932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Blasius M, Sommer S, Hübscher U (2008) Deinococcus radiodurans: what belongs to the survival kit? Crit Rev Biochem Mol Biol 43: 221-238. doi:10.1080/10409230802122274. PubMed: 18568848. [DOI] [PubMed] [Google Scholar]
- 26. Slade D, Radman M (2011) Oxidative stress resistance in Deinococcus radiodurans . Microbiol Mol Biol Rev 75: 133-191. doi:10.1128/MMBR.00015-10. PubMed: 21372322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Hansen MT (1982) Rescue of mitomycin C- or psoralen-inactivated Micrococcus radiodurans by additional exposure to radiation or alkylating agents. J Bacteriol 152: 976-982. PubMed: 6815165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Kitayama S (1982) Adaptive repair of cross-links in DNA of Micrococcus radiodurans . Biochim Biophys Acta 697: 381-384. doi:10.1016/0167-4781(82)90103-8. PubMed: 7104365. [DOI] [PubMed] [Google Scholar]
- 29. Kitayama S, Asaka S, Totsuka K (1983) DNA double-strand breakage and removal of cross-links in Deinococcus radiodurans . J Bacteriol 155: 1200-1207. PubMed: 6411683. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Lawley PD, Brookes P (1968) Cytotoxicity of alkylating agents towards sensitive and resistant strains of Escherichia coli in relation to extent and mode of alkylation of cellular macromolecules and repair of alkylation lesions in deoxyribonucleic acids. Biochem J 109: 433-447. PubMed: 4879534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31. Howard-Flanders P, Boyce RP (1966) DNA repair and genetic recombination: studies on mutants of Escherichia coli defective in these processes. Radiat Res Suppl: 6:156+. PubMed; : 5334390 [PubMed] [Google Scholar]
- 32. Howard-Flanders P, Theriot L (1966) Mutants of Escherichia coli K-12 defective in DNA repair and in genetic recombination. Genetics 53: 1137-1150. PubMed: 5335129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. de Groot A, Dulermo R, Ortet P, Blanchard L, Guérin P et al. (2009) Alliance of proteomics and genomics to unravel the specificities of Sahara bacterium Deinococcus deserti . PLOS Genet 5: e1000434 PubMed: 19370165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Makarova KS, Aravind L, Wolf YI, Tatusov RL, Minton KW et al. (2001) Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics. Microbiol Mol Biol Rev 65: 44-79. doi:10.1128/MMBR.65.1.44-79.2001. PubMed: 11238985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Yuan M, Zhang W, Dai S, Wu J, Wang Y et al. (2009) Deinococcus gobiensis sp. nov., an extremely radiation-resistant bacterium. Int J Syst Evol Microbiol 59: 1513-1517. doi:10.1099/ijs.0.004523-0. PubMed: 19502345. [DOI] [PubMed] [Google Scholar]
- 36. Narumi I, Satoh K, Cui S, Funayama T, Kitayama S et al. (2004) PprA: a novel protein from Deinococcus radiodurans that stimulates DNA ligation. Mol Microbiol 54: 278-285. doi:10.1111/j.1365-2958.2004.04272.x. PubMed: 15458422. [DOI] [PubMed] [Google Scholar]
- 37. Kota S, Misra HS (2006) PprA: A protein implicated in radioresistance of Deinococcus radiodurans stimulates catalase activity in Escherichia coli . Appl Microbiol Biotechnol 72: 790-796. doi:10.1007/s00253-006-0340-7. PubMed: 16586106. [DOI] [PubMed] [Google Scholar]
- 38. Misra HS, Khairnar NP, Kota S, Shrivastava S, Joshi VP et al. (2006) An exonuclease I-sensitive DNA repair pathway in Deinococcus radiodurans: a major determinant of radiation resistance. Mol Microbiol 59: 1308-1316. doi:10.1111/j.1365-2958.2005.05005.x. PubMed: 16430702. [DOI] [PubMed] [Google Scholar]
- 39. Daly MJ, Ling O, Minton KW (1994) Interplasmidic recombination following irradiation of the radioresistant bacterium Deinococcus radiodurans . J Bacteriol 176: 7506-7515. PubMed: 8002574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40. Zahradka K, Slade D, Bailone A, Sommer S, Averbeck D et al. (2006) Reassembly of shattered chromosomes in Deinococcus radiodurans . Nature 443: 569-573. PubMed: 17006450. [DOI] [PubMed] [Google Scholar]
- 41. Moeller R, Stackebrandt E, Reitz G, Berger T, Rettberg P et al. (2007) Role of DNA repair by nonhomologous-end joining in Bacillus subtilis spore resistance to extreme dryness, mono- and polychromatic UV, and ionizing radiation. J Bacteriol 189: 3306-3311. doi:10.1128/JB.00018-07. PubMed: 17293412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Iyer LM, Koonin EV, Aravind L (2002) Classification and evolutionary history of the single-strand annealing proteins, RecT, Redβ, ERF and RAD52. BMC Genomics 3: 8. doi:10.1186/1471-2164-3-8. PubMed: 11914131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Mortensen UH, Bendixen C, Sunjevaric I, Rothstein R (1996) DNA strand annealing is promoted by the yeast Rad52 protein. Proc Natl Acad Sci U S A 93: 10729-10734. doi:10.1073/pnas.93.20.10729. PubMed: 8855248. [DOI] [PMC free article] [PubMed] [Google Scholar]