

Abstract
Bone is the most common site of breast cancer metastasis and once established, it is frequently incurable. Critical to our ability to prevent and treat bone metastasis is the identification of the key factors mediating its establishment and understanding their biological function. To address this issue we previously carried out an in vivo selection process to isolate murine mammary tumor sublines possessing an enhanced ability to colonize the bone. A comparison of gene expression between parental cells and sublines by genome-wide cDNA microarray analysis revealed several potential mediators of bone metastasis, including the pyrophosphate-generating ectoenzyme Enpp1. By qRT-PCR and Western analysis we found that expression of Enpp1 was elevated in human breast cancer cell lines known to produce bone metastasis in animal models compared to non-metastatic and normal mammary epithelial cell lines. Further, in clinical specimens, levels of Enpp1 were significantly elevated in human primary breast tumors relative to normal mammary epithelium, with highest levels observed in breast-bone metastasis as determined by qRT-PCR and immunohistochemical analysis. To examine the potential role of Enpp1 in the development of bone metastasis, Enpp1 expression was stably increased in the breast cancer cell line MDA-MB-231 and the ability to colonize the bone following intracardiac and direct intratibial injection of athymic nude mice was determined. By both routes of administration, increased expression of Enpp1 enhanced the ability of MDA-MB-231 cells to form tumors in the bone relative to cells expressing vector alone, as determined by digital radiography and histological analysis. Taken together, these data suggest a potential role for Enpp1 in the development of breast cancer bone metastasis.
Introduction
Metastasis is the ultimate cause of mortality in breast cancer patients, developing in the bone more frequently than any other site. Bone metastasis occurs in approximately 80% of patients with advanced breast cancer and causes considerable morbidity in the form of bone pain, pathological fractures, nerve compression, and life-threatening hypercalcemia. Sadly, bone metastasis is frequently incurable and the median survival time following diagnosis is just 2 years. Despite ongoing research efforts, the molecular and cellular mechanisms that regulate the development of bone metastasis and resultant osteolysis remain poorly understood. Identification of factors regulating this process may reveal novel targets for preventative and therapeutic interventions against this devastating disease.
In this study, we identified the ectoenzyme ectonucleotide pyrophosphatase/phosphodiesterase I (Enpp1) as being overexpressed in human primary breast cancer relative to normal mammary epithelium and provide the first evidence of its potential to foster the development of bone metastasis. Enpp1 is a type II transmembrane glycoprotein with pyrophosphatase and phosphodiesterase activity, expressed highly in bone and cartilage [1]. Enpp1 is located at 6q22-q23, a region reportedly amplified in breast cancer [2], [3], and its mutation has been associated with several disorders including infantile arterial calcification [4], [5], ossification of the posterior longitudinal ligament of the spine [6], and insulin resistance [7]. Biologically, Enpp1 is well known for its role in regulating bone mineralization, serving as the principal ectoenzyme responsible for the generation of extracellular inorganic pyrophosphate (PPi) [1], a potent inhibitor of hydroxyapatite formation [8], [9]. In addition, Enpp1 has been shown to modulate insulin signaling, recently reported to regulate bone acquisition and energy metabolism through effects on osteoblasts [10], while also functioning in purinergic signaling [11].
Results and Discussion
Association of Enpp1 with Breast Cancer Bone Metastasis
Previously, we utilized the murine mammary tumor cell line NT2.5 in an in vivo selection process to generate sublines having an increased propensity to develop metastasis to bone as compared to other sites [12]. To identify genes potentially important for the development of bone metastasis, microarray analysis was conducted comparing gene expression between parental NT2.5 cells and sublines having a moderately (BO3) and highly (BO6) increased preference for the colonization of bone following intracardiac injection. These microarray data have been deposited in the National Center for Biotechnology Information (NCBI) Gene Expression Omnibus (GEO) under accession number GSE47714. While many candidates were identified in this analysis [12], of particular interest was the pyrophosphate-generating ectoenzyme Enpp1, a central regulator of extracellular PPi levels, which have a substantial impact on bone physiology [13]. By microarray analysis, Enpp1 mRNA levels were incrementally increased from parental NT2.5 cells to BO3 and finally BO6 sublines (p = 0.008), which was confirmed in a separate experiment by qRT-PCR (Fig. 1A).
Figure 1. Expression of Enpp1 in breast cancer cell lines.
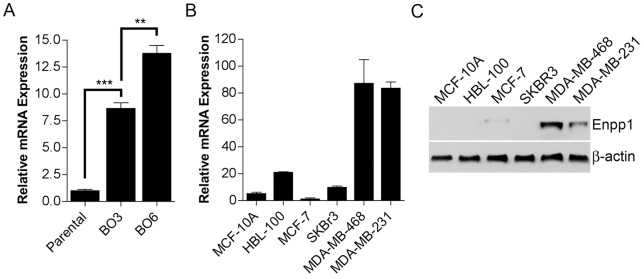
Enpp1 (A, B) mRNA and (C) protein expression was determined in (A) NT2.5 murine breast cancer cells and bone sublines, and (B, C) immortalized normal human mammary epithelial cell lines and human breast cancer cell lines by (A, B) qRT-PCR and (C) Western analysis. (A, B) Data are representative of three independent experiments performed in triplicate and expressed as the mean ± s.e.m (**p<0.01, ***p<0.001). (C) Protein expression was determined by Western analysis performed on equal amounts of protein from total cell lysates.
Expression of Enpp1 in Human Breast Cancer Cell Lines and Tissues
To determine whether the expression pattern of Enpp1 observed using the NT2.5 murine mammary tumor model translated to human breast cancer, we examined the expression of Enpp1 in a panel of human breast cancer cell lines with and without the ability to establish bone metastasis in animal models (our unpublished observations). Similar to our observation in NT2.5 cells and bone metastasis sublines, Enpp1 expression was elevated in human breast cancer cell lines that are capable of forming bone metastasis in animal models (MDA-MB-231/MDA-MB-468) as compared to those incapable of colonizing the bone (MCF-7/SKBR3), which displayed expression levels similar to that in immortalized normal mammary epithelial cell lines (MCF-10A/HBL-100) (Fig. 1B, C). Extending our investigation to clinical samples, we found that Enpp1 mRNA expression was significantly elevated in primary breast tumors relative to normal mammary epithelium (p<0.05), with the highest levels observed in breast-bone metastasis (Fig. 2A). Correspondingly, immunohistochemical (IHC) analysis revealed strong Enpp1 protein expression in 32% (n = 37) of primary breast tumors while little to no Enpp1 expression was detected in adjacent normal mammary epithelium (Fig. 2B, C). Further, Enpp1 was strongly expressed in 100% (n = 6) of breast-bone metastases (Fig. 2D). Importantly, the results of our IHC analysis not only correlated with our mRNA findings, but confirmed that Enpp1 was expressed in tumor cells as opposed to other cell types within these tissues.
Figure 2. Expression of Enpp1 in human primary breast cancer and bone metastasis tissues.

Enpp1 (A) mRNA and (B–D) protein expression in human normal mammary epithelium (N), primary breast tumor (T), and breast cancer bone metastasis (BBM) was determined by (A) qRT-PCR and (B–D) IHC analysis. (A) Data are representative of three independent experiments performed in triplicate (*p<0.05 for T versus N). (B–D) Proteins were identified using DAB (brown). Sections were counterstained with hematoxylin and visualized by light microscopy (200X). N – Normal, T – Tumor.
Effect of Enpp1 on the Establishment of Bone Metastasis
To investigate the potential importance of Enpp1 in the establishment of bone metastasis, we utilized the MDA-MB-231 breast cancer cell line, which is capable of forming osteolytic bone metastasis in animal models and displayed moderate expression of Enpp1 by Western analysis (Fig. 1C). MDA-MB-231 cells were stably infected with a retroviral expression vector containing full-length Enpp1 cDNA or with vector alone (EV). Levels of biologically active Enpp1 were increased by greater than two-fold as determined by Western analysis (Fig. 3A) and nucleotide phosphodiesterase activity assay (Fig. 3B). Further, the rate of cell proliferation in MDA-MB-231-Enpp1 cells was equivalent to that in MDA-MB-231-EV cells as determined by MTS assay (data not shown). It should be noted that in an attempt to compliment this gain-of-function approach with a loss-of-function approach employing RNAi, we observed that all control shRNA tested, whether scrambled Enpp1 sequence or unrelated sequence, resulted in reduced Enpp1 expression, preventing use of this strategy and indicating that Enpp1 may be subject to off-target RNAi effects.
Figure 3. Generation of breast cancer cells with increased Enpp1 expression and enzymatic activity.
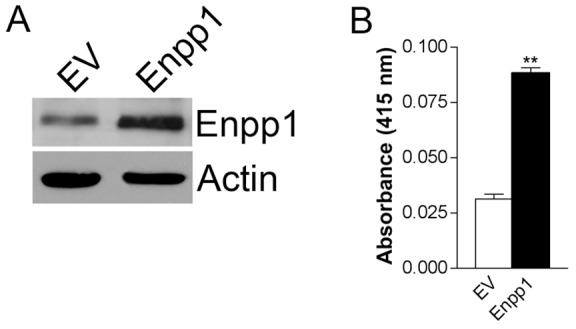
MDA-MB-231 cells were stably transduced with Enpp1 or empty vector (EV). Enpp1 (A) protein expression was determined by Western analysis performed on equal amounts of protein from total cell lysates. (B) Nucleotide phosphodiesterase activity was determined using the nucleotide derivative p-nitrophenyl thymidine 5′-monophosphate (pNP-TMP) as a substrate. Data are representative of three independent experiments performed in triplicate and expressed as the mean ± s.e.m (**p<0.01).
To determine whether increased levels of Enpp1 influence the formation of bone metastasis in vivo, both intratibial and intracardiac xenograft model systems were used. As expected, both the MDA-MB-231-Enpp1 and MDA-MB-231-EV group displayed a 100% overall incidence of bone metastasis. However, following both intratibial and intracardiac tumor cell administration, animals that received MDA-MB-231-Enpp1 cells demonstrated more rapid disease progression, evidenced by a significant increase in both osteolysis and tumor area as measured on radiographic images (Fig. 4A, C) and histological sections (Fig. 4 B, D), respectively. Taken together, these data indicate that increased expression of Enpp1 can enhance the development of bone metastasis.
Figure 4. Effect of Enpp1 on the development of bone metastasis.

MDA-MB-231 cells stably expressing Enpp1 or empty vector (EV) were injected into the (A, B) tibia (n = 9/group) and (C, D) left cardiac ventricle (n = 5/group) of athymic nude mice and digital radiographic imaging was performed at weekly intervals. (A, C) Osteolytic area and (B, D) tumor area were measured on radiographic images and histological sections, respectively (*p<0.05, **p<0.01). Representative images are shown at 4 weeks following tumor cell administration. Black outlined areas on histological images indicate areas of tumor.
Although the mechanism through which Enpp1 enhances the development of osteolytic bone metastasis remains unknown, given its function as a nucleotide pyrophosphatase/phosphodiesterase, it may be linked to the production of extracellular inorganic pyrophosphate. The balance of PPi and inorganic phosphate (Pi) is critical to bone physiology, as PPi acts as an inhibitor of mineral deposition [14], [15]. In addition, PPi has been shown to increase production of osteopontin in osteoblasts, another key mineralization inhibitor [16]. In the case of bone metastasis, it is possible that levels of PPi within the bone environment become elevated by the presence of tumor cells expressing Enpp1. This imbalance in PPi could lead to reduced mineralization, potentially creating an environment favoring bone resorption, which has been shown to be critical to the development of bone metastasis as it liberates growth factors from the bone, fueling tumor growth [17]. Beyond its effects on PPi/Pi balance, the involvement of Enpp1 in insulin and purinergic signaling represent additional avenues worthy of investigation, all having the potential to yield novel insights regarding the interaction between tumor cells and the bone environment.
In summary, we have provided evidence that Enpp1 expression is elevated in human primary breast tumors relative to normal mammary epithelium, with highest levels observed in breast-bone metastasis. Further, consistent with the expression patterns observed in both murine and human breast cancer cell lines, increased expression of Enpp1 enhanced the development of bone metastasis in animal models. These data not only support a potential role for Enpp1 in breast-bone metastasis, but suggest that Enpp1 may be useful as a prognostic indicator for breast cancer.
Materials and Methods
Ethics Statement
All experiments were carried out in accordance with the National Research Council's “Guide to the Care and Use of Laboratory Animals”. Animal use was approved by the Johns Hopkins Animal Care and Use Committee, animal welfare assurance #A3272-01, protocol #MO10M450.
Cell lines and tissues
The murine mammary carcinoma cell line NT2.5 and sublines, BO3 and BO6, were obtained as previously described [12]. NT2.5 parental cells and sublines were maintained in RPMI supplemented with 20% fetal bovine serum (FBS), 10 mM HEPES, 1% nonessential amino acids, 1 mM sodium pyruvate and 10 lg/ml insulin. All other cell lines were obtained from American Type Culture Collection (Rockville, MD) and cultured according to conditions specified. Mammary organoid samples, kindly provided by Dr. Saraswati Sukumar (Johns Hopkins University School of Medicine, Baltimore, MD), were prepared from reduction mammoplasty specimens of women with no breast abnormalities. Breast tumor and bone metastasis tissues were obtained from the Surgical Pathology Division of the Johns Hopkins Hospital following the approval of the institutional review board (IRB) of the Johns Hopkins University School of Medicine. For all specimens, required written informed patient consents were obtained as approved by the IRB.
Quantitative (q)RT-PCR
Total RNA was extracted using Trizol (Invitrogen) and cDNA was generated by reverse transcription. 25 μl reactions contained 1X SYBR Green Reaction Mix (Applied Biosystems), 1 μl cDNA, and 100 nm of each primer: Enpp1 (sense) 5′-GAAACGCCTCCTACCCTCTT-3′, (antisense) 5′-ATCCTGGCCAGAAAAATGTG-3′; GAPDH (sense) 5′-GTCAGTGGTGGACCTGACCT-3′, (antisense) 5′-TGCTGTAGCCAAATTCGTTG-3′. qRT-PCR parameters were: 1 cycle (95°C for 3 minutes) and 40 cycles (95°C for 30 seconds, 61.9°C for 30 seconds, and 72°C for 45 seconds). Amplification of GAPDH was used as an internal control. Relative expression between samples was calculated by the comparative CT method.
Western analysis
Total protein was extracted from cell lines using lysis buffer consisting of 15% glycerol, 5% SDS and 250 mM Tris-HCl, pH 6.7. Equal amounts of protein were resolved using 10% SDS- PAGE. Protein was transferred to ECL nitrocellulose membranes (Amersham) and probed with anti-Enpp1 (Everest Biotech) and β-actin (Sigma-Aldrich) antibody. Membranes were then incubated with horseradish peroxidase-conjugated antibody against rabbit IgG (Amersham) and binding was revealed by chemiluminescence (Amersham).
Immunohistochemistry
Paraffin-embedded sections were deparaffinized in xylene and rehydrated through graded ethanol. Antigen retrieval was achieved by immersing sections in 0.01 mol/L sodium citrate (pH 6.0) and heating in a steamer for 20 min. Sections were cooled to room temperature (RT), and endogenous peroxidase activity was quenched by immersing in 0.3% hydrogen peroxide. Blocking was carried out by incubation in diluted normal rabbit serum (Vector Laboratories) as per the manufacturer's instructions. Sections were then incubated with goat polyclonal anti-Enpp1 (Everest Biotech) at a 1:750 dilution for 16 hours at 4°C. Diluted biotinylated anti-goat IgG (Vector Laboratories) was added to the sections and incubated for 30 min. at RT followed by Vectastain ABC reagent. Enpp1 proteins were visualized using 3, 3′-diaminobenzidine (DAB) as per the manufacturer's instructions (Vector Laboratories). Sections were subsequently washed in water and counterstained in hematoxylin (Richard-Allan Scientific). Images were acquired by light microscopy.
Nucleotide Phosphodiesterase activity assay
Cells were plated in triplicate in a 96-well plate at 3000 cells/well and cultured in complete media for 72 hours. 1 μg/ml p-nitrophenylthymidine monophosphate was then added to each well as a substrate for phosphodiesterase and the reaction was allowed to proceed for 1 hour at 37°C. Formed p-nitrophenol was then quantified by measuring the absorbance at a wavelength of 415 nm in a microplate reader.
In vivo assessment of bone metastasis and osteolysis
To determine the effect of Enpp1 on the establishment of bone metastasis, Enpp1 cDNA was cloned into the retroviral expression vector, pBabe-puro (Addgene plasmid 1764), and MDA-MB-231 cells were infected with pBabe-puro or pBabe-puro-Enpp1. Stable pools were selected in the presence of 1 µg/ml puromycin (Sigma-Aldrich) for one week.
In separate experiments, tumor cells were injected into the right tibia (2.5×104 cells) and left cardiac ventricle (1×105 cells) of 5-week-old female athymic nude mice (NCI-Frederick Cancer Research Facility). To monitor osteolysis, digital radiographic images were obtained once per week using a Faxitron MX-20 X-ray unit (Faxitron X-ray Corp.) and osteolytic area was measured using MetaMorph image analysis software (Meta Imag- ing Series version 6.1, Universal Imaging Corp.). The experiment was terminated at 4 weeks when there was marked tibial osteolysis with or without soft tissue extension of the tumor in any group. To assess tumor area, tibiae were harvested, fixed in formalin for 24 hours, and decalcified in 10% EDTA, pH 7.4 for 48 hours. Paraffin-embedded sections (5 μm) were generated laterally throughout the tibiae at 100 μm intervals. A minimum of five sections was stained with hematoxylin and eosin, and light microscopic images were acquired. The total area occupied by intraosseous and extraosseous tumor was then measured using MetaMorph image analysis software.
Funding Statement
These authors have no support or funding to report.
References
- 1. Huang R, Rosenbach M, Vaughn R, Provvedini D, Rebbe N, et al. (1994) Expression of the murine plasma cell nucleotide pyrophosphohydrolase PC-1 is shared by human liver, bone, and cartilage cells. Regulation of PC-1 expression in osteosarcoma cells by transforming growth factor-beta. The Journal of clinical investigation 94: 560–567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Rodriguez C, Causse A, Ursule E, Theillet C (2000) At least five regions of imbalance on 6q in breast tumors, combining losses and gains. Genes, chromosomes & cancer 27: 76–84. [PubMed] [Google Scholar]
- 3. Marchio C, Iravani M, Natrajan R, Lambros MB, Savage K, et al. (2008) Genomic and immunophenotypical characterization of pure micropapillary carcinomas of the breast. The Journal of pathology 215: 398–410. [DOI] [PubMed] [Google Scholar]
- 4. Rutsch F, Ruf N, Vaingankar S, Toliat MR, Suk A, et al. (2003) Mutations in ENPP1 are associated with ‘idiopathic’ infantile arterial calcification. Nature genetics 34: 379–381. [DOI] [PubMed] [Google Scholar]
- 5. Rutsch F, Vaingankar S, Johnson K, Goldfine I, Maddux B, et al. (2001) PC-1 nucleoside triphosphate pyrophosphohydrolase deficiency in idiopathic infantile arterial calcification. The American journal of pathology 158: 543–554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Saito T, Shimizu Y, Hori M, Taguchi M, Igarashi T, et al. (2011) A patient with hypophosphatemic rickets and ossification of posterior longitudinal ligament caused by a novel homozygous mutation in ENPP1 gene. Bone 49: 913–916. [DOI] [PubMed] [Google Scholar]
- 7. Goldfine ID, Maddux BA, Youngren JF, Reaven G, Accili D, et al. (2008) The role of membrane glycoprotein plasma cell antigen 1/ectonucleotide pyrophosphatase phosphodiesterase 1 in the pathogenesis of insulin resistance and related abnormalities. Endocrine reviews 29: 62–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Fleisch H, Russell RG, Straumann F (1966) Effect of pyrophosphate on hydroxyapatite and its implications in calcium homeostasis. Nature 212: 901–903. [DOI] [PubMed] [Google Scholar]
- 9. Meyer JL, Nancollas GH (1973) The influence of multidentate organic phosphonates on the crystal growth of hydroxyapatite. Calcified tissue research 13: 295–303. [DOI] [PubMed] [Google Scholar]
- 10. Clemens TL, Karsenty G (2011) The osteoblast: an insulin target cell controlling glucose homeostasis. Journal of bone and mineral research: the official journal of the American Society for Bone and Mineral Research 26: 677–680. [DOI] [PubMed] [Google Scholar]
- 11. Belleannee C, Da Silva N, Shum WW, Brown D, Breton S (2010) Role of purinergic signaling pathways in V-ATPase recruitment to apical membrane of acidifying epididymal clear cells. American journal of physiology Cell physiology 298: C817–830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Lau WM, Weber KL, Doucet M, Chou YT, Brady K, et al. (2010) Identification of prospective factors promoting osteotropism in breast cancer: a potential role for CITED2. International journal of cancer Journal international du cancer 126: 876–884. [DOI] [PubMed] [Google Scholar]
- 13. Harmey D, Hessle L, Narisawa S, Johnson KA, Terkeltaub R, et al. (2004) Concerted regulation of inorganic pyrophosphate and osteopontin by akp2, enpp1, and ank: an integrated model of the pathogenesis of mineralization disorders. The American journal of pathology 164: 1199–1209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Fedde KN, Blair L, Silverstein J, Coburn SP, Ryan LM, et al. (1999) Alkaline phosphatase knock-out mice recapitulate the metabolic and skeletal defects of infantile hypophosphatasia. Journal of bone and mineral research: the official journal of the American Society for Bone and Mineral Research 14: 2015–2026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Russell RG, Bisaz S, Donath A, Morgan DB, Fleisch H (1971) Inorganic pyrophosphate in plasma in normal persons and in patients with hypophosphatasia, osteogenesis imperfecta, and other disorders of bone. The Journal of clinical investigation 50: 961–969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Addison WN, Azari F, Sorensen ES, Kaartinen MT, McKee MD (2007) Pyrophosphate inhibits mineralization of osteoblast cultures by binding to mineral, up-regulating osteopontin, and inhibiting alkaline phosphatase activity. The Journal of biological chemistry 282: 15872–15883. [DOI] [PubMed] [Google Scholar]
- 17. Chirgwin JM, Guise TA (2000) Molecular mechanisms of tumor-bone interactions in osteolytic metastases. Critical reviews in eukaryotic gene expression 10: 159–178. [PubMed] [Google Scholar]