

Abstract
Invasive species, range-expanding species, genetically modified organisms (GMOs), synthetic organisms, and emerging pathogens increasingly affect the human environment. We propose a framework that allows comparison of consecutive stages that such novel organisms go through. The framework provides a common terminology for novel organisms, facilitating knowledge exchange among researchers, managers, and policy makers that work on, or have to make effective decisions about, novel organisms. The framework also indicates that knowledge about the causes and consequences of stage transitions for the better studied novel organisms, such as invasive species, can be transferred to more poorly studied ones, such as GMOs and emerging pathogens. Finally, the framework advances understanding of how climate change can affect the establishment, spread, and impacts of novel organisms, and how biodiversity affects, and is affected by, novel organisms.
Keywords: Biological invasions, Ecological novelty, Emerging diseases, Genetically modified organisms, Range-expanding species, Synthetic organisms
Introduction
Recent efforts have attempted to integrate research on “ecological novelty” (Kueffer et al. 2011), “novel ecosystems” (Hobbs et al. 2009), and “novel organisms”, where the latter are organisms that are either novel anywhere [genetically modified organisms (GMOs), synthetic organisms, some emerging pathogens] or novel in a given environment (invasive non-native species, range-expanding species). Novel organisms are an important part of global change and increasingly affect the human environment. In Europe, for instance, there are over 10,000 non-native species, many of which are known to have an ecological or economic impact, e.g., the Chinese mitten crab (Eriocheir sinensis), insects such as the thrips Frankliniella occidentalis and Heliothrips haemorrhoidalis, or the Japanese knotweed (Fallopia japonica) (Vilà et al. 2010). Kettunen et al. (2008) estimated that total costs caused by invasive species in Europe probably exceed €20 billion per year.
Novel organisms have largely been studied in isolation, e.g., invasion biologists have focused on invasive species, and disease ecologists have focused on emerging pathogens. Integrating research on novel organisms promises to reveal previously overlooked similarities between such organisms, and it facilitates exchanges of knowledge, ideas, and research methods among researchers, managers, and policy makers working on different novel organisms. We aim to advance this integration by providing a framework that illustrates similarities among novel organisms (Fig. 1). We suggest that an integrated understanding of novel organisms will allow for improved monitoring, evaluation, and management of novel organisms.
Fig. 1.
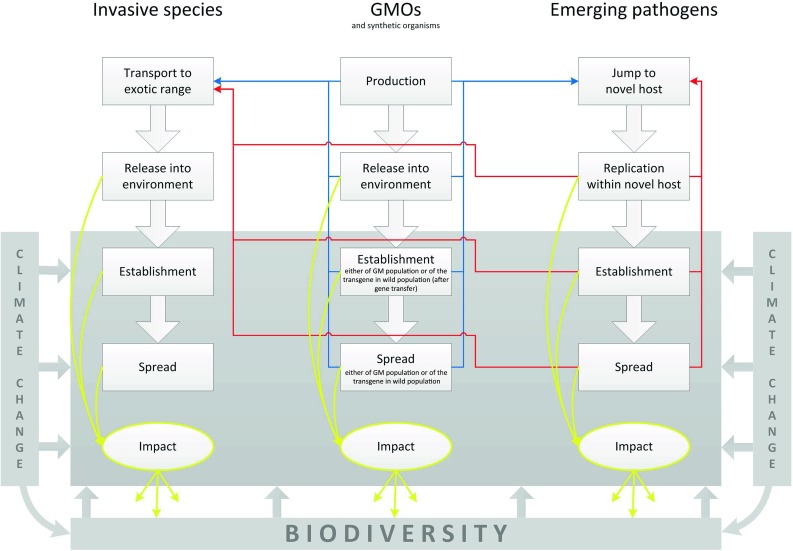
Proposed framework for comparing novel organisms. Invasive species, range-expanding species, GMOs, synthetic organisms, and emerging pathogens go through consecutive stages drawn as framed boxes. For example, transport, release, establishment, and spread for invasive species. The process can be stopped at each transition between two stages (drawn as thick white arrows), for example, a pathogen that has jumped to and replicated within a novel host (stage 2) may not establish itself in the novel host (e.g., Lyme disease with humans as novel host species). Those species that are released or established, and especially those that have spread can have severe environmental and economic impacts (yellow arrows), for example, by influencing biodiversity. To improve clarity, range-expanding species are not depicted explicitly. They are similar to invasive species, except they are not transported to a non-native range and released there, but instead are naturally dispersing to a non-native range where climate change may facilitate their establishment and spread. Blue (for GMOs) and red (for pathogens) arrows starting from different stages and pointing to the transport stage of invasive species illustrate that both GMOs and pathogens can become invasive, too: they can be transported beyond the region where they were first produced (GMOs) or where they first jumped to and replicated within a novel host (emerging pathogens). The probability that they will be transported beyond this region increases with their abundance, so it is higher for later stages of their processes (establishment and spread). Arrows pointing at the jump-to-novel-host stage of emerging pathogens illustrate that GMOs can serve as novel hosts (blue arrows) and that emergent pathogens can be a source for further novel pathogens (red arrows), again with higher probabilities in later stages of the processes. Invasive species can serve as novel hosts as well (not illustrated to improve the figure’s clarity). The grey-shaded boxes illustrate that altered climate and biodiversity affect probabilities of species to establish, spread, and have impacts. The effect of climate change is therefore both direct (because the new climate can, for example, better match a species’ fundamental ecological niche) and indirect (e.g., by reducing biodiversity). Biodiversity itself is also changed by impacts of novel organisms
The framework is based on the rich theory and conceptual foundation that has been developed for invasive species in the last decades (e.g., Lockwood et al. 2007). The literature about other types of novel organisms is not as well developed conceptually, and we argue that some concepts developed for invasive species are also suitable for other novel organisms. A particularly useful idea in the invasion literature has been the “invasion process” which distinguishes the consecutive stages that invasive species go through (Jeschke and Strayer 2005; Hellmann et al. 2008; Walther et al. 2009; Blackburn et al. 2011; and references therein). Although it is somewhat arbitrary into how many stages the invasion process should be divided, a typical sequence of stages is: (1) transport (intentionally or accidentally) of the species from its native to a non-native range → (2) release or escape of the species from captivity in the non-native range → (3) establishment of one or more self-sustaining populations → (4) spread. The concept of the invasion process has proven fruitful not only for researchers but also managers and policy makers, as it has, for instance, helped develop cost-effective barriers and management actions at different stages of the invasion process (Clout and Williams 2009). As we will show here, similar processes also apply to other types of novel organisms, including emerging pathogens which are not typically considered in the new field of the ecology of novel organisms (Kueffer et al. 2011).
Similarities Among Novel Organisms
All novel organisms go through similar stages. Compared to the stages that invasive species go through (described above and illustrated in Fig. 1), range-expanding species naturally disperse into a new range rather than being actively transported and released by humans. Hence, the first stage for range-expanding species is natural dispersal (replacing the stages “transport” and “release”), whereas the later stages are the same as for invasive species: establishment and spread. Range-expanding species are expanding or shifting their range, e.g., as a result of climate change, which enables them to establish and spread in environments that were previously unsuitable for them. Some authors have suggested that the definition of “invasive species” be extended to include range-expanding species, stressing the similarities between these two types of novel organisms (Hellmann et al. 2008; Walther et al. 2009; Engel et al. 2011).
The consecutive stages that GMOs and synthetic organisms go through are similar as well: they undergo production, release, establishment, and spread (Fig. 1; Sharples 1982; Regal 1986, 1993; Williamson 1993; Kowarik 2010). A GMO is defined as an organism in which the genetic material has been altered in a way that does not occur naturally by mating and/or natural recombination (EC 2001). Typically, this genetic alteration is due to the introduction of one or more foreign genes, which are called transgenes. Once released, populations of GMOs might establish or spread themselves, or the transgene(s) might establish or spread in non-GM populations via gene transfer. Consideration of the potential release, establishment, and spread of GMOs is highly relevant for their environmental risk assessment [see e.g., the Cartagena Protocol on Biosafety (Secretariat of the Convention on Biological Diversity 2000) or EFSA 2010]. Several studies have been published with evidence for the establishment of transgenes in the wild (e.g., Reichman et al. 2006; Warwick et al. 2008). Synthetic organisms are completely synthesized by humans; their genomes are typically built by assembling short DNA sequences (“bio-bricks”) (Preston 2008; Deplazes and Huppenbauer 2009). No case of a synthetic organism establishing in the wild is currently known. Nevertheless, considering how different synthetic organisms can be from other organisms, it seems important to assess their potential environmental risks.
Pathogens can emerge via a variety of pathways. For instance, rapid evolution of a microbe can result in a major change in its pathogenicity, transmissibility, or ability to invade a new host species. “Emergence events” can also be caused by long-distance dispersal of a pathogen or its host or vector. Others are caused by the movements of a potential host, previously unexposed to a pathogen, into areas where it is vulnerable to infection.
Emerging pathogens can be novel everywhere, similar to GMOs and synthetic organisms, whereas invasive and range-expanding species are novel only in their non-native range. One example is the rapid evolution of antibiotic-resistant bacteria, which are functionally distinct from their wild-type ancestors. Although the first stages that emerging pathogens go through do not exactly match those of other types of novel organisms, the overall process is similar: (1) jump to a novel host → (2) replication within the novel host → (3) establishment within the host population → (4) spread (Fig. 1; see also Hudson et al. 2008; Hatcher et al. 2012). Most of the emerging pathogens of humans are, or were, zoonotic, meaning that they are transmitted to humans from non-human vertebrates (Taylor et al. 2001; Woolhouse et al. 2005; Jones et al. 2008). Zoonotic pathogens, therefore, must “jump” from the native, non-human host to humans; this stage is analogous to anthropogenic transport of a species to a novel habitat. Indeed, some cases of emergence (e.g., that of West Nile virus in North America) consist of both the human transport of the pathogen (probably via the accidental importation of mosquito vectors from the Middle East to New York City) and species jumps from avian reservoir hosts, via mosquitoes, to humans (LaDeau et al. 2008). For emerging pathogens of non-human animals, plants, and microbes, the situation is the same—species jumps with or without long-distance transport often cause the emergence event. Following the reservoir-to-human species jump, the pathogen typically must replicate in the novel human host for it to cause disease (Wolfe et al. 2007; Lloyd-Smith et al. 2009). Genetic changes in the pathogen that facilitate replication in the novel host can either precede or follow the species jump. These genetic changes, often in response to a different suite of immune defenses in the novel host, can lead to tight co-adaptation between the pathogen and novel host. The same or additional genetic changes can modify a pathogen so that it becomes transmissible directly between individual humans. For example, HIV/AIDS is caused by pathogens that circulated within non-human primate hosts, jumped to humans, and eventually evolved to specialize on humans. Establishment of the emerging disease can accompany either sustained transmission from zoonotic hosts to humans (with or without a vector), or human-to-human transmission. Establishment can be followed by spatial spread, again in parallel to other novel organisms.
Organisms Can be Novel in Multiple Ways
An important insight from comparing types of novel organisms is that some can be novel in multiple ways. In particular, GMOs, synthetic organisms, and emerging pathogens can become invasive, just as more “traditional” invasive species can (cf. Perkins et al. 2008). As mentioned above, GMOs, synthetic organisms, and some emerging pathogens have a higher level of novelty than invasive species, as they are novel to any region, whereas invasive and range-expanding species are only novel to regions beyond their native range. As a result, GMOs, synthetic organisms, and emerging pathogens can go through two processes: process 1 can lead to their establishment and spread in their original environment (drawn as thick white arrows in Fig. 1), and process 2 can lead to their transport, release, establishment, and spread in other regions of the world [as invasive organisms; drawn as blue (for GMOs, synthetic organisms) and red (for pathogens) arrows in Fig. 1].
Although we are unaware of a GMO completing all stages of process 2, examples exist of GMOs completing either the first stage or stages one and two. For instance, GloFish, fluorescent GM fish, have been produced in the U.S. and sold on the U.S. market since 2003. Even though they have not been approved for the European market, they have often been illegally transported to and sold in Europe, where they can now be found in many people’s aquaria. They have thus completed the first stage (transport) of process 2. Seeds and pollen of GM crops can be passively dispersed (e.g., by wind), causing their unintentional transport over possibly large distances (Warwick et al. 2008). They have thus completed stages one and two (transport and release) of process 2.
In the case of emerging pathogens, it is well known that on airplanes, pathogens can be transported to almost any place in the world within 1–2 days (similarly to other microbes or insects). They are thus able to rapidly complete stages one and two of process 2. The importance of the large-scale spread of pathogens is reflected by the inclusion of many pathogens in listings of particularly harmful invaders (e.g., in the Global Invasive Species Database’s “100 of the World’s Worst Invasive Alien Species”; www.issg.org/database; see also Hatcher et al. 2012). Invasive species, range-expanding species, GMOs, and synthetic organisms can all also serve as novel hosts for emerging pathogens (indicated in Fig. 1 for GMOs and synthetic organisms by blue arrows pointing at the jump-to-novel-host stage of emerging pathogens; not shown in Fig. 1 for invasive species and range-expanding species in order to improve the figure’s clarity). Thus, different types of novel organisms are highly interconnected.
Knowledge Exchange About Novel Organisms
This section develops promising examples for knowledge exchange about novel organisms among researchers, managers, and policy makers. The examples are organized around two broad questions: Which organisms and with which traits tend to become successful novel organisms? And, Which ecosystems are especially susceptible to novel organisms?
Which Organisms and with Which Traits Tend to Become Successful Novel Organisms?
In invasion ecology, recent research has shown that the traits determining transition probabilities in the invasion process are stage-dependent. In particular, traits that facilitate transport and release often do not facilitate establishment and spread (Cassey et al. 2004; Jeschke and Strayer 2006; Blackburn and Jeschke 2009). Typically with invasive species, the transport and release stages are directly influenced by humans, whereas the establishment and spread stages depend more on environmental conditions (which may, however, be influenced by humans, e.g., due to human-driven climate change, Fig. 1 and below). The same is true for GMOs and synthetic organisms, where humans produce and release (intentionally or accidentally) organisms, but establishment and spread of organisms is to a greater degree influenced by environmental conditions. In the case of emerging pathogens, we might expect, a priori, similar patterns. For example, the first two stages of emergence (jump to and replication within a novel host) are either directly influenced by humans (e.g., human activities putting pathogens in closer contact with novel hosts) or chance events, whereas establishment and spread might be largely influenced by environmental conditions. In other words, the factors that promote initial species jumps might not also promote establishment or subsequent spread.
Due to these differences in human influence among the stages, it is not surprising that the traits correlated with high transition probabilities between stages vary depending on the stage. For example, for invasive vertebrates in Europe and North America, species being hunted by humans have a significantly higher probability of being transported and released into a non-native range than species that are not hunted by humans (Jeschke and Strayer 2006). However, their probability of becoming established and spreading tends to be lower than for other species. Overall, hunted vertebrates (e.g., muskrat, Ondatra zibethicus, Canada goose, Branta canadensis, or brook trout, Salvelinus fontinalis) are more often invasive than non-hunted vertebrates, but this is only due to their higher probability of being transported and released (stages 1 and 2), and cannot be explained by differences in establishment and spread (stages 3 and 4; Jeschke and Strayer 2006). Similar observations were made for other characteristics of species, for example, vertebrates with a large native range are more often invasive than vertebrates with a smaller native range, because the former have a higher probability of being transported and released than the latter (Cassey et al. 2004; Jeschke and Strayer 2006; Blackburn and Jeschke 2009; and references therein). In the case of emerging pathogens, we are not aware of similar studies. Based on the outlined stage-dependent differences for invasive species, it would be promising to investigate also which traits in pathogens make them more likely to jump to and replicate within a novel host (Woolhouse et al. 2005), and to establish themselves and spread following a species jump. Similarly for GMOs, it would be fruitful to focus on each stage when investigating traits that increase the likelihood of high impacts.
The probability that an organism will successfully transition from one stage to the next—its transition probability—has received considerable attention in invasion ecology. Most notably, the tens rule has proposed that transition probabilities are around 10%, that is of 100 species released into a non-native range, about 10 species will establish themselves, and 1 species will spread (Williamson 1993). These suggested transition probabilities have also been used as rough estimates of how many GMOs released into the wild will become established (Regal 1993; Williamson 1993). However, subsequent research about invasive species has suggested that animals, particularly vertebrates, have generally higher transition probabilities than plants (Jeschke and Strayer 2005; Jeschke et al. 2012) and that taxonomic differences in transition probabilities also exist within vertebrates, as mammals appear to have higher transition probabilities than birds (Jeschke 2008). Such differences should be investigated for other novel organisms as well, as risk assessment needs to be adjusted to taxonomic differences.
Especially for emerging pathogens, general differences in transition probabilities between the different stages might be expected. For example, one might expect pathogens with hosts that are ecological generalists or human commensals to be most likely to jump to novel hosts, including humans, due to higher contact rates with organisms belonging to other species. One might expect pathogens with high rates of mutation or other genetic change, such as viruses or bacteria under selection via antimicrobials, to be most likely to replicate and establish within novel hosts following the species jump. And one might expect pathogens infecting highly generalized and geographically widespread hosts or vectors to be most likely to spread following establishment.
Which Ecosystems Are Especially Susceptible to Novel Organisms?
Less diverse ecosystems may be more susceptible to invaders, and likewise, pathogens may be transmitted more readily in ecological communities with reduced diversity. In invasion ecology, the biotic resistance hypothesis, also known as diversity-invasibility hypothesis, posits that diverse ecosystems and communities (e.g., measured as species richness) are more resistant against invaders than less diverse ecosystems and communities (Elton 1958; Fridley et al. 2007; Jeschke and Genovesi 2011; Jeschke et al. 2012). In disease ecology, a similar idea has been formulated: more diverse communities often have lower rates of pathogen transmission than less diverse communities (Ostfeld and Keesing 2000; Keesing et al. 2010); genetically diverse populations have been shown in some cases to be more resistant to pathogens than less diverse populations (Lively 2010 and references therein); and individuals with a diverse immune system (e.g., MHC diversity) are more resistant to pathogens than other individuals (Sommer 2005). In invasion ecology, resistance has not yet been investigated at these different ecological scales. We suggest that future studies investigate the relationship between the genetic diversity of populations within a community and genotypic (genomic) diversity of individuals within populations on the one hand, and their resistance to biotic invasions on the other. This could be very informative, as studies varying in spatial scale have found different levels of support for biotic resistance against biological invasions (Fridley et al. 2007).
Aside from biodiversity, many other characteristics of ecosystems could potentially influence their susceptibility to novel organisms. When investigating such characteristics, it is again important to separate different stages. An illustrative example is the case of HIV. When the ailment later termed “acquired immunodeficiency syndrome” (AIDS) was first described in the 1980s, it was assumed that the locations and human populations where it was initially reported, i.e., urban communities of mostly gay males and intravenous drug users in large American and European cities, somehow facilitated the emergence of this new disease. Consequently, risk factors such as urban living, homosexuality, sexual promiscuity, and drug use were identified in early studies as leading to emergence (Fauci 2003). In fact, however, evidence now indicates that the initial species jump of what was probably a simian immunodeficiency virus from apes to humans occurred decades earlier in central Africa, and probably not in homosexual populations (Fauci 2003). Scientists had mistakenly treated what we now know was the post-establishment spread of a new pathogen as one of the initial stages of emergence (species jump followed by replication). It is now evident that the factors eliciting the initial emergence had nothing to do with urbanization or sexual promiscuity. Only subsequent spread is facilitated by unprotected sex between infected individuals and many uninfected ones of either sex. This can occur in rural as well as urban environments. Currently, incidence of HIV/AIDS is vastly higher in sub-Saharan Africa and south Asia than in European or North American cities (Illife 2006).
A similar phenomenon has been suggested by P. Genovesi (pers. comm.) for invasive species, namely that patterns of apparent ecosystem susceptibility to invasive species may change with time: shortly after the transport to and release into a non-native environment, non-native species will be found mainly where they were released. Indeed, currently observed numbers of invasive species in different regions are mainly driven by what has been termed community-level propagule pressure, that is the number of different species transported to and released in non-native environments (Jeschke and Genovesi 2011 and references therein). But similarly to HIV/AIDS and other pathogens, it might be true for invasive species that currently observed patterns will change with time when invasive species have dispersed to locations where they find the best conditions for post-establishment spread.
Climate Change, Biodiversity, and Novel Organisms
Novel organisms are both drivers and consequences of global environmental change and are highly connected with current environmental problems. Climate change affects the probabilities that novel organisms establish, spread, and have high environmental impacts (Fig. 1). Some invasive species benefit from climate change, e.g., the Asian tiger mosquito (Stegomyia albopicta syn. Aedes albopictus) and the waterflea Daphnia lumholtzi (Hellmann et al. 2008; Walther et al. 2009; Engel et al. 2011). Such organisms are both invasive and range-expanding. It is obvious that many other range-expanding species benefit from climate change as well. Further examples for novel organisms benefiting from climate change are emerging pathogens, e.g., avian malaria (Lapointe et al. 2012). Climate change also directly affects biodiversity, for example when environmental conditions change too quickly for species to be able to adapt to them or migrate to areas with more suitable conditions (if such areas still exist; e.g., Bellard et al. 2012).
As pointed out in the previous section (Which ecosystems are especially susceptible to novel organisms?), biodiversity can affect novel organisms. The reverse is true as well: novel organisms can affect biodiversity (Fig. 1). For instance, invasive species are known to be able to reduce species richness (e.g., rats, cats, and other predators have eliminated endemic island species, especially birds; Clavero and García-Berthou 2005; Loehle and Eschenbach 2012). The same is true for emerging pathogens: a current example is chytridiomycosis which is threatening amphibians worldwide (Pounds et al. 2006). Compared to biological invasions and emerging pathogens, effects of GMOs and synthetic organisms on biodiversity have been much less thoroughly investigated (Ricciardi et al. 2011). Although such studies do exist (e.g., Chambers et al. 2010), there are too few to allow for general conclusions. Also, not many GMOs have been released into the wild thus far, especially not animals. Such a lack of knowledge is critical, as policy makers need guidance for making decisions whether or not a given GMO should be approved for market release. Here again, comparing GMOs with other types of novel organisms can be helpful.
Conclusions
Besides promoting the fruitful exchange of ideas and approaches among researchers working on different types of novel organisms, the framework outlined in Fig. 1 also eases communication among other natural and social scientists as well as managers and policy makers when discussing novel organisms, for instance when possible management actions are devised. Such discussions are often complicated by different perspectives and terminologies, and a common framework allows usage of a common language. In this way, we hope that both researchers and practitioners will benefit from the framework proposed here. Furthermore, by explicating similarities among types of novel organisms, our framework suggests ways to integrate policy and management of novel organisms. For example, given that invasive species, GMOs, synthetic organisms, and emerging pathogens are all potentially transported worldwide, the framework suggests that border controls and similar measures against unrecorded transport and release of novel organisms should be discussed and implemented jointly for all types of novel organisms rather than separately for each type. Europe has a system in place that deals with applications for market approval of GMOs (e.g., EC 2001; EFSA 2010), but it has no system that would effectively avoid undeclared transport and release of GMOs into Europe. We suggest the development of a system that is effective against undeclared transport and release of GMOs as well as other types of novel organisms. Similarly for the later stages, establishment and spread (Fig. 1), our framework suggests that a joint policy and management for all types of novel organisms will be more cost-effective than if they are done separately for each type. Thus to avoid negative impacts of novel organisms, integration of research and policy for all types of novel organisms seems most promising. The framework presented here may contribute to such an integration and may generally serve as a useful tool for effective biosafety and biosecurity.
Acknowledgments
JMJ thanks Wolf-Christian Saul and Mark van Kleunen for discussions about novel organisms, and Sara Hammerstein and Justyna Wolinska for discussions about pathogens. He acknowledges financial support from the Deutsche Forschungsgemeinschaft (DFG; JE 288/4-1). FK and RSO thank the National Science Foundation’s Ecology and Evolution of Infectious Diseases program and the United States Environmental Protection Agency for support. Paul Ewald, Brendon Larson, Gary Lovett, Wolf-Christian Saul, and anonymous reviewers provided helpful comments on the manuscript.
Biographies
Jonathan M. Jeschke
is Principal Investigator at Technische Universität München, Germany, and Visiting Scientist at the Cary Institute of Ecosystem Studies, Millbrook, New York, USA. He held research positions at Ludwig-Maximilians-Universität München, Germany; Cary Institute of Ecosystem Studies, Millbrook, New York, USA; and University of Helsinki, Finland. His research interests focus on biological invasions, other novel organisms, biodiversity, predator–prey interactions, and other basic and applied ecological topics.
Felicia Keesing
is David and Rosalie Rose Distinguished Professor in Science, Mathematics, and Computing and Professor of Biology at Bard College, New York, USA. Her research interests include ecology of African savannas, ecology of infectious disease, and pedagogical issues in undergraduate biology education.
Richard S. Ostfeld
is Senior Scientist at the Cary Institute of Ecosystem Studies, Millbrook, New York, USA. His early training was in mammalian ecology and behavioral ecology at the University of California, Berkeley and Boston University. Later his focus shifted to community ecology and disease ecology. He is currently conducting research on the effects of biodiversity change, land use change, and climate change on various vector-borne, zoonotic diseases.
References
- Bellard C, Bertelsmeier C, Leadley P, Thuiller W, Courchamp F. Impacts of climate change on the future of biodiversity. Ecology Letters. 2012;15:365–377. doi: 10.1111/j.1461-0248.2011.01736.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blackburn TM, Jeschke JM. Invasion success and threat status: Two sides of a different coin? Ecography. 2009;32:83–88. doi: 10.1111/j.1600-0587.2008.05661.x. [DOI] [Google Scholar]
- Blackburn TM, Pyšek P, Bacher S, Carlton JT, Duncan RP, Jarošík V, Wilson JRU, Richardson DM. A proposed unified framework for biological invasions. Trends in Ecology & Evolution. 2011;26:333–339. doi: 10.1016/j.tree.2011.03.023. [DOI] [PubMed] [Google Scholar]
- Cassey P, Blackburn TM, Jones KE, Lockwood JL. Mistakes in the analysis of exotic species establishment: Source pool designation and correlates of introduction success among parrots (Aves: Psittaciformes) of the world. Journal of Biogeography. 2004;31:277–284. doi: 10.1046/j.0305-0270.2003.00979.x. [DOI] [Google Scholar]
- Chambers CP, Whiles MR, Rosi-Marshall EJ, Tank JL, Royer TV, Griffiths NA, Evans-White MA, Stojak AR. Responses of stream macroinvertebrates to Bt maize leaf detritus. Ecological Applications. 2010;20:1949–1960. doi: 10.1890/09-0598.1. [DOI] [PubMed] [Google Scholar]
- Clavero M, García-Berthou E. Invasive species are a leading cause of animal extinctions. Trends in Ecology & Evolution. 2005;20:110. doi: 10.1016/j.tree.2005.01.003. [DOI] [PubMed] [Google Scholar]
- Clout MN, Williams PA, editors. Invasive species management: A handbook of principles and techniques. Oxford: Oxford University Press; 2009. [Google Scholar]
- Deplazes A, Huppenbauer M. Synthetic organisms and living machines. Systems and Synthetic Biology. 2009;3:55–63. doi: 10.1007/s11693-009-9029-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- EC. 2001. Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the deliberate release into the environment of genetically modified organisms and repealing Council Directive 90/220/EEC. Official Journal of the European Communities L106: 1–39.
- EFSA. 2010. Guidance on the environmental risk assessment of genetically modified plants. EFSA Journal 8: 1879 (111 pp.).
- Elton CS. The ecology of invasions by animals and plants. London: Methuen; 1958. [Google Scholar]
- Engel, K., R. Tollrian, and J.M. Jeschke. 2011. Integrating biological invasions, climate change and phenotypic plasticity. Communicative & Integrative Biology 4: 247–250. [DOI] [PMC free article] [PubMed]
- Fauci AS. HIV and AIDS: 20 years of science. Nature Medicine. 2003;9:839–843. doi: 10.1038/nm0703-839. [DOI] [PubMed] [Google Scholar]
- Fridley JD, Stachowicz JJ, Naeem S, Sax DF, Seabloom EW, Smith MD, Stohlgren TJ, Tilman D, et al. The invasion paradox: Reconciling pattern and process in species invasions. Ecology. 2007;88:3–17. doi: 10.1890/0012-9658(2007)88[3:TIPRPA]2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Hatcher M, Dick JTA, Dunn AM. Disease emergence and invasions. Functional Ecology. 2012;26:1275–1287. doi: 10.1111/j.1365-2435.2012.02031.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hellmann JJ, Byers JE, Bierwagen BG, Dukes JS. Five potential consequences of climate change for invasive species. Conservation Biology. 2008;22:534–543. doi: 10.1111/j.1523-1739.2008.00951.x. [DOI] [PubMed] [Google Scholar]
- Hobbs RJ, Higgs E, Harris JA. Novel ecosystems: Implications for conservation and restoration. Trends in Ecology & Evolution. 2009;24:599–605. doi: 10.1016/j.tree.2009.05.012. [DOI] [PubMed] [Google Scholar]
- Hudson PJ, Perkins SE, Cattadori IM. The emergence of wildlife disease and the application of ecology. In: Ostfeld RS, Keesing F, Eviner VT, editors. Infectious disease ecology: Effects of ecosystems on disease and of disease on ecosystems. Princeton: Princeton University Press; 2008. pp. 347–367. [Google Scholar]
- Illife J. The African AIDS epidemic: A history. Athens: Ohio University Press; 2006. [Google Scholar]
- Jeschke JM. Across islands and continents, mammals are more successful invaders than birds. Diversity and Distributions. 2008;14:913–916. doi: 10.1111/j.1472-4642.2008.00488.x. [DOI] [Google Scholar]
- Jeschke JM, Genovesi P. Do biodiversity and human impact influence the introduction or establishment of alien mammals? Oikos. 2011;120:57–64. doi: 10.1111/j.1600-0706.2010.18621.x. [DOI] [Google Scholar]
- Jeschke JM, Strayer DL. Invasion success of vertebrates in Europe and North America. Proceedings of the National Academy of Sciences of the United States of America. 2005;102:7198–7202. doi: 10.1073/pnas.0501271102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeschke JM, Strayer DL. Determinants of vertebrate invasion success in Europe and North America. Global Change Biology. 2006;12:1608–1619. doi: 10.1111/j.1365-2486.2006.01213.x. [DOI] [Google Scholar]
- Jeschke JM, Gómez Aparicio L, Haider S, Heger T, Lortie CJ, Pyšek P, Strayer DL. Support for major hypotheses in invasion biology is uneven and declining. NeoBiota. 2012;14:1–20. doi: 10.3897/neobiota.14.3435. [DOI] [Google Scholar]
- Jones KE, Patel NG, Levy MA, Storeygard A, Balk D, Gittleman JL, Daszak P. Global trends in emerging infectious diseases. Nature. 2008;451:990–993. doi: 10.1038/nature06536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keesing F, Belden LK, Daszak P, Dobson A, Harvell CD, Holt RD, Hudson P, Jolles A, et al. Impacts of biodiversity on the emergence and transmission of infectious diseases. Nature. 2010;468:647–652. doi: 10.1038/nature09575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kettunen, M., P. Genovesi, S. Gollasch, S. Pagad, U. Starfinger, P. ten Brink, and C. Shine. 2008. Technical support to EU strategy on invasive species (IAS)—Assessment of the impacts of IAS in Europe and the EU (final module report for the European Commission). Institute for European Environmental Policy (IEEP), Brussels, Belgium (44 pp. + Annexes).
- Kowarik I. Biologische Invasionen: Neophyten und Neozoen in Mitteleuropa. 2. Stuttgart: Ulmer; 2010. [Google Scholar]
- Kueffer, C., A. Hilbeck, P. Edwards, A. Fischlin, B. Wynne, and J. Scott. 2011. Understanding and managing ecological novelty—Towards an integrative framework of the socio-ecological risks of novel organisms. Monte Verita Conference, 4–9 September 2011. Retrieved 1 February, 2013, from http://www.en2011.ethz.ch.
- LaDeau SL, Marra PP, Kilpatrick AM, Calder CA. West Nile virus revisited: Consequences for North American ecology. BioScience. 2008;58:937–946. doi: 10.1641/B581007. [DOI] [Google Scholar]
- Lapointe DA, Atkinson CT, Samuel MD. Ecology and conservation biology of avian malaria. The Year in Ecology and Conservation Biology 2012. Annals of the New York Academy of Sciences. 2012;1249:211–226. doi: 10.1111/j.1749-6632.2011.06431.x. [DOI] [PubMed] [Google Scholar]
- Lively CM. The effect of host genetic diversity on disease spread. American Naturalist. 2010;175:E149–E152. doi: 10.1086/652430. [DOI] [PubMed] [Google Scholar]
- Lloyd-Smith JO, George D, Pepin KM, Pitzer VE, Pulliam JRC, Dobson AP, Hudson PJ, Grenfell BT. Epidemic dynamics at the human–animal interface. Science. 2009;326:1362–1367. doi: 10.1126/science.1177345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lockwood JL, Hoopes MF, Marchetti MP. Invasion ecology. Malden: Blackwell; 2007. [DOI] [PubMed] [Google Scholar]
- Loehle C, Eschenbach W. Historical bird and terrestrial mammal extinction rates and causes. Diversity and Distributions. 2012;18:84–91. doi: 10.1111/j.1472-4642.2011.00856.x. [DOI] [Google Scholar]
- Ostfeld RS, Keesing F. The function of biodiversity in the ecology of vector-borne zoonotic diseases. Canadian Journal of Zoology. 2000;78:2061–2078. doi: 10.1139/z00-172. [DOI] [Google Scholar]
- Perkins SE, Altizer S, Bjornstad O, Burdon JJ, Clay K, Gómez-Aparicio L, Jeschke JM, Johnson PTJ, et al. Invasion biology and parasitic infections. In: Ostfeld RS, Keesing F, Eviner VT, et al., editors. Infectious disease ecology: Effects of ecosystems on disease and of disease on ecosystems. Princeton: Princeton University Press; 2008. pp. 179–204. [Google Scholar]
- Pounds JA, Bustamante MR, Coloma LA, Consuegra JA, Fogden MPL, Foster PN, La Marca E, Masters KL, et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature. 2006;439:161–167. doi: 10.1038/nature04246. [DOI] [PubMed] [Google Scholar]
- Preston CJ. Synthetic biology: Drawing a line in Darwin’s sand. Environmental Values. 2008;17:23–39. doi: 10.3197/096327108X271932. [DOI] [Google Scholar]
- Regal PJ. Models of genetically engineered organisms and their ecological impact. In: Mooney HA, Drake JA, editors. Ecology of biological invasions of North America and Hawaii. New York: Springer; 1986. pp. 111–129. [Google Scholar]
- Regal PJ. The true meaning of “exotic species” as a model for genetic engineered organisms. Experientia. 1993;49:225–234. doi: 10.1007/BF01923530. [DOI] [PubMed] [Google Scholar]
- Reichman JR, Watrud LS, Lee EH, Burdick CA, Bollman MA, Storm MJ, King GA, Mallory-Smith C. Establishment of transgenic herbicide-resistant creeping bentgrass (Agrostis stolonifera L.) in nonagronomic habitats. Molecular Ecology. 2006;15:4243–4255. doi: 10.1111/j.1365-294X.2006.03072.x. [DOI] [PubMed] [Google Scholar]
- Ricciardi A, Palmer ME, Yan ND. Should biological invasions be managed as natural disasters? BioScience. 2011;61:312–317. doi: 10.1525/bio.2011.61.4.11. [DOI] [Google Scholar]
- Secretariat of the Convention on Biological Diversity. 2000. Cartagena Protocol on Biosafety to the Convention on Biological Diversity: Text and annexes. Secretariat of the Convention on Biological Diversity, Montreal, Canada, 30 pp.
- Sharples, F.E. 1982. Spread of organisms with novel genotypes: Thoughts from an ecological perspective. ORNL/TM-8473, Oak Ridge National Laboratory Environmental Sciences Division Publication No. 2040, 50 pp.
- Sommer S. The importance of immune gene variability (MHC) in evolutionary ecology and conservation. Frontiers in Zoology. 2005;2:16. doi: 10.1186/1742-9994-2-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taylor LH, Latham SM, Woolhouse MEJ. Risk factors for human disease emergence. Philosophical Transactions of the Royal Society of London, Series B. 2001;356:983–989. doi: 10.1098/rstb.2001.0888. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vilà M, Basnou C, Pyšek P, Josefsson M, Genovesi P, Gollasch S, Nentwig W, Olenin S, et al. How well do we understand the impacts of alien species on ecosystem services? A pan-European, cross-taxa assessment. Frontiers in Ecology and the Environment. 2010;8:135–144. doi: 10.1890/080083. [DOI] [Google Scholar]
- Walther G-R, Roques A, Hulme PE, Sykes MT, Pyšek P, Kühn I, Zobel M, Bacher S, et al. Alien species in a warmer world: Risks and opportunities. Trends in Ecology & Evolution. 2009;24:686–693. doi: 10.1016/j.tree.2009.06.008. [DOI] [PubMed] [Google Scholar]
- Warwick SI, Légère A, Simard M-J, James T. Do escaped transgenes persist in nature? The case of an herbicide resistance transgene in a weedy Brassica rapa population. Molecular Ecology. 2008;17:1387–1395. doi: 10.1111/j.1365-294X.2007.03567.x. [DOI] [PubMed] [Google Scholar]
- Williamson M. Invaders, weeds and the risk from genetically modified organisms. Experientia. 1993;49:219–224. doi: 10.1007/BF01923529. [DOI] [Google Scholar]
- Wolfe ND, Panosian Dunavan C, Diamond J. Origins of major human infectious diseases. Nature. 2007;447:279–283. doi: 10.1038/nature05775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woolhouse MEJ, Haydon DT, Antia R. Emerging pathogens: The epidemiology and evolution of species jumps. Trends in Ecology & Evolution. 2005;20:238–244. doi: 10.1016/j.tree.2005.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]