

Abstract
Prompt and accurate diagnosis of malarial patients is a crucial factor in controlling the morbidity and mortality of the disease. Effective treatment decisions require a correct diagnosis among mixed-species malarial patients. Differential diagnosis is particularly important in cases of Plasmodium vivax, a species that shares endemicity with P. falciparum in most endemic areas. Moreover, it is difficult to identify P. knowlesi on the basis of morphology alone, and rapid diagnostic tests are still not available for this malaria species. Therefore, the development of diagnostic tests applicable to the field is urgently needed. 1-Cys peroxiredoxin (1-Cys-Prx) in P. falciparum is abundantly expressed in the mature asexual stages, making it a promising candidate as a diagnostic antigen. In this study, we produced five monoclonal antibodies (mAbs) against P. falciparum 1-Cys-Prx (Pf1-Cys-Prx) by immunizing BALB/c mice with recombinant Pf1-Cys-Prx and subsequent hybridoma production. Cross reactivity of established mAbs with the orthologous molecule of Pf1-Cys-Prx in P. vivax (Pv1-Cys-Prx) and P. knowlesi (Pk1-Cys-Prx) was examined. Western blot analyses showed that three mAbs reacted with Pv1-Cys-Prx and Pk1-Cys-Prx but two mAbs did not. These results indicate that the two mAbs were effective in differentiating P. falciparum from P. vivax and P. knowlesi and could be used in differential diagnosis as well as comparative molecular studies of human Plasmodium species.
Keywords: malaria diagnosis, monoclonal antibody, peroxiredoxin, Plasmodium knowlesi, P. vivax
Introduction
Malaria is still considered to be one the most serious infectious diseases in the world. There were about 216 million cases of clinical malaria in 2010, 655,000 of which led to malaria related deaths [1]. Prompt and accurate diagnosis of malarial patients is fundamental in reducing the morbidity and mortality of the disease, avoiding unnecessary use of antimalarial agents and minimizing the spread of resistance to antimalarial drugs. Malaria elimination has been considered in countries with declining human malaria cases, and 32 countries have been categorized as malaria-eliminating countries in the summary report of Feachem et al. [2]. There is a need, therefore, to improve current diagnostic techniques as well as to address the new challenges in malaria control, including the presence of asymptomatic Plasmodium vivax carriers [3].
P. knowlesi, “the fifth human malaria parasite,” is originally a parasite of Old World monkeys, and naturally acquired human infections were believed to be very rare and sporadic until a few years ago [4]. Several studies conducted in various countries in Southeast Asia showed that P. knowlesi human malaria was underestimated and misdiagnosed because of morphological similarities with P. falciparum and P. malariae [5, 6], underlining the difficulty of identifying knowlesi malaria on the basis of morphology alone [6]. The lack of any available rapid diagnostic test to detect P. knowlesi further indicates the urgent need to develop a diagnostic test applicable to the field.
Peroxiredoxins (Prxs) are cysteine dependent antioxidant enzymes which are ubiquitous and frequently present in abundance from prokaryotes to eukaryotes [7]. The critical role and abundance of Prxs in parasites makes them potentially valuable candidates for drugs and vaccines as well as diagnostic targets [8]. It has been shown that Prx may have diagnostic value for the detection of Leishmania spp., Echinococcus granulosus, Fasciola gigantica, Taenia spp. [8] and Schistosoma japonicum [9]. 1-Cys peroxiredoxin (1-Cys-Prx) in P. falciparum, the orthologous molecule of human Prx VI, is abundantly expressed in the trophozoite stage of P. falciparum [10] making it a promising candidate as a diagnostic antigen. 1-Cys-Prx in P. falciparum, P. vivax and P. knowlesi has already been characterized by our group [11, 12]. In this study, we produced several monoclonal antibodies (mAbs) against P. falciparum 1-Cys-Prx (Pf1-Cys-Prx) and checked their cross reactivity with the orthologous molecule in P. vivax and P. knowlesi.
Materials and Methods
Recombinant protein production and purification
Recombinant proteins were produced as described previously [12, 13]. Briefly, the PCR products were digested with restriction enzymes (Supplementary Table S1) and then inserted into pET-28a expression vector (EMD Biosciences, San Diego, CA) by overnight incubation with ligation mixture (Takara, Otsu, Japan) at 15°C. The recombinant plasmids containing the coding sequence of Pf1-Cys-Prx, P. vivax 1-Cys-Prx (Pv1-Cys-Prx) and P. knowlesi 1-Cys-Prx (Pk1-Cys-Prx) were transfected into Escherichia coli (strain BL21). Expression of the recombinant proteins (rPf1-Cys-Prx, rPv1-Cys-Prx and rPk1-Cys-Prx) as a fusion protein with N-terminal histidine-tag was induced by adding 1 mM isopropyl thio-β-D-galactoside (IPTG) and purified using Ni-NTA agarose beads (Qiagen Inc., Valencia, CA). The expression and purification of the recombinant proteins were evaluated by 12% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS/PAGE) under reducing conditions and subsequent Coomassie Brilliant Blue R-250 (CBB) staining. The concentration of each expressed protein was measured using bicinchoninic acid (BCA) assay (Thermo Scientific, Rockford, IL).
Preparation of monoclonal antibodies
In order to produce mAbs, rPf1-Cys-Prx was used to immunize 8–10 week-old BALB/c mice. rPf1-Cys-Prx was mixed with an equal volume of complete Freund’s adjuvant (Sigma-Aldrich, St. Louis, MO) for initial immunization and with incomplete Freund’s adjuvant (Sigma-Aldrich, St. Louis, MO) for two subsequent immunizations. The antibody titer was measured by indirect enzyme-linked immunosorbent assay (ELISA). Prefusion boost was conducted three days before harvesting the spleen by injecting 0.2 ml of rPf1-Cys-Prx into the tail vein. Hybridomas were developed by fusion of harvested splenocytes to Sp2/0 myeloma cells and screened using ELISA and Western blot. Isotyping of mAbs was performed using IsoStrip mouse monoclonal antibody isotyping kit (Roche Diagnostics, Indianapolis, IN). The animal experiments in this study were carried out in compliance with the Obihiro University of Agriculture and Veterinary Medicine Guidelines for Animal Experimentation (permission numbers: 23–155 and 24–43).
Epitope mapping
The full length of the Pf1-Cys-Prx was split into four equal parts (Pf1-Cys-Prx (amino acid nos. 1–55), Pf1-Cys-Prx (56–110), Pf1-Cys-Prx (111–165) and Pf1-Cys-Prx (166–220)) with 55 amino acids each and primers were designed to amplify these DNA fragments (Supplementary Table S1). The amplified DNA fragments were digested with restriction enzymes and inserted into pGEX4T-1 (GE Healthcare Bio-Sciences AB, Uppsala, Sweden) by overnight incubation with ligation mixture (Takara, Otsu, Japan) at 15°C. The recombinant plasmids were transfected into E. coli (strain BL21). The transformed colony was incubated in LB broth (Sigma-Aldrich, St. Louis, MO) for expression of the glutathione S-transferase (GST)-tagged protein. The recombinant protein was purified with Glutathione Sepharose 3B following the manufacturer’s protocol (GE Healthcare Bio-Sciences AB). The purity of the recombinant protein was evaluated by 12% SDS/PAGE under reducing conditions and subsequent CBB staining (Fig. 1).
Fig. 1.
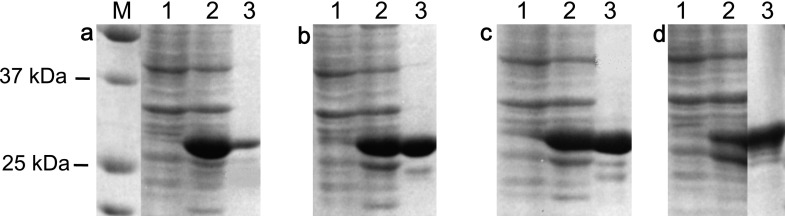
SDS/PAGE image of production of recombinant proteins for epitope mapping. The full length of Pf1-Cys-Prx was split into four parts (Pf1-Cys-Prx (amino acid nos. 1–55), Pf1-Cys-Prx (56–110), Pf1-Cys-Prx (111–165) and Pf1-Cys-Prx (166–220). Recombinant proteins were expressed in E. coli. M, Marker. Lane 1: E. coli culture before adding IPTG; lane 2: 4h after adding IPTG; and lane 3: purified recombinant proteins. Panel a: Pf1-Cys-Prx (1–55); panel b: Pf1-Cys-Prx (56–110); panel c: Pf1-Cys-Prx (111–165); and panel d: Pf1-Cys-Prx (166–220).
Results and Discussion
In this study, five different mAbs were produced against Pf1-Cys-Prx by screening the hybridoma clones with ELISA and Western blot. Isotyping showed that all of the mAbs are IgG1 with kappa light chains. In order to clarify the cross reactivity of established mAbs with the orthologous molecule of Pf1-Cys-Prx in P. vivax (Pv1-Cys-Prx) and P. knowlesi (Pk1-Cys-Prx), Western blot analysis was conducted using recombinant Pv1-Cys-Prx (rPv1-Cys-Prx) and rPk1-Cys-Prx protein (Fig. 2). As a result, mAbs 1, 2 and 3 reacted with rPv1-Cys-Prx and rPk1-Cys-Prx while mAbs 4 and 5 did not, indicating that mAbs 4 and 5 could differentiate P. falciparum from P. vivax and P. knowlesi.
Fig. 2.

Reactivity of produced mAbs against Pf1-Cys-Prx with Pv1-Cys-Prx and Pk1-Cys-Prx. Western blot analysis using mAbs (lanes 1–5) and specific polyclonal antibody against rPf1-Cys-Prx (lane P). Panel a: rPv1-Cys-Prx; and panel b: rPk1-Cys-Prx.
Epitope mapping to determine the target epitope of each mAbs showed that mAbs 1, 2 and 3 reacted with Pf1-Cys-Prx (166–220), which is the C-terminal part of Pf1-Cys-Prx, while mAbs 4 and 5 reacted with Pf1-Cys-Prx (111–165) (Fig. 3). Moreover, none of the mAbs reacted with GST. Deduced amino acid sequence analysis of Pf1-Cys-Prx (111–165) and Pf1-Cys-Prx (166–220) with the corresponding part in Pv1-Cys-Prx and Pk1-Cys-Prx showed 93% and 60% identity, respectively (Supplementary Fig. S1). Epitopes can be divided into linear and conformational residues wherein linear epitopes are a sequence of amino acids, while conformational epitopes result from a three-dimensional structure [14]. Although mAbs 4 and 5-targeting epitopes are located in Pf1-Cys-Prx (111–165), which had a high identity with the corresponding part in Pv1-Cys-Prx and Pk1-Cys-Prx, their binding epitope residues may exist in the non-identical part, or possibly they may be targeting conformational epitopes.
Fig. 3.

Epitope mapping. Western blot analysis shows the reactivity of each mAbs (lanes 1–5) to recombinant proteins. Panel a: Pf1-Cys-Prx (1–55); panel b: Pf1-Cys-Prx (56–110); panel c: Pf1-Cys-Prx (111–165); panel d: Pf1-Cys-Prx (166–220); and panel e: GST.
The binding affinities of mAbs were determined by measuring the dissociation constant (Kd) as described before [15]. Briefly, constant amounts of mAbs were incubated with various concentrations of rPf1-Cys-Prx until the equilibrium was reached. The remaining unsaturated mAbs were measured by indirect ELISA. Kd was determined using Klotz plot [15]. As a result, mAbs 1, 2 and 3 showed the highest affinity with Kd, namely 0.39 ± 0.16, 2.23 ± 0.83 and 1.64 ± 0.66 nM, respectively (Table 1). These findings are comparable with mAbs from previous studies showing a Kd of 12.03 ± 3.1 and 24.21 ± 2.07 nM against Plasmodium lactate dehydrogenase (pLDH) [16], and 0.16 ± 0.13 and 1.0 ± 0.049 nM against histidine-rich protein 2 (HRP-2) [17]. However, mAbs 4 and 5 showed a lower affinity (Table 1). These results suggest that mAbs 1, 2 and 3 are good candidates for use in the diagnosis of falciparum malaria, while mAbs 4 and 5 are useful for species differentiation in smear-confirmed malarial patients.
Table 1.
Dissociation constants of monoclonal antibodies determined by ELISA
| mAb* | Isotype | Kd (nM)# | SD¶ |
|---|---|---|---|
| 1 | IgG1 | 0.39 | 0.16 |
| 2 | IgG1 | 2.23 | 0.83 |
| 3 | IgG1 | 1.64 | 0.66 |
| 4 | IgG1 | 8743 | 2263 |
| 5 | IgG1 | 4873 | 851 |
* Monoclonal antibodies tested.
# The dissociation constants (Kd) are calculated by Klotz plot [15].
¶ Standard deviation.
Diagnosis of mixed-species malarial patients is a crucial factor in decisions regarding treatment [16]. P. knowlesi co-exists with other human Plasmodium species in Southeast Asian countries [4]. Diagnosis of P. knowlesi through microscopy alone is difficult because of its morphological similarities with other Plasmodium species. The early trophozoites of P. knowlesi are similar to those of P. falciparum while the late trophozoites and schizonts morphologically resemble those of P. malaria [6]. Some researchers have investigated the use of commercial RDTs as a way to detect knowlesi infections. McCutchan et al. [18] showed that mAbs targeting pLDH of P. falciparum and P. vivax can also detect P. knowlesi and attributed this to the highly similar amino acid sequences among these malaria parasites. In another study using two different commercial RDTs, P. knowlesi did not react with HRP-2-based RDT but was detected with pLDH-based RDT [19]. These results suggest that pLDH-based RDTs are not suitable for knowlesi malaria detection or differentiation. In the present study, however, mAbs 4 and 5 could differentiate falciparum from knowlesi malaria, indicating that they are good candidates for uses as differential diagnosis tools in P. knowlesi endemic Southeast Asian countries.
Since the diagnosis of malaria is based on direct detection of the parasites by microscopy or the parasites’ antigens, mAbs may play an important role in the detection of malaria parasites. Commercial antigen detection ELISAs and RDTs use mAbs that target one of three antigens (HRP-2, pLDH and aldolase) [3] being used extensively for case management and prevalence surveys. However, the available RDTs are known to have certain deficiencies, such as the fact that HRP-2 is only expressed in P. falciparum and persists even after parasite clearance, thus causing false positive results [20]. The genetic variation of HRP-2 in different geographical regions has also been reported, a problem that affects the sensitivity of tests in the field [21, 22], and a large proportion of field isolates in the Peruvian Amazon lack this antigen [23]. On the other hand, pLDH-based tests were less efficient in detecting low parasite densities and yielded variable results in different studies [24], while aldolase targeting tests have been shown to lack sensitivity in recent comparative studies, possibly due to the low concentration of this antigen in the parasite [24]. The development of RDTs targeting other antigens may overcome some of the deficiencies of current RDTs and improve the identification of human malaria parasites [3]. Pf1-Cys-Prx is highly expressed in the asexual stage of P. falciparum and accounts for 0.5% of total cellular protein during the trophozoite stage [11]. Moreover, Pf1-Cys-Prx does not exhibit genetic diversity and therefore maybe a great potential candidate for diagnostic use.
In summary, we produced five different mAbs targeting Pf1-Cys-Prx, two of which were able to differentiate P. falciparum from P. vivax and P. knowlesi. These mAbs can therefore be used in differential diagnosis as well as comparative molecular biology studies.
Acknowledgements
The authors are grateful to Dr. Yasuyuki Goto of the University of Tokyo for his technical help. This work was supported by a Grant-in-Aid for Scientific Research (23390098) from the Japan Society for the Promotion of Sciences.
Supplementary Material
References
- 1.World Health Organization World malaria report 2011. Geneva: World Health Organization, Available: http://www.who.int/malaria/world_malaria_report_2011/9789241564403_eng.pdf
- 2.Feachem RG, Phillips AA, Hwang J, Cotter C, Wielgosz B, Greenwood BM, Sabot O, Rodriguez MH, Abeyasinghe RR, Ghebreyesus TA, Snow RW. Shrinking the malaria map: progress and prospects. Lancet 2010, 376: 1566–1578 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.The malERA Consultative Group on Diagnoses and Diagnostics A research agenda for malaria eradication: diagnoses and diagnostics. PLoS Med 2011; 8(1): e1000396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Cox-Singh J, Davis TM, Lee KS, Shamsul SS, Matusop A, Ratnam S, Rahman HA, Conway DJ, Singh B. Plasmodium knowlesi malaria in humans is widely distributed and potentially life threatening. Clin Infect Dis 2008; 46: 165–171 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.White NJ. Plasmodium knowlesi: the fifth human malaria parasite. Clin Infec Dis 2008; 46: 172–173 [DOI] [PubMed] [Google Scholar]
- 6.Lee KS, Cox-Singh J, Singh B. Morphological features and differential counts of Plasmodium knowlesi parasites in naturally acquired human infections. Malar J 2009; 8: 73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rhee SG, Chae HZ, Kim K. Peroxiredoxins: a historical overview and peculative preview of novel mechanisms and emerging concepts in cell signaling. Free Radic Biol Med 2005; 38: 1543–1552 [DOI] [PubMed] [Google Scholar]
- 8.Gretes MC, Poole LB, Karplus PA. Peroxiredoxins in parasites. Antioxid Redox Signal 2012; 17(4): 608–633 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Angeles JM, Goto Y, Kirinoki M, Leonardo L, Tongol-Rivera P, Villacorte E, Inoue N, Chigusa Y, Kawazu SI. Human antibody response to thioredoxin peroxidase-1 and tandem repeat proteins as immunodiagnostic antigen candidates for Schistosoma japonicum infection. Am J Trop Med Hyg 2011; 85: 674–675 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kawazu S, Komaki-Yasuda K, Oku H, Kano S. Peroxiredoxins in malaria parasites: parasitologic aspects. Parasitol Int 2008; 57(1): 1–7 [DOI] [PubMed] [Google Scholar]
- 11.Kawazu S, Tsuji N, Hatabu T, Kawai S, Matsumoto Y, Kano S. Molecular cloning and characterization of a peroxiredoxin from the human malaria parasite Plasmodium falciparum. Mol Biochem Parasitol 2000; 109: 165–169 [DOI] [PubMed] [Google Scholar]
- 12.Hakimi H, Asada M, Angeles JMM, Kawai S, Inoue N, Kawazu S. Plasmodium vivax and P. knowlesi: cloning, expression and functional analysis of 1-Cys peroxiredoxin. Exp Parasitol 2013; 133: 101–105 [DOI] [PubMed] [Google Scholar]
- 13.Hakimi H, Asada M, Angeles JMM, Inoue N, Kawazu S. Cloning and characterization of Plasmodium vivax thioredoxin peroxidase-1. Parasitol Res 2012; 111: 525–529 [DOI] [PubMed] [Google Scholar]
- 14.Sun J, Xu T, Wang S, Li G, Wu D, Cao Z. Does difference exist between epitope and non-epitope residues? Analysis of the physicochemical and structural properties on conformational epitopes from B-cell protein antigens. Immunome Res 2011; 7: 1–1122126823 [Google Scholar]
- 15.Friguet B, Chaffotte AF, Djavadi-Ohaniance L, Goldberg ME. Measurements of the true affinity constant in solution of antigen-antibody complexes by enzyme-linked immunosorbent assay. J Immunol Methods 1985; 77: 305–319 [DOI] [PubMed] [Google Scholar]
- 16.Lee GC, Jeon ES, Le DT, Kim TS, Yoo JH, Kim HY, Chong CK. Development and evaluation of a rapid diagnostic test for Plasmodium falciparum, P. vivax, and mixed-species malaria antigens. Am J Trop Med Hyg 2011; 85(6): 989–993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kattenberg JH, Versteeg I, Migchelsen SJ, González IJ, Perkins MD, Mens PF, Schallig HD. New developments in malaria diagnostics: Monoclonal antibodies against Plasmodium dihydrofolate reductase-thymidylate synthase, heme detoxification protein and glutamate rich protein. mAbs 2012; 4(1): 120–126 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.McCutchan TF, Piper RC, Makler MT. Use of malaria rapid diagnostic test to identify Plasmodium knowlesi infection. Emerg Infect Dis 2008; 14: 1750–1752 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kawai S, Hirai M, Haruki K, Tanabe K, Chigusa Y. Cross-reactivity in rapid diagnostic tests between human malaria and zoonotic simian malaria parasite Plasmodium knowlesi infections. Parasitol Int 2009; 58: 300–302 [DOI] [PubMed] [Google Scholar]
- 20.Moody A. Rapid diagnostic tests for malaria parasites. Clin Microbiol Rev 2002; 15: 66–78 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Baker J, McCarthy J, Gatton M, Kyle DE, Belizario V, Luchavez J, Bell D, Cheng Q. Genetic diversity of Plasmodium falciparum histidine-rich protein 2 (PfHRP2) and its effect on the performance of PfHRP2-based rapid diagnostic tests. J Infect Dis 2005; 192: 870–877 [DOI] [PubMed] [Google Scholar]
- 22.Kumar N, Singh JP, Pande V, Mishra N, Srivastava B, Kapoor R, Valecha N, Anvikar AR. Genetic variation in histidine rich proteins among Indian Plasmodium falciparum population: possible cause of variable sensitivity malaria of rapid diagnostic tests. Malar J 2012; 11: 298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gamboa D, Ho MF, Bendezu J, Torres K, Chiodini PL, Barnwell JW, Incardona S, Perkins M, Bell D, McCarthy J, Cheng Q. A large proportion of P. falciparum isolates in the Amazon region of Peru lack pfhrp2 and pfhrp3: implications for malaria rapid diagnostic tests. PLoS One 2010; 5: e8091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.WHO-FIND-CDC-TDR Malaria rapid diagnostic test performance: results of WHO product testing of malaria RDTs: Round 1(2008). Geneva: World Health Organization; 2009
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.