

Abstract
Few attempts have been made to study the alleviating effects of signal molecules on zoysiagrass ( Zoysia japonica ) under drought stress. Calcium chloride has been shown to ameliorate the adverse effects of drought stress on many plants. It is necessary to investigate how to enhance drought tolerance of zoysiagrass using calcium chloride. The study elucidated the effects of calcium chloride on zoysiagrass under drought conditions by investigating the following parameters: biomass, chlorophyll (Chl) content, net photosynthetic rate (Pn), chlorophyll fluorescence, antioxidant enzymes, proline content, and malondialdehyde (MDA) content. Experimental conditions consisted of an aqueous CaCl2 solution at 5, 10, and 20 mM sprayed on zoysiagrass leaves for 3 d, following by an inducement of drought conditions by withholding water for 16 d. Under drought conditions, all CaCl2 pretreatments were found to increase the above-ground fresh biomass, as well as below-ground fresh and dry biomass. The resulting Chl (a, b, a+b) contents of the 5 and 10 mM CaCl2 pretreatment groups were higher than those of the control. In the later stages of drought conditions, the chlorophyll fluorescence parameter Fv/Fm was higher in leaves treated with 10 mM CaCl2 than in the leaves of the other two treatment groups. Zoysiagrass pretreated with 10 mM CaCl2 possessed both the maximum observed Pn and antioxidant enzyme activities. Meanwhile, lower MDA and proline contents were recorded in the plants pretreated with 5 and 10 mM CaCl2 under drought conditions. As a whole, the drought tolerance of zoysiagrass was improved to some extent by the application of a moderate calcium concentration.
Introduction
Drought is a major environmental factor that adversely affects the productivity and survival of plants [1]. Plants that experience drought conditions often appear serious physiological and biochemical dysfunctions, including: reduction in turgor, growth, photosynthetic rate, and stomatal conductance, as well as damage to various cellular components [2,3]. Plants utilize various mechanisms to adapt to and withstand drought conditions [4]. For example, in plants subjected to drought, the generation of a variety of active oxygen species (ROS) has been observed. These include: superoxide anion (O- 2), hydrogen peroxide (H2O2), and hydroxyl radical (·OH), all of which can cause oxidative damage to plants, including membrane lipid peroxidation [5]. When plant capacity to detoxify ROS increases, an increase in drought tolerance can be observed [6,7]. The rapid accumulation of proline, an osmoregulation substance in cells, constitutes another biochemical mechanism in plants that acts to achieve drought tolerance [8,9].
Various physiological practices have been applied to alleviate the adverse effects of a water deficit on normal plant functioning. For example, researchers have applied the following in an effort to improve plant growth under drought conditions: calcium ion [10,11], plant growth regulators [12–15], and other substances [16–18].
Importantly, calcium is involved in the regulatory mechanisms that plants activate to adjust to adverse environmental conditions of drought [10,11,19], heat [20,21], cold [22,23], salt [24,25], and heavy metal [26,27]. Further, calcium has been shown to ameliorate the adverse effects of water stress on plants [28], and is involved in signaling anti-drought responses [10]. Calcium appears to play a central role in many defense mechanisms that are induced by drought, and Ca2+ signaling is required for the acquisition of drought tolerance or resistance [29].
Zoysia japonica (zoysiagrass) is an important turfgrass species in East Asia, and particularly in China, Korea, and Japan [30]. In turfgrass management, drought constitutes a major factor limiting grass growth. Some researchers have studied the morphological and physiological responses of zoysiagrass to drought conditions [3,31–33]. However, to our knowledge, few attempts have been made to study the alleviation of such negative effects to drought stress. It is therefore necessary to investigate how to enhance drought tolerance using signal molecules.
As a result, we studied the effects of calcium applications on the physiological and biochemical mechanisms in leaves of Z . japonica under drought conditions. In the present study, we analyzed plant growth, as well as photosynthesis, chlorophyll fluorescence, lipid peroxidation, proline content, and activities of antioxidant enzymes.
Materials and Methods
Plant materials and treatments
Zoysiagrass plants were collected from 1-year-old turfgrass plots at the Ecological Research Center, Liaoning University. Plants were grown in plastic pots (diameter: 13 cm diameter, depth: 14.5 cm) filled with a mixture of topsoil and coarse river sand (1:1) in a greenhouse for one week (25°C day / 20 °C night, 16 hr/8 hr light / dark period, 800 µmol m-2 s-1 photosynthetically active radiation, and 75% relative humidity). The pots were then transferred to the experiment field and grown under natural conditions at the Liaoning University for one month. During this period, plants were watered to field capacity (≥ 75%) every 2 d and were fertilized once a week using compound fertilizer. Once the assay began, fertilization was stopped.
Plants were randomized and divided into four groups of 60 individuals. Aqueous calcium chloride solutions at 5, 10, and 20 mM containing 0.1% Tween 20 were sprayed on the leaves of three of the four groups until runoff thrice a day for 3 d. Control plants were sprayed with distilled water containing 0.1% Tween 20. Drought was induced through withholding water for one week. The soil water content then reached 7-8%, and maintained this degree until 16 d after CaCl2 application. Thus, the treatments were as follows: (1) Control: non-CaCl2 and drought, (2) 5 mM: 5 mM CaCl2 pretreatment and drought, (3) 10 mM: 10 mM CaCl2 pretreatment and drought, and (4) 20 mM: 20 mM CaCl2 pretreatment and drought. Each treatment was carried out in triplicate.
Soil water content
After the treatment period was complete, the soil was taken from each pot and weighed immediately to obtain its wet weight. The soil was then dried for 48 hr at 90 °C, and wet and dry weights were then used to estimate soil water content (SWC). SWC was calculated as [(soil wet weight) - (soil dry weight)] / (soil wet weight) × 100.
Estimation of plant biomass
The above-ground plants in the 13 cm diameter pots were clipped at 16 d after CaCl2 application. The roots in the pots were washed with distilled water. Above- and below-ground fresh and dry biomass weights were recorded, and dry biomass was obtained by oven-drying samples at 80 °C for 24 hr.
Determination of Chl content and net photosynthetic rate
The Chl content was quantified using the method of Agrawal and Rathore [34]. Chl content was extracted from 0.1 g of leaf discs with 10 ml 80% acetone, and the absorbance of the solution was measured at 663 and 645 nm.
Between 10:00 and 12:00 on each sampling day, gas exchange by leaves (one leaf per plant, three plants per replicate) was measured with a portable photosynthesis system (Li-6400, Li-Cor, Lincoln, NE, USA). Net photosynthetic rate (Pn) was measured under ambient CO2 (370 µmol mol-1). Photosynthetic photon flux density (PPFD) was set at 800 µmol m−2 s−1 in the cuvette containing the leaf for Pn measurement.
Measurement of chlorophyll fluorescence
Fluorescence parameters of intact leaves (three leaves per plant, three plants per replicate) were measured using Li-6400-40LCF (Li-Cor, Lincoln, NE, USA). The minimal chlorophyll fluorescence (Fo) level when photosystem II centres are open was measured after applying a far-red pulse for 6 s. The maximal fluorescence (Fm) after 30 min of dark adaptation was measured after applying a saturating flash for 0.8 s. Maximal photochemical efficiency of PSII (Fv/Fm) was expressed as: Fv/Fm=(Fm-Fo)/Fm.
Extraction and assay of antioxidant enzymes
Fresh leaf sample (0.5 g) was homogenized in 5 ml extraction buffer (0.1 M phosphate buffer pH 6.8) using a mortar and pestle on ice. The homogenate was then centrifuged at 12,000 × g for 15 min at 4 °C, and the supernatant was used as the crude extract for superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD).
SOD activity was measured according to the method of Beyer and Fridovich [35]. SOD activity was assayed by measuring the ability of the enzyme in the crude extract to inhibit the photochemical reduction of nitroblue tetrazolium (NBT) by photochemically-generated superoxide radicals. One unit of SOD was defined as the amount of enzyme required to inhibit the rate of reduction NBT by 50% at 25 °C.
POD and CAT activities were assayed following the method of Chance and Maehly [36] with some modification. The POD reaction solution contained 50 mM phosphate buffer (pH 7.8), 25 mM guaiacol, 200 mM H2O2, and the enzyme extract. Changes in absorbance of the reaction solution at 470 nm were determined. CAT activity was assayed in a reaction mixture containing 50 mM potassium phosphate buffer (pH 7.0, containing 0.1 mM EDTA), 200 mM H2O2, and the enzyme extract. The reaction was started with the addition of the enzyme extract, and the decomposition rate of H2O2 was followed at 240 nm.
Estimation of MDA content
MDA content was determined using the method of Fu and Huang [37]. Fresh leaf sample (1.0 g) was homogenized with 4 ml of 0.1% (w/v) trichloroacetic acid (TCA) in an ice bath. The homogenate was centrifuged at 12,000 × g for 20 min, and the supernatant was used for lipid peroxidation analysis. A total of 4 ml of 0.5% thiobarbituric acid (TBA) in 20% TCA was added to 1 ml aliquot of the supernatant. The mixture was incubated in boiling water for 30 min. MDA content was then detected spectrophotometrically at 532 nm and corrected for nonspecific turbidity at 600 nm.
Estimation of proline content
Free proline content was estimated using the method described by Bates et al. [38]. Fresh leaves (0.5 g) were extracted in 5 ml of 3% sulphosalicylic acid, and the homogenates were centrifuged at 3,000 × g for 20 min. A total of 2 ml of the supernatant was reacted with 4 ml of 2.5% acid ninhydrin reagent and 2 ml of glacial acetic acid in a test tube. After boiling the mixture in a water bath at 100 °C for 60 min, the reaction was stopped by cooling the tubes in ice bath for 5 min. The chromophore formed was extracted with 4 ml of toluene and mixed vigorously by vortexing for 0.5 min. Absorbance of the resulting organic layer was measured at 520 nm. The concentration of proline was estimated by referring to a standard curve for L-proline.
Statistical analysis
All experiments were conducted with three replicates, and results were expressed as mean ± standard deviation (SD). All data were subjected to one-way analysis of variance (ANOVA) and LSD multiple comparison test (p < 0.05) using the SPSS statistical package.
Results
SWC during drought stress
The SWC values observed during the treatment period are shown in Figure 1. During treatment, the SWC values for all CaCl2 pretreatments and the control significantly decreased to approximately 8% at 7 days after drought. The SWC values for all CaCl2 pretreatments and the control were not significantly different from one another, suggesting that the level of drought was the same for all treatments.
Figure 1. Soil water content during drought.
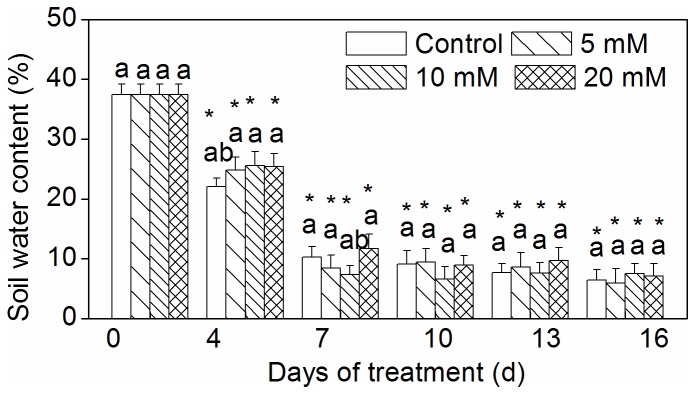
Vertical bars represent standard deviations (n=3). The different letters indicate significant difference among different treatments on a given day (p < 0.05, LSD multiple test). The asterisks indicate significant difference between given day and 0 d at the same treatment (p < 0.05, LSD multiple test).
Changes in above- and below-ground biomass
An increase in the above- and below-ground fresh biomass was observed for all CaCl2 pretreatment groups, and the greatest increase was observed for the 10 mM CaCl2 pretreatment group (Figure 2A). All CaCl2 pretreatment groups had below-ground dry biomass values that were higher than those of the control, while above-ground dry biomass increased in the 10 mM CaCl2 pretreatment group (Figure 2B).
Figure 2. Effect of CaCl2 on above- and below-ground biomass of Zoysia japonica under drought.

(A) Fresh weight; (B) Dry weight. Vertical bars represent standard deviations (n=3). The different letters indicate significant difference at p < 0.05 (LSD multiple test).
Changes in Chl content
The Chl content (Chl a, Chl b and Chl a+b) values decreased with drought treatment (Figure 3). An increased Chl a content was observed for the 10 mM CaCl2 pretreatment group alone (Figure 3A). Meanwhile, Chl b and Chl a+b contents of the 5 and 10 mM CaCl2 pretreatment groups were higher than those of the control (Figure 3B, C), while Chl b and Chl a+b contents for the 10 mM CaCl2 pretreatment group was higher than those of the 5 mM CaCl2 pretreatment group.
Figure 3. Effect of CaCl2 on chlorophyll content of Zoysia japonica leaves under drought.

(A) Chl a content; (B) Chl b content; (C) Chl a+b content. Vertical bars represent standard deviations (n=3). The different letters indicate significant difference among different treatments on a given day (p < 0.05, LSD multiple test). The asterisks indicate significant difference between given day and 0 d at the same treatment (p < 0.05, LSD multiple test).
Changes in Pn and chlorophyll fluorescence
Compared to the control, Pn of the 10 and 20 mM CaCl2 pretreatment groups significantly increased at the early stages of drought (from 1 to 7 days after drought) (Figure 4). At the later stages of drought, Pn of the 10 mM CaCl2 pretreatment group alone was positive, because it also exhibited an increased Pn compared to the control.
Figure 4. Effect of CaCl2 on net photosynthetic rate of Zoysia japonica leaves under drought.
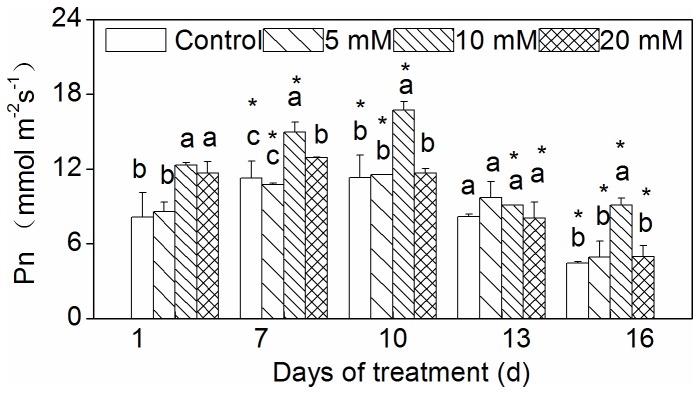
Vertical bars represent standard deviations (n=3). The different letters indicate significant difference among different treatments on a given day (p < 0.05, LSD multiple test). The asterisks indicate significant difference between given day and 0 d at the same treatment (p < 0.05, LSD multiple test).
The chlorophyll fluorescence parameter Fv/Fm ratio decreased as drought was prolonged. The Fv/Fm ratio was higher in leaves treated with 10 mM CaCl2 than in those of the other treatments (Figure 5).
Figure 5. Effect of CaCl2 on Fv/Fm of Zoysia japonica leaves under drought.

Vertical bars represent standard deviations (n=3). The different letters indicate significant difference among different treatments on a given day (p < 0.05, LSD multiple test). The asterisks indicate significant difference between given day and 0 d at the same treatment (p < 0.05, LSD multiple test).
Changes in antioxidant enzymes activities
The CaCl2 pretreatments had significant effects on antioxidant enzymes under drought conditions (Figure 6). SOD activity was increased in the 5 and 10 mM CaCl2 pretreatment groups, and there was no significant change in the 20 mM CaCl2 pretreatment group (Figure 6A). As drought was prolonged, SOD activity in the stressed leaves of all treatments increased. During drought stress, the SOD activity of the 10 mM CaCl2 pretreatment group was higher than that of the control, and that of the 20 mM CaCl2 pretreatment was lower. No significant difference in the SOD activity of the stressed leaves of the 5 mM CaCl2 pretreatment group and the control was observed (except at day 9). The 20 mM CaCl2 pretreatment group alone showed decreased POD activity (Figure 6B). Under drought conditions, the 10 mM CaCl2 pretreatment group showed significantly increased POD activity in comparison to the control. The 10 mM CaCl2 pretreatment group showed significantly increased CAT activity under no stress and drought stress (Figure 6C). The 20 mM CaCl2 pretreatment group exhibited reduced CAT activity after 9 days of drought.
Figure 6. Effect of CaCl2 on SOD, POD, and CAT activity on Zoysia japonica leaves under drought.

(A) SOD activity; (B) POD activity; (C) CAT activity. Vertical bars represent standard deviations (n=3). The different letters indicate significant difference among different treatments on a given day (p < 0.05, LSD multiple test). The asterisks indicate significant difference between given day and 0 d at the same treatment (p < 0.05, LSD multiple test).
Changes in MDA and proline content
MDA content showed an increasing trend under drought conditions (Figure 7A). Lower MDA content was recorded in the plants pretreated with 5 and 10 mM CaCl2 under no or drought stress. There was no significant change observed between the 20 mM CaCl2 pretreatment group and the control, with the exception of the change observed at 12 days after drought.
Figure 7. Effect of CaCl2 on MDA and proline contents on Zoysia japonica leaves under drought.

(A) MDA content; (B) proline content. Vertical bars represent standard deviations (n=3). The different letters indicate significant difference among different treatments on a given day (p < 0.05, LSD multiple test). The asterisks indicate significant difference between given day and 0 d at the same treatment (p < 0.05, LSD multiple test).
When subjected to progressive water stress, the proline content in zoysiagrass increased (Figure 7B), and the 5 and 10 mM CaCl2 pretreatment groups exhibited a significant decrease in proline accumulation in comparison to the control. The lowest proline content was observed in plants pretreated with 10 mM CaCl2.
Discussion
The present investigation effectively confirmed that CaCl2 pretreatment could enhance the drought tolerance of zoysiagrass. Many studies have indicated that CaCl2 could act as a physiological treatment to increase plant tolerance. CaCl2 even at the relatively low concentrations used in this study, effectively contributed to the alleviation of drought stress.
Under stress conditions, the dry weight and Chl content are generally thought to be important parameters for growth measurement. In the present experiment, above- and below-ground fresh biomass and below-ground dry biomass increased in all CaCl2 pretreatment groups in comparison to the control (Figure 2). These results suggest that Ca2+ plays an important role in alleviating the damage to zoysiagrass incurred under drought conditions. Similar alleviation activity by applied CaCl2 has been reported for different plant species under different stress conditions [39–41]. Upadhyaya et al. [11] also reported that using foliar sprayed CaCl2 even after drought could increase the dry mass of leaves in the recovery phase. Increasing Ca2+ availability may reduce drought damage by increasing membrane integrity [19,42,43]. Our results showed a decrease in Chl content for all pretreatment groups and the control under drought (Figure 3). This decrease in Chl content under drought is due to the destruction of pigments and the instability of the pigment-protein complex [44]. The 5 and 10 mM CaCl2 pretreatment groups exhibited noticeably increased Chl content in the drought-stressed plants (Figure 3). It seems that the applied CaCl2 might prevent damage from cellular dehydration by balancing the osmotic strength of the cytoplasm [39]. In the present study, observed increases in biomass and Chl content values were greatest in the 10 mM CaCl2 pretreatment group (Figures 2, 3). The change in Pn for the 10 mM CaCl2 pretreatment group exhibited a similar trend (Figure 4). Further, similar results were reported by Amor et al. [24], who stated that the addition of 3.5 mM calcium was more effective than the addition of 20 mM in reducing NaCl stress for Cakile maritima .
Fv/Fm is an indicator of the efficiency of the photosynthetic apparatus [45]. In the present study, the Fv/Fm ratio was higher in plants pretreated with 10 mM CaCl2 than in other treatments (Figure 5). This demonstrates that the optimum CaCl2 concentration significantly enhances grass tolerance of drought without significantly affecting plant growth and development.
Oxidative stress is a key component of environmental stress, and increased SOD activity was correlated with increased protection from the damage associated with oxidative stress [7]. In the present study, SOD activity increased as drought was prolonged, and the SOD activity of the 10 mM CaCl2 pretreatment was higher than that of the control. The product of SOD activity is H2O2, which is still toxic and must be eliminated by conversion to H2O in subsequent reactions [24]. Some authors have indicated that the CAT/POD system might act cooperatively to remove H2O2 at a maximal rate and at a minimal expense of power reduction [27,46]. In fact, our results showed that POD activity of the 5 and 10 mM CaCl2 pretreatment groups increased compared to the control under drought conditions, and CAT activity of the 10 mM CaCl2 pretreatment group also increased. Interestingly, SOD, POD, and CAT activity levels of the 10 mM CaCl2 pretreatment were significantly higher than those of the control (Figure 6). This implies that an increase of antioxidant enzymes effectively scavenges ROS to provide protection from cellular oxidative damage. It has also been reported that external Ca2+ can induce significant increases in SOD, POD, and CAT activity in maize and cool season grasses seedlings [23,47].
We also observed that MDA contents in the leaves of pretreated and control groups gradually increased as drought continued (Figure 7A). According to Guimarães et al. [43], high MDA content may result in electrolyte leakage, indicating a loss of membrane integrity. However, the 5 and 10 mM CaCl2 pretreated sample groups showed lower MDA contents than did the control under drought. Some studies have shown that CaCl2 pretreatment decreased MDA content [20,47]. This result was in accordance with the increase in growth and antioxidant enzymes activities of zoysiagrass. This proved that pretreatment with the appropriate CaCl2 concentration alleviated and postponed oxidation damage that resulted from drought.
Free proline accumulation in the leaves under stress conditions is of utmost importance for plant adaptation during stress [48,49]. Our data suggests that proline content increased in zoysiagrass for all pretreatments under drought (Figure 7B). It was found that the availability of foliar CaCl2 could modulate endogenous proline accumulation under water stress, because the leaves of stressed plants that were pretreated with CaCl2 accumulated less proline than did the nontreated plants. Our results are consistent with those of Jaleel et al. (2007), who reported that addition of CaCl2 to drought stressed plants lowered the proline concentration by increasing the level of proline degrading enzyme and decreasing the proline synthesizing enzyme.
In summary, foliar application of CaCl2 can help zoysiagrass avoid the stress effects resulting from drought to some extent, as indicated by Chl content, Pn, chlorophyll fluorescence, antioxidant enzyme activities, lipid peroxidation, and proline content. These findings are very helpful for determining the ways in which conditions should be manipulated to secure the survival and improve the growth of turfgrass under drought conditions.
Funding Statement
This research was financially supported by the National Nature Science Foundation of China (31070285, 31270369). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Monclus R, Dreyer E, Villar M, Delmotte FM, Delay D et al. (2006) Impact of drought on productivity and water use efficiency in 29 genotypes of Populus deltoides x Populus nigra. New Phytol 169: 765-777. doi:10.1111/j.1469-8137.2005.01630.x. PubMed: 16441757. [DOI] [PubMed] [Google Scholar]
- 2. Xu X, Peng G, Wu C, Korpelainen H, Li C (2008) Drought inhibits photosynthetic capacity more in females than in males of Populus cathayana . Tree Physiol 28: 1751-1759. doi:10.1093/treephys/28.11.1751. PubMed: 18765380. [DOI] [PubMed] [Google Scholar]
- 3. Soares-Cordeiro AS, Carmo-Cilva AE, Bernardes da Silva A, Marques da Silva J, Keys AJ et al. (2009) Effects of rapidly imposed water deficit on photosynthetic parameters of three C4 grasses. Photosynthetica 47: 304-308. doi:10.1007/s11099-009-0047-y. [Google Scholar]
- 4. Diaz-Espejo A, Nicolás E, Fernández JE (2007) Seasonal evolution of diffusional limitations and photosynthetic capacity in olive under drought. Plant Cell Environ 30: 922-933. doi:10.1111/j.1365-3040.2007.001686.x. PubMed: 17617820. [DOI] [PubMed] [Google Scholar]
- 5. Sharma P, Dubey RS (2005) Drought induces oxidative stress and enhances the activities of antioxidant enzymes in growing rice seedlings. Plant Growth Regul 46: 209-221. doi:10.1007/s10725-005-0002-2. [Google Scholar]
- 6. de Carvalho MHC (2008) Drought stress and reactive oxygen species: production, scavenging and signaling. Plants Signal Behav 3: 156-165. doi:10.4161/psb.3.3.5536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Miller G, Suzuki N, Ciftci-yilmaz S, Mittler R (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ, 33: 453-467. PubMed: 19712065. [DOI] [PubMed] [Google Scholar]
- 8. Maggio A, Miyazaki S, Veronese P, Fujita T, Ibeas JI et al. (2002) Does proline accumulation play an active role in stress-induced growth reduction? Plant J 31: 699-712. doi:10.1046/j.1365-313X.2002.01389.x. PubMed: 12220262. [DOI] [PubMed] [Google Scholar]
- 9. Mafakheri A, Siosemardeh A, Bahramnejad B, Struik PC, Sohrabi E (2010) Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust J Crops Sci 4: 580-585. [Google Scholar]
- 10. Shao HB, Song WY, Chu LY (2008) Advances of calcium signals involved in plant anti-drought. C R Biol 331: 587-596. doi:10.1016/j.crvi.2008.03.012. PubMed: 18606388. [DOI] [PubMed] [Google Scholar]
- 11. Upadhyaya H, Panda SK, Dutta BK (2011) CaCl2 improves post-drought recovery potential in Camellia sinensis (L) O Kuntze. Plant Cell Rep 30: 495-503. doi:10.1007/s00299-010-0958-x. PubMed: 21153899. [DOI] [PubMed] [Google Scholar]
- 12. Ma QQ, Wang W, Li YH, Li DQ, Zou Q (2006) Alleviation of photoinhibition in drought-stressed wheat (Triticum aestivum) by foliar-applied glycinebetaine. J Plant Physiol 163: 165-175. doi:10.1016/j.jplph.2005.04.023. PubMed: 16399007. [DOI] [PubMed] [Google Scholar]
- 13. Shan CJ, Liang ZS (2010) Jasmonic acid regulates ascorbate and glutathione metabolism in Agropyron cristatum leaves under water stress. Plant Sci 178: 130-139. doi:10.1016/j.plantsci.2009.11.002. [Google Scholar]
- 14. Wang KH, Zhang XZ, Ervin E (2012) Antioxidative responses in roots and shoots of creeping bentgrass under high temperature: Effects of nitrogen and cytokinin. J Plant Physiol 169: 492-500. doi:10.1016/j.jplph.2011.12.007. PubMed: 22226339. [DOI] [PubMed] [Google Scholar]
- 15. Hojati M, Modarres-Sanavy SAM, Ghanati F, Panahi M (2011) Hexaconazole induces antioxidant protection and apigenin-7-glucoside accumulation in Matricaria chamomilla plants subjected to drought stress. J Plant Physiol 168: 782-791. doi:10.1016/j.jplph.2010.11.009. PubMed: 21208683. [DOI] [PubMed] [Google Scholar]
- 16. Zhao HJ, Tan JF, Qi CM (2007) Photosynthesis of Rehmannia glutinosa subjected to drought stress is enhanced by choline chloride through alleviating lipid peroxidation and increasing proline accumulation. Plant Growth Regul 51: 255-262. doi:10.1007/s10725-007-9167-1. [Google Scholar]
- 17. Ahmed M (2011) Fayyaz-ul-Hassen Khurshid Y. Does Silicon Irrig Have Implant Drought Tolerance Mech Sorghum? Agr Water Manage 98: 1808-1812. [Google Scholar]
- 18. Ishibashi Y, Yamaguchi H, Yuasa T, Iwaya-Inoue M, Arima S et al. (2011) Hydrogen peroxide spraying alleviates drought stress in soybean plants. J Plant Physiol 168: 1562-1567. doi:10.1016/j.jplph.2011.02.003. PubMed: 21377755. [DOI] [PubMed] [Google Scholar]
- 19. Ma R, Zhang M, Li B, Du G, Wang J et al. (2005) The effects of exogenous Ca2+ on endogenous polyamine levels and drought-resistant traits of spring wheat grown under arid conditions. J Arid Environ 63: 177-190. doi:10.1016/j.jaridenv.2005.01.021. [Google Scholar]
- 20. Wang Y, Yu QY, Tang XX, Wang LL (2009) Calcium pretreatment increases thermotolerance of Laminaria japonica sporophytes. Prog Nat Sci 19: 435-442. doi:10.1016/j.pnsc.2008.07.016. [Google Scholar]
- 21. Tan W, Meng QW, Brestic M, Olsovska K, Yang XH (2011) Photosynthesis is improved by exogenous calcium in heat-stressed tobacco plants. J Plant Physiol 168: 2063-2071. doi:10.1016/j.jplph.2011.06.009. PubMed: 21803445. [DOI] [PubMed] [Google Scholar]
- 22. Zhou B, Guo Z (2009) Calcium is involved in the abscisic acid-induced ascorbate peroxidase, superoxide dismutase and chilling resistance in Stylosanthes guianensis . Biol Plant 53: 63-68. doi:10.1007/s10535-009-0009-z. [Google Scholar]
- 23. Wang Y, Yang ZM, Zhang QF, Li JL (2009) Enhanced chilling tolerance in Zoysia matrella by pre-treatment with salicylic acid, calcium chloride, hydrogen peroxide or 6-benzylaminopurine. Biol Plant 53: 179-182. doi:10.1007/s10535-009-0030-2. [Google Scholar]
- 24. Amor NB, Megdiche W, Jimenez A, Sevilla F, Abdelly C (2010) The effect of calcium on the antioxidant systems in the halophyte Cakile maritima under salt stress. Acta Physiol Plants 32: 453-461. doi:10.1007/s11738-009-0420-2. [Google Scholar]
- 25. Zehra A, Gul B, Ansari R, Khan MA (2012) Role of calcium in alleviating effect of salinity on germination of Phragmites karka seeds. South Afr J Bot 78: 122-128. doi:10.1016/j.sajb.2011.05.016. [Google Scholar]
- 26. Antosiewicz DM, Hennig J (2004) Overexpression of LCT1 in tobacco enhances the protective action of calcium against cadmium toxicity. Environ Pollut 129: 237-245. doi:10.1016/j.envpol.2003.10.025. PubMed: 14987809. [DOI] [PubMed] [Google Scholar]
- 27. Siddiqui MH, Al-Whaibi MH, Basalah MO (2011) Interactive effect of calcium and gibberellin on nickel tolerance in relation to antioxidant systems in Triticum aestivum L. Protoplasma 248: 503-511. doi:10.1007/s00709-010-0197-6. PubMed: 20730631. [DOI] [PubMed] [Google Scholar]
- 28. Jaleel CA, Manivannan P, Sankar B, Kishorekumar A, Gopi R et al. (2007) Water deficit stress mitigation by calcium chloride in Catharanthus roseus: Effects on oxidative stress, proline metabolism and indole alkaloid accumulation. Colloids Surf B 60: 110-116. doi:10.1016/j.colsurfb.2007.06.006. [DOI] [PubMed] [Google Scholar]
- 29. Cousson A (2009) Involvement of phospholipase C-independent calcium-mediated abscisic acid signaling during Arabidopsis response to drought. Biol Plant 53: 53-62. doi:10.1007/s10535-009-0008-0. [Google Scholar]
- 30. Mao PS, Wang XG, Wang YH, Han JG (2009) Effect of storage temperature and duration on the vigor of zoysiagrass (Zoysia japonica Steud.) seed harvested at different maturity stages. Grassland Sci 55: 1-5. doi:10.1111/j.1744-697X.2009.00129.x. [Google Scholar]
- 31. Liang X, Su DR, Yin SX, Wang Z (2009) Leaf water absorption and desorption functions for three turfgrasses. J Hydrol 376: 243-248. doi:10.1016/j.jhydrol.2009.07.035. [Google Scholar]
- 32. Huang BR (1999) Water relations and root activities of Buchloe dactyloides and Zoysia japonica in response to localized soil drying. Plant Soil 208: 179-186. doi:10.1023/A:1004472314631. [Google Scholar]
- 33. Du H, Wang Z, Huang B (2008) Differential responses of tall fescue and zoysiagrass to heat and drought stress. Acta Hort (ISHS) 783: 207-214. [Google Scholar]
- 34. Agrawal SB, Rathore D (2007) Changes in oxidative stress defense in wheat (Triticum aestivum L.) and mung bean (Vigna radiata L.) cultivars grown with and without mineral nutrients and irradiated by supplemental ultraviolet-B. Environ Exp Bot 59: 21-33. doi:10.1016/j.envexpbot.2005.09.009. [Google Scholar]
- 35. Beyer WF Jr, Fridovich I (1987) Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Anal Biochem 161: 559-566. doi:10.1016/0003-2697(87)90489-1. PubMed: 3034103. [DOI] [PubMed] [Google Scholar]
- 36. Chance M, Maehly AC (1955) Assay of catalases and peroxidases. Methods Enzymol 2: 764-775. doi:10.1016/S0076-6879(55)02300-8. [DOI] [PubMed] [Google Scholar]
- 37. Fu J, Huang B (2001) Involvement of antioxidants and lipid peroxidant in the adaptation of two cool-season grasses to localized drought stress. Environ Exp Bot 45: 105-114. doi:10.1016/S0098-8472(00)00084-8. PubMed: 11275219. [DOI] [PubMed] [Google Scholar]
- 38. Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39: 205-207. doi:10.1007/BF00018060. [Google Scholar]
- 39. Arshi A, Abdin MZ, Iqbal M (2006) Effect of CaCl2 on growth performance, photosynthetic efficiency and nitrogen assimilation of Cichorium intybus L. grown under NaCl stress. Acta Physiol Plants 28: 137-147. doi:10.1007/s11738-006-0040-z. [Google Scholar]
- 40. Tattini M, Traversi ML (2009) On the mechanism of salt tolerance in olive (Olea europaea L.) under low- or high-Ca2+ supply. Environ Exp Bot 65: 72-81. doi:10.1016/j.envexpbot.2008.01.005. [Google Scholar]
- 41. Shoresh M, Spivak M, Bernstein N (2011) Involvement of calcium-mediated effects on ROS metabolism in the regulation of growth improvement under salinity. Free Radic Biol Med 51: 1221-1234. doi:10.1016/j.freeradbiomed.2011.03.036. PubMed: 21466848. [DOI] [PubMed] [Google Scholar]
- 42. Nayyar H, Kaushal SK (2002) Alleviation of negative effects of water stress in two contrasting wheat genotypes by calcium and abscisic acid. Biol Plant 45: 65-70. doi:10.1023/A:1015132019686. [Google Scholar]
- 43. Guimarães FVA, de Lacerda CF, Marques EC, de Miranda MRA, de Abreu CEB et al. (2011) Calcium can moderate changes on membrane structure and lipid composition in cowpea plants under salt stress. Plant Growth Regul 65: 55-63. doi:10.1007/s10725-011-9574-1. [Google Scholar]
- 44. Levitt J (1980) Responses of plants to environmental stresses. New York: Academic Press. [Google Scholar]
- 45. Israr M, Jewell A, Kumar D, Sahi SV (2011) Interactive effects of lead, copper, nickel and zinc on growth, metal uptake and antioxidative metabolism of Sesbania drummondii . J Hazard Mater 186: 1520-1526. doi:10.1016/j.jhazmat.2010.12.021. PubMed: 21216094. [DOI] [PubMed] [Google Scholar]
- 46. Willekens H, Chamnongpol S, Davey M, Schraudner M, Langebartels C et al. (1997) Catalase is a sink for H2O2 and is indispensable for stress defense in C3 plants. EMBO J 16: 4806-4816. doi:10.1093/emboj/16.16.4806. PubMed: 9305623. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Issam N, Kawther M, Haythem M, Moez J (2012) Effects of CaCl2 pretreatment on antioxidant enzyme and leaf lipid content of faba bean (Vicia faba L.) seedlings under cadmium stress. Plant Growth Regul 68: 37-47. doi:10.1007/s10725-012-9691-5. [Google Scholar]
- 48. Dobrá J, Vanková R, Havlová M, Burman AJ, Libus J et al. (2011) Tobacco leaves and roots differ in the expression of proline metabolism-related genes in the course of drought stress and subsequent recovery. J Plant Physiol 168: 1588-1597. doi:10.1016/j.jplph.2011.02.009. PubMed: 21481968. [DOI] [PubMed] [Google Scholar]
- 49. Ku HM, Tan CW, Su YS, Cy Chiu, Chen CT et al. (2012) The effect of water deficit and excess copper on proline metabolism in Nicotiana benthamiana . Biol Plant 56: 337-343. doi:10.1007/s10535-012-0095-1. [Google Scholar]