

Abstract
Study Objectives:
Narcolepsy is caused by selective loss of the orexin/hypocretin-producing neurons of the hypothalamus. For patients with narcolepsy, chronic sleepiness is often the most disabling symptom, but current therapies rarely normalize alertness and do not address the underlying orexin deficiency. We hypothesized that the sleepiness of narcolepsy would substantially improve if orexin signaling were restored in specific brain regions at appropriate times of day.
Design:
We used gene therapy to restore orexin signaling in a mouse model of narcolepsy. In these Atx mice, expression of a toxic protein (ataxin-3) selectively kills the orexin neurons.
Interventions:
To induce ectopic expression of the orexin neuropeptides, we microinjected an adeno-associated viral vector coding for prepro-orexin plus a red fluorescence protein (AAV-orexin) into the mediobasal hypothalamus of Atx and wild-type mice. Control mice received an AAV coding only for red fluorescence protein. Two weeks later, we recorded sleep/wake behavior, locomotor activity, and body temperature and examined the patterns of orexin expression.
Measurements and Results:
Atx mice rescued with AAV-orexin produced long bouts of wakefulness and had a normal diurnal pattern of arousal, with the longest bouts of wake and the highest amounts of locomotor activity in the first hours of the night. In addition, AAV-orexin improved the timing of rapid eye movement sleep and the consolidation of nonrapid eye movement sleep in Atx mice.
Conclusions:
These substantial improvements in sleepiness and other symptoms of narcolepsy demonstrate the effectiveness of orexin gene therapy in a mouse model of narcolepsy. Additional work is needed to optimize this approach, but in time, AAV-orexin could become a useful therapeutic option for patients with narcolepsy.
Citation:
Kantor S; Mochizuki T; Lops SN; Ko B; Clain E; Clark E; Yamamoto M; Scammell TE. Orexin gene therapy restores the timing and maintenance of wakefulness in narcoleptic mice. SLEEP 2013;36(8):1129–1138.
Keywords: AAV-orexin, ataxin-3, cataplexy, gene therapy, hypothalamus, locomotor activity, narcolepsy, wake maintenance
INTRODUCTION
Narcolepsy is characterized by daytime sleepiness and abnormal manifestations of rapid eye movement (REM) sleep, including cataplexy, a pathological intrusion of REM sleep atonia into wakefulness.1 In most people with narcolepsy, chronic sleepiness is the most disruptive symptom; these individuals often struggle to remain awake, especially under sedentary conditions during school and work,2 and they have a threefold to fourfold increased risk of motor vehicle accidents, probably from lapses in attention and dozing off.3–5 Narcolepsy typically begins in adolescence or young adulthood, and the proper management of such patients usually requires lifelong treatment with medications.6
During the past decade, researchers discovered that narcolepsy is caused by selective loss of the hypothalamic neurons producing the orexin/hypocretin neuropeptides,7–9 and this crucial observation has led to great progress in understanding the neurobiology of narcolepsy. During the active period, the orexin peptides (orexin-A and -B) increase the activity of neurons in many brain regions that promote wakefulness and regulate REM sleep.10–12 In mouse models of narcolepsy, a lack of orexin signaling results in low levels of arousal during the active period with much shorter bouts of wakefulness, reduced locomotor activity, and cataplexy.13–15 Most likely, in the absence of orexins, the activities of wake-promoting and REM sleep-regulating brain regions are unstable, resulting in frequent transitions into sleep and intrusions of cataplexy.16
These discoveries have shed much light on the neurobiology of narcolepsy, but they have not yet translated into better clinical therapies. For example, excessive daytime sleepiness is often the most disabling symptom for patients with narcolepsy, but even with optimal treatment, few patients achieve fully normal levels of alertness.17 Clinical improvements are suboptimal most likely because these medications simply address the symptoms but not the underlying orexin deficiency. The ideal therapy for narcolepsy would restore orexin signaling in wake-promoting brain regions during the active period.
We hypothesized that the sleepiness of narcolepsy would substantially improve with orexin gene therapy. Thus, to induce robust ectopic expression of the orexin peptides, we microinjected an adeno-associated viral vector coding for the orexins (AAV-orexin) into mice with degeneration of the orexin neurons. Our main goal was to determine whether AAV-orexin could restore the ability of these mice to maintain long periods of wakefulness. In addition, we examined if restoration of orexin signaling could promote wakefulness at the correct times of day and if it could rescue other aspects of the narcolepsy phenotype such as REM sleep abnormalities and cataplexy.
MATERIALS AND METHODS
We carried out these experiments in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (National Institutes of Health Publication 8023, revised 1978). These studies were approved by the Institutional Animal Care and Use Committees of Beth Israel Deaconess Medical Center and Harvard Medical School.
Construction of Vector
We used a recombinant AAV produced by the Harvard Gene Therapy Initiative. We chose to use an AAV because they have high transduction efficiency in neurons, produce almost no inflammation, and produce long-lasting gene expression.18,19 These desirable properties have made AAV vectors very popular for genetic rescue experiments in mice and in humans.20–22
The experimental AAV (serotype 8) contained the genes coding for prepro-orexin and mCherry (a red fluorescence protein), linked by an internal ribosome entry site sequence. The control vector contained only the mCherry sequence. The AAV were assembled on a pAAV-MCS8 backbone with a constitutively active cytomegalovirus promoter followed by a human beta-globin intron 2 sequence that drives expression of the prepro-orexin and mCherry genes. These genes were followed by a polyadenylation sequence. The vectors were generated using tripartite transfection (AAV-rep/cap expression plasmid, adenovirus miniplasmid, and AAV vector plasmid) into 293A cells, followed by purification by iodixanol gradient and Q sepharose column chromatography, dialyzation against phopshate buffered saline, concentration by Amicon spin column (Merck Millipore, Billerica, MA), and titration by dot blot hybridization. The final titers were ∼4 × 1012 pfu/mL.
Animals
As a mouse model of narcolepsy, we used transgenic (Atx) mice in which the human orexin promoter drives the expression of a mutant form of ataxin-3, a toxic protein that selectively kills nearly all the orexin neurons by 8-12 weeks of age.14 This selective cell loss closely mirrors the loss of the orexin neurons in people with narcolepsy, and it provides a rigorous test of whether expression of orexin peptides in “non-orexin” neurons can rescue the narcolepsy phenotype. These Atx mice were a gift from T. Sakurai at Kanazawa University. To maximize genetic homogeneity, we backcrossed the Atx line with C57BL/6J mice for six to eight generations.
We examined the effects of AAV gene therapy in four groups of male mice (age 15-19 weeks; 25-35 g): Atx mice rescued with injection of AAV-orexin (n = 10); an unrescued control group of Atx mice injected with AAV-mCherry (n = 9); a normal control group of wild-type littermates (WT) injected with AAV-mCherry (n = 10); and a group of WT mice injected with AAV-orexin (n = 7).
We identified mice using polymerase chain reaction (PCR) genotyping of tail biopsies. Primers used were 5′-CAT GAA GGA AGA AGG TCC TGG and 3′-CCT TGC ACC CAG GAA TCT GG against the orexin/ataxin-3 transgene.
Surgery and Protocol
We anesthetized mice with ketamine-xylazine (100 and 10 mg/kg, intraperitoneally) and placed them in a stereotaxic apparatus. To induce orexin expression in a large number of neurons, we microinjected 50 nl of AAV-orexin or AAV-mCherry unilaterally over 5 min. Coordinates for the injections were: AP −1.82 mm, DV −5.3 mm, RL + 0.5 mm, a region just below the dorsomedial nucleus of the hypothalamus (DMH). We targeted this area because previous studies indicated that when driven by a nonspecific promoter, many neurons in the ventral hypothalamus robustly produce orexin-A.23
After injection of AAV, we implanted each mouse with electroencephalogram (EEG) and electromyogram (EMG) electrodes, as described previously.24 In brief, we placed stainless steel screw electrodes epidurally over the left frontal cortex (1.5 mm lateral and 1 mm anterior to bregma) and left parietal cortex (1.5 mm lateral and 1.0 mm anterior to lambda). EMG signals were acquired by a pair of stainless steel spring wires (Plastics One Inc., Roanoke, VA) inserted into the neck extensor muscles. We placed a telemetry transmitter (TA-F20, Data Sciences International, St. Paul, MN) into the peritoneal cavity to record body temperature and locomotor activity (LMA). These transmitters were factory-calibrated to an accuracy of 0.1°C. The signal from the telemetry transmitter was received and digitally acquired by an antenna located below the recording cage (RPC-1 and Data-quest, Data Sciences International, St. Paul, MN).
Recording and Analysis of Sleep
After surgery, we housed mice in individual recording cages with food and water available ad libitum. After a recovery period of 10 days, we connected the mice to recording cables within a sound-attenuated chamber with a 12:12 h light-dark (LD) cycle (30 lux daylight-type fluorescent tubes with lights on at 07:00) and constant temperature (23 ± 1°C). The recording cable was attached to a low torque electrical swivel above the cage that allowed free movement. After mice acclimated to the recording environment for 4 days, we recorded EEG, EMG, infrared video, body temperature, and LMA for 24 h. We recorded LMA and body temperature in 5 min bins.
The EEG/EMG signals were acquired using Grass Model 12 amplifiers (West Warwick, RI), digitized at 128 Hz, digitally filtered (EEG: 0.3-30 Hz, EMG: 5-60 Hz), and semi-automatically scored as wake, nonrapid eye movement (NREM) sleep, rapid eye movement (REM) sleep, or cataplexy in 10 s epochs using SleepSign (Kissei Comtec, Matsumoto, Japan). S.K. then visually inspected this preliminary scoring and made corrections when appropriate.
A behavior was scored as cataplexy at any time during a 24 h period if it fulfilled certain criteria.25 Specifically, we scored an epoch as cataplexy when: (1) the mouse had one or more epochs of EEG theta activity and neck muscle atonia immediately preceded and followed by active wake; (2) at least 40 s of wake preceded cataplexy to exclude any REM sleep that might follow a brief awakening26; and (3) the animal was immobile during the episode. Whenever behavior met criterion 1, we examined integrated infrared video recordings (SleepSign) to determine if all criteria were fulfilled.24,25 No specific triggers of cataplexy were used in this study.
Immunostaining and Cell Counts
After polysomnographic recordings, we deeply anesthetized the mice with ketamine-xylazine (150 and 15 mg/kg, intraperitoneally) and perfused them transcardially with 50-100 ml 0.9% saline followed by 50-100 ml of neutral buffered 10% formalin (pH 7.0; Fisher, Agawam, MA). We fixed the brains overnight in formalin and cryoprotected them in 20% sucrose with 0.02% sodium azide (Sigma-Aldrich, St. Louis, MO). Afterward, we sectioned brains at 30 μm into a 1:3 series on a freezing microtome.
To confirm that AAV-orexin induced the expression of both orexin-A and orexin-B peptides, we immunostained brain sec-tions as described previously.27 In brief, after three 5 min rinses in PBS, we treated the sections with PBT (phosphate buffered saline, pH 7.4, with 0.25% Triton X-100) containing 0.3% hydrogen peroxide for 30 min. Following three more 5 min rinses in PBS, we incubated sections overnight in goat antiorexin-A or antiorexin-B antisera (1:10,000 and 1:500, respectively; Santa Cruz Biotechnology, Santa Cruz, CA). After six 5 min washes in PBS, we incubated sections for 2 h in biotinylated secondary antiserum (donkey antigoat immunoglobulin G (IgG) (H + L), 1:500; Jackson ImmunoResearch, West Grove, PA). After three 10 min rinses in PBS, we incubated sections with avidin-biotin complex (Vectastain Elite ABC Kit, Vector Labs, Burlingame, CA) for 1 h followed by four 5 min rinses in PBS. We then visualized orexin-immunoreactive (IR) neurons by reaction with 3,3′-diaminobenzidine (DAB; SK-4100; Vector Labs) for 3-5 min. For cell counts and mapping of AAV-orexin injection sites, we immunostained one series of sections for orexin-B. As a control, we mapped AAV-mCherry injection sites using immunostaining for dsRed (rabbit anti-dsRed antiserum, 1:5,000, Clontech, Mountain View, CA; donkey antirabbit, 1:500, Jackson ImmunoResearch) and DAB.
Atx mice injected with the control vector (AAV-mCherry) had approximately 80% fewer orexin-B IR neurons than their WT littermates injected with the same AAV (P < 0.001) (Figure 1). This is slightly less than the 95% cell loss described in other studies,14,24,28 perhaps because we used younger mice and possibly more sensitive antisera. All the remaining orexin-IR cells in the Atx mice showed highly abnormal morphology, with small and fragmented cell bodies, weakly IR soma, and much less orexin immunoreactivity in axons and terminal fields.
Figure 1.

Distributions of orexin immunoreactive neurons. A, The orexin field in a wild-type (WT) mouse injected with AAV-mCherry shows the normal, moderately dense cluster of orexin-immunoreactive (IR) neurons above the fornix (f) and in the lateral hypothalamus (LH). B, An Atx mouse injected with AAV-mCherry has a few scattered orexin-IR neurons in this region, but the cells are small and weakly IR and axonal labeling is nearly absent. C and D, An AAV-orexin injection site in an Atx mouse shows many neurons expressing orexin in the posterior hypothalamus (PH), dorsomedial nucleus of the hypothalamus (DMH), tuberomammillary nucleus (TMN), and other parts of the mammillary region. Axons from these neurons course around the mammillothalamic tract (mt) and densely innervate the amygdalohippocampal area (AHiA) plus other sites. AAV, adeno-associated viral vector; Atx, transgenic (ataxin); opt, optic tract; ic, internal capsule; 3V, third ventricle; cp, cerebral peduncle.
For fluorescent double staining of mCherry and orexin-A, we first incubated sections in goat antiorexin-A antiserum, followed by Alexa Fluor 488 secondary antiserum (donkey antigoat, 1:500; Invitrogen, Grand Island, NY). The next day, we incubated sections in rabbit anti-dsRed antiserum (1:2,000) and then in Alexa Fluor 555 secondary antiserum (donkey anti-rabbit, 1:500; Invitrogen).
The orexin-A and -B antisera produced no immunolabeling in brain sections from prepro-orexin knockout mice, and the dsRed antisera produced no labeling in sections from uninjected WT mice.
Data Analysis and Statistics
We analyzed the duration of bouts, number of bouts, percentage of time spent in each behavioral state, and LMA. We used two-way analysis of variance (ANOVA) with repeated measures to analyze changes in each vigilance state and LMA as a function of time and group (WT AAV-mCherry, WT AAV-orexin, Atx AAV-mCherry, and Atx AAV-orexin). We used the same test with two main factors (group and day-night cycle) to compare vigilance state parameters and LMA. The Bonferroni test was used for post hoc comparisons.
We examined the ability of mice to maintain wakefulness by analyzing the distribution of wake as a function of bout length. We separated all wake bouts into eight bins according to length (< 30, 40-70, 80-150, 160-310, 320-630, 640-1270, 1280-2550, and > 2550 sec).29–31 The percentage of wake occurring in each bin was used to produce a time-weighted frequency histogram. The primary endpoint was to compare the amounts of wake occurring in bouts of various lengths between AAV-orexin or AAV-mCherry treated WT and Atx mice during the dark period. The results were compared by ANOVA for repeated measures followed by Bonferroni post hoc test.
To examine the diurnal variations in the longest wake bout lengths in mice, we calculated the longest duration of discrete bouts of wakefulness across 24 h.32 A wake bout was defined as two or more consecutive epochs of polygraphically defined wakefulness. We identified the maximum length bout within a 1 h window moved stepwise in 2 min increments through the sleep-wake time series of each animal. In each step, the longest wake bout within the window was identified and recorded. If the length of a wake bout exceeded the size of the moving window the entire length of the bout was used. We averaged bout lengths at corresponding times across mice and plotted them as a function of clock time (means ± standard error of the mean). We then compared results across groups using ANOVA with repeated measures and the Bonferroni post hoc test.
To measure the diurnal variations in sleep/wakefulness and LMA we used the nocturnality ratio.24 Nocturnality ratios were defined as the percentage of total amount of a behavior (wake, NREM sleep, REM sleep, or LMA) occurring during the night. Thus, high nocturnality ratios represent high incidence of the behavior during the dark period, whereas ratios close to 50% indicate behavior evenly distributed between day and night. We compared the results across groups using one-way ANOVA followed by the Bonferroni post hoc test.
RESULTS
AAV-Orexin Induced Robust Ectopic Expression of the Orexin Peptides
We microinjected an AAV containing the prepro-orexin and mCherry genes (AAV-orexin), or a control AAV containing only the mCherry gene (AAV-mCherry) into the mediobasal hypothalamus of Atx and WT mice. In Atx mice injected with AAV-orexin, immunostaining showed robust expression of orexin-A and -B (Figure 1). Double fluorescent labeling demonstrated that more than 98% of orexin-A IR neurons at the injection sites also contained mCherry. These observations demonstrate that AAV-orexin induces strong ectopic expression of both orexin peptides and mCherry.
AAV injection sites extended from the dorsomedial nucleus of the hypothalamus (DMH) and lateral hypothalamus (LH), caudally into the ventral parts of the posterior hypothalamus (PH), and ventrally into the arcuate nucleus, retrochiasmatic region, supramammillary nuclei, and tuberomammillary nucleus (TMN) (Figure 2). Interestingly, neurons in the ventromedial nucleus of the hypothalamus often expressed mCherry but rarely contained orexin immunoreactivity, suggesting that unlike many other hypothalamic neurons, these cells lack key factors for producing the orexin peptides. The distributions of injection sites were similar in the other groups that received AAV-orexin and AAV-mCherry, though there was some scatter within groups (data not shown). These microinjections of AAV-orexin resulted in a similar number of orexin-IR neurons (1,000-1,400 per mouse) across the mediobasal hypothalamus in each Atx mice.
Figure 2.
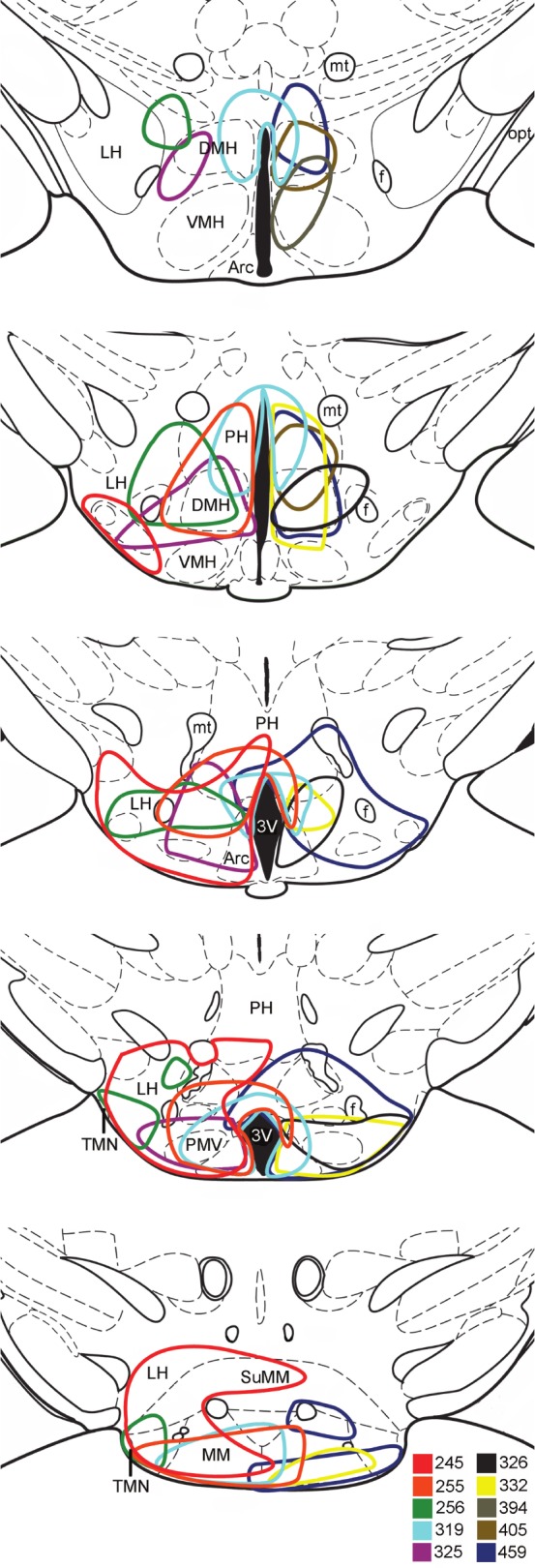
Distribution of AAV-orexin injection sites in Atx mice. Each mouse received a unilateral injection into the right side of the hypothala-mus, but injection sites are depicted on either side for clarity. The drawn injection site boundaries encompass approximately 90% of the orexin-immunoreactive neurons. AAV, adeno-associated viral vector; Atx, transgenic (ataxin); DMH, dorsomedial nucleus; Arc, arcuate nucleus; PMV, ventral premammillary nucleus; SuMM, medial supramammillary nucleus; LH, lateral hypothalamus; MM, medial mammillary nucleus; mt, mammillothalamic tract; 3V, third ventricle; TMN, tuberomammillary nucleus. Injected sites are plotted on drawings adapted from Paxinos and Franklin.50
Orexin-IR fibers projected from the injection sites to many nuclei throughout the brain, including the anterior paraventricular thalamic nuclei, basal forebrain, bed nucleus of the stria terminalis, DMH, PH, supramammillary nucleus, and TMN. The orexin fiber projections varied across animals because their injection sites were slightly different. The best improvements in maintenance of wakefulness occurred in Atx mice with dense orexin-IR innervation of the bed nucleus of the stria terminalis, preoptic area, anterior thalamus, arcuate nucleus, supramammillary nucleus, and TMN.
Expression of Orexins in the Hypothalamus Restored the Timing and Maintenance of Wakefulness
Mice treated with AAV-orexin continued to have normal diurnal variations in sleep/wake behavior (Figure 3). In Atx mice, AAV-orexin increased the amount of wake by 13% (F3,32 = 9.93, P < 0.01) and reduced the amount of NREM sleep during the dark period (Table 1). Although WT mice showed similar shifts, the changes were not statistically significant. AAV-orexin did not alter the amounts of sleep/wake stages in the light period.
Figure 3.
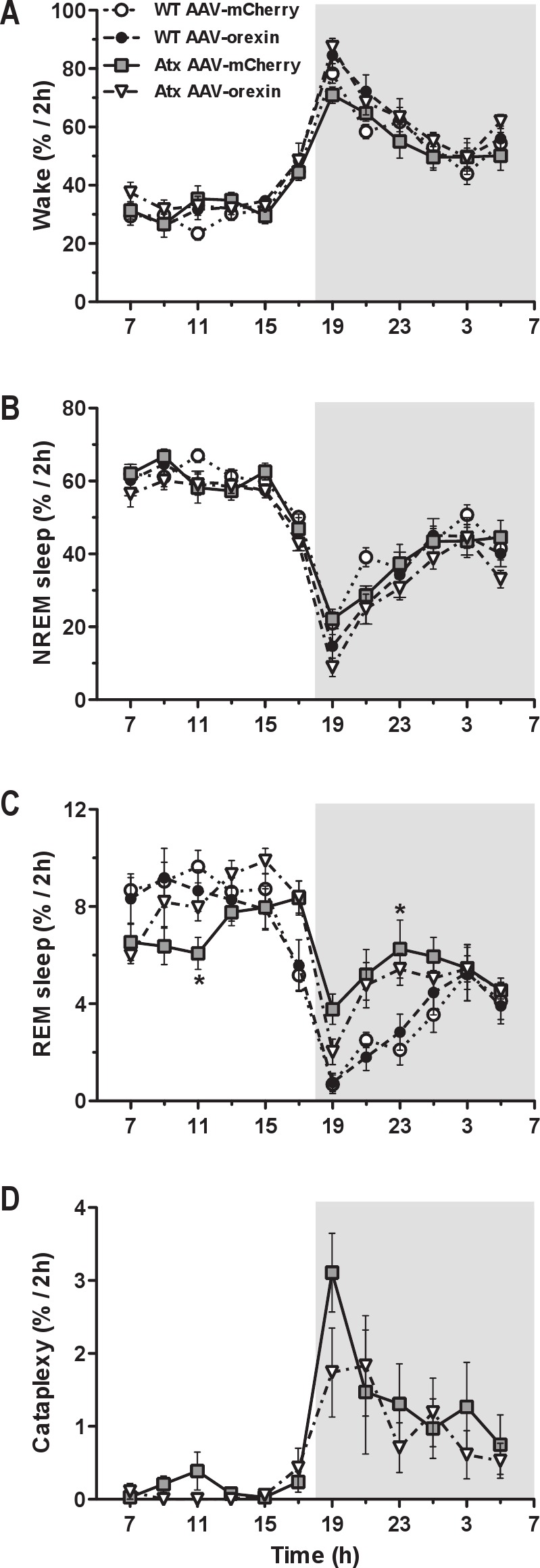
Diurnal variations in sleep/wake states. A and B, The diurnal pattern of wake and NREM sleep was almost identical in WT and Atx mice irrespective of the treatment (AAV-mCherry or AAV-orexin). C, However, Atx mice treated with AAV-mCherry had much less circadian variation in REM sleep than their WT littermates due to an increase in their REM sleep amount during the dark period. D, Injections of AAV-orexin caused a small and statistically insignifi cant decrease in cataplexy after dark onset in Atx mice. There were no changes in the amounts of sleep-wake states in WT mice treated with AAV-orexin. Data are shown in 2 h intervals. *P < 0.01 compared to WT AAV-mCherry mice. AAV, adeno-associated viral vector; Atx, transgenic (ataxin); NREM, nonrapid eye movement; REM, rapid eye movement; WT, wild type.
Table 1.
Vigilance state parameters in WT and Atx mice microinjected with AAV-mCherry or AAV-orexin
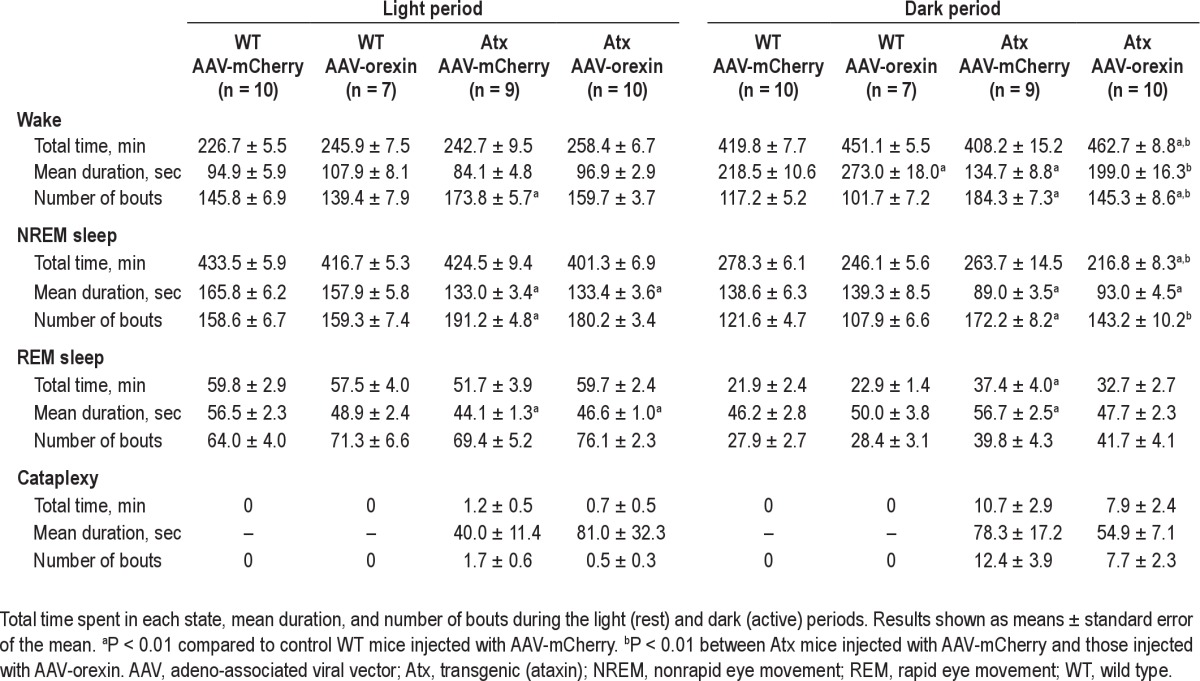
During the dark period, unrescued Atx mice injected with AAV-mCherry had wake bouts much shorter than those seen in control WT mice. In contrast, the mean duration of wake bouts in AAV-orexin-injected Atx mice was 48% longer and nearly the same length as in WT mice injected with AAV-mCherry (F3,32 = 12.63, P < 0.01). In addition, Atx mice treated with AAV-orexin had more consolidated wakefulness as evidenced by 21% fewer wake bouts (F3,32 = 6.99, P < 0.01). Thus, we can conclude that AAV-orexin markedly improved the consolidation of wakefulness in Atx mice.
AAV-orexin had similar but smaller effects in WT mice. Compared with WT mice injected with AAV-mCherry, WT mice treated with AAV-orexin had 25% longer wake bouts during the dark period (F3,32 = 12.63, P < 0.01). During the light period, AAV-orexin did not alter the number or duration of wake bouts in Atx and WT mice.
To better understand the ability of mice to maintain wakefulness, we analyzed wake as a function of bout length. During the dark period, most wakefulness in WT mice occurs in very long wake bouts, but unrescued Atx mice spent very little time in these long wake bouts and instead most of their wake occurred in midlength bouts (Figure 4). Specifi cally, in WT mice injected with AAV-mCherry, 33% of wake occurred in bouts lasting more than 2,550 s (42.5 min). In contrast, Atx mice injected with AAV-mCherry spent only 4% of their wake in such long bouts. However, Atx mice rescued with AAV-orexin spent 20% of their time awake in these long bouts (F21,224 = 10.08, P < 0.01). This observation further demonstrates that AAV-orexin markedly improved the maintenance of wakefulness in Atx mice.
Figure 4.
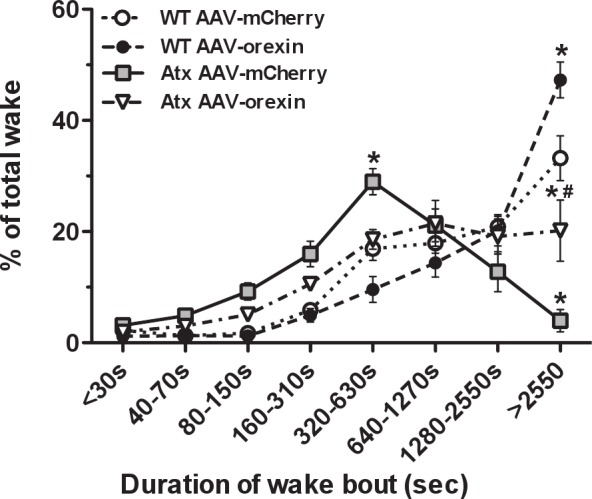
AAV-orexin improved maintenance of wakefulness. Time-weighted frequency histograms of wake bouts during the dark period show that in WT mice treated with AAV-mCherry or AAV-orexin, 33-47% of wakefulness occurred in bouts longer than 2,550 s (42.5 min). In contrast, most wakefulness in Atx mice treated with AAV-mCherry occurred in bouts lasting only 320-630 s (5.3-10.5 min) with almost no wakefulness occurring in very long bouts. In Atx mice treated with AAV-orexin, 20% of all wake occurred in very long wake bouts. *P < 0.01 compared to WT AAV-mCherry mice. #P < 0.01 between Atx AAV-mCherry and Atx AAV-orexin mice. AAV, adeno-associated viral vector; Atx, transgenic (ataxin); WT, wild type.
Mice normally produce their longest bouts of wakefulness in the fi rst few hours of the dark period when feeding and locomotor activity are most abundant.15,33,34 To determine if AAV-orexin promotes long bouts of wake at the correct times of day, we examined the longest wake bouts across time. WT mice injected with AAV-mCherry or AAV-orexin had very long wake bouts in the fi rst hours of the dark period, but Atx mice injected with AAV-mCherry showed no lengthening of wake bouts, even just after dark onset (Figure 5). In contrast, injection of AAV-orexin restored the normal diurnal pattern of wakefulness in Atx mice by producing very long wake bouts in the fi rst 2 h of the dark period in a pattern very similar to that of WT mice (F2157,23008 = 1.55, P < 0.01).
Figure 5.

AAV orexin rescued the timing of wakefulness. A, WT mice injected with AAV-mCherry produce very long wake bouts in the fi rst 2 h of the dark period. B, A similar pattern is seen in WT mice injected with AAV-orexin though these very long wake bouts persist for about 4 h. C, Unrescued Atx mice injected with AAV-mCherry have short wake bouts at all times, even at dark onset. D, In contrast, Atx mice injected with AAV-orexin produce long wake bouts in the fi rst 2 h of the dark period, much like normal WT mice. Maximum wake bouts were calculated using a 1 h-wide moving window as described in the Methods section. Data are shown as group mean (solid lines) ± standard error of the mean (dashed lines). The triangles represent signifi cant differences from WT mice injected with AAV-mCherry, whereas the vertical bars denote signifi cant differences between AAV-orexin and AAV-mCherry treated Atx mice after Bonferroni post hoc tests (P < 0.05). AAV, adeno-associated viral vector; Atx, transgenic (ataxin); WT, wild type.
Expression of Orexins in the Hypothalamus Improved Sleep and Locomotor Activity, but Had Little Effect on Cataplexy
In addition to poor maintenance of wakefulness, Atx mice, just like people with narcolepsy, also have fragmented sleep, poor circadian control of REM sleep, and cataplexy.14,23,24,35,36 In Atx mice treated with AAV-orexin, NREM sleep bouts were still shorter than in WT mice (F3,32 = 25.56, P < 0.01; Table 1), but they had fewer NREM sleep bouts (F3,32 = 18.14, P < 0.01; Table 1), providing evidence of less fragmented sleep.
AAV-orexin also improved the diurnal pattern of REM sleep in Atx mice. REM sleep is under strong circadian control, and normally, most REM sleep occurs during the inactive period.37–39 Compared with WT mice, unrescued Atx mice had longer REM sleep bouts and more REM sleep during the night (F3,32 = 11.35, P < 0.01 and F3,32 = 8.71, P < 0.01, respectively; Table 1 and Figure 3). Moreover, almost half (42%) of their total daily REM sleep occurred during the dark period: a considerably higher proportion than the 26% seen in control WT mice treated with AAV-mCherry (F3,32 = 9.66, P < 0.01; Figure 6). Injection of AAV-orexin into Atx mice reduced the proportion of REM sleep in the dark period to 35%, an amount closer to that of WT mice, further supporting the role of orexins in the circadian control of REM sleep.24
Figure 6.
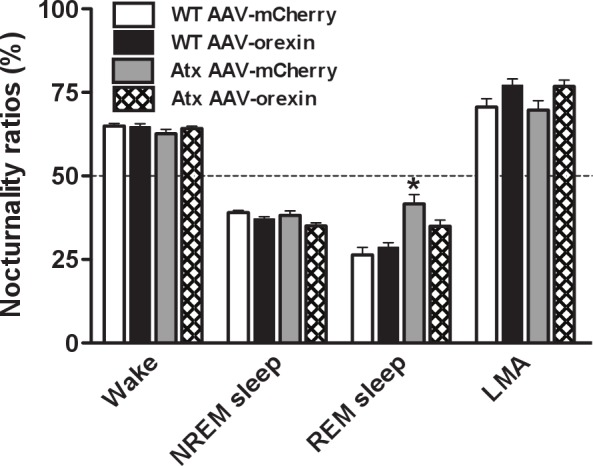
AAV-orexin improved the diurnal rhythm of REM sleep. Nocturnality ratios (amount of a state occurring during the dark period / total amount in 24 h) show normal patterns for wake and NREM sleep in all groups. Atx mice treated with AAV-mCherry have an excessive proportion of REM sleep in the dark period, indicated by a nocturnality ratio close to 50%, but Atx mice injected with AAV-orexin show normal proportions. *P < 0.01 compared to WT mice injected with AAV-mCherry. AAV, adeno-associated viral vector; Atx, transgenic (ataxin); NREM, nonrapid eye movement; REM, rapid eye movement; WT, wild type.
In contrast to a prior study that injected AAV-orexin into the zona incerta,28 our more ventral injections of AAV-orexin did not reduce cataplexy (Table 1). The frequency and the total amount of cataplexy appeared less, but the reductions were not statistically significant.
In Atx mice, AAV-orexin increased locomotion in the fi rst 2 h of the dark period (F33,352 = 1.83, P < 0.01; Figure 7A), and it increased the total amount of LMA during the dark period (F3,32 = 3.13, P < 0.05; Figure 7B). The diurnal patterns of body temperature did not differ between groups (data not shown).
Figure 7.
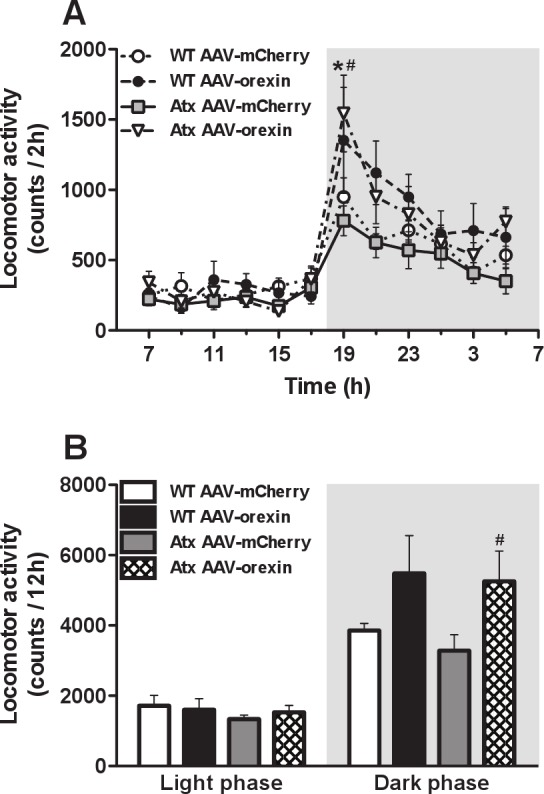
AAV-orexin increased locomotor activity (LMA) during the dark period. A, The general timing and the amounts of LMA was similar in WT and Atx mice treated with AAV-mCherry. B, Injections of AAV-orexin strongly increased LMA in Atx mice during the dark period. WT mice injected with AAV-orexin showed a less striking increase that did not reach statistical signifi cance. *P < 0.05 compared to WT mice injected with AAV-mCherry. #P < 0.05 between Atx mice injected with AAV-mCherry and AAV-orexin. AAV, adeno-associated viral vector; Atx, transgenic (ataxin); WT, wild type.
DISCUSSION
We found that orexin gene therapy substantially improved the sleepiness of mice with narcolepsy. Hypothalamic expression of the orexin peptides in mice lacking the orexin neurons markedly increased their ability to maintain long bouts of wakefulness. In addition, mice rescued with AAV-orexin had a normal diurnal pattern of arousal, with the longest bouts of wake and the highest amounts of locomotor activity in the fi rst hours of the dark period, just as in WT mice. Although the expression of orexins in the hypothalamus had little effect on cataplexy, both NREM and REM sleep improved in orexin neuron-deficient mice. These observations provide strong evidence that orexin gene therapy considerably improved the symptoms of narcolepsy in mice, even when the orexin peptides were produced by a variety of “nonorexin” hypothalamic neurons.
AAV-Orexin Improved Wakefulness
Sleepiness is the most disabling symptom for most people with narcolepsy, and a key goal when treating narcolepsy is to achieve normal alertness during the waking hours. Several studies have shown that intracerebroventricular administration of orexin peptides transiently increases wakefulness in mice with narcolepsy and healthy rodents, but the amounts of wake-fulness are often much higher than normal and ventricular injections are impractical in people.23,40,41 Restoration of orexin signaling using viral vectors is a more promising approach, but prior studies have shown no improvements in wakefulness even though cataplexy was improved.28,42
In our study, orexin gene therapy improved many behaviors that likely refl ect the excessive sleepiness of narcolepsy. Atx mice injected with AAV-orexin had more consolidated wake-fulness, with more time awake and longer wake bouts during the dark period. This response is similar to that seen in Atx mice treated with a very high dose of modafi nil (200 mg/kg).43 In addition, we found that expression of orexins in the mediobasal hypothalamus produces a surprisingly normal diurnal pattern of wakefulness in Atx mice. Similar to WT mice, Atx mice injected with AAV-orexin had their highest levels of arousal in the fi rst few hours of the dark period, as shown by very long wake bouts and high levels of locomotor activity during this time.
Several factors may explain why AAV-orexin increased arousal at the beginning of the active period. Normally, extracellular levels of orexins vary in a circadian pattern, with high levels during the active period and low levels during the rest period.44 Our AAV used a constitutively active promoter to drive production of prepro-orexin messenger RNA, so restoration of this normal pattern of arousal suggests that the timing of orexin peptide release is more important than the timing of orexin synthesis. Perhaps “nonorexin” hypothalamic neurons transduced with AAV-orexin have patterns of synaptic activity similar to that of the orexin neurons. Indeed, some of the neurons ectopically expressing orexins (e.g., TMN) normally help regulate wakefulness, and other neurons (e.g., arcuate, supramammillary, PH, LH) may release orexins in the course of mediating behaviors such as feeding that occur during wake. Alternatively, ectopic orexins may be released in a disorganized pattern, but they enhance the effects of other wake-promoting mechanisms, amplifying signals that increase arousal and activity during the active period yet having little effect at other times. Future studies will need to defi ne the exact nature of these neurons ectopically expressing orexins and determine whether they produce higher extracellular levels of orexins during the dark period.
In comparison with prior gene transfer studies, injection of AAV-orexin into the brain of Atx mice produced marked improvement in wakefulness in our study. Liu and colleagues28 injected vectors coding for orexins into the zona incerta, LH, or dorsolateral pons of mice with narcolepsy, and though the mice had less cataplexy, they showed little improvement in their ability to maintain wakefulness.,42,45,46 In contrast, we targeted our AAV-orexin injections at more ventral regions because prior research showed that neurons in the mediobasal hypothalamus can produce very high levels of orexin-A when driven by a nonspecific promoter.23 After transduction with AAV-orexin, these neurons may have simply released greater amounts of orexins, or their axons may have been more effective at delivering orexins to key wake-promoting brain regions.
AAV-Orexin in the Hypothalamus Improved Sleep but Not Cataplexy
Ectopic expression of orexins in the hypothalamus also improved sleep in mice with narcolepsy. Unrescued Atx mice injected with AAV-mCherry had fragmented NREM sleep and excessive amounts of REM sleep during the dark period as seen in prior studies.14,23,24,47 In contrast, Atx mice treated with AAV-orexin had more consolidated NREM sleep and a partially improved diurnal pattern of REM sleep. The dark period suppression of REM sleep by orexins reported here is consistent with previous studies in which the prepro-orexin gene was expressed in orexin-neuron ablated Atx mice,23 or overexpressed in mice with intact orexin neurons.46,48 However, mice in these previous studies had widespread ectopic expression of orexins and highly fragmented sleep, especially during the light phase, perhaps as a consequence of excessively high orexin tone.23,46,48 In addition, the partial improvement in REM sleep in our study suggests that targeting AAV-orexin to different brain regions, or restoring the signaling of other molecules normally released by orexin neurons (e.g., dynorphin and/or glutamate) will be necessary to fully correct the REM sleep abnormalities in Atx mice. Still, these findings further support the hypothesis that orexins, by suppressing REM sleep during the active period, play an important role in the circadian control of REM sleep.24
It is very likely that the diverse symptoms of narcolepsy, such as excessive daytime sleepiness and cataplexy, are mediated by different brain pathways. This idea is supported by the finding that drugs with different mechanisms of action are required for the treatment of each symptom.1 Our injections of AAV-orexin into the mediobasal hypothalamus improved the timing and maintenance of wakefulness, sleep fragmentation, and diurnal pattern of REM sleep, but produced smaller reductions in cataplexy than reported in prior gene transfer studies.28,42,45 Orexin gene transfer into the LH or dorsolateral pons of orexin peptide knockout mice improved cataplexy in recent studies, but the usefulness of this approach was limited by the preservation of the “orexin neurons” and the modest or no improvement in wake maintenance.42,45 Microinjections of AAVorexin into the zona incerta or LH of orexin neuron-deficient Atx mice produced similar reductions in cataplexy with no improvements in wakefulness.28 Most likely, these differences are a consequence of targeting different brain regions. It is possible that neurons ectopically expressing orexins in our study more heavily innervate brain sites that promote wakefulness whereas in other studies neurons of the injected area better innervate regions that regulate cataplexy.
Future Directions
Our results demonstrate that injection of AAV-orexin into the mediobasal hypothalamus markedly improved the maintenance of wakefulness in mice lacking the orexin neurons. In principle, AAV-orexin could be developed into a novel and effective therapy for people with narcolepsy. AAV vectors are relatively safe, cannot replicate, and produce almost no inflammatory response. In addition, genes encoded by AAV can be expressed for many years, so AAV-orexin could produce long-lasting improvements in narcolepsy.49 These properties have made AAV vectors very attractive for genetic rescue experiments in rodents and early stage clinical trials of Parkinson disease and other neurologic disorders.20–22 Thus far, clinical applications have focused on neurological disorders with limited life expectancy, but as more is learned about how to optimize the safety and efficacy of AAV-orexin, it may become a potent option for many patients with narcolepsy.
DISCLOSURE STATEMENT
The listed authors independently analyzed all data and wrote the entire manuscript. The authors have indicated no conflicts of interest.
ACKNOWLEDGMENTS
The Atx mice were a gift from Takeshi Sakurai (Kanazawa University) and Masashi Yanagisawa (University of Texas Southwestern). Veronique van der Horst provided helpful advice on mapping AAV injection sites. This research was supported by a gift from Wake Up Narcolepsy, and grants from the NIH (NS055367, HL095491) and the Harvard College Research Program.
REFERENCES
- 1.Nishino S. Clinical and neurobiological aspects of narcolepsy. Sleep Med. 2007;8:373–99. doi: 10.1016/j.sleep.2007.03.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Fronczek R, Middelkoop HA, van Dijk JG, Lammers GJ. Focusing on vigilance instead of sleepiness in the assessment of narcolepsy: high sensitivity of the Sustained Attention to Response Task (SART) Sleep. 2006;29:187–91. [PubMed] [Google Scholar]
- 3.Broughton R, Ghanem Q, Hishikawa Y, Sugita Y, Nevsimalova S, Roth B. Life effects of narcolepsy in 180 patients from North America, Asia and Europe compared to matched controls. Can J Neurol Sci. 1981;8:299–304. doi: 10.1017/s0317167100043419. [DOI] [PubMed] [Google Scholar]
- 4.Findley LJ, Suratt PM, Dinges DF. Time-on-task decrements in “steer clear” performance of patients with sleep apnea and narcolepsy. Sleep. 1999;22:804–9. doi: 10.1093/sleep/22.6.804. [DOI] [PubMed] [Google Scholar]
- 5.Ozaki A, Inoue Y, Nakajima T, et al. Health-related quality of life among drugnaive patients with narcolepsy with cataplexy, narcolepsy without cataplexy, and idiopathic hypersomnia without long sleep time. J Clin Sleep Med. 2008;4:572–8. [PMC free article] [PubMed] [Google Scholar]
- 6.Scammell TE. The neurobiology, diagnosis, and treatment of narcolepsy. Ann Neurol. 2003;53:154–66. doi: 10.1002/ana.10444. [DOI] [PubMed] [Google Scholar]
- 7.Peyron C, Faraco J, Rogers W, et al. A mutation in a case of early onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains. Nat Med. 2000;6:991–7. doi: 10.1038/79690. [DOI] [PubMed] [Google Scholar]
- 8.Thannickal TC, Moore RY, Nienhuis R, et al. Reduced number of hypocretin neurons in human narcolepsy. Neuron. 2000;27:469–74. doi: 10.1016/s0896-6273(00)00058-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Crocker A, Espana RA, Papadopoulou M, et al. Concomitant loss of dynorphin, NARP, and orexin in narcolepsy. Neurology. 2005;65:1184–8. doi: 10.1212/01.wnl.0000168173.71940.ab. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bourgin P, Huitron-Resendiz S, Spier AD, et al. Hypocretin-1 modulates rapid eye movement sleep through activation of locus coeruleus neurons. J Neurosci. 2000;20:7760–5. doi: 10.1523/JNEUROSCI.20-20-07760.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Adamantidis AR, Zhang F, Aravanis AM, Deisseroth K, de Lecea L. Neural substrates of awakening probed with optogenetic control of hypocretin neurons. Nature. 2007;450:420–4. doi: 10.1038/nature06310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Mochizuki T, Arrigoni E, Marcus JN, et al. Orexin receptor 2 expression in the posterior hypothalamus rescues sleepiness in narcoleptic mice. Proc Natl Acad Sci U S A. 2011;108:4471–6. doi: 10.1073/pnas.1012456108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chemelli RM, Willie JT, Sinton CM, et al. Narcolepsy in orexin knockout mice: molecular genetics of sleep regulation. Cell. 1999;98:437–51. doi: 10.1016/s0092-8674(00)81973-x. [DOI] [PubMed] [Google Scholar]
- 14.Hara J, Beuckmann CT, Nambu T, et al. Genetic ablation of orexin neurons in mice results in narcolepsy, hypophagia, and obesity. Neuron. 2001;30:345–54. doi: 10.1016/s0896-6273(01)00293-8. [DOI] [PubMed] [Google Scholar]
- 15.Espana RA, McCormack SL, Mochizuki T, Scammell TE. Running promotes wakefulness and increases cataplexy in orexin knockout mice. Sleep. 2007;30:1417–25. doi: 10.1093/sleep/30.11.1417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Saper CB, Fuller PM, Pedersen NP, Lu J, Scammell TE. Sleep state switching. Neuron. 2010;68:1023–42. doi: 10.1016/j.neuron.2010.11.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mitler MM, Hayduk R. Benefits and risks of pharmacotherapy for narcolepsy. Drug Saf. 2002;25:791–809. doi: 10.2165/00002018-200225110-00004. [DOI] [PubMed] [Google Scholar]
- 18.Tal J. Adeno-associated virus-based vectors in gene therapy. J Biomed Sci. 2000;7:279–91. doi: 10.1007/BF02253246. [DOI] [PubMed] [Google Scholar]
- 19.Scammell TE, Arrigoni E, Thompson MA, Ronan PJ, Saper CB, Greene RW. Focal deletion of the adenosine A1 receptor in adult mice using an adeno-associated viral vector. J Neurosci. 2003;23:5762–70. doi: 10.1523/JNEUROSCI.23-13-05762.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Griffey M, Macauley SL, Ogilvie JM, Sands MS. AAV2-mediated ocular gene therapy for infantile neuronal ceroid lipofuscinosis. Mol Ther. 2005;12:413–21. doi: 10.1016/j.ymthe.2005.04.018. [DOI] [PubMed] [Google Scholar]
- 21.Pawlyk BS, Smith AJ, Buch PK, et al. Gene replacement therapy rescues photoreceptor degeneration in a murine model of Leber congenital amaurosis lacking RPGRIP. Invest Ophthalmol Vis Sci. 2005;46:3039–45. doi: 10.1167/iovs.05-0371. [DOI] [PubMed] [Google Scholar]
- 22.High KA, Aubourg P. rAAV human trial experience. Methods Mol Biol. 2011;807:429–57. doi: 10.1007/978-1-61779-370-7_18. [DOI] [PubMed] [Google Scholar]
- 23.Mieda M, Willie JT, Hara J, Sinton CM, Sakurai T, Yanagisawa M. Orexin peptides prevent cataplexy and improve wakefulness in an orexin neuron-ablated model of narcolepsy in mice. Proc Natl Acad Sci U S A. 2004;101:4649–54. doi: 10.1073/pnas.0400590101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kantor S, Mochizuki T, Janisiewicz AM, Clark E, Nishino S, Scammell TE. Orexin neurons are necessary for the circadian control of REM sleep. Sleep. 2009;32:1127–34. doi: 10.1093/sleep/32.9.1127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Scammell TE, Willie JT, Guilleminault C, Siegel JM. A consensus definition of cataplexy in mouse models of narcolepsy. Sleep. 2009;32:111–6. [PMC free article] [PubMed] [Google Scholar]
- 26.Fujiki N, Cheng T, Yoshino F, Nishino S. Specificity of direct transitions from wake to REM sleep in orexin/ataxin-3 narcoleptic mice. Sleep. 2006;29:A225. doi: 10.1016/j.expneurol.2009.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yoshida K, McCormack S, Espana RA, Crocker A, Scammell TE. Afferents to the orexin neurons of the rat brain. J Comp Neurol. 2006;494:845–61. doi: 10.1002/cne.20859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Liu M, Blanco-Centurion C, Konadhode R, et al. Orexin gene transfer into zona incerta neurons suppresses muscle paralysis in narcoleptic mice. J Neurosci. 2011;31:6028–40. doi: 10.1523/JNEUROSCI.6069-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Trachsel L, Tobler I, Achermann P, Borbely AA. Sleep continuity and the REM-nonREM cycle in the rat under baseline conditions and after sleep deprivation. Physiol Behav. 1991;49:575–80. doi: 10.1016/0031-9384(91)90283-t. [DOI] [PubMed] [Google Scholar]
- 30.Franken P, Malafosse A, Tafti M. Genetic determinants of sleep regulation in inbred mice. Sleep. 1999;22:155–69. [PubMed] [Google Scholar]
- 31.Mochizuki T, Crocker A, McCormack S, Yanagisawa M, Sakurai T, Scammell TE. Behavioral state instability in orexin knockout mice. J Neurosci. 2004;24:6291–300. doi: 10.1523/JNEUROSCI.0586-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Edgar DM, Dement WC, Fuller CA. Effect of SCN lesions on sleep in squirrel monkeys: evidence for opponent processes in sleep-wake regulation. J Neurosci. 1993;13:1065–79. doi: 10.1523/JNEUROSCI.13-03-01065.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Xu B, Goulding EH, Zang K, et al. Brain-derived neurotrophic factor regulates energy balance downstream of melanocortin-4 receptor. Nat Neurosci. 2003;6:736–42. doi: 10.1038/nn1073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Mochizuki T, Klerman EB, Sakurai T, Scammell TE. Elevated body temperature during sleep in orexin knockout mice. Am J Physiol Regul Integr Comp Physiol. 2006;291:R533–40. doi: 10.1152/ajpregu.00887.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Lavie P. REM periodicity under ultrashort sleep/wake cycle in narcoleptic patients. Can J Psychol. 1991;45:185–93. doi: 10.1037/h0084284. [DOI] [PubMed] [Google Scholar]
- 36.Dantz B, Edgar DM, Dement WC. Circadian rhythms in narcolepsy: studies on a 90 minute day. Electroencephalogr Clin Neurophysiol. 1994;90:24–35. doi: 10.1016/0013-4694(94)90110-4. [DOI] [PubMed] [Google Scholar]
- 37.Dijk DJ, Czeisler CA. Contribution of the circadian pacemaker and the sleep homeostat to sleep propensity, sleep structure, electroencephalo-graphic slow waves, and sleep spindle activity in humans. J Neurosci. 1995;15:3526–38. doi: 10.1523/JNEUROSCI.15-05-03526.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Wurts SW, Edgar DM. Circadian and homeostatic control of rapid eye movement (REM) sleep: promotion of REM tendency by the suprachias-matic nucleus. J Neurosci. 2000;20:4300–10. doi: 10.1523/JNEUROSCI.20-11-04300.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lee ML, Swanson BE, de la Iglesia HO. Circadian timing of REM sleep is coupled to an oscillator within the dorsomedial suprachiasmatic nucleus. Curr Biol. 2009;19:848–52. doi: 10.1016/j.cub.2009.03.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Piper DC, Upton N, Smith MI, Hunter AJ. The novel brain neuropeptide, orexin-A, modulates the sleep-wake cycle of rats. Eur J Neurosci. 2000;12:726–30. doi: 10.1046/j.1460-9568.2000.00919.x. [DOI] [PubMed] [Google Scholar]
- 41.Espana RA, Baldo BA, Kelley AE, Berridge CW. Wake-promoting and sleep-suppressing actions of hypocretin (orexin): basal forebrain sites of action. Neuroscience. 2001;106:699–715. doi: 10.1016/s0306-4522(01)00319-0. [DOI] [PubMed] [Google Scholar]
- 42.Liu M, Thankachan S, Kaur S, et al. Orexin (hypocretin) gene transfer diminishes narcoleptic sleep behavior in mice. Eur J Neurosci. 2008;28:1382–93. doi: 10.1111/j.1460-9568.2008.06446.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Okuro M, Fujiki N, Kotorii N, Ishimaru Y, Sokoloff P, Nishino S. Effects of paraxanthine and caffeine on sleep, locomotor activity, and body temperature in orexin/ataxin-3 transgenic narcoleptic mice. Sleep. 2010;33:930–42. doi: 10.1093/sleep/33.7.930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Yoshida Y, Fujiki N, Nakajima T, et al. Fluctuation of extracellular hypocretin-1 (orexin A) levels in the rat in relation to the light-dark cycle and sleep-wake activities. Eur J Neurosci. 2001;14:1075–81. doi: 10.1046/j.0953-816x.2001.01725.x. [DOI] [PubMed] [Google Scholar]
- 45.Blanco-Centurion C, Liu M, Konadhode R, Pelluru D, Shiromani PJ. Effects of orexin gene transfer in the dorsolateral pons in orexin knockout mice. Sleep. 2013;36:31–40. doi: 10.5665/sleep.2296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Makela KA, Wigren HK, Zant JC, et al. Characterization of sleep-wake patterns in a novel transgenic mouse line overexpressing human prepro-orexin/hypocretin. Acta Physiol (Oxf) 2010;198:237–49. doi: 10.1111/j.1748-1716.2009.02068.x. [DOI] [PubMed] [Google Scholar]
- 47.Zhang S, Zeitzer JM, Sakurai T, Nishino S, Mignot E. Sleep/wake fragmentation disrupts metabolism in a mouse model of narcolepsy. J Physiol. 2007;581:649–63. doi: 10.1113/jphysiol.2007.129510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Willie JT, Takahira H, Shibahara M, et al. Ectopic overexpression of orexin alters sleep/wakefulness states and muscle tone regulation during REM sleep in mice. J Mol Neurosci. 2011;43:155–61. doi: 10.1007/s12031-010-9437-7. [DOI] [PubMed] [Google Scholar]
- 49.Mandel RJ, Manfredsson FP, Foust KD, et al. Recombinant adeno-associated viral vectors as therapeutic agents to treat neurological disorders. Mol Ther. 2006;13:463–83. doi: 10.1016/j.ymthe.2005.11.009. [DOI] [PubMed] [Google Scholar]
- 50.Paxinos G, Franklin KBJ. The mouse brain in stereotaxic coordinates. 2nd ed. San Diego: Academic Press; 2001. [Google Scholar]