

Abstract
Turtles are one of the least explored clades regarding endocranial anatomy with few available descriptions of the brain and inner ear of extant representatives. In addition, the paleoneurology of extinct turtles is poorly known and based on only a few natural cranial endocasts. The main goal of this study is to provide for the first time a detailed description of the neuroanatomy of an extinct turtle, the Late Jurassic Plesiochelysetalloni , including internal carotid circulation, cranial endocast and inner ear, based on the first digital 3D reconstruction using micro CT scans. The general shape of the cranial endocast of P. etalloni is tubular, with poorly marked cephalic and pontine flexures. Anteriorly, the olfactory bulbs are clearly differentiated suggesting larger bulbs than in any other described extinct or extant turtle, and indicating a higher capacity of olfaction in this taxon. The morphology of the inner ear of P . etalloni is comparable to that of extant turtles and resembles those of slow-moving terrestrial vertebrates, with markedly low, short and robust semicircular canals, and a reduced lagena. In P . etalloni the arterial pattern is similar to that found in extant cryptodires, where all the internal carotid branches are protected by bone. As the knowledge of paleoneurology in turtles is scarce and the application of modern techniques such as 3D reconstructions based on CT scans is almost unexplored in this clade, we hope this paper will trigger similar investigations of this type in other turtle taxa.
Introduction
While the study of endocranial casts has a long history in paleontology (see 1 and references herein), it was only in the past 20 years that the development of non-invasive technologies such as computed tomography made it possible to reconstruct endocranial structures in extinct reptile forms on a broader scale (e.g. [2–11]). Most of these studies, which focused largely on dinosaurs, included not only brain morphology, but also inner ear structure, and cranial vasculature, allowing for detailed comparisons with extant representatives (e.g. [8,12–14]). Despite these methodological advances, however, knowledge on the morphology and evolution of the turtle brain still remains poor.
The turtle brain is housed in a tubular braincase, composed of the prootic, opisthotic, epipterygoid (present in basal and pancryptodiran turtles), basisphenoid, basioccipital, and exoccipital, and roofed by the supraoccipital, parietal and frontal. As in other reptiles, the brain is smaller than the cranial cavity and therefore the endocast only vaguely reflects its parts [15]. In extant turtles, the cranial endocast shows poorly marked cephalic and pontine flexures, and an extremely short olfactory tract (e.g. [16–19]). While the inner ear morphology has been described for a few extant turtles such as Chelydraserpentina , Chrysemys scripta , Carettochelysinsculpta and Chelonoidis chilensis [20,21], there is no description of the inner ear of any extinct form. However, the general morphology of the turtle inner ear consists of a labyrinth with short and dorsoventrally low semicircular canals and a short and conical lagena.
Natural or artificial cranial endocasts of extinct turtles are known for only a few taxa such as the chelonioid Corsochelyshaliniches from the Late Cretaceous of Alabama [16], the pelomedusoids Bothremyscooki and B . barberi from the Late Cretaceous of North America [17,18], and the Late Cretaceous baenid Plesiobaenaantiqua from Canada [19]. Studies of the endocasts of extant turtles are also scarce, including only the chelonioids Chelonia mydas [16,22], Caretta caretta [16], Dermochelys coriaceae , Eretmochelysimbricata , and Lepidochelys kempii [23], the chelydrid Macrochelys temminckii [24], and the pelomedusoid Erymnochelys madagascariensis [17].
The purpose of the present study is to provide a detailed description of the endocranial cavity and inner ear of the extinct turtle Plesiochelysetalloni using micro-computed tomography (CT), to compare it with other available information on endocranial casts of turtles. Plesiochelysetalloni is a pancryptodiran turtle [25] from the Late Jurassic (Upper Kimmeridgian–Lower Tithonian) lithographic limestones of Western Europe. Previous authors suggested plesiochelyids were living either in freshwater or coastal marine habitats [26–28], and oxygen isotope composition of plesiochelyid turtles recently supported the latter hypothesis given the taxon’s marine isotopic signature [29]. The new data from this study will serve as a basis for a better understanding of turtle brain anatomy and evolution.
Materials and Methods
Plesiochelysetalloni (MH 435) is represented by an almost complete skull (missing the occipital region) with the lower jaw [30,31] (Figure 1). This specimen was found in Glovelier (Canton de Berne) in Switzerland in 1950 and is housed in the collections of the Naturhistorisches Museum, in Basel, Switzerland. The species was described in detail by Gaffney [30,31] who based his description on the same specimen. Later, Sterli et al. [32] re-studied the basicranial region exploring the parabasisphenoid complex and the carotid circulation. To acquire the CT scans used in the present study, a loan of the specimen MH 435 was obtained from the Naturhistorisches Museum (Switzerland) to one of the authors (J. Müller) in March 2008. All necessary permits were obtained, and the study complied with all relevant regulations. For this study a high-resolution X-ray micro CT scan of the skull of MH 435 was performed at the Helmholtz-Zentrum für Materalien und Energie in Berlin, Germany, with 1000 projections over 360°, and exposure time of 0.6 s, a voltage of 100.0 kV, and current of 100 µA. Virtual three-dimensional inner ear and cranial endocasts were generated at the University of Alberta Paleovertebrate Laboratory (Alberta, Canada) using the software Materialise Mimics (14.0); and the resulting 3D models were then imported into the software Geomagic (10.0) to create 3D PDFs (see supplementary data). The final illustrations were generated using the software Illustrator (CS5). The endocast of Plesiochelysetalloni was compared with published turtle endocasts (e.g., [16,17,19]) and with an unpublished latex endocast of the terrestrial turtle Chelonoidis chilensis (Testudinidae, MPEF-AC 411). The CT data are archived at the Museo Carmen Funes (Plaza Huincul, Neuquén, Argentina) and at the Museum für Naturkunde Leibniz-Institut für Evolutions-und Biodiversitätsforschung an der Humboldt-Universität zu Berlin (Berlin, Germany).
Figure 1. Volume-rendered CT-based reconstruction of the skull of the extinct turtle Plesiochelysetalloni (MH 435).

In the images of the left side the bone is rendered opaque, whereas in the right side the bone is rendered semitransparent to show the endocranial cavity. The bone is shown in green, the cranial endocast and inner ear in light blue, the internal carotid arteries in pink, and the columella in fuchsia. Skull in left lateral (A) dorsal (B) and ventral (C) views. Abbreviations: col, columella auris; endo, cranial endocast; fpcci, foramen posterius canalis caroticus internus; fst, foramen staedio-temporale. Scale bar equals 10 mm.
Description
Cranial endocast
As in all extant and most extinct turtles, the anterior walls of the braincase such as ethmoidal elements, orbitosphenoids and laterosphenoids are not ossified, preventing the reconstruction of cranial nerves II to IV (cranial nerve I leaves an impression on the ventral aspect of the frontals). Endocranially, there is no floccular recess and the vestibular eminence is not ossified; consequently, the inner ear cavity is confluent with the endocranial cavity medially (hiatus acusticus, sensu [33]).
The cranial endocast of P . etalloni is almost complete, missing the posterior section of the medulla oblongata, together with the roots for cranial nerves IX to XII (Figure 1B, C). The shape of the endocast is generally similar to those of other extinct and extant turtles (e.g., [16,17,19]), being tubular and with poorly marked cephalic and pontine flexures. The preserved length of the endocast is 58.6 mm. The total estimated length would be 60.1 mm, based on the length of the endocasts of extant turtles (personal observations on an artificial endocast of Chelonoidis chilensis ). The hind-brain, midbrain and forebrain are more or less horizontally aligned as in the extinct Corsochelyshaliniches ( [15]: Figure 6B) and the extant Dermochelys coriaceae ( [23]: Figure 192); meaning the dorsal border of the medulla oblongata is in the same horizontal plane with most of the olfactory tract and nasal cavity when viewed from lateral (Figure 2). This differs from Bothremys ( [15]: Figure 7) and the extant sea turtles Chelonia mydas , Lepidochelys kempii , Caretta caretta , and Eretmochelysimbricata ( [23]: Figure 192) where the medulla is below the level of the olfactory tract.
Figure 2. Surface-rendered CT-based reconstructions of the cranial endocast and endosseous labyrinth of Plesiochelysetalloni (MH 435).

In dorsal (A–B), lateral (C–D), and ventral (E–F) views. The cavum acustico-jugulare is rendered opaque in (A, C, and E); to show the location of the columella it is rendered semitransparent in (B, F), and is absent in (D). The images show the cranial endocast in light blue, the inner ear in light violet, the cavum acustico-jugulare in purple, the columella in fuchsia, and internal carotids in pink. Abbreviations: asc, anterior semicircular canal; ca, cerebral artery; caj, cavum acustico-jugulare; ccv, canalis cavernosus; cer, cerebral hemisphere; col, columella auris; col. f, columellar foot; cr, cartilaginous ridge (= cartilaginous rider of [17,19]); cst, canalis stapedio-temporalis; fst, foramen stapedio-temporale; ic, internal carotid; ica, incisura columella auris; lab, endosseous labyrinth; lsc, lateral semicircular canal; med, medulla oblongata; nc, nasal cavity; ob, olfactory bulb; ot, olfactory tract; pa, palatine artery; pit, pituitary fossa; psc, posterior semicircular canal; sa, stapedial artery; V–VIII, cranial nerves. Scale bar equals 10 mm.
Forebrain
The lateral and ventral walls of the olfactory tract cavity are not ossified. The olfactory tract is short, almost nonexistent, and 3.4 mm in width (maximum width). The impression of the olfactory bulb on the frontal, situated anteriorly to the tract, is shallow but clear enough to delimit its morphology. The bulbs are oval-shaped and slightly divergent from the midline (Figure 2A–D). The olfactory bulbs leave no impressions on the ventral aspect of the frontals in extant turtles (e.g. Chelonioidis chilensis personal observation) and for this reason the bulbs are not discernible in cranial endocasts. In the endocast of P . etalloni , the presence of olfactory bulbs indicates that those are relatively larger than in other studied turtles, suggesting that this taxon had a better olfaction. Cranial nerve I projects anteriorly in front of the olfactory bulbs in a trajectory that is slightly convex, and ends in the large nasal cavities. The nasal cavities are large and medially connected.
The cerebral hemispheres are easily discernible (Figure 2A). The maximum width of the endocast (13.3 mm) is at the level of the cerebral hemispheres, which are less laterally expanded than in Bothremys and Corsochelyshaliniches ( [15]: Figures 6A, 7B). On the ventral aspect of the endocast, the pituitary cast is small and dorsoventrally depressed (Figure 2E, F). The cerebral carotid arteries enter it posteriorly, through separated foramina (see below).
Midbrain
The optic lobes are not discernible in the endocast. The roots for cranial nerves II–IV cannot be reconstructed since the bones enclosing their foramina (orbitosphenoids) are not ossified in turtles [33].
Hindbrain
Dorsal to the cerebellum, there is a large triangular area marked by sharp ridges that are thickened laterally and connecting to the root of the trigeminal nerve, which are here interpreted as the cartilaginous ridge (= cartilaginous “rider” [17]) (Figure 2A–D). In other reptiles, this dorsal protuberance is related to the dorsal sagittal sinus, the dorsal head veins and the rostral middle cerebral vein (see 8. In P. etalloni, the cartilaginous ridge has marked lateral expansions that reach ventrally the root of CN V. The presence of this lateral ridge suggests that the dorsal head vein and the rostral middle cerebral vein drain through the same foramen as CN V. The dorsal longitudinal sinus is not well developed, although thick enough to hide the shape of the cerebral hemispheres. In the extant Chelonia mydas , Hopson [15] interpreted this area in the endocast as the cartilaginous anterior end of the supraoccipital (=cartilaginous “rider” of [17]).
Cranial nerve V leaves the endocranial cavity through a single large foramen (=foramen nervi trigemini [33]), enclosed mainly by the parietal with small contributions of the epipterygoid and the pterygoid [30,31]. The trigeminal root is large and posteroventrally in connection with the canalis cavernosus, which, at this level, is ventromedially projected (Figures 2A–C, 3).
Figure 3. Detail of posterior region of the CT-based reconstructions of the cranial endocast of Plesiochelysetalloni (MH 435) in left anteroventral view.

The figure shows the cranial endocast in light blue, the columella in fuchsia, the internal carotid arteries in pink, the CN VI in yellow, and the cavum acustico-jugulare and the canalis stapedio-temporalis (rendered semitransparent to show the Columella ) in purple. Abbreviations: ca, cerebral artery; ccv, canalis cavernosus; col, columella auris; ic, internal carotid; pa, palatine artery; pit, pituitary fossa; sa, stapedial artery; V–VIII, cranial nerves. Scale bar equals 10 mm.
The passage of CN VI has a very small diameter. In the endocast, CN VI exits at the base of the medulla and it passes through the basisphenoid in a dorsally convex trajectory to exit at the base of the processus clinoideus [33] close to the base of the most anteroventral section of the canalis cavernosus at the level of CN V (Figure 3). Both rami of CN VI pass almost parallel into anterior direction. The total length of the nerve is 5.5 mm.
Endocranially, cranial nerves VII and VIII are located posteroventrally to CN V (Figures 2B, 4). As in most turtles [33], they are housed in an oval, single recess (= fossa acustico-facialis [33]), and as revealed by the endocast, both nerves seem to share a single space, as product of the filling of the recess in the cast (Figure 3). CN VII leaves the recess anterior to CN VIII and has a larger diameter and a longer passage that passes laterally to enter the canalis cavernosus (Figure 2E). On the contrary, CN VIII has a smaller diameter and a shorter passage to enter the anterior ampulla of the inner ear. In turtles, usually two branches of CN VIII pierce the prootic, and sometimes a third branch enters the inner ear through the cartilaginous hiatus acusticus [33]. In the extant Chelydraserpentina , there are three branches of CN VIII, which are innervating the anterior ampulla, the macula sacculi and the posterior ampulla, respectively ( [34]: Figure 49). In P . etalloni , however, the preservation of a single foramen may be related to the lack of ossification of the vestibular eminence, although there is no evidence of a separated ganglion for other branches.
Figure 4. Detail of posterior region of the CT-based reconstructions of the cranial endocast and inner ear of Plesiochelysetalloni (MH 435) showing cranial nerves VII and VIII.

The image shows the cranial endocast in light blue and the inner ear in light violet. Abbreviations: ccv, canalis cavernosus; lab, endosseous labyrinth; med, medulla oblongata; V–VIII, cranial nerves. Scale bar equals 10 mm.
Only a short part of the medulla oblongata is preserved in P . etalloni , which is horizontally oriented (Figure 2C, D). As in all turtles, CN IX pierces the processus interfenestralis of the opisthotic (Figure 5). Due to the missing posterior part of the skull of P . etalloni , the trajectories of cranial nerves X to XII are not preserved.
Figure 5. Digital reconstruction of the skull of Plesiochelysetalloni (MH 435) in lateroposteroventral view.
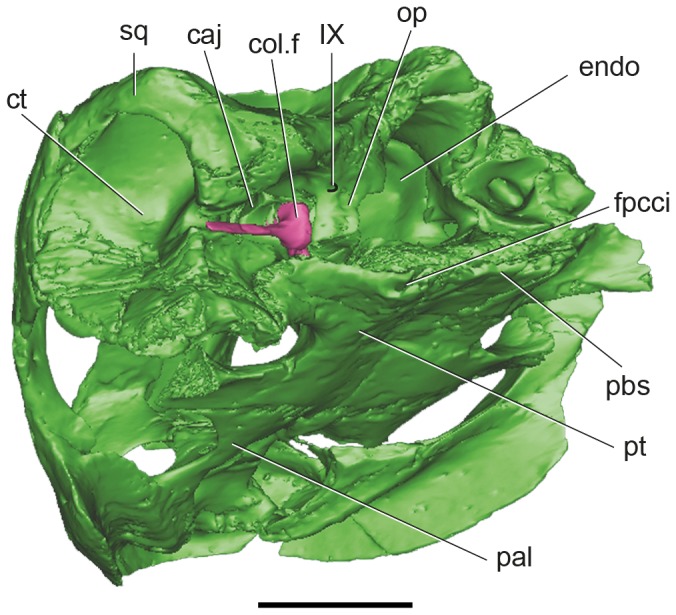
Detail of the posterior part of the middle ear showing the foramen for cranial nerve IX. The image shows the bone in green and the columella in fuchsia. Abbreviations: cavum acustico-jugulare; col. f, columellar foot; ct, cavum tympani; endo, endocranial cavity; fpcci, foramen posterius canalis caroticus internus; op, opisthotic; pal, palatine; pbs, parabasisphenoid complex; pt, pterygoid; sq, squamosal; IX, cranial nerve. Scale bar equals 10 mm.
Inner ear
The complete left inner ear of P . etalloni was digitally reconstructed (Figure 6). The overall inner ear morphology corresponds to that described in extant turtles such as Chelydraserpentina ( [21]: Figure 1) and Trachemys scripta ( [35]: Figure 23-3), in which the lagena is globose and extremely short, and the semicircular canals are short, subequal in size and oval-shaped.
Figure 6. Digital reconstruction of the left inner ear of Plesiochelys etalloni (MH 435).

In lateral (A, E), dorsal (B, F), anterior (C) and posterior (D) views. The image shows the inner ear in light violet and the columella in fuchsia. The columella is shown in anatomical position (E, F) and isolated in posteroventral view (G). Abbreviations: aa, anterior ampulla; asc, anterior semicircular canal; cc, common crus; col, columella auris; col. f, columellar foot; dpl, ductus perilymphaticus; fo, fenestra ovalis; fpl, fenestra perylimphatica; lag, lagena; lsc, lateral semicircular canal; psc, posterior semicircular canal; Scale bar equals 10 mm.
In P . etalloni , the anterior and posterior semicircular canals are markedly depressed dorsoventrally and the internal diameter is eye-shaped (Figure 6A, C, D). The diameter of the tube of the semicircular canals is as follows: 1.1 mm for the anterior semicircular canal (ASC), 1.2 mm for the posterior semicircular canal (PSC) and 1.7 mm for the lateral semicircular canal (LSC) (Figure 6B, F). The ASC is slightly larger than the PSC, whereas the LSC is the most robust and the smallest. The internal diameter of the ASC is 4.1 mm, while the PSC is 3.4 mm, and the LSC is 2.3 mm. The angle between ASC and the PSC is approximately 80° in dorsal view (Figure 6 B, F), which is slightly wider than the angle described for the extant turtle Trachemys scripta [2]. The anterior ampulla (=recessus labyrinthicus prooticus [33]) is well developed and larger than the posterior ampulla (=recessus labyrinthicus opisthoticus [33]). There is a small passage passing from the posterior ampulla to exit through a small foramen on the posterior part of the skull, the fenestra perilymphatica, opening between the cavum labyrinthicum and the recessus scalae tympani of the cavum acustico-jugulare formed by the opisthotic (Figure 6A, D). The periotic sac of the inner ear extends through this fenestra [33].
The fenestra ovalis is large and circular, occupying more than the 50% of the width of the vestibulum (Figure 6A). The diameter of the fenestra ovalis is 2.5 mm. The limit between the cavum labyrinthicum (inner ear) and the cavum acustico−jugulare (middle ear) is marked by the fenestra ovalis [33]. As in Proganochelysquenstedti , Palaeochersistalampayensis , and Australochelys africanus [36–38] and contrary to most extant turtles, the fenestra ovalis is completely surrounded by bone, anteriorly by the prootic and posteriorly by the opisthotic. In turtles, the elements forming the floor of the medial aspect of the inner ear are variable. In pancryptodires the floor of the inner ear is usually formed by the prootic and opisthotic, with occasional contributions from the basisphenoid, basioccipital, or pterygoid, while in many pleurodires, the floor of the cavum labyrinthicum is mainly cartilaginous with osseous contributions of the basisphenoid and quadrate [33]. In P . etalloni the floor of the cavum labyrinthicum is formed by bone. The basisphenoid forms the anteromedial quarter of the flooring of the inner ear, whereas the basioccipital and the pterygoid are closing the cavity posteromedially and ventrally, respectively (Figure 5).
As in other turtles, the lagena in P . etalloni is globose and markedly short (Figure 6A, D). The lagena houses the cochlear tube, which serves for sound perception [9]. Its length is directly related to the length of the sensory epithelium, the distance of which is used to estimate auditory capacity [9]. Recent studies on hearing showed that turtles have the lowest hearing frequency range within extant reptiles [21].
Columella auris
The columella is almost completely preserved and in its natural position (Figure 2 B, D). It is nail-shaped, with a large and circular foot that is concave towards the fenestra ovalis. This foot is 2.5 mm in diameter. In P . etalloni , the shaft of the columella is a rod-like element (cylindrical) and 0.9 mm in diameter. The preserved length of the columella is 9.2 mm, and is only missing the extracolumella, which was probably cartilaginous, as in extant turtles [33].
Canalis stapedio-temporalis and cavum acustico-jugulare
Regarding the middle ear cavity proper, the canalis stapedio-temporalis is posterodorsal and the canalis cavernosus of the cavum acustico-jugulare is lateral and anteroventral. The canalis stapedio-temporalis and the canalis cavernosus are confluent at the level of the inner ear (laterally to it), forming a single cavity located ventrolaterally to the hindbrain, as shown by the 3D reconstruction (Figure 2A–C, E, F).
The posterodorsal branch is named canalis stapedio-temporalis, and is a dorsally projecting passage that ends in the large foramen stapedio-temporalis on the posterior margin of the supratemporal fossa (Figures 1B, 2A, B). The canalis stapedio-temporalis and the foramen stapedio-temporale are formed by the prootic and quadrate. The arteria stapedialis passes through this canal [33], as probably did the hyomandibular branch of the nerve VII.
The cavum acustico-jugulare in turtles, is L-shaped in dorsal view (Figure 2A, B). The cavum acustico-jugulare is located between the cavum tympani (a funnel-shaped cavity formed by the quadrate in turtles) and the cavum labyrinthicum [33]. The elongated anteroventral part of the cavum acustico-jugulare corresponds to the canalis cavernosus (Figure 2A, C, F) and represents the cranio-quadrate space of basal tetrapods [33]. The posteromedial section of the cavum-acustico jugulare is the recessus scalae-tympani [33]. The recessus scalae-tympani is separated from the remaining parts of the cavum acustico-jugulare by the processus interfenestralis of opisthotic. The columella auris is housed by the cavum tympani (laterally) and in part by the cavum acustico-jugulare (Figures 2B, F, 5).
In Plesiochelysetalloni as in other turtles, the canalis cavernosus is the anteroventral branch and the largest (Figure 2A–C). It is arc-shaped and passes anteriorly to reach the level of the trigeminal nerve. The canalis cavernosus in P. etalloni passes then anteroventrally and medially and merges with its counterpart anteriorly to the pituitary fossa on the ventral aspect of the cranial endocast (Figure 2E, F). Cranial nerves V, VII, and the stapedial artery (see below) enter the lateral section of the canalis cavernosus, whereas CN VI and the palatine artery enter the ventral section of the canal (Figure 3), and the vena capitis lateralis passes inside the canal (cranio-quadrate space of other vertebrates sensu [33]).
Carotid circulation
The internal carotid artery enters the skull at the posterior end of the pterygoid [32,39] (Figures 1C, 5). It passes anteriorly for a short length before it bifurcates into two branches (Figure 3). The medial branch continues anteriorly, whereas the lateral (arteria stapedialis) curves towards the canalis cavernosus. The arteria stapedialis enters the ventral aspect of the cavum acustico-jugulare. We assume that it passes posteriorly turning dorsally and entering the aditus canalis stapedio-temporalis, running along the canalis stapedio-temporalis to exit through the stapedio-temporalis foramen (Figures 2E, F, 3). The medial branch of the internal carotid continues anteriorly for a short distance and bifurcates again into lateral and medial branches. The medial branch (arteria cerebralis) enters the pituitary fossa posteriorly (Figures 2E, F, 3). The lateral branch (arteria palatina) passes anterodorsally and enters the canalis cavernosus anteriorly to the pituitary fossa (Figures 2E, F, 3). This arterial pattern, where all the internal carotid branches are enclosed by bone, corresponds to pattern I as recognized by Sterli and de la Fuente [40].
Discussion and Conclusions
The scarce knowledge of endocranial casts in turtles prevents a more detailed comparison of the endocranial cast of P . etalloni with that of other turtles. We hope this work will be the starting point for more descriptions of endocranial reconstructions using extinct and extant turtles.
In general aspects, the shape of the brain of P. etalloni is similar to that of Dermochelys coriaceae . This marine species has the most horizontally developed brains, closely resembling that of P. etalloni. Other marine species such as Caretta caretta , Eretmochelysimbricata , Chelonia mydas and Lepidochelys kempi have brains with well-marked cephalic flexures and nasal cavities that are long and ventrally convex [23] although a cranial endocast would probably not reflect this morphology. As observed in skull dissections on extant taxa [23] and as described for archosaurian reptiles, the brain parts are obscured by the overlying dural venous sinuses and the dura itself [8]. In this sense, the endocranial cavity does not reflect well the morphology of the brain and other soft tissues. For example, the pineal gland is well developed in most turtles, but only in D . coriaceae [23] is it correlated with a dorsal projection of the endocranial cavity (identified as the cartilaginous “rider” by Gaffney and Zangerl [17] (Figure 7A). In P . etalloni the cartilaginous rider is one of the most striking features on the dorsal surface of the endocast. This structure could indicate the location of the pineal organ, but also could be a part of the dorsal longitudinal sinus. Among extinct turtle species, the largest known cartilaginous ridge is observed in Corsochelyshaliniches and Brothremyscooki (Figure 7A), whereas it is less prominent in P . etalloni and Plesiobaenaantiqua .
Figure 7. Brain endocasts and left endosseous inner ears of turtles available in the literature displayed in a simplified cladogram (based on Joyce [25]).

The black circle indicates a synapomorphy based clade definition while the white circle indicates a crown-group definition following Joyce et al. [54]. A, Brain endocasts comparison based on line drawings of extinct and extant turtles in right lateral view represented by (left to right) (1) Bothremys cooki (redrawn from [17]), (2) Podocnemis unifilis (after [33]), Chelidae Emydura macquarii (after [33]), Baenidae Plesiobaena antiqua (redrawn from [19]), Plesiochelys etalloni (this study), (3) Corsochelys haliniches (redrawn from [16]), (4) Chelonia mydas, (5) Eretmochelys imbricata, (6) Caretta caretta, (7) Lepidochelys kempi, (8) Dermochelys coriacea (after [23]), Chelydridae Macrochelys temminckii (after [33]), Emydidae Pseudemys concinna (after [33]), Testudinidae Chelonoidis chilensis (this study), Kinosternidae Sternotherus odoratus (after [33]), and Trionychia Apalone ferox (after [33]). Dashed line indicates the shape of the nasal cavity. Dotted line in Plesiochelys etalloni indicates reconstructed area of the endocast. Dark gray, extinct species. Light gray, extant species. Not to scale. B, Inner ear endocasts were redrawn from the following sources: Plesiochelys etalloni from this study and Chelydridae, Emydidae, Testudinidae, and Trionychia from Thewissen and Nummela [20] (Fig. 15.11 K-N). Chelydridae is represented by Macrochelys temminckii; Emydidae is represented by Trachemys scripta; Testudinidae is represented by Chelonoidis nigra; Trionychia is represented by Carettochelys insculpta. Proganochelys quenstedti, Meiolaniformes, and Plesiochelys etalloni are extinct taxa. Not to scale.
Regarding the olfactory region, in extant turtles the small olfactory bulbs do not leave impressions on the frontals and cannot be clearly identified on the artificial endocasts [41]. However, the olfactory bulbs are clearly differentiated in the endocast of P . etalloni , suggesting larger bulbs than in any other described extinct or extant turtle, and indicating a relatively higher capacity of olfaction in this taxon.
In vertebrates it has been proven that the endosseous labyrinth closely follows the path and shape of the membranous semicircular ducts (see 42 and references cited there). Therefore, interpretations on hearing and balance in extinct taxa based on extant species seem to be adequate at this level. As in other vertebrates, the vestibular system is involved in the coordination of movement, gaze control, and balance, detecting head movement (head rotation) in space and maintaining visual and postural stability. For a given angular acceleration, the maximum achieved bending depends on the sensitivity of the semicircular canal (which in turns depends on the length of the canal, on the area enclosed by the canal in its maximal response plane, and on the cross-section of the semicircular duct and the ampulla) (see 42 and references cited there). In P . etalloni the labyrinth is dorsoventrally depressed and the diameters of the tubes are ASC<PSC<LSC (e.g. the internal diameter of the LSC is one and a half smaller than the ASC). Also, the fenestra ovalis seems to be more dorsally located (closer to the LSC) than in other turtles. The ASC is dorsoventrally depressed and markedly developed anteroposteriorly forming an angled margin with the anterior ampulla. This disposition is similar to the one observed in Carettochelysinsculpta and Chelonoidis chilensis (this study). The anterior ampulla is well developed. It housed the utriculus, being the section of the vestibulum that senses motion in the horizontal plane (e.g., forward, backward, and left–right movements).
The morphology of the inner ear of P . etalloni resembles those of other slow-moving terrestrial vertebrates such as the tuatara and the non-arboreal chamaeleon Brookesia, in which this morphology is optimized to detect weak accelerations, probably important to ensure postural stability [43]. Recent studies showed that in most turtle species, the turtle ear is less sensitive to airborne sound than those of other tympanate tetrapods [44]. However, aquatic turtles have better hearing than terrestrial turtles (i.e. the range of sound perception is higher) since the ear responds to much lower sound energy in water, and therefore being more efficient. Recent studies also showed that the turtle ear is adapted for underwater hearing, explaining both the poor sensitivity to airborne sound and the turtle middle ear structure [21,44]. For the turtle Trachemys scripta , the underwater audiograms had the same shape as audiograms in air, with best frequency of 400–500 Hz [44]. In terms of environmental adaptation the inner ear morphology in turtles is yet unclear. For Lenhardt et al. [45] the auditory process in turtles is even more complex. They suggested that the sound process in turtles involves the traditional air conduction, but also bone conduction. The sound captured by bone conduction happens through the vibration of skull and/or shell bones ( [45]), while air conduction occurs through the tympanum. Manley and Kraus [46] stated that hearing limits are much less specific to systematic groups than previously thought, depending less on phylogenetic position than on species-specific physical (size) and ecological (temperature regime, lifestyle) constraints. As stated by Christensen-Dalsgaard et al. [44] the behavioral significance of hearing in turtles still remains to be discovered, but navigation, prey detection and predator avoidance are probably important in this group, as in many other aquatic vertebrates. With reference to studies suggesting that the inner ear system of aquatic turtles is elongated anteroposteriorly relative to terrestrial turtles [20], our comparison of the endosseous labyrinth of P . etalloni indicates that the inner ear morphology is not useful to identify the taxon’s ancient habitat (Figure 7B). However, the sample of known inner ears of extinct turtles is so small that new descriptions are needed to understand the implications of its morphology (Figure 7B).
Another interesting structure clearly observed in the 3D reconstruction is the carotid circulation pattern. The different patterns of carotid circulation have long been used for turtle systematics (e.g., [25,39,40,47,48]). In recent years many new extinct turtles have been discovered and new studies have been made, highlighting that the complexity of the carotid circulation in turtles is greater than previously thought (e.g., [40,49–53]). The 3D endocranial reconstruction of P . etalloni reinforces the previous descriptions of its carotid circulation [30–32]. In P . etalloni the carotid circulation is similar to that found in extant cryptodires (pattern I of [40]). In these turtles the carotid artery enters the skull through the foramen posterius canalis caroticus internus located in the posterior part of the pterygoid. The entire path of the carotid artery, and its bifurcation into cerebral and palatine branches, is surrounded by bone.
Supporting Information
(PDF)
Acknowledgments
We thank C. Meyer (MH) for allowing the study and scanning of the specimen described here. P.J. Currie and E. Koppelhus (University of Alberta) supported the trip of one of the authors (APC) to Canada, and V. Arbour (U of A) instructed her in the use of the software Mimics. P. Puerta (MEF) made the latex endocast of Chelonoidis chilensis . We also thank G. Sobral (Museum für Naturkunde Berlin) for critically reading an earlier draft of the manuscript and J. Anquetin (Office de la culture, République et Canton Du Jura, Switzerland) and an anonymous reviewer whose comments improved the manuscript. Richard Butler is thanked for his editorial work.
Funding Statement
This study was partially financed by an Ale-Arg fellowship to JS. No additional funding was received for this study. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Buchholtz EA, Seyfarth E-A (1999) The gospel of the fossil brain: Tilly Edinger and the science of paleoneurology. Brain Res Bull 48: 351–361. doi:10.1016/S0361-9230(98)00174-9. PubMed: 10357066. [DOI] [PubMed] [Google Scholar]
- 2. Rogers SW (1998) Exploring dinosaur neuropaleobiology: computed tomography scanning and analysis of an Allosaurus fragilis endocast. Neuron 21: 673–679. doi:10.1016/S0896-6273(00)80585-1. PubMed: 9808455. [DOI] [PubMed] [Google Scholar]
- 3. Brochu CA (2000) A digitally-rendered endocast for Tyrannosaurus rex . J Vert Paleontol 20: 1-6
- 4. Franzosa J, Rowe T (2005) Cranial endocast of the Cretaceous theropod dinosaur Acrocanthosaurus atokensis . J Vert Paleontolol 25: 859–864
- 5. Sanders KR, Smith DK (2005) The endocranium of the theropod dinosaur Ceratosaurus studied with computed tomography. Acta Palaeontol Pol 50: 601–616. [Google Scholar]
- 6. Evans CD, Ridgely R, Witmer LM (2009) Endocranial anatomy of lambeosaurine hadrosaurids (Dinosauria: Ornithischia): a sensorineural perspective of cranial crest function. Anat Rec 292: 1315–1337. doi:10.1002/ar.20984. [DOI] [PubMed] [Google Scholar]
- 7. Sereno PC, Wilson JA, Witmer LM, Whitlock JA, Maga A et al. (2007) Structural extremes in a Cretaceous dinosaur. PLOS ONE 2(11): 21230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Witmer LM, Ridgely RC, Dufeau DL (2008) Using CT to peer into the past: 3D visualization of the brain and ear regions of birds, crocodiles and nonavian dinosaurs. In: Endo H, Frey R. Anatomical Imaging: Towards a New Morphology. Tokyo: Springer-Verlag; pp. 67–87. [Google Scholar]
- 9. Witmer LM, Ridgely RC (2009) New insights into the brain, braincase, and ear region of Tyrannosaurus (Dinosauria, Theropoda), with implications for sensory organization and behavior. Anat Rec 292: 1266–1296. doi:10.1002/ar.20983. [DOI] [PubMed] [Google Scholar]
- 10. Paulina Carabajal A, Currie PJ (2012) New information on the braincase and endocast of Sinraptor dongi (Theropoda: Allosauroidea): Ethmoidal region, endocranial anatomy and pneumaticity. Vert PalAs 50: 85–101
- 11. Paulina Carabajal A (2012) Neuroanatomy of titanosaurid dinosaurs from the Upper Cretaceous of Patagonia, with comments on endocranial variability within Sauropoda. Anat Rec 295: 2141–2156. doi:10.1002/ar.22572. [DOI] [PubMed] [Google Scholar]
- 12. Sedlmayr JC (2002) Anatomy, evolution, and functional significance of cephalic vasculature in Archosauria. Unpublished PhD thesis, Ohio University, Ohio, 398 pp. . [Google Scholar]
- 13. Witmer LM, Chatterjee S, Franzosa J, Rowe T (2003) Neuroanatomy of flying reptiles and implications for flight, posture and behavior. Nature 425: 950–953. doi:10.1038/nature02048. PubMed: 14586467. [DOI] [PubMed] [Google Scholar]
- 14. Holliday CM, Gardner NM (2012) A new eusuchian crocodyliform with novel cranial integument and its significance for the origin and evolution of crocodylia. PLOS ONE 7(1): e30471. doi:10.1371/journal.pone.0030471. PubMed: 22303441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Hopson JA (1979) Paleoneurology. In: Gans C. Biology of the Reptilia. vol 9 New York: Academic Press; pp 39–146. [Google Scholar]
- 16. Zangerl R (1960) The vertebrate fauna of the Selma Formation of Alabama. V. An advanced cheloniid sea turtle. Fieldiana Geol Mem 3: 281–312. [Google Scholar]
- 17. Gaffney ES, Zangerl R (1968) A revision of the chelonian genus Bothremys (Pleurodira: Pelomedusidae). Fieldiana Geol 16: 193–239. [Google Scholar]
- 18. Gaffney ES (1977) An endocranial cast of the side-necked turtle, Bothremys, with a new reconstruction of the palate. Am Museum Nov 2639: 1–12. [Google Scholar]
- 19. Gaffney ES (1982) Cranial morphology of the baenid turtles. Am Museum Nov 2737: 1–22. [Google Scholar]
- 20. Thewissen JGM, Nummela S (2008) Sensory Evolution on the Threshold: Adaptations in Secondarily Aquatic Vertebrates. California: University of California Press. 351pp. [Google Scholar]
- 21. Walsh SA, Barrett PM, Milner AC, Manley G, Witmer LM (2009) Inner ear anatomy is a proxy for deducing auditory capability and behavior in reptiles and birds. Proc R Soc Lond B: 1355–1360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Edinger T (1929) Die fossilen Gehirne. Ergeb, Anat, Entwicklungsgesch B. 28 Berlin: Julius Springer Verlag; p. 249. [Google Scholar]
- 23. Wyneken J (2001) The Anatomy of Sea Turtles. U.S. Department of Commerce NOAA Technical Memorandum NMFS-SEFSC-470. pp. 1–172.
- 24. Edinger T (1934) Anton Fritsch’s "Grosshirn von Polyptychodon" ist der Steinkern eines Schildkrötenschadels. Psych Neurol Bladen 3–4: 396–404. [Google Scholar]
- 25. Joyce WG (2007). Phylogenet Relat Mesozoic Turtles Bull Peabody Mus Nat Hist 48: 3–102. doi:10.3374/0079-032X(2007)48[3:PROMT]2.0.CO;2. [Google Scholar]
- 26. de Broin F (1994) Données préliminaires sur les chéloniens du Tithonien inférieur des calcaires lithographiques de Canjuers (Var, France). Geobios Mem Spec 16: 167–175. [Google Scholar]
- 27. Buffetaut E (1994) Tetrapods from the Late Jurassic and Early Cretaceous lithographic limestones of Europe: a comparative review. Geobios Mem Spec 16: 259–265. [Google Scholar]
- 28. Bardet N (1995) Évolution et extinction des reptiles marins au cours du Mésozoique. Palaeovertebrata 24: 177–283. [Google Scholar]
- 29. Billon-Bruyat JP, Lécuyer C, Martineau F, Mazin J-M (2005) Oxygen isotope compositions of Late Jurassic vertebrate remains from lithographic limestones of western Europe: implications for the ecology of fish, turtles, and crocodilians. Palaeogeogr Palaeoclimatol Palaeoecol 216: 359–375. doi:10.1016/j.palaeo.2004.11.011. [Google Scholar]
- 30. Gaffney ES (1975) A taxonomic revision of the Jurassic turtles Portlandemys and Plesiochelys . Am Museum Nov 2574: 1–19. [Google Scholar]
- 31. Gaffney ES (1976) Cranial morphology of the European Jurassic turtles Portlandemys and Plesiochelys . Bull Am Museum Nat Hist 157: 487–544. [Google Scholar]
- 32. Sterli J, Müller J, Anquetin J, Hilger A (2010) The parabasisphenoid complex in Mesozoic turtles and the evolution of the testudinate basicranium. Can J Earth Sci 47: 1337–1346. doi:10.1139/E10-061. [Google Scholar]
- 33. Gaffney ES (1979) Comparative cranial morphology of recent and fossil turtles. Bull Am Museum Nat Hist 164: 65–376. [Google Scholar]
- 34. Wersäll J, Bagger-Sjöbäck D (1974) Morphology of the vestibular sense organ. In: Komhuber HH. Handbook of Sensory Physiology, Vestibular System. Basic Mechanisms. Berlin: Springer-Verlag; pp. 123–170. [Google Scholar]
- 35. Weber EG (1978) The Reptile Ear. Princeton: Princeton Press. 1038pp. [Google Scholar]
- 36. Gaffney ES (1990) The comparative osteology of the Triassic turtle Proganochelys . Bull Am Museum Nat Hist 194: 1–263. [Google Scholar]
- 37. Gaffney ES, Kitching JW (1995) The morphology and relationships of Australochelys, an Early Jurassic turtle from South Africa. Am Museum Nov 3130: 1–29. [Google Scholar]
- 38. Sterli J, de la Fuente MS, Rougier GW (2007) Anatomy and relationships of Palaeochersis talampayensis, a Late Triassic turtle from Argentina. Palaeontogr A, 281: 1–61. [Google Scholar]
- 39. Gaffney ES (1975) A phylogeny and classification of the higher categories of turtles. Bull Am Museum Nat Hist 155: 387–436. [Google Scholar]
- 40. Sterli J, de la Fuente MS (2010) Anatomy of Condorchelys antiqua Sterli 2008 and the origin of modern jaw closure mechanism in turtles. J Vert Paleont 30: 251–366
- 41. Parsons TS (1959) Nasal anatomy and the phylogeny of reptiles. Evolution 13: 175–187. doi:10.2307/2405871. [Google Scholar]
- 42. Romain D, Droulezb J, Allain R, Berthozb A, Janvier P et al. (2010) Motion from the past. A new method to infer vestibular capacities of extinct species. C R Palevol 9: 397–410. doi:10.1016/j.crpv.2010.07.012. [Google Scholar]
- 43. Boistel R, Herrel A, Daghfous G, Libourel P-A, Boller E et al. (2010) Asisted walking in Malagasy dwarf chamaeleons. Biol Lett 6: 740–743. doi:10.1098/rsbl.2010.0322. PubMed: 20462882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Christensen-Dalsgaard J, Brandt C, Willis KL, Christensen CB, Ketten D et al. (2012) Specialization for underwater hearing by the tympanic middle ear of the turtle, Trachemys scripta elegans . Proc R Soc Lond B 279: 2816–2824. doi:10.1098/rspb.2012.0290. PubMed: 22438494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Lenhardt ML, Klinger RC, Musick JA (1985) Marine turtle middle-ear anatomy. J Aud Res 25: 66–72. PubMed: 3836997. [PubMed] [Google Scholar]
- 46. Manley GA, Kraus JEM (2010) Exceptional high-frequency hearing and matched vocalizations in Australian pygopod geckos. J Exp Biol 213: 1876–1885. doi:10.1242/jeb.040196. PubMed: 20472775. [DOI] [PubMed] [Google Scholar]
- 47. McDowell SB Jr (1961) On the major arterial canals in the ear region of testudinoid turtles and the classification of the Testudinoidea. Bull Museum Comp Zool 125: 23–39. [Google Scholar]
- 48. Albrecht PW (1967) The cranial arteries and cranial arterial foramina of the turtle genera Chrysemys, Sternotherus, and Trionyx: a comparative study with analysis of posible evolutionary implications. Tulane Stud Zoology 14: 81–89. [Google Scholar]
- 49. Brinkman DB, Nicholls EL (1991) Anatomy and relationships of the turtle Boremys pulchra (Testudines: Baenidae). J Vert Paleontol 11: 302–315
- 50. Jamniczcky HA, Russell AP (2004) Cranial arterial foramen diameter in turtles: Quantitative assessment of size-independent phylogenetic signal. Anim Biol 54: 417–436. doi:10.1163/1570756042729537. [Google Scholar]
- 51. Jamniczky HA, Russell AP (2007) Reappraisal of patterns of nonmarine cryptodiran turtle carotid circulation: evidence from osteological correlates and soft tissues. J Morphol 268: 571–587. doi:10.1002/jmor.10539. PubMed: 17437297. [DOI] [PubMed] [Google Scholar]
- 52. Brinkman DB, Eberth DA, Xu X, Clark, JM, Wu X-C (2013) Turtles from the Jurassic Shishugou Formation of the Junggar Basin, People’s Republic of China, with comments on the basicranial region of basal eucryptodires : Brinkman DB, Holroyd PA, Gardner JD, editors. Morphology and Evolution of Turtles, Vertebrate Paleobiology and Paleoanthropology. Dorsrecht: Springer Verlag; . pp. 147–172 [Google Scholar]
- 53. Miyashita T (2013) Geometric and developmental perspectives on the evolution of the skull and internal carotid circulation in turtles. In: Brinkman DB, Holroyd PA, Gardner JD. Morphology and Evolution of Turtles, Vertebrate Paleobiology and Paleoanthropology. Dorsrecht: Springer Verlag; pp. 71–101. [Google Scholar]
- 54. Joyce WG, Parham JF, Gauthier JA (2004) Developing a protocol for the conversion of rank-based taxon names to phylogenetically defined clade names, as exemplified by turtles. J Paleontol 78: 989–11013. doi:10.1666/0022-3360(2004)078. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(PDF)