

Abstract
Background & objectives:
Banaba (Lagerstroemia speciosa L.) extracts have been used as traditional medicines and are effective in controlling diabetes and obesity. The aim of this study was to evaluate the anti-HIV property of the extracts prepared from the leaves and stems of banaba, and further purification and characterization of the active components.
Methods:
Aqueous and 50 per cent ethanolic extracts were prepared from leaves and stems of banaba and were evaluated for cytotoxicity and anti-HIV activity using in vitro reporter gene based assays. Further, three compounds were isolated from the 50 per cent ethanolic extract of banaba leaves using silica gel column chromatography and characterization done by HPLC, NMR and MS analysis. To delineate the mode of action of the active compounds, reverse transcriptase assay and protease assay were performed using commercially available kits.
Results:
All the extracts showed a dose dependent inhibition of HIV-1-infection in TZM-bl and CEM-GFP cell lines with a maximum from the 50 per cent ethanolic extract from leaves (IC50= 1 to 25 μg/ml). This observation was confirmed by the virus load (p24) estimation in infected CEM-GFP cells when treated with the extracts. Gallic acid showed an inhibition in reverse transcriptase whereas ellagic acid inhibited the HIV-1 protease activity.
Interpretation & conclusions:
The present study shows a novel anti-HIV activity of banaba. The active components responsible for anti-HIV activity were gallic acid and ellagic acid, through inhibition of reverse transcriptase and HIV protease, respectively and hence could be regarded as promising candidates for the development of topical anti-HIV-1 agents.
Keywords: Anti-HIV-1 activity, ellagic acid, gallic acid, Lagerstroemia speciosa L., protease, reverse transcriptase, rutin
Human immunodeficiency virus (HIV) infection is initiated by the binding of the envelope protein gp120 to a set of cell-surface molecules comprising the primary receptor CD4 and a co-receptor, usually either CCR5 or CXCR41,2,3. In the host cell, viral RNA is reverse transcribed by reverse transcriptase (RT). This viral DNA is integrated in the host genetic material by the integrase enzyme. HIV-1 protease also plays an essential role by promoting virus maturation and thus promoting infection of new cells. The activity of this enzyme is essential for virus infectivity, rendering the protein a major therapeutic target for AIDS treatment. The probable inhibitors may target many stages in the virus life cycle: virus adsorption, virus-cell fusion, virus uncoating, HIV regulatory proteins and HIV enzymes (reverse transcriptase, integrase and protease). The problem of drug resistance in HIV infection is a consequence of the virus propensity to mutate4. Mutations arise because HIV's replication machinery lacks the proofreading mechanism and copies itself with low fidelity resulting in one mistake every time it replicates. The selective effect of these drugs, also favours emergence of mutations that can establish clinical drug resistance5. Additionally, the dose-limiting side-effects and the necessity for long-term anti-HIV treatments are the limitations to standard HIV therapy6. Therefore, it is essential to focus on isolation of novel anti-HIV therapeutics from natural resources which are well tolerated, convenient and relatively cheaper. HIV reverse transcriptase inhibitors are important drugs for the treatment of AIDS and many natural products from plants belonging to a wide range of different structural classes, e.g., coumarins, flavonoids, tannins, alkaloids, lignans, terpenes, naphtho- and anthraquinones, and polysaccharides have been shown to be active as RT inhibitors7,8. Different plant extracts and compounds isolated from Leguminoseae, Loranthaceae and Malvaceae family inhibited protease activity9.
Lagerstroemia speciosa L. (Lythraceae), commonly known as ‘Banaba’ grows widely in tropical countries, including Australia, India, China, Malaysia and Philipines. The tea prepared from the leaves of this plant has traditionally been used in several countries for treating diabetes and obesity10,11,12. Administration of the aqueous and methanolic extracts prepared from the leaves of this plant was shown to be effective in reducing blood glucose and insulin levels in hereditary type 2 diabetic KK-Ay/Ta Jcl mice13. These extracts were shown to stimulate glucose uptake in 3T3-L1 cells in a manner similar as insulin14. In obese diabetic mice, its leaf extract led to a significant reduction in their body weights15. Corosolic acid (2-α-hydroxyursoloic acid; C30H48O4) and ellagitannins have been identified as active principles responsible for stimulating glucose transport16,17. Standardized extract prepared from the leaves of this plant containing 1 per cent corosolic acid, Glucose™, when administered orally in the form of gelatin capsule has shown its efficacy to lower blood glucose levels in a randomized clinical trial involving type 2 diabetic patients18. In addition to its traditional use for controlling diabetes and obesity, recent studies suggest that fruit extracts modulates biofilm formation by Pseudomonas aeruginosa19. Further, orobol 7-O-D-glucoside isolated from this plant inhibits human rhinovirus replication20.
In the present study, the aqueous and 50 per cent ethanolic extracts from the leaves and stems as well as different fractions prepared from 50 per cent ethanolic extract of banaba leaves have been evaluated for anti-HIV activity using different in vitro reporter gene based cell assay systems. Attempts have been made to isolate bioactive compounds and delineate their mechanism of action.
Material & Methods
Collection of plant materials: L. speciosa L. material was collected three times from Lucknow, India, during June 2007, May 2008 and November 2008. Air and shed dried plant materials (leaves and stems) were grinded and strained through 30 mesh (0.5 mm). The voucher specimen (accession number 264305) has been submitted to Herbarium of National Botanical Research Institute, Lucknow, India.
Preparation of extracts: The finely grinded leaves and stems (1 kg) were extracted with either MilliQ water (5 l × 3) at 60-75°C for 6-8 h or by ethanol: water (1:1 v/v, 5 l × 3) at 25-30°C for overnight to obtain aqueous as well as 50 per cent ethanolic extracts of leaves and stems. These extracts were filtered and concentrated under reduced pressure to afford aqueous and ethanolic (50%) crude extracts (A-D).
Preparation of fractions: The dried 50 per cent alcoholic extract (100 g) prepared from the leaves was suspended in 1 l petroleum ether (60-80°C), followed by stirring at 10-20 rpm on rotavapor at 30-40°C for 30-60 min and was subsequently left for overnight at room temperature (25-30°C). The clear petroleum ether soluble fraction was collected after filtration through Whatman filter paper number 1. The marc (residual petroleum ether insoluble extract) was treated three more times with the petroleum ether as above, to collect the total petroleum ether soluble fraction. It was concentrated at 30-40°C and finally dried under vacuum to collect 7-9 per cent of dried petroleum ether soluble fraction. The residual petroleum ether insoluble extract, left after extraction by petroleum ether, was air dried for 20-30 h. It was then suspended in 1 l of chloroform followed by stirring at 10-20 rpm on rotavapor at 30-40°C for 30-60 min. It was left overnight at room temperature (25-30°C). The clear chloroform soluble fraction was collected after filtration through Whatmann filter paper number 1. The residual chloroform insoluble extract was treated thrice with the chloroform as above, to collect the total chloroform soluble fraction. It was concentrated at 35-45°C and finally vacuum dried at -20°C to collect 17-20 per cent of dried chloroform soluble fraction. The residual chloroform insoluble extract was air dried for 20-30 h, treated in a similar way with n-butanol to prepare the n-butanol soluble fraction.
Isolation of compounds: The 50 per cent ethanolic extract (A, 10 g) prepared from leaves was chromatographed on silica gel (100–200 mesh, Merck, Darmstadt, Germany), using petroleum ether, chloroform and methanol as eluants. Methanol soluble fractions on repeat chromatography using chloroform and methanol (95:5 to 80:20 v/v) afforded compound E (rutin, yellowish green powder; 2.00 to 6.00 mg/g), compound F (gallic acid, pale white powder; 0.58 to 1.50 mg/g), and compound G (ellagic acid, buff coloured powder; 1.07 to 4.09 mg/g). These compounds were identified by direct comparison with the spectroscopic data (NMR and MS) of authentic sample, procured from MP Biomedicals, Ohio, Solon, USA as well as those reported in the literature21.
Cells and virus: TZM-bl cells [a recombinant HeLa cell line expressing high levels of CD4, HIV-1 co-receptors CCR5 & CXCR4 with β-galactosidase and luciferase as reporter genes under HIV-1 long terminal repeat (LTR) promoter] were maintained in Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich Inc., USA) supplemented with 10 per cent foetal bovine serum (FBS; Biological Industries, Kibbutz beit Haemek, Israel), and an antibiotic-antimycotic cocktail [penicillin (100 units/ml), streptomycin (100 μg/ml) and amphotericin B (250 ng/ml); Pen-Strep-Ampho sol, Biological Industries]. CEM-GFP [a CD4+ T-lymphocytic reporter cell line expressing green fluorescent protein (GFP) under HIV-1 LTR promoter] suspension cells were cultured in RPMI-1640 medium (Sigma-Aldrich Inc.) supplemented with 10 per cent FBS, G418 (500 μg/ml; Gibco, Grand Island, NY, USA) and antibiotic-antimycotic cocktail as described for TZM-bl cells22. HIV-1 NL4.3 was prepared by transfection of HEK-293T cells with pNL4.3 plasmid (Catalog number 114; AIDS Research and Reference Reagent Program, Division of AIDS, National Institute of Allergy and Infectious Diseases, USA) using CaPO4 method23.
Cytotoxicity assay: Cytotoxicity of extracts/compounds was assessed using MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; Sigma-Aldrich Inc., USA] assay24. Briefly, TZM-bl cells (6 × 103/well/100 μl were seeded in a 96-well culture plate (Greiner Bio-One, GmbH, Frickenhausen, Germany) and grown overnight at 37°C in a humidified atmosphere of 5 per cent CO2. Next day, culture medium with increasing concentrations of various extracts/compounds was added in duplicate and further incubated for 48 h. Appropriate solvents, used to prepare various extracts/compounds were included as negative controls. After incubation, 20 μl of MTT reagent (5 mg/ml) was added per well and incubated at 37°C for 3 h followed by addition of MTT solvent (100 μl/well; 20% SDS and 50% dimethyl formamide in 50 mM PBS). The absorbance (OD) was read at 570 nm with reference filter at 690 nm. Similarly, CEM-GFP cells (5 × 104/0.2 ml culture medium/well) were also assessed for their viability as described for TZM-bl cells. Cell viability was calculated using the equation,
% Viability = [(OD extract/compound treated cultures)/(OD untreated cell control cultures)] × 100
Inhibition of HIV infection in TZM-bl cells-based assay: TZM-bl cells (4 × 104/well) were seeded in 24-well plate and cultured overnight. In separate vials, HIV-1 NL4.3 at a multiplicity of infection (MOI) of 0.05 were treated with extracts/compounds or solvents used to prepare above extracts/compounds for 1 h at 37°C. Subsequently, pretreated viruses were added in duplicate to TZM-bl cells and cultured for 4 h. The cells were washed once with cold 50 mM PBS after incubation to remove the cell free viruses followed by addition of fresh culture medium with extracts/compounds. Azidothymidine (AZT; Sigma-Aldrich Inc) was used as positive control whereas negative control comprised cells with untreated HIV. Cells were further cultured for 48 h, washed twice with PBS, and lysed with 1X lysis buffer (Promega Corporation, Sweden, USA) by freeze-thaw. The supernatant was analyzed for luciferase activity by adding substrate (Promega Corporation) using white optiplate in the fluorimeter (BMG Labtech GmbH, Offenberg, Germany). The results were expressed as percentage inhibition, calculated by taking the luminescence in experimental group (i.e. in presence of test extract/compound/AZT) divided by the luminescence in infected cells in absence of test extract/compound/AZT multiplied by hundred. Per cent inhibition was calculated by subtracting the above value from hundred.
Inhibition of HIV infection in CEM-GFP cells-based assay: CEM-GFP cells were infected with HIV-1 NL4.3 virus at an MOI of 0.05 as described previously25. Post-infection, the cells (2.0 × 105/well) were incubated with varying concentrations of plant extracts/compounds for up to 8 days. AZT was used as a positive control whereas solvents used to prepare extracts were used as negative controls. Post-incubation, cells were lysed with 150 μl of 1X Promega cell culture lysis buffer, 100 μl/well was transferred to black optiplate and absorbance read at an excitation wavelength of 485 nm and emission at 520 nm using fluorimeter. The results were expressed as percentage inhibition, calculated by taking the GFP fluorescence in experimental group divided by GFP fluorescence in infected cells in the absence of test extract/compound/AZT multiplied by hundred. Per cent inhibition was obtained by subtracting the above value from hundred.
p24 ELISA: The virus load in the supernatant of CEM-GFP cells treated with plant extracts was measured by estimating p24 using an ELISA kit (SAIC-Frederick Inc., NCI-Frederick, USA), following the instruction manual. Per cent inhibition in virus load was calculated by dividing the p24 concentration in the presence of plant extracts/AZT by p24 value observed in control (infected cells without any treatment), multiplied by hundred and the obtained value was subtracted from hundred.
Reverse transcriptase assay, protease assay and integrase assay: The inhibitory activity on HIV-1 RT, protease and integrase was determined as per the manufacturer's instructions of the respective kits (Reverse Transcriptase assay kit from Roche Applied Sciences, Mannheim, Germany; Protease assay kit from Anaspec, CA, USA and Integrase Assay kit from XpressBio, Life Science Products, MD, USA).
Statistical analysis: Analyses of concentration-response data were performed by non linear curve-fitting programme Prism (GraphPad Software, Inc., CA, USA) to determine CC50 and IC50 values. The results pertaining to inhibition of RT, protease and integrase activity are presented as mean ± SEM of three independent experiments.
Results
The 50 per cent ethanolic and aqueous extracts prepared from leaves and stems (A-D) from all three collections were evaluated for anti-HIV activity using TZM-bl as well as CEM-GFP cells and were found to possess promising anti-HIV activity. The typical example of anti-HIV activity of the extracts prepared from L. speciosa L. collected in November 2008 is described below.
The cytotoxicity of the above extracts by MTT assay was assessed on the reporter cell lines used for determination of anti-HIV activity. The CC50 values of 50 per cent ethanolic as well as aqueous extracts prepared from stems and leaves of L. speciosa L. were > 438 μg/ml on TZM-bl cells (Table I) and > 693 μg/ml on CEM-GFP cells (Table II).
Table I.
In vitro cytotoxicity and anti-HIV activity of the extracts/isolated compounds of L. speciosa L. using TZM-bl cells
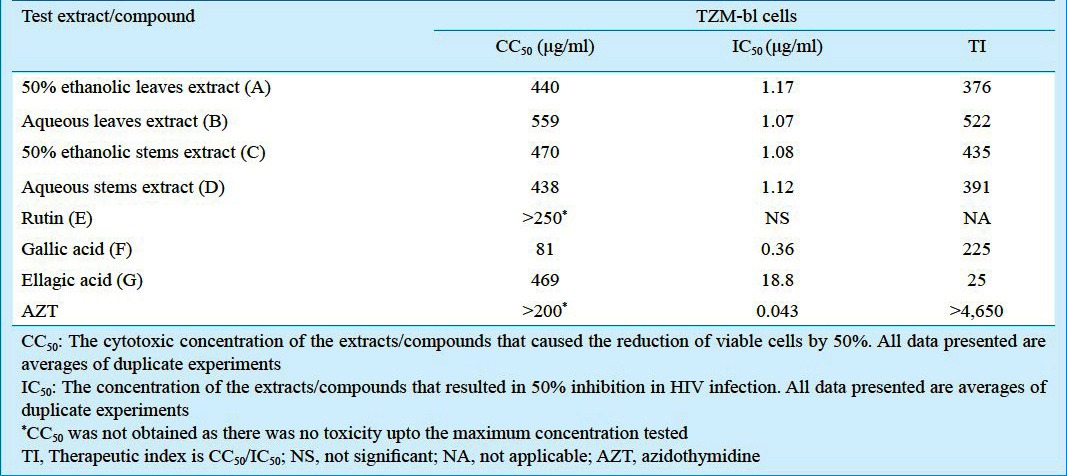
Table II.
In vitro cytotoxicity and anti-HIV activity of the extracts/isolated compounds of L. speciosa L. using CEM-GFP cells
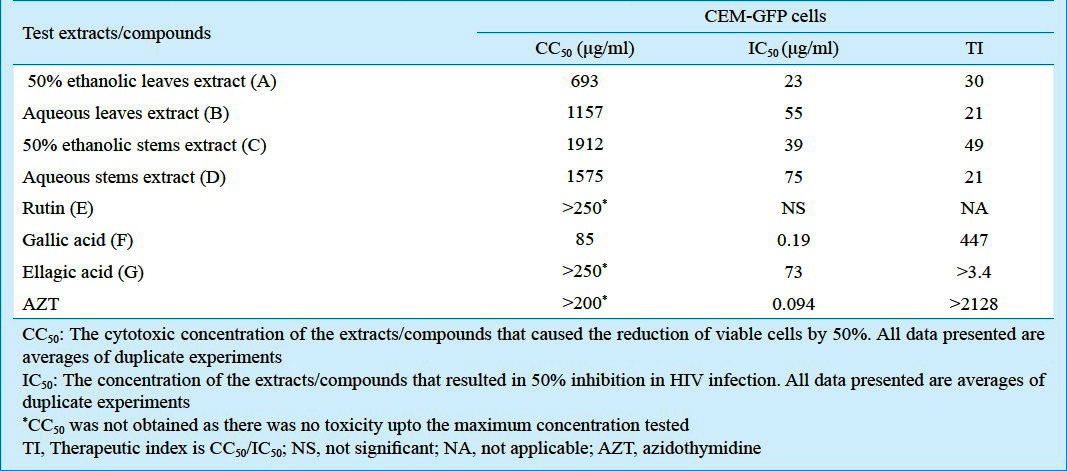
The aqueous as well as 50 per cent ethanolic extracts from both the leaves and stems of the plant inhibited HIV infection in TZM-bl cells in a dose dependent fashion (Fig. 1A). The IC50 obtained by using non linear curve-fitting programme ranged from 1.07 to 1.17 μg/ml with respect to the extracts (A-D) (Table I). These extracts also showed potent anti-HIV activity in CEM-GFP cells previously infected with HIV-1 and treated with varying concentrations of these extracts (Fig. 1B). The IC50 values of extracts (A-D) varied from 23-75 μg/ml (Table II). The highest anti-HIV activity was observed in extract (A) i.e. 50 per cent ethanolic extract of leaves (IC50 value; 23 μg/ml) (Table II). The leaf extracts as compared to the stem extracts were more potent in decreasing the virus load in the supernatant of infected cells (Fig. 1C).
Fig. 1.

Different fractions prepared from the 50 per cent ethanolic extract of leaves showed different phytochemical constituents by HPLC (data not shown). The anti-HIV activity evaluation of these fractions was performed at concentrations much below than that showing cytotoxicity. There was no anti-HIV activity observed in the petroleum ether fraction whereas both chloroform and n-butanol soluble fractions showed a dose dependent inhibition in HIV infection (Fig. 2).
Fig. 2.

Anti-HIV activity of different fractions prepared from L. speciosa L. (A) shows anti-HIV activity at varying concentrations of chloroform soluble fraction and n-butanol soluble fraction in TZM-bl cells. (B) shows a dose dependent inhibition in green fluorescent protein (GFP) expression using NL4.3 HIV infected CEM-GFP cells whereas (C) shows the virus load (p24) in the supernatant. Values shown as mean ± SEM of two independent experiments performed in duplicate.
The 50 per cent ethanol extract of leaves was processed to isolate the active compounds (E, F and G). NMR data along with other spectroscopic data suggest the compound E to be rutin, which did not show any significant anti-HIV activity (Tables I and II).
NMR data along with HPLC and MS data suggested compound F to be gallic acid. It has shown a potent anti-HIV activity in both the assays. The cytotoxicity of this compound on both the cell lines was >80 μg/ml. The anti-HIV activity of this compound was at concentrations much lower than the cytotoxic concentration. The IC50 values in TZM-bl and CEM-GFP cells based assays were 0.36 and 0.19 μg/ml, respectively (Tables I and II).
Using MS, HPLC and NMR data, Compound G, was characterized as ellagic acid. It also showed a consistent anti-HIV activity in both the assays. The anti-HIV activity observed was not due to cellular toxicity as much higher concentration was required to obtain the CC50 value of the compound G (Tables I and II).
The n-butanol fraction and compounds were further evaluated for their anti-RT and anti-protease activity. The n-butanol fraction and gallic acid (compound F) when tested at different concentrations showed a dose dependent inhibition in HIV-1 RT activity as compared with the reference control nevirapine (1 μM) (Fig. 3A & 3B).
Fig. 3.
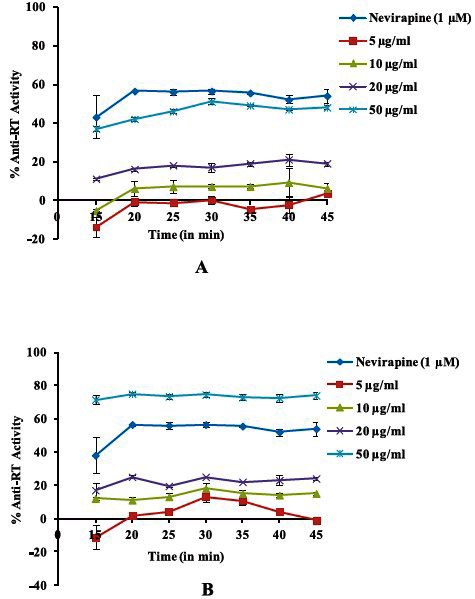
Inhibitory activity of n-butanol fraction of Lagerstroemia speciosaL. and Compound F (Gallic acid) on HIV reverse transcriptase. (A) shows a dose dependent inhibition in HIV RT activity by the n-butanol fraction as compared with the reference control nevirapine as a function of time. Similarly (B) shows the inhibitory activity of compound F (Gallic acid). Values shown as mean ± SEM of three independent experiments.
The n-butanol fraction and the ellagic acid (compound G) showed a dose-dependent inhibition of HIV-1 protease activity, when compared with the vehicle control. Saquinavir (1 μM), a known inhibitor of HIV-protease activity was used as a reference control (Fig. 4A & 4B). These extracts and compounds were also screened for their inhibition of HIV-1 integrase activity using commercially available ELISA kit (data not shown) and were found to be ineffective when tested upto a concentration of 50 μg/ml as compared with the positive control (sodium azide).
Fig. 4.

Inhibitory activity of n-butanol fraction of Lagerstroemia speciosa L. and Compound G (Ellagic acid) on HIV-1 protease activity. (A) shows a dose dependent inhibition in HIV-1 protease activity by the n-butanol fraction as compared with Saqinavir used as a reference control with time. (B) shows the inhibitory activity of compound G (Ellagic acid) on HIV-1 protease. Values shown as mean ± SEM of three independent experiments.
Discussion
Banaba has been used as a folk medicine to treat diabetes in various parts of the world. The anti-hyperglycaemic effect of its aqueous and methanolic extracts has been demonstrated in several animal models as well in human studies without any adverse effects10,11,12. The extract also possesses anti-hyperlipidaemic, anti-oxidant, anti-inflammatory, anti-fungal, anti-viral and anti-neoplastic properties19,20,26. The aqueous as well as 50 per cent ethanolic extracts from both the leaves and stems of the plant inhibited HIV infection in dose dependent manner in TZM-bl cells and CEM-GFP reporter gene based assays. The supernatant from the infected and extract treated cells showed the maximum inhibition in virus load by 50 per cent ethanolic leaves extract. The therapeutic index (TI) of a drug is the ratio between the toxic and the therapeutic dose and is used as a measure of its relative safety. The TI of the extracts (A-D) were >20 and ranged up to 400. A high TI of these extracts also suggests the plant as an effective candidate for isolation of natural compounds possessing anti-HIV activity. Since, the 50 per cent ethanol extract from leaves was found to be most effective, it was further fractionated into three different fractions by macerating with petroleum ether, chloroform and n-butanol successively. All fractions were evaluated for anti-HIV activity. Because maximum inhibition was localized in chloroform and n-butanol fractions 50 per cent ethanolic crude extract prepared from leaves was chromatographed over a silica gel column, and elution with chloroform-methanol yielded three different compounds. These compounds were characterized as rutin, gallic acid and ellagic acid. Further evaluation of these compounds for their cytotoxicity and anti-HIV activity showed that gallic acid and ellagic acid possessed anti-HIV activity. Gallic acid and ellagic acid from other plant sources have been reported for their anti-HIV activity27,28,29. Observed anti-HIV activity of the extracts/purified compounds in CEM-GFP cells based assay suggested that these may be acting as post-entry inhibitors by inhibiting the activities of reverse transcriptase (RT)/HIV specific protease/integrase. Due to the essential role of HIV RT in synthesizing the double-stranded proviral DNA from single-stranded HIV RNA genome, it is a major target among the current anti-HIV therapies. Drugs such as AZT, lamivudine, stavudine, zalcitabine, and didanosine found useful in slowing HIV infections are RT inhibitors. In our study n-butanol fraction prepared from L speciosa L. and gallic acid (Compound F) showed inhibition of HIV-1 RT activity.
Protease inhibitors bind to the active site of the viral protease enzyme, preventing the processing of viral particles into functional form. Protease inhibitors lead to the production of non-infectious (non-functional) viral particles and serve as a potent inhibitor of HIV propagation. Hence, testing these extracts as well as the compounds for their anti-protease activity was imperative. The n-butanol fraction and the ellagic acid (Compound G) showed inhibition of HIV-1 protease activity. The n-butanol fraction as well as gallic acid and ellagic acid did not show any significant inhibition of HIV integrase activity. However, in one of the previous studies, ellagic acid has been shown to have HIV integrase inhibitory activity26. At this stage, the possible reasons for discrepancy with respect to HIV integrase inhibitory activity of ellagic acid are not clear. One of the possibilities may be, that we have used a commercial kit, which may have different sensitivity and specificity as compared to the assay system used in earlier study27.
In this study, the active fractions and compounds showed minimal haemolytic effect on RBCs upto a maximum concentration of 50 μg/ml (data not shown). The extracts obtained from this plant, is a mixture of several antiviral phytocompounds which may provide beneficial remedies in the form of combination therapy for the management of HIV infection and AIDS30. Gallic acid acts by inhibiting RT activity whereas ellagic acid acts by inhibiting HIV protease activity. Microbicides based on polyherbal creams have been proposed to prevent sexually transmitted infections, including HIV31,32. The present results suggest that L. speciosa L. has a potential to be considered for the development of a plant-based microbicide for prevention of HIV infection. Further in vivo studies need to be done to confirm these findings.
Acknowledgment
This study was supported by the research grants from the Department of Biotechnology, Government of India and Indian Council of Medical Research, Government of India. The authors thank Dr Debashis Mitra, National Centre for Cell Science, Pune, for making available TZM-bl and CEM-GFP cell lines and NIH AIDS Research & Reference Reagent program, Division of AIDS, NIAID, NIH for providing the molecular clone of HIV (NL4.3).
References
- 1.Klatzmann D, Champagne E, Chamaret S, Gruest J, Guetard D, Hercend T, et al. T-lymphocyte T4 molecule behaves as the receptor for human retrovirus LAV. Nature. 1984;312:767–8. doi: 10.1038/312767a0. [DOI] [PubMed] [Google Scholar]
- 2.Alkhatib G, Combadiere C, Broder CC, Feng Y, Kennedy PE, Murphy PM, et al. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Science. 1996;272:1955–8. doi: 10.1126/science.272.5270.1955. [DOI] [PubMed] [Google Scholar]
- 3.Feng Y, Broder CC, Kennedy PE, Berger EA. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science. 1996;272:872–7. doi: 10.1126/science.272.5263.872. [DOI] [PubMed] [Google Scholar]
- 4.Flexner C. HIV drug development: the next 25 years. Nat Rev Drug Disco. 2007;6:959–66. doi: 10.1038/nrd2336. [DOI] [PubMed] [Google Scholar]
- 5.O’Brien WA. Resistance against reverse transcriptase inhibitors. Clin Infect Dis. 2000;30(Suppl 2):S185–92. doi: 10.1086/313858. [DOI] [PubMed] [Google Scholar]
- 6.Hupfeld J, Efferth T. Drug resistance of human immunodeficiency virus and overcoming it by natural products. In Vivo. 2009;23:1–6. [PubMed] [Google Scholar]
- 7.el-Mekkawy S, Meselhy MR, Kusumoto IT, Kadota S, Hattori M, Namba T. Inhibitory effects of Egyptian folk medicines on human immunodeficiency virus (HIV) reverse transcriptase. Chem Pharm Bull (Tokyo) 1995;43:641–8. doi: 10.1248/cpb.43.641. [DOI] [PubMed] [Google Scholar]
- 8.Matthee G, Wright AD, Konig GM. HIV reverse transcriptase inhibitors of natural origin. Planta Med. 1999;65:493–506. doi: 10.1055/s-1999-14004. [DOI] [PubMed] [Google Scholar]
- 9.Filho JR, de Sousa Falcao H, Batista LM, Filho JM, Piuvezam MR. Effects of plant extracts on HIV-1 protease. Curr HIV Res. 2010;8:531–44. doi: 10.2174/157016210793499204. [DOI] [PubMed] [Google Scholar]
- 10.Garcia F. On the hypoglycemic effect of decoction of Lagerstroemia speciosa (banaba) administered orally. J Phil Med Assoc. 1940;20:395–402. [Google Scholar]
- 11.Quisumbing E. Medicinal plants of the Philippines. Quezon, Philippines: Katha Publishing; 1978. pp. 640–2. [Google Scholar]
- 12.Matsuyama F. Composition for inhibiting increase in blood sugar level or lowering blood sugar level. United States Patent Application No. 09/730. 2008:74. [Google Scholar]
- 13.Kakuda T, Sakane I, Takihara T, Ozaki Y, Takeuchi H, Kuroyanagi M. Hypoglycemic effect of extracts from Lagerstroemia speciosa L. leaves in genetically diabetic KK-AY mice. Biosci Biotechnol Biochem. 1996;60:204–8. doi: 10.1271/bbb.60.204. [DOI] [PubMed] [Google Scholar]
- 14.Liu F, Kim J, Li Y, Liu X, Li J, Chen X. An extract of Lagerstroemia speciosa L. has insulin-like glucose uptake-stimulatory and adipocyte differentiation-inhibitory activities in 3T3-L1 cells. J Nutr. 2001;131:2242–7. doi: 10.1093/jn/131.9.2242. [DOI] [PubMed] [Google Scholar]
- 15.Suzuki Y, Unno T, Ushitani M, Hayashi K, Kakuda T. Antiobesity activity of extracts from Lagerstroemia speciosa L. leaves on female KK-Ay mice. J Nutr Sci Vitaminol (Tokyo) 1999;45:791–5. doi: 10.3177/jnsv.45.791. [DOI] [PubMed] [Google Scholar]
- 16.Murakami C, Myoga K, Kasai R, Ohtani K, Kurokawa T, Ishibashi S, et al. Screening of plant constituents for effect on glucose transport activity in Ehrlich ascites tumour cells. Chem Pharm Bull (Tokyo) 1993;41:2129–31. doi: 10.1248/cpb.41.2129. [DOI] [PubMed] [Google Scholar]
- 17.Hayashi T, Maruyama H, Kasai R, Hattori K, Takasuga S, Hazeki O, et al. Ellagitannins from Lagerstroemia speciosa as activators of glucose transport in fat cells. Planta Med. 2002;68:173–5. doi: 10.1055/s-2002-20251. [DOI] [PubMed] [Google Scholar]
- 18.Judy WV, Hari SP, Stogsdilla WW, Judy JS, Naguib YMA, Passwater R. Antidiabetic activity of a standardized extract (Glucosol) from Lagerstroemia speciosa leaves in type II diabetics. A dose-dependence study. J Ethnopharmacol. 2003;87:115–7. doi: 10.1016/s0378-8741(03)00122-3. [DOI] [PubMed] [Google Scholar]
- 19.Singh B, Singh HB, Singh A, Singh B, Mishra A, Nautiyal C. Lagerstroemia speciosa fruit extract modulates quorum sensing-controlled virulence factor production and biofilm formation in Pseudomonas aeruginosa. Microbiology. 2012;158:529–38. doi: 10.1099/mic.0.052985-0. [DOI] [PubMed] [Google Scholar]
- 20.Choi HJ, Bae EY, Song JH, Baek SH, Kwon DH. Inhibitory effects of orobol 7-O -D-glucoside from banaba (Lagerstroemia speciosa L.) on human rhinoviruses replication. Lett Appl Microbiol. 2010;51:1–5. doi: 10.1111/j.1472-765X.2010.02845.x. [DOI] [PubMed] [Google Scholar]
- 21.Mahajan A, Pai N. Simultaneous isolation and identification of phytoconstituents from Terminalia chebula by preparative chromatograph. J Chem Pharm Res. 2010;2:97–103. [Google Scholar]
- 22.Gervaix A, West D, Leoni LM, Richman DD, Wong-Staal F, Corbeil J. A new reporter cell line to monitor HIV infection and drug susceptibility in vitro. Proc Natl Acad Sci USA. 1997;94:4653–8. doi: 10.1073/pnas.94.9.4653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pear WS, Nolan GP, Scott ML, Baltimore D. Production of high-titer helper-free retroviruses by transient transfection. Proc Natl Acad Sci USA. 1993;90:8392–6. doi: 10.1073/pnas.90.18.8392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mosmann T. Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods. 1983;65:55–63. doi: 10.1016/0022-1759(83)90303-4. [DOI] [PubMed] [Google Scholar]
- 25.Kumar M, Mitra D. Heat shock protein 40 is necessary for human immunodeficiency virus-1 Nef-mediated enhancement of viral gene expression and replication. J Biol Chem. 2005;280:40041–50. doi: 10.1074/jbc.M508904200. [DOI] [PubMed] [Google Scholar]
- 26.Stohs SJ, Miller H, Kaats GR. A review of the efficacy and safety of banaba (Lagerstroemia speciosa L.) and corosolic acid. Phytother Res. 2012;26:317–24. doi: 10.1002/ptr.3664. [DOI] [PubMed] [Google Scholar]
- 27.Farnet CM, Wang B, Hansen M, Lipford JR, Zalkow L, Robinson WE, Jr, et al. Human immunodeficiency virus type 1 cDNA integration: new aromatic hydroxylated inhibitors and studies of the inhibition mechanism. Antimicrob Agents Chemother. 1998;42:2245–53. doi: 10.1128/aac.42.9.2245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Tuchinda P, Kornsakulkarn J, Pohmakotr M, Kongsaeree P, Prabpai S, Yoosook C, et al. Dichapetalin-type triterpenoids and lignans from the aerial parts of Phyllanthus acutissima. J Nat Prod. 2008;71:655–63. doi: 10.1021/np7007347. [DOI] [PubMed] [Google Scholar]
- 29.Bedoya LM, Abad MJ, Sanchez-Palomino S, Alcami J, Bermejo P. Ellagitannins from Tuberaria lignosa as entry inhibitors of HIV. Phytomedicine. 2010;17:69–74. doi: 10.1016/j.phymed.2009.08.008. [DOI] [PubMed] [Google Scholar]
- 30.Houghton PJ. Compounds with anti-HIV activity from plants. Trans R Soc Trop Med Hyg. 1996;90:601–4. doi: 10.1016/s0035-9203(96)90403-4. [DOI] [PubMed] [Google Scholar]
- 31.Talwar GP, Dar SA, Rai MK, Reddy KV, Mitra D, Kulkarni SV, et al. A novel polyherbal microbicide with inhibitory effect on bacterial, fungal and viral genital pathogens. Int J Antimicrob Agents. 2008;32:180–5. doi: 10.1016/j.ijantimicag.2008.03.004. [DOI] [PubMed] [Google Scholar]
- 32.Usha PR, Naidu MU, Raju YS. Evaluation of the antiretroviral activity of a new polyherbal drug (Immu-25) in patients with HIV infection. Drugs R D. 2003;4:103–9. doi: 10.2165/00126839-200304020-00003. [DOI] [PubMed] [Google Scholar]