

Abstract
AIM: To determine the effects of BN52021 on platelet-activating factor receptor (PAFR) signaling molecules under lipopolysaccharide (LPS)-induced inflammatory conditions in MS1 cells.
METHODS: MS1 cells (a mouse pancreatic islet endothelial cell line) were grown in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum, 2 mmol/L glutamine and 100 μg/mL penicillin/streptomycin in 5% CO2 at 37 °C. After growth to confluency in media, the cells were processed for subsequent studies. The MS1 cells received 0, 0.1, 1 and 10 μg/mL LPS in this experiment. The viability/proliferation of the cells induced by LPS was observed using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide colorimetric assay. Apoptosis and necrosis of the cells under the inflammatory condition described previously were observed using Hoechst 33342-propidium iodide staining. Adenylate cyclase (AC), phospholipase A2 (PLA2), phospholipase Cβ (PLCβ), protein tyrosine kinase (PTK), G protein-coupled receptor kinases (GRK) and p38-mitogen-activated protein kinase (p38 MAPK) mRNA in the PAFR signaling pathway were measured by real-time polymerase chain reaction. The protein expression level of phosphorylated AC (p-AC), phosphorylated PLA2 (p-PLA2), phosphorylated PTK (p-PTK), phosphorylated p38 MAPK (p-p38 MAPK), PLCβ and GRK was measured using Western blotting analysis.
RESULTS: The activity of MS1 cells incubated with different concentrations of LPS for 6 h decreased significantly in the 1 μg/mL LPS group (0.49 ± 0.10 vs 0.67 ± 0.13, P < 0.05) and 10 μg/mL LPS group (0.44 ± 0.10 vs 0.67 ± 0.13, P < 0.001), but not in 0.1 μg/mL group. When the incubation time was extended to 12 h (0.33 ± 0.05, 0.32 ± 0.03 and 0.25 ± 0.03 vs 0.69 ± 0.01) and 24 h (0.31 ± 0.01, 0.29 ± 0.03 and 0.25 ± 0.01 vs 0.63 ± 0.01), MS1 cell activity decreased in all LPS concentration groups compared with the blank control (P < 0.001). BN52021 significantly improved the cell activity when its concentration reached 50 μmol/L compared with the group that received LPS treatment alone, which was consistent with the results obtained from fluorescence staining. The mRNAs levels of AC (4.02 ± 0.14 vs 1.00 ± 0.13), GRK (2.63 ± 0.03 vs 1.00 ± 0.12), p38 MAPK (3.87 ± 0.07 vs 1.00 ± 0.17), PLA2 (3.31 ± 0.12 vs 1.00 ± 0.12), PLCβ (2.09 ± 0.08 vs 1.00 ± 0.06) and PTK (1.85 ± 0.07 vs 1.00 ± 0.11) were up-regulated after LPS stimulation as compared with the blank control (P < 0.05). The up-regulated mRNAs including AC (2.35 ± 0.13 vs 3.87 ± 0.08), GRK (1.17 ± 0.14 vs 2.65 ± 0.12), p38 MAPK (1.48 ± 0.18 vs 4.30 ± 0.07), PLCβ (1.69 ± 0.10 vs 2.41 ± 0.13) and PLA2 (1.87 ± 0.11 vs 2.96 ± 0.08) were significantly suppressed by BN52021 except for that of PTK. The level of p-AC (1.11 ± 0.12 vs 0.65 ± 0.08), GRK (0.83 ± 0.07 vs 0.50 ± 0.03), PLCβ (0.83 ± 0.16 vs 0.50 ± 0.10) and p-p38 MAPK (0.74 ± 0.10 vs 0.38 ± 0.05) was up-regulated after LPS stimulation as compared with the blank control (P < 0.05). The up-regulated proteins, including p-AC (0.65 ± 0.15 vs 1.06 ± 0.14), GRK (0.47 ± 0.10 vs 0.80 ± 0.06), PLCβ (0.47 ± 0.04 vs 0.80 ± 0.19) and p-p38 MAPK (0.30 ± 0.10 vs 0.97 ± 0.05), was significantly suppressed by BN52021, but p-PLA2 and p-PTK protein level were not suppressed.
CONCLUSION: BN52021 could effectively inhibit LPS-induced inflammation by down-regulating the mRNA and protein levels of AC, GRK, p38 MAPK, PLA2 and PLCβ in the PAFR signaling pathway.
Keywords: BN52021, Platelet-activating factor receptor, Signaling pathway, Inflammation, Pancreatitis
Core tip: Microcirculatory disorder is considered to be one of the possible mechanisms of pathogenesis of severe acute pancreatitis (SAP). Platelet-activating factor (PAF) is known to mediate microcirculatory disturbance and inflammation. Although BN52021, a PAF receptor antagonist, has demonstrated significant treatment effects on SAP, its mechanism has not been elucidated in detail. In this study, we examined the signaling molecules of the PAF receptor pathway to evaluate whether BN52021 has any influence on the inflammatory effects induced by lipopolysaccharide in MS1 cells, hoping to elucidate the mechanism underlying microcirculatory disturbances in the pathogenesis of SAP in vitro.
INTRODUCTION
Acute pancreatitis (AP) is an inflammatory disease that can develop into severe AP (SAP)[1]. SAP refers to AP associated with organ failure and/or local complications such as necrosis, pseudocyst or abscess, which is a disease of high morbidity and mortality with an unpredictable clinical course[2,3]. There is no clinically effective therapeutic strategy for SAP, because the pathogenesis of the disease remains largely unclarified. The possible explanations for the pathogenesis of SAP include theories of self-digestion, leukocyte overactivation, microcirculatory disorder, bacterial shifting, and secondary infection, which is a second attack by immune functional change, cell apoptosis, oxygen-free radicals, and others from different aspects[4]. Accumulated evidence has proven that microcirculatory disorders are the key pathogenesis of AP. Many complications of SAP are due to the amplifying effects of microcirculatory disruption[5-10]. The inflammation of pancreatic microvascular endothelial cells induced by lipopolysaccharide is a suitable pancreatitis model to simulate the microcirculatory disturbances in vitro.
Platelet-activating factor (PAF), a bioactive phospholipid synthesized and secreted by a variety of cells including pancreatic acini and microvascular endothelium cells[11], is known to mediate many physiological responses such as microcirculatory disturbance and inflammation. AP causes the release of PAF, which induces systemic effects that contribute to circulatory disturbance and multiple organ failure[1]. PAF can significantly potentiate pancreatic tissue damage, increase serum amylase and lipase levels, cause scattered hemorrhages and may serve as a primary mediator of inflammation in the pathological progress of SAP[1,7,12]. A single injection of PAF into the superior pancreaticoduodenal artery of rabbits induces dose-dependent morphologic alterations of the pancreatic tissue and increased serum amylase levels[13]. Our previous research revealed that PAF was stably expressed in the rat pancreas tissue and played an important role in inflammatory response during the procession of SAP[4,14]. PAF could produce physiological and pathological effects by binding to its cell surface receptor, PAF receptor (PAFR). Flickinger et al[15] revealed specific localization of PAFR in the pancreatic vascular endothelium but not in other pancreatic cell types. Recent studies have demonstrated that bacterial lipopolysaccharide (LPS) can induce an increase in the surface expression of PAF receptors[16]. Our recent study demonstrated that BN52021 exerted biological effects through inhibiting the increased PAF level and binding potential with PAFR rather than through decreasing PAFR expression in the pancreatic tissue[17]. Through binding with PAFR, PAF may, through G-protein transduction, activate phospholipase C, phospholipase A2, adenylate cyclase and tyrosine protein kinase, leading to the occurrence and development of SAP[18].
PAFR antagonists can block a series of inflammatory injuries caused by PAF, thereby improving the AP prognosis as a preventive treatment[19]. Research on such a potential therapy has helped elucidate the role of PAF in AP[20]. It was observed that BN52021 extracted from Ginkgo biloba leaves could act as a potent antagonist of PAFR[21], and BN52021 can inhibit the PAF-induced cascade effect in inflammatory reactions, exhibiting an anti-shock effect by reducing the portal vein pressure of liver cirrhosis[22,23]. In experimental pancreatitis models and clinical trials, the administration of several PAF antagonists significantly reduced the level of serum amylase, leukocyte infiltration, and improved capillary blood flow in the pancreas and distant organs, as well as the renal and respiratory functions and the survival rate. BN52021 could significantly reduce vascular permeability, pancreatic edema, hyperamylasemia, diminute superoxide dismutase activity, and inhibit lipid peroxidation in the pancreatic tissue. These changes were accompanied by a significant reduction of acinar cell vacuolization and a remarkable inhibition of inflammatory cell infiltration in the interacinar space[24,25]. Our recent studies have also shown a therapeutic effect of BN52021 on experimental SAP[26-28], but its mechanism is not yet fully understood.
In this study, we examined signaling molecules of the PAFR pathway to evaluate whether a PAF receptor antagonist (BN52021) had any influence on the inflammatory effects induced by lipopolysaccharide in MS1 cells, hoping to elucidate the mechanism underlying the microcirculatory disturbances in the pathogenesis of SAP in vitro.
MATERIALS AND METHODS
Chemicals and reagents
Chemicals and reagents used in this study included BN52021, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) and LPS (Sigma-Aldrich, St. Louis, MO, United States); Dulbecco’s modified Eagle’s medium (DMEM, Gibco/Invitrogen, Carlsbad, CA, United States); the mouse primers for the Adcy1 [adenylate cyclase (AC)], Pla2g4a [phospholipase A2 (PLA2)], Plcb3 [phospholipase Cβ (PLCβ)], Ptk7 [protein tyrosine kinase (PTK)], Adrbk1 [G protein-coupled receptor kinases (GRK)], Mapk14 [p38-mitogen-activated protein kinase (p38 MAPK)] and Gapdh (glyceraldehyde 3-phosphate dehydrogenase) genes (Beijing AuGCT DNA-SYN Biotechnology Co., Ltd., Beijing, China); rabbit polyclonal antibodies of phosphorylated PTK (p-PTK) and phosphorylated AC (p-AC) (Abcam, Cambridge, MA, United States); rabbit polyclonal antibodies for GRK2 and phosphorylated p38 MAPK (p-p38 MAPK) (Epitomics, Burlingame, CA, United States); rabbit polyclonal antibody for phosphorylated PLA2 (p-PLA2) (Cell Signaling Technology, Beverly, MA, United States); rabbit polyclonal antibody for PLCβ (Santa Cruz, Dallas, TX, United States); rabbit polyclonal antibody for β-actin (Abmart, Arlington, MA, United States); protein molecular weight markers, reverse transcription-polymerase chain reaction (RT-PCR) kit and quantitative PCR kit (Beijing TransGen Biotech Co., Ltd, Beijing, China); and polyvinylidene fluoride (PVDF) membranes (BD Biosciences, BD Corporation, MA, United States).
Cell culture
MS1 cell line (a mouse pancreatic islet endothelial cell line firstly established in 1994) was purchased from Shanghai Institute of Cell Biology of the Chinese Academy of Sciences (Shanghai, China). Cells were grown in DMEM supplemented with 10% fetal bovine serum (FBS), 2 mmol/L glutamine and 100 μg/mL penicillin/streptomycin in 5% CO2 at 37 °C. After grown to confluency in media, the cells were processed for subsequent studies.
MTT colorimetric assay
The viability/proliferation of the cells induced by LPS was observed using a MTT colorimetric assay as previously described[29]. MS1 cells received 0, 0.1, 1 and 10 μg/mL LPS in this experiment. Briefly, the cells were trypsinized with trypsin-ethylenediaminetetraacetic acid (EDTA), followed by incubation with DMEM in the presence of 10% FBS to inhibit trypsin activity. The cell pellets were then resuspended in DMEM with 10% FBS to a concentration of 1 × 104 cells/mL. Two hundred microliters of the cell suspension containing approximately 2000 cells was inoculated into selected wells of the 96-well plate. After the cells grew to 75% confluence, 20 μL of MTT solution was added to each well, and cultured for 4 h. Next, the medium was removed by inverting and tapping the plates, and 150 μL of dimethyl sulfoxide (DMSO) was added to each well. The spectrophotometric absorbance at 490 nm was measured by a Titertek Multiscan enzyme-linked immunosorbent assay reader. Each experiment was repeated at least three times. Every experimental condition was repeated at least in triplicate wells for each experiment.
Hoechst 33342/propidium iodide staining
The apoptosis and necrosis of the cells under the conditions described previously were observed by Hoechst 33342-propidium iodide (PI) staining[30]. MS1 cells were plated in a 6-well plate and co-incubated with media, LPS, LPS + DMSO and LPS + BN52021 when the cells achieved 90% confluence. The cells were washed twice with PBS. After the addition of 5 μL of Hoechst 33342 staining solution, the cells were stained with PI in the dark for 20-30 min at 4 °C and washed twice with PBS. Cells with blue and red fluorescence were examined under a fluorescence microscope.
Real-time quantitative RT-PCR
The mRNAs levels of AC, PLA2, PLCβ, PTK, GRK and p38 MAPK were measured by real-time PCR. In detail, MS1 cells were plated in a 6-well plate and co-incubated with media, LPS, LPS + DMSO and LPS + BN52021 when the cells achieved 90% confluence. The cells were collected, and the total RNA was extracted with a Trizol RNA reagent kit according to the manufacturer’s instructions. In addition, 2 μL (1 μg) of total RNA was added to the reverse transcription kit MIX system, and reverse-transcribed PCR was performed by random priming. The resulting complementary DNA amount was measured by quantitative PCR analysis using the GeneAmp 5700 Sequence Detection System and Step One Plus Real-Time PCR System (Applied Biosystems). The qPCR primer sequences are available online as indicated in Table 1. All expression data were normalized to the data for Gapdh. A no-template, double-distilled water control was included for each template. All samples were amplified simultaneously in triplicate in a single run. The relative quantitative gene expression was calculated as previously described and expressed as the percentage of the control level[31].
Table 1.
Specific primers for Adcy1, Pla2g4a, Plcb3, Ptk7, Adrbk1, Mapk14 and Gapdh genes
| Gene | Primer | Length (bp) | Annealing temperature (°C) |
| Adcy1 | |||
| Forward | 5'-GACTTTGTTCTCCGAGTTG-3' | 19 | 49 |
| Reverse | 5'-GTGCTATCCATCCGACTG-3' | ||
| Pla2g4a | |||
| Forward | 5'-GAATAAAGGCTCTACAATGG-3' | 20 | 49 |
| Reverse | 5'-GTTGTCGCTTTGGTACTC-3' | ||
| Plcb3 | |||
| Forward | 5'-CCTCAACTTCAACCGAGTT-3' | 19 | 49 |
| Reverse | 5'-CAGAGTGAGGTACGGCTTG-3' | ||
| Ptk7 | |||
| Forward | 5'-CACTGCGATGTCACATTG-3' | 18 | 49 |
| Reverse | 5'-CACTATGTTCGGGACTGG-3' | ||
| Adrbk1 | |||
| Forward | 5'-AAGCCAGCCAACATTCTC-3' | 18 | 51 |
| Reverse | 5'-CCCTTCTGTAGGACTTCG-3' | ||
| Mapk14 | |||
| Forward | 5'-GGACCTGAACAACATCGTG-3' | 19 | 50 |
| Reverse | 5'-CTAGGTTGCTGGGCTTTAG-3' | ||
| Gapdh | |||
| Forward | 5'-CATCTTCCAGGAGCGAGAC-3' | 19 | 50 |
| Reverse | 5'-GGCTAAGCAGTTGGTGGTG-3' |
Adcy1: Adenylate cyclase; Pla2g4a: Phospholipase A2; Plcb3: Phospholipase Cβ; Ptk7: Protein tyrosine kinase; Adrbk1: G protein-coupled receptor kinases; Mapk14: p38-mitogen-activated protein kinase; Gapdh: Glyceraldehyde 3-phosphate dehydrogenase.
Western blotting
The protein expression level of p-AC, p-PLA2, p-PTK, p-p38 MAPK, PLCβ and GRK was measured using Western blotting analysis. In detail, MS1 cells were plated in a 6-well plate and co-incubated with media, LPS, LPS + DMSO and LPS + BN52021 for 24 h when the cells achieved 90% confluence. The cells were washed twice with 0.1 mol/L PBS and then lysed in RIPA lysis buffer (Tris-HCl 10 mmol/L, pH 7.4; NaCl 0.15 mmol/L; EDTA 0.5 mmol/L; phenylmethylsulfonyl fluoride 10 mmol/L; Tritonx-100 1%; dithiothreitol 40 mmol/L). The protein concentration of the lysate was determined using a BCA protein assay kit (Beyotime Institute of Biotechnology, Beijing, China). Cell lysates containing 60 mg of protein were subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis using 12% polyacrylamide resolving gels. After electrophoresis, the proteins were transferred onto PVDF membranes, which were then blocked with 5% nonfat dry milk in TBS-0.05% Tween 20 (TBST) for 1 h at room temperature, washed in TBST for 10 min × 3, and incubated at 4 °C with gentle shaking overnight with rabbit primary antibodies against the protein of interest at corresponding dilutions, followed by incubation with horseradish peroxidase conjugated to goat anti-rabbit immunoglobulin G at 1:2000 dilution, incubation with 1 mL of enhanced chemiluminescence reagent for 3 min, and exposure to the film. The optical density of the protein of interest relative to that of β-actin was analyzed using Quantity One 4.6.2.
Statistical analysis
The data are expressed as the mean ± SE. The dose and time effects of LPS on the activity of MS1 cells were evaluated with a two-way analysis of variance (ANOVA). The differences between three or more groups were evaluated by one-way ANOVA. A P value less than 0.05 (2-tailed) was considered statistically significant. All tests were performed using the statistical software package GraphPad 5.0 (GraphPad Software Inc., San Diego, CA, United States).
RESULTS
Dose and time effect of LPS on MS1 cell activity
MS1 cells received 0, 0.1, 1 and 10 μg/mL LPS to mimic the inflammation condition of AP in vitro. The optimal dose and duration of LPS stimulation were determined using the MTT method. As shown in Figure 1, there was no significant difference in MS1 cell activity between cells co-incubated with the different concentrations of LPS and control cells 3 h after culture (P > 0.05), but when the incubation time was extended to 6 h, MS1 cell activity decreased significantly in the 1 μg/mL LPS group (0.49 ± 0.10 vs 0.67 ± 0.13, P < 0.05) and 10 μg/mL LPS group (0.44 ± 0.10 vs 0.67 ± 0.13, P < 0.001), but not in the 0.1 μg/mL group (P > 0.05) compared with the control group. When the incubation time was extended to 12 h (0.33 ± 0.05, 0.32 ± 0.03 and 0.25 ± 0.03 vs 0.69 ± 0.01) and 24 h (0.31 ± 0.01, 0.29 ± 0.03 and 0.25 ± 0.01 vs 0.63 ± 0.01), MS1 cell activity decreased in all LPS concentration groups compared with the blank control (P < 0.001). Therefore, we chose the concentration 10 μg/mL LPS for the 24 h stimulation as the optimal protocol in the following experiments.
Figure 1.
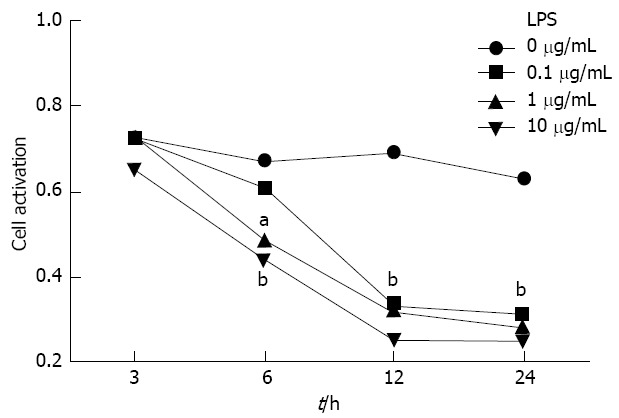
The optimal dose and duration of lipopolysaccharide stimulation were determined using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide method. The cell survival rate was determined after incubation with 0 (saline) and 0.1, 1 and 10 μg/mL lipopolysaccharide (LPS) for 3, 6, 12 and 24 h. aP < 0.05, bP < 0.01 vs the saline group.
Dose effect of BN52021 on LPS-induced inflammation
The dose effect of BN52021 on LPS-induced inflammation was determined using the MTT method and Hoechst 33342/PI staining. The MS1 cell activity was significantly decreased 24 h after administration of 10 μg/mL LPS compared with the control group (P < 0.01). Pretreatment with BN52021 20 min before incubation with LPS significantly improved the cell activity compared with the group receiving LPS only when its concentration reached 50 μmol/L, which was consistent with the results obtained by Hoechst 33342/PI staining (P < 0.05) (Figure 2). Therefore, the concentration of 50 μmol/L BN52021 was used for pretreatment in the following experiments.
Figure 2.
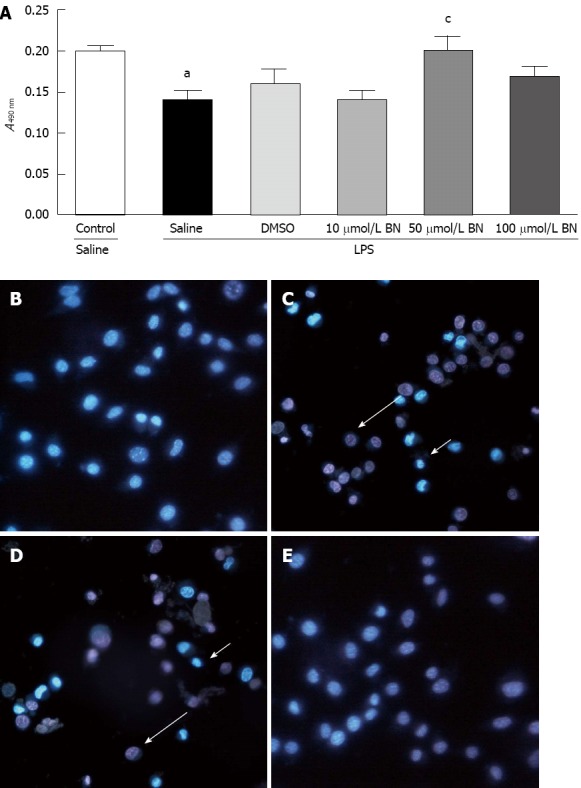
The dose effect of BN52021 on lipopolysaccharide-induced inflammation was determined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide method and Hoechst 33342/propidium iodide staining. MS1 cell activity at A490 nm was significantly decreased 24 h after administration of 10 μg/mL lipopolysaccharide (LPS) vs the control group (aP < 0.05). Pretreatment with BN52021 for 20 min before incubation with LPS significantly improved the MS1 cell activity at A490 nm vs the group that received LPS treatment only when its concentration reached 50 μmol/L (cP < 0.05) (A). Pretreatment with 50 μmol/L BN52021 for 20 min before incubation with LPS significantly improved MS1 cell activity vs the LPS + saline group, and the LPS + dimethyl sulfoxide (DMSO) group as determined Hoechst 33342/propidium iodide staining (B, C, D and E). The arrows indicate the apoptosis (short) and necrosis (long) of the cells.
Effect of BN52021 on PAFR signaling molecules at the mRNA level in LPS-induced inflammation
The mRNAs levels of AC (to 4.02 ± 0.14 folds), GRK (to 2.63 ± 0.03 folds), p38 MAPK (to 3.87 ± 0.07 folds), PLA2 (to 3.31 ± 0.12 folds), PLCβ (to 2.09 ± 0.08 folds) and PTK (to 1.85 ± 0.07 folds) were up-regulated after LPS stimulation compared with the blank control (P < 0.05). The up-regulated mRNAs were significantly suppressed by BN52021, except for that of PTK (fold-change relative to control, 1.83 ± 0.13, P > 0.05), including that of AC (fold-change relative to control, down to 2.35 ± 0.13), GRK (down to 1.17 ± 0.14), p38 MAPK (down to 1.49 ± 0.18), PLCβ (down to 2.09 ± 0.08) and PLA2 (down to 1.87 ± 0.11), as shown in Figure 3.
Figure 3.
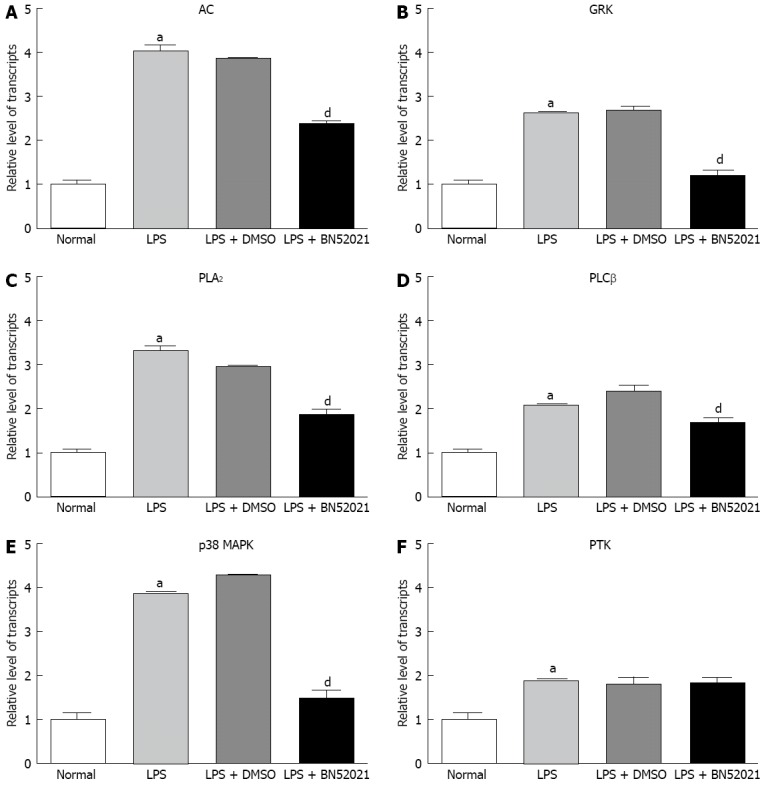
The effect of BN52021 on platelet-activating factor receptor signaling molecules at the mRNA level under lipopolysaccharide-induced inflammation. The mRNA level of adenylate cyclase (AC) (A), G protein-coupled receptor kinases (GRK) (B), phospholipase A2 (PLA2) (C), phospholipase Cβ (PLCβ) (D), p38-mitogen-activated protein kinase (p38 MAPK) (E) and protein tyrosine kinase (PTK) (F) was up-regulated after lipopolysaccharide (LPS) stimulation. The up-regulation of AC, GRK, p38 MAPK, PLCβ and PLA2 mRNA was significantly suppressed by BN52021 except for that of PTK. aP < 0.05 vs control; dP < 0.01 vs the LPS + dimethyl sulfoxide (DMSO) groups.
Effect of BN52021 on PAFR signaling molecules at the protein level in LPS-induced inflammation
The level of p-AC (fold-change relative to control, increase from 0.65 ± 0.08 to 1.11 ± 0.12), GRK (increase from 0.50 ± 0.03 to 0.83 ± 0.07), PLCβ (increase from 0.50 ± 0.10 to 0.83 ± 0.16) and p-P38 MAPK (increase from 0.38 ± 0.05 to 0.74 ± 0.10) was up-regulated after LPS stimulation compared with the blank control (P < 0.05). The up-regulated protein level was significantly suppressed by BN52021 for p-AC (decrease from 1.11 ± 0.12 to 0.65 ± 0.15), GRK (decrease from 0.83 ± 0.07 to 0.47 ± 0.10), PLCβ (decrease from 0.83 ± 0.16 to 0.47 ± 0.04) and p-p38 MAPK (decrease from 0.74 ± 0.10 to 0.30 ± 0.10). However, the level of p-PLA2 and p-PTK was not significantly up-regulated after LPS stimulation and was not significantly altered by BN52021, as shown in Figure 4.
Figure 4.

The effect of BN52021 on platelet-activating factor receptor signaling molecules at the protein level under lipopolysaccharide-induced inflammation. The protein level of p-adenylate cyclase (p-AC) (A), G protein-coupled receptor kinases (GRK) (B), p-phospholipase A2 (p-PLA2) (C), phospholipase Cβ (PLCβ) (D) and p-p38-mitogen-activated protein kinase (p-p38 MAPK) (E) was up-regulated after lipopolysaccharide (LPS) stimulation vs the blank control (aP < 0.05). The up-regulation of p-AC, p-p38 MAPK, GRK and PLCβ protein levels was significantly suppressed by BN52021. However, p-PLA2 and phosphorylated protein tyrosine kinase (p-PTK) protein levels were insignificantly up-regulated after LPS stimulation and were not significantly changed by BN52021 (F). cP < 0.05, dP < 0.01 vs LPS + dimethyl sulfoxide (DMSO) groups.
DISCUSSION
In this study, we examined the signaling molecules of the PAFR pathway to evaluate whether the PAFR antagonist BN52021 had any influence on LPS-induced inflammation in MS1 cells. It was observed that BN52021 could sufficiently inhibit the inflammation, apoptosis and necrosis induced by LPS in pancreatic vascular endothelial cells. BN52021 could inhibit the up-regulation of signaling molecules in the PAFR pathway, which may help to explaining the mechanism underlying microcirculatory disturbance in the pathogenesis of AP.
PAF-induced microcirculatory disruption plays a key role in the pathogenesis of AP
Platelet-activating factor is a proinflammatory lipid mediator that plays a key role in many pathophysiological conditions, including asthma, ischemia, gastrointestinal ulceration, pancreatitis and multiple organ failure[32]. A number of experimental studies suggest that the pathogenesis of AP correlates with microcirculatory disorders. An experiment that constricted interlobular pancreatic arteries 2 min after intraductal infusion of sodium taurocholate indicated that microcirculatory changes are closely related to the process of AP[6]. Many complications of SAP are due to the amplifying effect of microcirculatory disruption[7]. PAF is one of the most important vasoactive mediators activated during the inflammatory response to pancreatic injury that can cause microcirculatory disorders in AP. Recent data suggest that PAF can directly modulate microvascular permeability and increase venular permeability[10]. Increased microvessel permeability induced by PAF may be related directly to endothelial cell activation, adhesion molecule expression, and leukocyte activation[7,8]. Increased capillary permeability permits the sequestration of macromolecules and fluid, causing deficiency of circulating blood volume and microcirculatory disorders[7]. In addition, vasospasm and microthrombus formation due to hypercoagulability can also lead to the deterioration of pancreatic microcirculation and pancreatic necrosis[7]. The treatment of AP with PAF antagonists can significantly improve capillary blood flow in the pancreas and colon, renal and respiratory function, and the survival rate and can stabilize capillary permeability and decrease fluid loss into the third space[33,34]. As a preventive treatment, PAFR antagonists such as BN52021 can block a series of PAF-mediated inflammatory injuries, thus improving the prognosis of AP[1]. This protective effect of PAF antagonists further supports the role of PAF in microcirculatory disorders.
LPS-induced inflammation of pancreatic microvascular endothelial cells is a suitable pancreatitis model to simulate microcirculatory disturbance in vitro. The MS1 cell line is a mouse pancreatic islet endothelial cell line first established in 1994. It can represent the pancreatic microvascular endothelium because previous studies[35] have verified that the pancreatic lobule is a structured and functional basic unit of pancreatic microcirculation, and insulo-acinar portal circulation represents the basic feature of the pancreatic microcirculation. Therefore, in this study, we examined the signaling molecules of the PAFR pathway in MS1 cells to evaluate whether the PAF receptor antagonist BN52021 had any influence on the LPS-induced inflammatory effect, hoping that it could help elucidate the mechanism underlying microcirculatory disturbance in the pathogenesis of SAP in vitro. Our results indicated that pretreatment with BN52021 for 20 min before incubation with LPS could significantly improve the MS1 cell activity compared with the group that received LPS treatment only.
PAFR signaling pathway plays a pivotal role in pancreatic proinflammatory response
In recent years, researchers have become concerned with the significance of the signal transduction pathway of PAF in the pathogenesis of AP[4,22,28], because it has been reported to induce morbidity and unacceptably high mortality[18]. However, the impact of a PAF receptor antagonist (BN52021) on the signaling molecules of the PAFR signaling pathway in pancreatic microvascular endothelial cells under the LPS-induced inflammatory condition remains unclear.
PAFR is almost ubiquitous in diverse type cells and acts not only on the local pancreas cells, including the pancreatic vascular endothelium, but also on distant organs, inducing systemic inflammatory response and multiple organ injury[15]. PAFR belongs to the G protein-coupled receptor subfamily[36]. By binding to its receptor, PAF activates the associated G protein, which, in turn, activates phosphoinositide hydrolysis by phosphoinositide specific phospholipase C, arachidonic acid release by phospholipase A2, increases in intracellular Ca2+ concentration, activation of protein kinase C and PTK[37]. PAF has also been shown to activate MAPKs, including extracellular signal-regulated kinase[38-42], p38 MAPK[38,40,41], and c-Jun N-terminal kinase[43]. Deo et al[44] reported that PAF activated pertussis toxin-insensitive Gαq protein upon binding to its seven transmembrane receptors and adenylate cyclase, elevating cAMP levels, and thus activating protein kinase A in human umbilical vein endothelial cells. GRK plays a key role in the homologous desensitization of G protein-coupled receptor (GPCR) and GRK phosphorylate activated receptors, promoting high affinity binding of arrestins, thus precluding G protein coupling. Direct binding to active GPCRs activates GRKs so that they selectively phosphorylate only the activated form of the receptor regardless of the accessibility of the substrate peptides within it and their Ser/Thr-containing sequence[45]. Most GPCRs display a rapid loss of responsiveness in the continuing presence of chemoattractants in a process of desensitization that involves the phosphorylation of agonist-occupied GPCR by GRK[46]. The inflammation in pancreatic vascular endothelial cells induced by LPS was suppressed by BN52021. This finding might contribute to an understanding of the mechanism underlying the microcirculatory disturbances in the pathogenesis of SAP.
According to our results, the mRNA and protein levels of AC, GRK, p38 MAPK, PLA2 and PTK were up-regulated after LPS stimulation compared with the blank control. The up-regulated AC, GRK, p38 MAPK and PLA2 mRNA and protein levels were significantly suppressed by BN52021, suggesting that BN52021 could effectively inhibit the apoptosis and necrosis of MS1 cells under the LPS-induced inflammatory condition. The mechanism underlying the inhibition might relate to the suppression effect of BN52021 on the up-regulation of AC, GRK, p38 MAPK and PLA2 mRNA and protein levels in the PAFR signaling pathway.
Other potential mechanisms of PAFR antagonism in AP treatment
It is known that PAFR is also able to interact with components of the bacterial wall, such as lipopolysaccharides[47] and phosphorylcholine[48]. The cell wall components exit the vasculature into the heart and brain, accumulating within endothelial cells, cardiomyocytes, and neurons in a PAFR-dependent way. The physiological consequences of the cell wall/PAFR interaction are cell specific, being noninflammatory in endothelial cells and neurons but causing a rapid loss of cardiomyocyte contractility that contributes to death. Thus, PAFR shepherds phosphorylcholine-containing bacterial components such as the cell wall into host cells from where the response ranges from quiescence to severe pathophysiology[48]. The explanation for the protective effect of BN-52021 cannot simply be attributed to the antagonism of LPS binding to PAFR or the prevention of PAF binding to its receptor. Therefore other potential mechanisms of PAFR antagonism in AP treatment must exist.
Bacterial translocation from the gastrointestinal tract to mesenteric lymph nodes and other extra intestinal organs is an important source of infection in AP. Preventing bacterial dissemination in early AP may have beneficial effects on the evolution of this disease[26,49]. PAF antagonist treatment decreases the bacterial spread to distant sites, suppresses elevation of interleukin (IL)-6 level, and has a significant effect on serum pancreatic enzymes and the histologic score of pancreatitis without reducing serum amylase and tumor necrosis factor alpha levels or ameliorating pancreatic damage in rats with AP[7,50]. In addition, BN52021 has been shown to have protective effect on slow mesenterioangial small arteriolar and venular blood flow velocity and dilated mesenterioangial small venular diameter in the early phase of AP[51]. Pretreatment with lexipafant could reduce the pancreatic endothelial barrier dysfunction and severity of pancreatitis-associated intestinal dysfunction as well as systemic concentrations of IL-1 and local leukocyte recruitment in experimental AP rats[52-54]. PAFR antagonism appears to be involved in the maintenance of intestinal barrier integrity and the inhibition of cytokines release, such as IL-1 and IL-6[32]. Moreover, PAFR antagonists can also exert their effects by inhibiting the activity of neutrophils and depressing pulp peroxidase, competing for targets with PAF and inhibiting the activity of PAF, inhibiting increases in PAF in AP, and reducing plasma cytokines and inflammatory mediators, enzyme activity and the role of self-digestion of pancreatic tissue[1]. The involvement of the PAFR signaling pathway in these mechanisms needs to be further investigated.
The PAFR antagonist BN52021 could effectively inhibit LPS-induced inflammation, apoptosis and necrosis in pancreatic vascular endothelial cells. The mechanisms underlying the inhibition might be related to the suppression effect of BN52021 on the up-regulation of AC, GRK, p38 MAPK and PLCβ mRNA and protein levels in the PAFR signaling pathway, which may help to explain the mechanism underlying the microcirculatory disturbance in the pathogenesis of AP.
COMMENTS
Background
Microcirculatory disorder is considered to be one of the possible mechanisms of severe acute pancreatitis (SAP) pathogenesis. Platelet-activating factor (PAF), a bioactive phospholipid synthesized and secreted by a variety of cells including pancreatic acini and microvascular endothelium cells, is known to mediate many physiological responses, including microcirculatory disturbance and inflammation.
Research frontiers
Recent studies have demonstrated that PAF plays an important role in the pathological progress of SAP. Although BN52021, a PAF receptor antagonist, has demonstrated significant treatment effects against SAP, its effects on PAF receptor (PAFR) signaling molecules have not been elucidated in detail.
Innovations and breakthroughs
The authors found that BN52021 could effectively inhibit the apoptosis and necrosis of MS1 cells under lipopolysaccharide (LPS)-induced inflammatory conditions. The mechanism underlying the inhibitory effect may relate to the inhibitory effect of BN52021 on the up-regulation of adenylate cyclase, G protein-coupled receptor kinases, p38-mitogen-activated protein kinase, phospholipase A2 and phospholipase Cβ mRNA and protein levels in the PAFR signaling pathway.
Applications
This study may contribute to a future strategy involving SAP treatment with BN52021 by investigating how PAF is induced and blocking its expression.
Terminology
PAF is a biologically active phospholipid mediator that plays its role by binding to PAFR, which is a unique G-protein-coupled seven transmembrane receptor, and the binding activates multiple intracellular signaling pathways. Ginkgolide B (code: BN52021) is one of the four Ginkgolide constituents (Ginkgolide A, B, C and J) that are present in the whole extract of Ginkgo biloba leaves.
Peer review
This article attempts to elucidate the protective role of BN-52021 against LPS-induced apoptosis and necrosis in a pancreatic islet endothelial cell line. PAF is a crucial mediator of acute pancreatitis. Therefore the inhibition of its actions by BN-52021 is interesting from a pharmaceutical point of view. Because BN-52021 is a well-established antagonist of PAFR, the authors investigated its effect on certain members of the signal transduction pathways initiated by PAFR activation. The results are novel and interesting.
Footnotes
Supported by The National Natural Science Foundation of China, No. 81173393; the Natural Science Foundation of Tianjin City, Grant No. 12YFJZJC00800; the Scientific Research Foundation for PhD grant to Xia SH, No. WYB201010; and the Innovation Team Program (WHTD201310) from the Logistics University of the Chinese People’s Armed Police Force
P- Reviewers Nomikos T, Sahu RP S- Editor Wen LL L- Editor A E- Editor Li JY
References
- 1.Chen C, Xia SH, Chen H, Li XH. Therapy for acute pancreatitis with platelet-activating factor receptor antagonists. World J Gastroenterol. 2008;14:4735–4738. doi: 10.3748/wjg.14.4735. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Büchler MW, Gloor B, Müller CA, Friess H, Seiler CA, Uhl W. Acute necrotizing pancreatitis: treatment strategy according to the status of infection. Ann Surg. 2000;232:619–626. doi: 10.1097/00000658-200011000-00001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Malangoni MA, Martin AS. Outcome of severe acute pancreatitis. Am J Surg. 2005;189:273–277. doi: 10.1016/j.amjsurg.2004.11.013. [DOI] [PubMed] [Google Scholar]
- 4.Xia SH, Hu CX, Fang JM, Di Y, Zhao ZL, Liu LR. G[alpha]i2 and G[alpha]q expression change in pancreatic tissues and BN52021 effects in rats with severe acute pancreatitis. Pancreas. 2008;37:170–175. doi: 10.1097/MPA.0b013e3181661b07. [DOI] [PubMed] [Google Scholar]
- 5.Cuthbertson CM, Christophi C. Disturbances of the microcirculation in acute pancreatitis. Br J Surg. 2006;93:518–530. doi: 10.1002/bjs.5316. [DOI] [PubMed] [Google Scholar]
- 6.Kusterer K, Poschmann T, Friedemann A, Enghofer M, Zendler S, Usadel KH. Arterial constriction, ischemia-reperfusion, and leukocyte adherence in acute pancreatitis. Am J Physiol. 1993;265:G165–G171. doi: 10.1152/ajpgi.1993.265.1.G165. [DOI] [PubMed] [Google Scholar]
- 7.Liu LR, Xia SH. Role of platelet-activating factor in the pathogenesis of acute pancreatitis. World J Gastroenterol. 2006;12:539–545. doi: 10.3748/wjg.v12.i4.539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Montrucchio G, Lupia E, De Martino A, Silvestro L, Savu SR, Cacace G, De Filippi PG, Emanuelli G, Camussi G. Plasmin promotes an endothelium-dependent adhesion of neutrophils. Involvement of platelet activating factor and P-selectin. Circulation. 1996;93:2152–2160. doi: 10.1161/01.CIR.93.12.2152. [DOI] [PubMed] [Google Scholar]
- 9.Sugimoto M, Takada T, Yasuda H. A new experimental pancreatitis by incomplete closed duodenal loop: the influence of pancreatic microcirculation on the development and progression of induced severe pancreatitis in rats. Pancreas. 2004;28:e112–e119. doi: 10.1097/00006676-200405000-00023. [DOI] [PubMed] [Google Scholar]
- 10.Victorino GP, Newton CR, Curran B. Modulation of microvascular hydraulic permeability by platelet-activating factor. J Trauma. 2004;56:379–384. doi: 10.1097/01.TA.0000042156.89779.6C. [DOI] [PubMed] [Google Scholar]
- 11.Zhou W, Levine BA, Olson MS. Platelet-activating factor: a mediator of pancreatic inflammation during cerulein hyperstimulation. Am J Pathol. 1993;142:1504–1512. [PMC free article] [PubMed] [Google Scholar]
- 12.Marrache AM, Gobeil F, Bernier SG, Stankova J, Rola-Pleszczynski M, Choufani S, Bkaily G, Bourdeau A, Sirois MG, Vazquez-Tello A, et al. Proinflammatory gene induction by platelet-activating factor mediated via its cognate nuclear receptor. J Immunol. 2002;169:6474–6481. doi: 10.4049/jimmunol.169.11.6474. [DOI] [PubMed] [Google Scholar]
- 13.Emanuelli G, Montrucchio G, Gaia E, Dughera L, Corvetti G, Gubetta L. Experimental acute pancreatitis induced by platelet activating factor in rabbits. Am J Pathol. 1989;134:315–326. [PMC free article] [PubMed] [Google Scholar]
- 14.Brown SL, Jala VR, Raghuwanshi SK, Nasser MW, Haribabu B, Richardson RM. Activation and regulation of platelet-activating factor receptor: role of G(i) and G(q) in receptor-mediated chemotactic, cytotoxic, and cross-regulatory signals. J Immunol. 2006;177:3242–3249. doi: 10.4049/jimmunol.177.5.3242. [DOI] [PubMed] [Google Scholar]
- 15.Flickinger BD, Olson MS. Localization of the platelet-activating factor receptor to rat pancreatic microvascular endothelial cells. Am J Pathol. 1999;154:1353–1358. doi: 10.1016/S0002-9440(10)65389-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liu H, Chao W, Olson MS. Regulation of the surface expression of the platelet-activating factor receptor in IC-21 peritoneal macrophages. Effects of lipopolysaccharide. J Biol Chem. 1992;267:20811–20819. [PubMed] [Google Scholar]
- 17.Xia SH, Hu CX, Zhao ZL, Xia GD, Di Y. Significance of platelet activating factor receptor expression in pancreatic tissues of rats with severe acute pancreatitis and effects of BN52021. World J Gastroenterol. 2007;13:2992–2998. doi: 10.3748/wjg.v13.i21.2992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Miike S, Kurasawa K, Saito Y, Iwamoto I. Platelet-activating factor activates mitogen-activated protein kinases through the activation of phosphatidylinositol 3-kinase and tyrosine kinase in human eosinophils. J Leukoc Biol. 2000;67:117–126. doi: 10.1002/jlb.67.1.117. [DOI] [PubMed] [Google Scholar]
- 19.Kingsnorth AN. Platelet-activating factor. Scand J Gastroenterol Suppl. 1996;219:28–31. doi: 10.3109/00365529609104996. [DOI] [PubMed] [Google Scholar]
- 20.Johnson CD. Platelet-activating factor and platelet-activating factor antagonists in acute pancreatitis. Dig Surg. 1999;16:93–101. doi: 10.1159/000018699. [DOI] [PubMed] [Google Scholar]
- 21.Mauri P, Simonetti P, Gardana C, Minoggio M, Morazzoni P, Bombardelli E, Pietta P. Liquid chromatography/atmospheric pressure chemical ionization mass spectrometry of terpene lactones in plasma of volunteers dosed with Ginkgo biloba L. extracts. Rapid Commun Mass Spectrom. 2001;15:929–934. doi: 10.1002/rcm.316. [DOI] [PubMed] [Google Scholar]
- 22.Ji RL, Xia SH, Di Y, Xu W. Mechanism and dose-effect of Ginkgolide B on severe acute pancreatitis of rats. World J Gastroenterol. 2011;17:2241–2247. doi: 10.3748/wjg.v17.i17.2241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Langley SM, Chai PJ, Jaggers JJ, Ungerleider RM. Platelet-activating factor receptor antagonism improves cerebral recovery after circulatory arrest. Ann Thorac Surg. 1999;68:1578–1584; discussion 1585. doi: 10.1016/S0003-4975(99)00998-4. [DOI] [PubMed] [Google Scholar]
- 24.Dabrowski A, Gabryelewicz A, Chyczewski L. The effect of platelet activating factor antagonist (BN 52021) on cerulein-induced acute pancreatitis with reference to oxygen radicals. Int J Pancreatol. 1991;8:1–11. doi: 10.1007/BF02930218. [DOI] [PubMed] [Google Scholar]
- 25.Jancar S, Abdo EE, Sampietre SN, Kwasniewski FH, Coelho AM, Bonizzia A, Machado MC. Effect of PAF antagonists on cerulein-induced pancreatitis. J Lipid Mediat Cell Signal. 1995;11:41–49. doi: 10.1016/0929-7855(94)00026-9. [DOI] [PubMed] [Google Scholar]
- 26.Bedirli A, Gokahmetoglu S, Sakrak O, Soyuer I, Ince O, Sozuer E. Beneficial effects of recombinant platelet-activating factor acetylhydrolase and BN 52021 on bacterial translocation in cerulein-induced pancreatitis. Eur Surg Res. 2004;36:136–141. doi: 10.1159/000077254. [DOI] [PubMed] [Google Scholar]
- 27.McKenna DJ, Jones K, Hughes K. Efficacy, safety, and use of ginkgo biloba in clinical and preclinical applications. Altern Ther Health Med. 2001;7:70–86, 88-90. [PubMed] [Google Scholar]
- 28.Xia SH, Fang DC, Hu CX, Bi HY, Yang YZ, Di Y. Effect of BN52021 on NFkappa-Bp65 expression in pancreatic tissues of rats with severe acute pancreatitis. World J Gastroenterol. 2007;13:882–888. doi: 10.3748/wjg.v13.i6.882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sun HY, Wei SP, Xu RC, Xu PX, Zhang WC. Sphingosine-1-phosphate induces human endothelial VEGF and MMP-2 production via transcription factor ZNF580: novel insights into angiogenesis. Biochem Biophys Res Commun. 2010;395:361–366. doi: 10.1016/j.bbrc.2010.04.019. [DOI] [PubMed] [Google Scholar]
- 30.McKeague AL, Wilson DJ, Nelson J. Staurosporine-induced apoptosis and hydrogen peroxide-induced necrosis in two human breast cell lines. Br J Cancer. 2003;88:125–131. doi: 10.1038/sj.bjc.6600675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Ding JL, Zhou ZG, Zhou XY, Zhou B, Wang L, Wang R, Zhan L, Sun XF, Li Y. Attenuation of acute pancreatitis by peroxisome proliferator-activated receptor-α in rats: the effect on Toll-like receptor signaling pathways. Pancreas. 2013;42:114–122. doi: 10.1097/MPA.0b013e3182550cc4. [DOI] [PubMed] [Google Scholar]
- 32.Sun Z, Wang X, Lasson A, Börjesson A, Leveau P, Haraldsen P, Andersson R. Roles of platelet-activating factor, interleukin-1beta and interleukin-6 in intestinal barrier dysfunction induced by mesenteric arterial ischemia and reperfusion. J Surg Res. 1999;87:90–100. doi: 10.1006/jsre.1999.5746. [DOI] [PubMed] [Google Scholar]
- 33.Eibl G, Buhr HJ, Foitzik T. Therapy of microcirculatory disorders in severe acute pancreatitis: what mediators should we block? Intensive Care Med. 2002;28:139–146. doi: 10.1007/s00134-001-1194-1. [DOI] [PubMed] [Google Scholar]
- 34.Foitzik T, Hotz HG, Eibl G, Hotz B, Kirchengast M, Buhr HJ. Therapy for microcirculatory disorders in severe acute pancreatitis: effectiveness of platelet-activating factor receptor blockade vs. endothelin receptor blockade. J Gastrointest Surg. 1999;3:244–251. doi: 10.1016/S1091-255X(99)80066-3. [DOI] [PubMed] [Google Scholar]
- 35.Zhou Z, Zeng Y, Yang P, Cheng Z, Zhao J, Shu Y, Gao X, Yan L, Zhang Z. Structure and function of pancreatic microcirculation. Shengwu Yixue Gongchengxue Zazhi. 2001;18:195–200. [PubMed] [Google Scholar]
- 36.Izumi T, Shimizu T. Platelet-activating factor receptor: gene expression and signal transduction. Biochim Biophys Acta. 1995;1259:317–333. doi: 10.1016/0005-2760(95)00171-9. [DOI] [PubMed] [Google Scholar]
- 37.Liu B, Nakashima S, Kanoh H, Takano T, Shimizu T, Nozawa Y. Activation of phospholipase D in Chinese hamster ovary cells expressing platelet-activating factor receptor. J Biochem. 1994;116:882–891. doi: 10.1093/oxfordjournals.jbchem.a124611. [DOI] [PubMed] [Google Scholar]
- 38.Chen LW, Lin MW, Hsu CM. Different pathways leading to activation of extracellular signal-regulated kinase and p38 MAP kinase by formyl-methionyl-leucyl-phenylalanine or platelet activating factor in human neutrophils. J Biomed Sci. 2005;12:311–319. doi: 10.1007/s11373-005-1704-1. [DOI] [PubMed] [Google Scholar]
- 39.Coffer PJ, Geijsen N, M’rabet L, Schweizer RC, Maikoe T, Raaijmakers JA, Lammers JW, Koenderman L. Comparison of the roles of mitogen-activated protein kinase kinase and phosphatidylinositol 3-kinase signal transduction in neutrophil effector function. Biochem J. 1998;329(Pt 1):121–130. doi: 10.1042/bj3290121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Marques SA, Dy LC, Southall MD, Yi Q, Smietana E, Kapur R, Marques M, Travers JB, Spandau DF. The platelet-activating factor receptor activates the extracellular signal-regulated kinase mitogen-activated protein kinase and induces proliferation of epidermal cells through an epidermal growth factor-receptor-dependent pathway. J Pharmacol Exp Ther. 2002;300:1026–1035. doi: 10.1124/jpet.300.3.1026. [DOI] [PubMed] [Google Scholar]
- 41.Nick JA, Avdi NJ, Young SK, Knall C, Gerwins P, Johnson GL, Worthen GS. Common and distinct intracellular signaling pathways in human neutrophils utilized by platelet activating factor and FMLP. J Clin Invest. 1997;99:975–986. doi: 10.1172/JCI119263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Shimizu T, Mori M, Bito H, Sakanaka C, Tabuchi S, Aihara M, Kume K. Platelet-activating factor and somatostatin activate mitogen-activated protein kinase (MAP kinase) and arachidonate release. J Lipid Mediat Cell Signal. 1996;14:103–108. doi: 10.1016/0929-7855(96)00515-9. [DOI] [PubMed] [Google Scholar]
- 43.DeCoster MA, Mukherjee PK, Davis RJ, Bazan NG. Platelet-activating factor is a downstream messenger of kainate-induced activation of mitogen-activated protein kinases in primary hippocampal neurons. J Neurosci Res. 1998;53:297–303. doi: 10.1002/(SICI)1097-4547(19980801)53:3<297::AID-JNR3>3.0.CO;2-5. [DOI] [PubMed] [Google Scholar]
- 44.Deo DD, Bazan NG, Hunt JD. Activation of platelet-activating factor receptor-coupled G alpha q leads to stimulation of Src and focal adhesion kinase via two separate pathways in human umbilical vein endothelial cells. J Biol Chem. 2004;279:3497–3508. doi: 10.1074/jbc.M304497200. [DOI] [PubMed] [Google Scholar]
- 45.Mushegian A, Gurevich VV, Gurevich EV. The origin and evolution of G protein-coupled receptor kinases. PLoS One. 2012;7:e33806. doi: 10.1371/journal.pone.0033806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Alves-Filho JC, de Freitas A, Spiller F, Souto FO, Cunha FQ. The role of neutrophils in severe sepsis. Shock. 2008;30 Suppl 1:3–9. doi: 10.1097/SHK.0b013e3181818466. [DOI] [PubMed] [Google Scholar]
- 47.Nakamura M, Honda Z, Waga I, Matsumoto T, Noma M, Shimizu T. Endotoxin transduces Ca2 signaling via platelet-activating factor receptor. FEBS Lett. 1992;314:125–129. doi: 10.1016/0014-5793(92)80957-I. [DOI] [PubMed] [Google Scholar]
- 48.Fillon S, Soulis K, Rajasekaran S, Benedict-Hamilton H, Radin JN, Orihuela CJ, El Kasmi KC, Murti G, Kaushal D, Gaber MW, et al. Platelet-activating factor receptor and innate immunity: uptake of gram-positive bacterial cell wall into host cells and cell-specific pathophysiology. J Immunol. 2006;177:6182–6191. doi: 10.4049/jimmunol.177.9.6182. [DOI] [PubMed] [Google Scholar]
- 49.de Souza LJ, Sampietre SN, Assis RS, Knowles CH, Leite KR, Jancar S, Monteiro Cunha JE, Machado MC. Effect of platelet-activating factor antagonists (BN-52021, WEB-2170, and BB-882) on bacterial translocation in acute pancreatitis. J Gastrointest Surg. 2001;5:364–370. doi: 10.1016/S1091-255X(01)80063-9. [DOI] [PubMed] [Google Scholar]
- 50.Liu Q, Djuricin G, Rossi H, Bewsey K, Nathan C, Gattuso P, Weinstein RA, Prinz RA. The effect of lexipafant on bacterial translocation in acute necrotizing pancreatitis in rats. Am Surg. 1999;65:611–616; discussion 617. doi: 10.1016/s0016-5085(98)85715-6. [DOI] [PubMed] [Google Scholar]
- 51.Ji Z, Wang B, Li S. The role of platelet activating factor in mesenterioangial microcirculatory disturbance complicated with acute pancreatitis in rats. Zhonghua Yixue Zazhi. 1995;75:139–140, 188. [PubMed] [Google Scholar]
- 52.Leveau P, Wang X, Sun Z, Börjesson A, Andersson E, Andersson R. Severity of pancreatitis-associated gut barrier dysfunction is reduced following treatment with the PAF inhibitor lexipafant. Biochem Pharmacol. 2005;69:1325–1331. doi: 10.1016/j.bcp.2005.01.023. [DOI] [PubMed] [Google Scholar]
- 53.Wang X, Sun Z, Börjesson A, Andersson R. Inhibition of platelet-activating factor, intercellular adhesion molecule 1 and platelet endothelial cell adhesion molecule 1 reduces experimental pancreatitis-associated gut endothelial barrier dysfunction. Br J Surg. 1999;86:411–416. doi: 10.1046/j.1365-2168.1999.01028.x. [DOI] [PubMed] [Google Scholar]
- 54.Wang X, Sun Z, Börjesson A, Haraldsen P, Aldman M, Deng X, Leveau P, Andersson R. Treatment with lexipafant ameliorates the severity of pancreatic microvascular endothelial barrier dysfunction in rats with acute hemorrhagic pancreatitis. Int J Pancreatol. 1999;25:45–52. doi: 10.1385/IJGC:25:1:45. [DOI] [PubMed] [Google Scholar]