

Abstract
Even though technologies involving nano/microparticles have great potential, it is crucial to determine possible toxicity of these technological products before extensive use. Fullerenes C60 are nanomaterials with unique physicochemical and biological properties that are important for the development of many technological applications. The aim of this study was to evaluate the consequences of nonphotoexcited fullerene C60 exposure in brain acetylcholinesterase expression and activity, antioxidant responses, and oxidative damage using adult zebrafish as an animal model. None of the doses tested (7.5, 15, and 30 mg/kg) altered AChE activity, antioxidant responses, and oxidative damage when zebrafish were exposed to nonphotoexcited C60 nano/microparticles during 6 and 12 hours. However, adult zebrafish exposed to the 30 mg/kg dose for 24 hours have shown enhanced AChE activity and augmented lipid peroxidation (TBARS assays) in brain. In addition, the up-regulation of brain AChE activity was neither related to the transcriptional control (RT-qPCR analysis) nor to the direct action of nonphotoexcited C60 nano/microparticles on the protein (in vitro results) but probably involved a posttranscriptional or posttranslational modulation of this enzymatic activity. Taken together these findings provided further evidence of toxic effects on brain after C60 exposure.
1. Introduction
Nanoscience investigates nanoscale phenomena and serves as the foundation for nanotechnology, which develops practical applications for nanomaterials (particles typically with sizes in the 1–100 nm range but not exclusively) [1]. Due to their composition, small size, and shape, nanomaterials exhibit novel properties for diverse applications that have already yielded in a variety of commercially available products [2]. As a consequence, it is expected that both humans and environmental systems will be increasingly exposed to nanomaterials in the next years. Nanotoxicology [3] and nanoecotoxicology [4] are emerging disciplines that arose to address the potential human and environmental health effects of nanomaterials' exposure.
Fullerenes are molecular materials that call attention after the first preparation of C60 which is an allotrope of carbon consisting of 60 carbon atoms joined to form a cage-like structure [5]. C60 exhibits unique physical and chemical properties for many technological applications, such as electronics, superconductors, cosmetics [6], and, more recently, drug and gene delivery [7]. Although the toxic effects of C60 are still mostly unknown, especially those related to neurotoxicity, some general aspects have already been addressed. For instance, C60 is reported to be cytotoxic to various mammalian cells [8] and it has been shown to induce lipid peroxidation in human cells [9], in brain of juvenile largemouth bass (Micropterus salmoides) [10], and in common carp (Cyprinus carpio) [11]. Moreover, C60 was classified as “very toxic” to aquatic organisms by the grid for the potential ecotoxicological hazard evaluation which is based mainly on toxicity to fish, Daphnia, and algae and in data about degradability of the substance [4].
In cholinergic neurotransmission, choline acetyltransferase (ChAT) is responsible for the synthesis of acetylcholine (ACh) in the presynaptic neuron. After its release into the synaptic cleft, ACh promotes the activation of metabotropic muscarinic and ionotropic nicotinic cholinergic receptors. The reaction responsible for the maintenance of levels of ACh is catalyzed by two cholinesterases (ChE): acetylcholinesterase (AChE) (E.C. 3.1.1.7) and butyrylcholinesterase (BuChE) (E.C. 3.1.1.8) [12]. The zebrafish (Danio rerio) has long been considered a powerful animal model for studying several biological events. More recently, zebrafish become also a valuable model to environmental and toxicological studies [13]. It has been demonstrated that BuChE is not encoded in the zebrafish genome, but AChE is encoded by a single gene that has been cloned, sequenced, and functionally detected in zebrafish brain [14].
AChE activity has been widely used as a bioindicator of environmental exposure to neurotoxic pollutants. For example, the inhibition of AChE as a biomarker for assessment of the exposure of organisms to organophosphate and carbamate insecticides is well known [15]. The inhibition of brain AChE activity of aquatic species by toxic substances such as methanol [16], heavy metals mercury, and lead [17], neurotoxins [18], and organochlorine pesticide endosulfan [19] also has been well established. On the other hand, AChE activation has also been demonstrated as a consequence of exposure to neurotoxic compounds such as aluminum [20] and ethanol [21].
Therefore, considering that (i) both humans and environmental systems will be increasingly exposed to C60 in a near future, (ii) the neurotoxic effects of C60 are far from being completely understood, (iii) measurement of AChE activity in organisms is used as a biomarker of environmental contamination due to neurotoxic substances, (iv) determination of oxidative stress parameters is recognized as a tool to evaluate toxicity mediated by small particles exposure, and (v) zebrafish is a well-established organism for toxicological analysis, the aim of the present study was to evaluate the nonphotoexcited C60 effects in brain AChE activity and its gene expression pattern. Furthermore, we have analyzed the effects of the nonphotoexcited C60 in parameters related to antioxidant defenses and lipid peroxidation in the brain of zebrafish.
2. Materials and Methods
2.1. Chemicals
Fullerene (C60, 99.5% purity) was purchased from Aldrich (Milwaukee, WI, USA); DMSO was purchased from Fisher Scientific (Pittsburgh, PA, USA) and Trizma Base; ethylenediaminetetraacetic acid (EDTA), ethylene glycol bis (beta aminoethylether)-N,N,N′,N′-tetraacetic acid (EGTA), sodium citrate, Coomassie Blue G, bovine serum albumin, acetylthiocholine, 5,5′-dithiobis-2-nitrobenzoic acid (DTNB) HEPES, BHT (99%), 2,2′-azobis(2-methylpropionamidine) dihydrochloride (ABAP), and 1,1,3,3-tetramethoxypropane were purchased from Sigma Chemical Co. (St. Louis, MO, USA). KCl and SDS (90%) were purchased from Labsynth (Brazil). Tetramethoxypropane (TMP) and 2′,7′-dichlorodihydrofluorescein diacetate were purchased from Acros Organics (Morris Plains, NJ, USA) and Molecular Probes Inc. (Eugene, OR, USA) respectively. MgCl2 and Acetic acid 99.7% were purchased from Isofar and Vetec (Brazil), respectively. TRIzol reagent, Platinum Taq DNA Polymerase, and SYBR Green I were purchased from Invitrogen (Carlsbad, CA, USA). ImProm-II Reverse Transcription System was purchased from Promega (Madison, WI, USA). All other reagents used were of analytical grade.
2.2. Animals
Adult wild-type zebrafish (Danio rerio, Cyprinidae) of both sexes (6–9 months old) were obtained from a specialized supplier (Redfish Agroloja, RS, Brazil). Animals were kept at a density of up to five animals per liter in 50 L housing tanks with tap water that was previously treated with Tetra's AquaSafe (to neutralize chlorine, chloramines, and heavy metals present in the water that could be harmful to fish) and continuously aerated (7.20 mg O2/L) at 26 ± 2°C, under a 14/10 h light/dark controlled photoperiod. Animals were acclimated for at least two weeks before the experiments and were fed three times a day with TetraMin Tropical Flake fish food. The fish were maintained healthy and free of any signs of disease and were used according to the Guide for the Care and Use of Laboratory Animalspublished by the US National Institutes of Health. All procedures in the present study were approved by the Animal Ethics Committee of the Pontifical Catholic University of Rio Grande do Sul (PUCRS), protocol number 10/00185-CEUA.
2.3. C60 Suspension
Suspensions of C60 in DMSO were prepared as previously described [23, 24] with modifications. Briefly, 7.6 mg of C60 was added to 0.5 mL of DMSO and sonicated for 3 h. The concentrated suspension of C60 obtained was diluted in water to result in suspensions that when intraperitoneally injected (mean injection volume was 10 μL) get the doses of 7.5, 15, and 30 mg/kg fish (12.5% DMSO). For this reason, the C60 suspensions were named as 7.5, 15, and 30. The doses and times of exposure were selected based on previous studies where multiwalled carbon nanotubes were intravenously injected in mice [25, 26], and functionalized single-walled carbon nanotubes were intraperitoneally injected in rats [27]. C60 suspensions were further sonicated for one hour prior to use. To avoid C60's photoexcitation, the C60 suspensions were prepared and stored in a dark condition.
2.4. Characterization of C60 Suspensions
The C60 suspensions (7.5, 15, and 30) were characterized in terms of particle size distribution [28]. The mean diameter over the volume and number distribution (d 4.3) was determined by laser diffractometry (Mastersizer 2000, Malvern Instruments, UK). The value of SPAN was utilized to determine particle size distribution according to (1), where d 0.9, d 0.1, and d 0.5 are the particle diameters determined at 90%, 10%, and 50% cumulative undersized volumes, respectively,
| (1) |
2.5. Animal Procedures
Intraperitoneal (i. p.) injection was adopted as administration route in the in vivo protocols (i) to avoid the photoexcitation of the C60 material and (ii) to ensure that exposure concentrations are in line with target values. Intraperitoneal injections were conducted using a 3/10-mL U-100 BD ultra-fine short insulin syringe 8 mm (5/16′′) × 31G short veedle (Becton Dickinson and Company, NJ, USA) according to the protocol established by Phelps and colleagues [29]. Briefly, the volume injected into the animal (mean injection volume was 10 μL) was adjusted to the fish bodyweight (mean mass of the animals was 0.75 g ± 0.06 g/mean ± SEM) to achieve 7.5, 15, and 30 mg/kg based on a previous study [30]. The animals of the control group received the same volume of saline solution, and the animals of the vehicle control received the same volume of 12.5% DMSO. Anesthesia of the animals prior to the injection was obtained by its immersion in a solution of benzocaine (1 mM in MeOH 1%) until the animal showed a lack of motor coordination and reduced respiratory rate. The anesthetized animal was gently placed in a water-soaked gauze-wrapped hemostat with the abdomen facing up and the head of the fish positioned at the hinge of the hemostat (the pectoral fins were used as a landmark on the abdomen). The needle was inserted parallel to the spine in the midline of the abdomen posterior to the pectoral fins. The injection procedure was conducted in such a way as to guarantee that the animal did not spend more than 10 s out of the water. After the injection, the animals were placed in a separate tank with highly aerated nonchlorinated tap water (25 ± 2°C) to facilitate recovery from the anesthesia. Saline solution was used as control. All the animals that recovered within 2-3 min following the injection continued in the experiment, while animals that did not recover during this period were discarded. Six, twelve, or twenty-four hours after the injection, the animals were euthanized.
2.6. In Vitro Assays of AChE Activity
In vitro assays were performed as previously described [31, 32] in order to evaluate if 7.5, 15, and 30 C60 suspensions might have a direct effect on the enzyme. Briefly, 33 μL of C60 suspensions were added to the reaction medium before the preincubation with the enzyme-containing lysate from zebrafish brain homogenate and maintained during the enzyme assays. Control treatments with equal volume of vehicle (DMSO 12.5%) were performed to exclude the DMSO effect on the enzyme activities.
2.7. Determination of AChE Activity
Zebrafish were euthanized, and their whole brains were removed by dissection. The brains (two whole brains for each sample) were homogenized on ice in 60 volumes (v/w) of Tris-citrate buffer (50 mM Tris, 2 mM EDTA, 2 mM EGTA, pH 7.4, adjusted with citric acid), in a glass-Teflon homogenizer. The rate of acetylthiocholine hydrolysis (ACSCh, 0.88 mM) was assessed in a final volume of 300 μL with 11 mM phosphate buffer, pH 7.5, and 0.22 mM DTNB using a method previously described [33]. Before the addition of substrate, samples containing protein (5 μg) and the reaction medium described above were preincubated for 10 min at 25°C. The hydrolysis of substrate was monitored by the formation of thiolate dianion of DTNB at 412 nm for 2-3 min (intervals of 30 s) in a microplate reader. Controls without the homogenate preparation were performed in order to determine the nonenzymatic hydrolysis of the substrate. The linearity of absorbance against time and protein concentration was previously determined. The AChE activity was expressed as micromoles of thiocholine (SCh) released per hour per milligram of protein. All enzyme assays were evaluated in triplicate, and at least four independent experiments were performed.
2.8. Gene Expression Analysis by Quantitative Real-Time RT-PCR (RT-qPCR)
Gene expression analysis was carried out only when kinetic alteration occurred. For this reason, immediately after 24 hours of intraperitoneal injection (C60 suspension 30), the animals were euthanized by decapitation. For each sample, a pool of three zebrafish whole brains was used. Total RNA was isolated with Trizol reagent (Invitrogen, Carlsbad, CA, USA) in accordance with the manufacturer's instructions. The total RNA was quantified by spectrophotometry and the cDNA was synthesized with ImProm-II Reverse Transcription System (Promega) from 1 μg total RNA, following the manufacturer's instructions. Quantitative PCR was performed using SYBR Green I (Invitrogen) to detect double-strand cDNA synthesis. Reactions were done in a volume of 25 μL using 12.5 μL of diluted cDNA (1 : 100 for EF1α and Rlp13α; and 1 : 20 for ache), containing a final concentration of 0.2x SYBR Green I (Invitrogen), 100 μM dNTP, 1x PCR Buffer, 3 mM MgCl2, 0.25 U Platinum Taq DNA Polymerase (Invitrogen), and 200 nM of each reverse and forward primers (Table 1). The PCR cycling conditions were an initial polymerase activation step for 5 min at 95°C, 40 cycles of 15 s at 95°C for denaturation, 35 s at 60°C for annealing, and 15 s at 72°C for elongation. At the end of cycling protocol, a melting curve analysis was included and fluorescence measured from 60 to 99°C. Relative expression levels were determined with 7500 Fast Real-Time System Sequence Detection Software v.2.0.5 (Applied Biosystems). The efficiency per sample was calculated using LinRegPCR 11.0 Software (http://LinRegPCR.nl) and the stability of the references genes, EF1α and Rlp13α (M-value), and the optimal number of reference genes according to the pairwise variation (V) were analyzed by GeNorm 3.5 Software (http://medgen.ugent.be/genorm/). Relative RNA expression levels were determined using the 2−ΔΔCT method.
Table 1.
PCR primers design.
| Proteins | Primer sequences (5′-3′) | Accession number (mRNA) |
|---|---|---|
| EF1α* |
F: CTGGAGGCCAGCTCAAACAT R: ATCAAGAAGAGTAGTACCGCTAGCATTAC |
NSDART00000023156 |
| Rpl13α* | F: TCTGGAGGACTGTAAGAGGTATGC R: AGACGCACAATCTTGAGAGCAG |
NM_212784 |
| ac he** | F: GCTAATGAGCAAAAGCATGTGGGCTTG R: TATCTGTGATGTTAAGCAGACGAGGCAGG |
NM_131846 |
*According to Tang et al. [22].
**Designed by the authors.
2.9. Antioxidant Capacity against Peroxyl Radicals
Total antioxidant capacity against peroxyl radicals was performed according Amado and colleagues [34], employing the thermal decomposition of 2,2′-azobis 2 methylpropionamidine dihydrochloride (ABAP; 4 mM) as peroxyl radical generator. Reactive oxygen species (ROS) concentration was detected with the fluorescent probe 2′,7′-dichlorofluorescein diacetate (H2DCF-DA) in a final concentration of 40 μM (480 and 525 nm forexcitation and emission, resp.). The relative difference between ROS area with and without ABAP was considered a measure of antioxidant capacity, with high area difference meaning low antioxidant capacity, since high fluorescence levels were obtaining after adding ABAP, meaning low competence to neutralize peroxyl radicals.
2.10. Measurement of Lipid Peroxidation
Lipid peroxidation was measured through the determination of thiobarbituric acid reactive substances (TBARS), following the methodology of Oakes and van der Kraak [35]. Brain homogenates (10 μL) were added to a reaction mixture made with 150 μL of 20% acetic acid, 150 μL of thiobarbituric acid (2.4%), 50 μL of Milli-Q water and 20 μL of sodium dodecyl sulfate (SDS, 8.1%). Samples were heated at 95°C during 30 min, and after cooling by 10 min, 100 μL of Milli-Q water and 500 μL of n-butanol were added. After centrifugation (3,000 ×g during 10 min at 15°C), the organic phase (150 μL) was placed in a microplate reader, and the fluorescence registered after excitation at 520 nm and emission of 580 nm. The concentration of TBARS (nmol/mg of wet tissue) was calculated employing tetramethoxypropane (TMP) as standard.
2.11. Protein Determination
Protein was measured by the Coomassie blue method [36] using bovine serum albumin as standard.
2.12. Statistical Analysis
AChE activity and antioxidant analyses were expressed as means ± S.E.M. and analyzed by one-way analysis of variance (ANOVA). Post hoc comparisons were made using Tukey's test and orthogonal comparisons. Before ANOVA, its assumptions (normality and variances homogeneity) were checked. Molecular data were expressed as means ± S.E.M. and analyzed by Student's t-test. In every case, the significance level was fixed in 5% (α = 0.05).
3. Results
3.1. C60 Suspensions of Nano/Microparticles
Although DMSO is known to show low toxicity by itself [24, 37], appropriate experimental controls must be employed to eliminate its influence. In this study, the DMSO was diluted to result in 12.5% DMSO as a final concentration. Any signal of toxicity, that is, mortality or even transient alterations in behavior, was observed in the vehicle control group (12.5% DMSO). In addition, control group (saline) and vehicle control group were never statistically different in the conditions tested.
The nano/microparticles mean diameters over the volume showed wide distributions (60 nm–316 μm; 69 nm–1,905 μm; 182 nm–208 μm, for suspensions 7.5, 15, and 30, resp.), with most abundant sizes in the micrometric range (Figures 1(a), 1(c), and 1(e)). The SPAN values increased with the dilution of suspensions (5.843, 3.003, and 1.607 to suspensions 7.5, 15, and 30, resp.), showing a narrow size distribution with lower dilutions. On the other hand, it is important to emphasize that when considering the mean diameters over the number distribution, we observed that the greater part of particles were under nanometric sizes (Figures 1(b), 1(d), and 1(f)).
Figure 1.
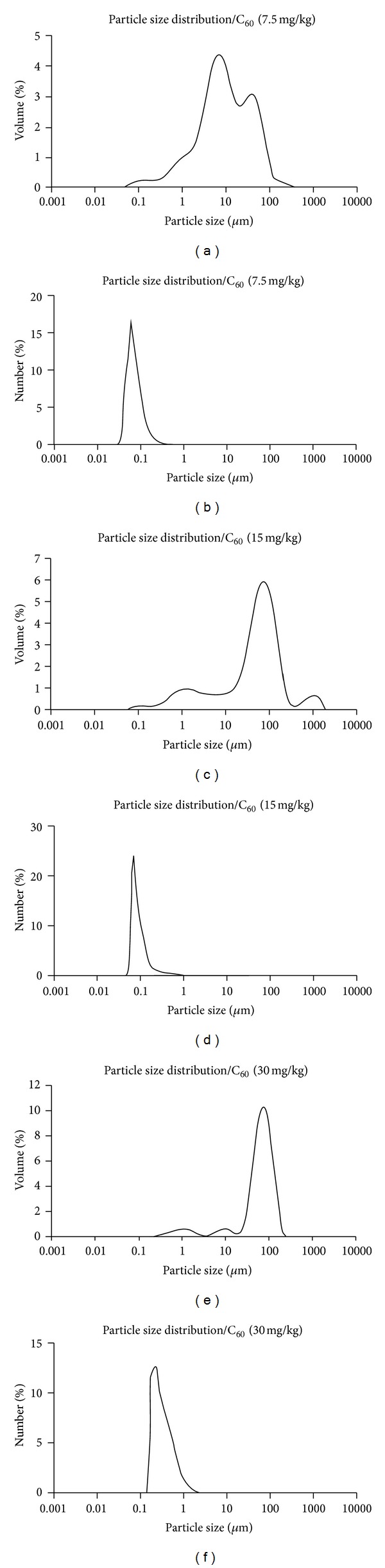
Size distribution in suspensions of nano/microparticles. Mean diameters distribution of fullerene C60 suspensions (7.5, 15, and 30) was determined over the volume ((a), (c), and (e)) and over the number ((b), (d), and (f)) of the nano/microparticles.
3.2. Acetylcholinesterase Enzymatic Activity and Gene Expression
The effect of different C60 concentrations and times of exposure on brain AChE activity was demonstrated by performing (in vivo) experiments using adult zebrafish. None of the concentrations tested (7.5, 15, and 30 mg/kg) altered AChE activity when zebrafish were exposed to C60 during 6 hours (Figure 2(a)) and 12 hours (Figure 2(b)). However, the analysis for 24 hours demonstrated that animals treated with the concentration of 30 mg/kg presented a significant increase in AChE activity (28.54 ± 3.72 μmol SCh·h−1·mg protein−1; P = 0.0001) when compared to saline (12.19 ± 0.55 μmol SCh·h−1·mg protein−1; P = 0.0001) and to the vehicle control group (15.46 ± 0.57 μmol SCh·h−1·mg protein−1; P = 0.0001) (Figure 2(c)). The upregulation of brain AChE activity after exposure to C60 (30 mg/kg for 24 hours) could be a consequence of transcriptional control. In order to determine if transcriptional regulation of AChE gene has occurred, a RT-qPCR analysis was performed. The results have shown that AChE transcript levels were not enhanced when compared to the vehicle control group (P = 0.6695; Figure 3) suggesting that the activation of brain AChE is not directly related with the transcriptional control.
Figure 2.
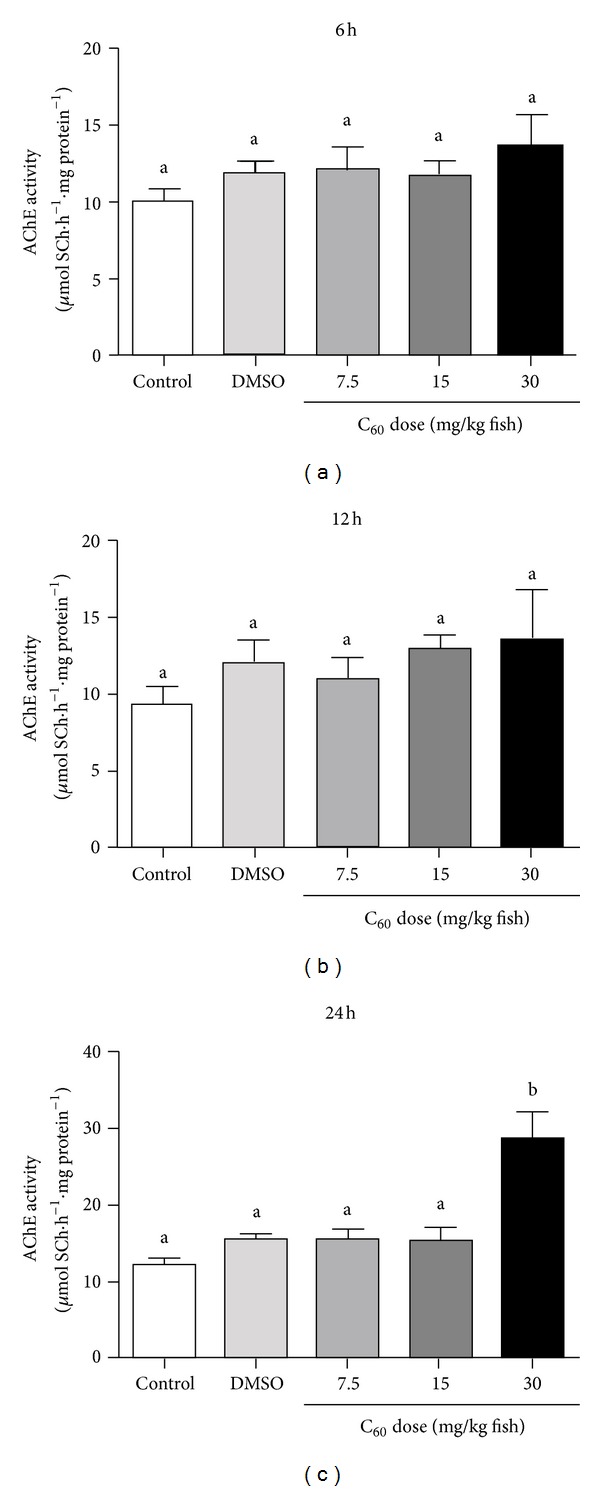
In vivo AChE activity in zebrafish brain after 06 (a), 12 (b), and 24 hours (c) of fullerene C60 exposure at distinct concentrations (7.5–30 mg/kg fish). Bars represent the mean ± SEM of at least three different experiments, each one performed in triplicate. The specific enzyme activity is reported as micromoles of thiocholine released per hour per milligram of protein. Bars represent the mean ± SEM of at least three independent experiments, each one performed in triplicate. Different letters indicate significant differences (P < 0.05) between groups.
Figure 3.
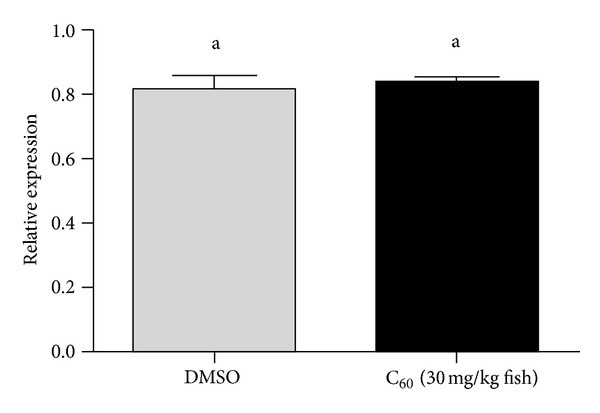
RT-qPCR analysis. Relative ache expression profile after fullerene C60 exposure (30 mg/kg fish for 24 hours) on zebrafish brain. Bars represent the mean ± SEM.
3.3. In Vitro Effects of C60 Suspensions on Acetylcholinesterase Activity
To verify whether C60 nano/microparticles might have a direct effect on the enzyme, we tested the in vitro effect of C60 suspensions on AChE activity in zebrafish brain. The results showed that C60 suspensions did not bring about any alteration in AChE activity (P = 0.7701; Figure 4).
Figure 4.
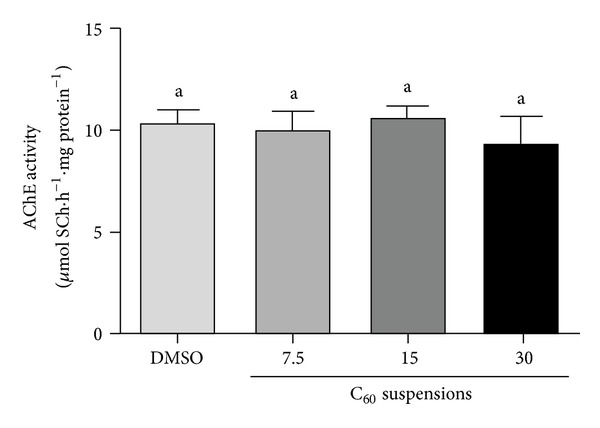
In vitro AChE activity. In vitro effect of different C60 suspensions (7.5, 15, and 30) on ACh hydrolysis in zebrafish brain. Bars represent the mean ± SEM of at least three different experiments, each one performed in triplicate.
3.4. Antioxidant Analysis
The total antioxidant competence against peroxyl radicals showed an augmented response (lower relative area) in brains of zebrafish exposed to C60 (15 mg/kg) for 6 hours when compared to zebrafish exposed to C60 (7.5 and 30 mg/kg) for 6 hours (P = 0.0209; Figure 5(a)). No other differences were observed under the experimental conditions (Figures 5(b) and 5(c)). Oxidative damage, measured by lipid peroxidation (TBARS assays), showed a pro-oxidant condition elicited by C60 at the highest dose (30 mg/kg) after 24 hours (P = 0.0194; Figures 6(a), 6(b), and 6(c)).
Figure 5.
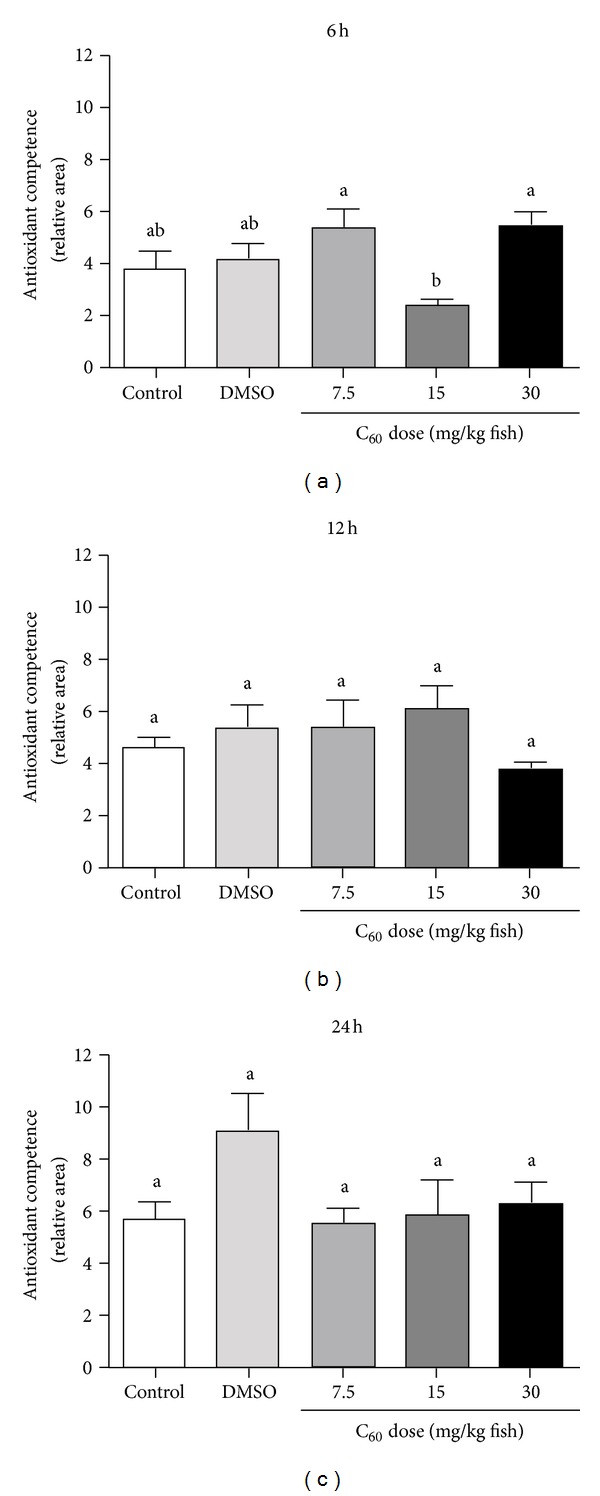
Antioxidant capacity. Total antioxidant capacity against peroxyl radical in zebrafish brain after 06 (a), 12 (b), and 24 hours (c) of fullerene C60 exposure at distinct concentrations (7.5–30 mg/kg fish). Bars represent the mean ± SEM of at least three independent experiments. Different letters indicate significant differences (P < 0.05) between groups.
Figure 6.
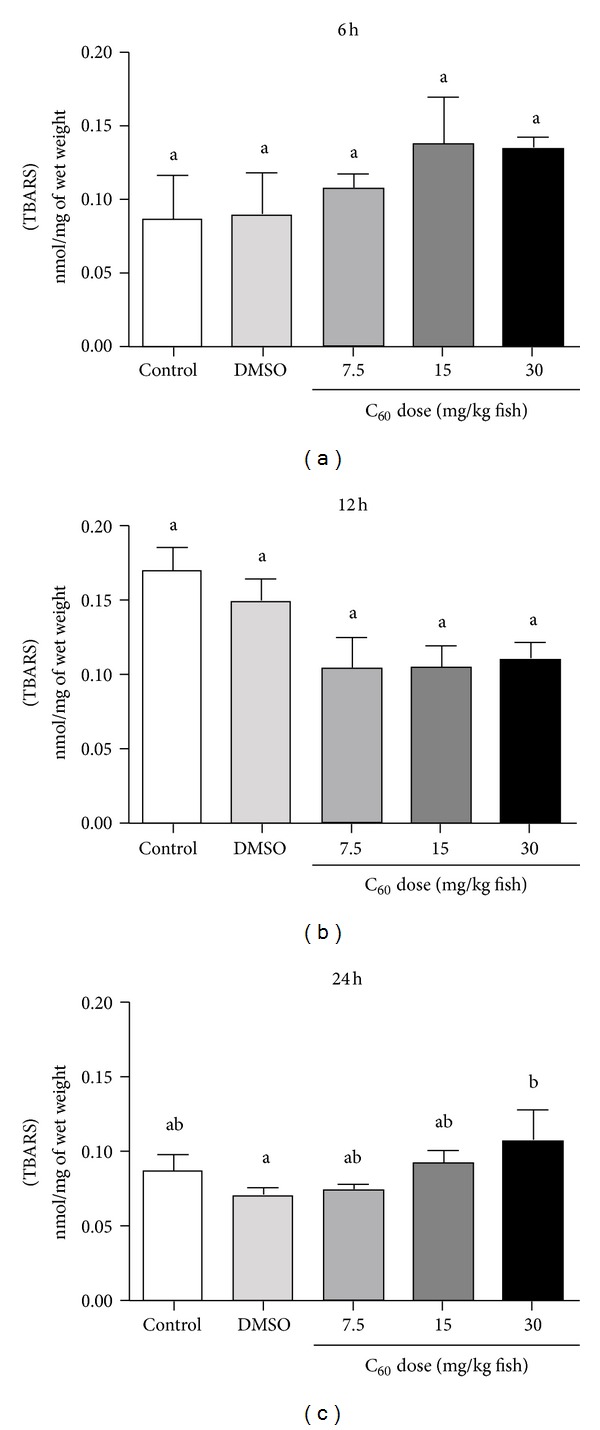
Lipid oxidative damage. Concentration of thiobarbituric acid reactive substances (TBARS; nmol/mg of wet weight) in zebrafish brain after 06 (a), 12 (b) and 24 hours (c) of fullerene C60 exposure at distinct concentrations (7.5–30 mg/kg fish). Bars represent the mean ± SEM of at least three independent experiments. Different letters indicate significant differences (P < 0.05) between groups.
4. Discussion
Although technologies evolving nano/microparticles have considerable potential in diverse applications, it is crucial to determine possible toxicity of these technological products before extensive use. Little is known about the toxic effects of fullerenes in brain. At present, only few studies presenting contradictory findings have evaluated possible neurotoxic effects of fullerenes exposure. For instance, it was already suggested that C60 did not cross the blood-brain barrier [38], whereas the results obtained by Mokrushin [39] suggested that fullerenes possess marked neurotropic properties and are neurotoxic substances irreversibly blocking the electrical activity of the nervous tissue.
Neurotoxicity of C60 in fish species has been previously reported [11, 40, 41]. Generation of reactive oxygen species (ROS) by C60 is influenced in part by the presence and type of illumination due to the photoexcitation of C60 by UV and visible light [42] or even to by-products of the organic solvents employed to prepare C60 suspensions [40]. For this reason, the C60 suspensions were prepared under the protection of light, and i.p. injection was adopted as administration route in the in vivo protocols, an experimental condition that avoids the influence of light in the analyzed variables. Also, in vitro assays were run in darkness.
The characterization of the size and stability of C60 nanoparticles in suspension is very important to evaluate their toxicity once particle size can change during the preparation of the suspension, dilution, and exposure [11]. In this study, the nano/microparticles mean diameters over the volume in the C60 suspensions showed wide distributions with most abundant sizes in the micrometric range. The C60 has a tendency to form aggregates very easily [7], and this may be a possible cause of this wide distribution. In contrast, the nano/microparticles mean diameters over the number of distribution in the C60 suspensions demonstrated that the greater part of particles was under nanometric sizes. Totsuka and colleagues [43] also observed wide distributions by dynamic light scattering in formulations manufactured with C60. Take into account that particle sizes limit their ability to translocate to different tissues [3, 44] the assessment of the distribution of C60 material on zebrafish brain would be useful to reinforce our findings. Nevertheless, studies have demonstrated that particles even larger than the nano/microparticles of this study were able to reach brain. For instance, the study of Sarlo and colleagues [45] showed that 1000 nm latex fluorospheres were recovered from rat brains one and 24 hours after intravenous (i.v.) injection. In addition, the study by Zhu and colleagues [41] showed that exposure to fullerene aggregates suspended in water (with average diameters of approximately 349 and/or 1,394 nm) decreased glutathione in brain of juvenile carps (Carassius auratus).
In the present study, we have evaluated the effect of different C60 doses (7.5, 15, and 30 mg/kg) and different times of exposure (6, 12, and 24 hours) on AChE activity and ache expression in zebrafish brain. In the concentrations tested, only the animals exposed to 30 mg/kg for 24 hours have shown enhanced AChE activity. The RT-qPCR results suggested that the activation of brain AChE is not directly related with the transcriptional control. The in vitro results indicated that none of the C60 suspensions had a direct effect on the enzyme. It is important to highlight that in vitro experiments do not evaluate the influence of other indirect mechanisms such as cell signaling pathways. Altogether, our results indicate that the effect of intraperitoneal exposure to nano/microparticles of fullerene (C60) on brain AChE activity was neither related to AChE gene activation nor to the direct action of this molecular material on this enzyme but probably involved a posttranscriptional or posttranslational modulation of this enzymatic activity.
Moreover, we have shown the effects of C60 exposure over the antioxidant competence and lipid peroxidation in zebrafish brain. The results demonstrated that the exposition to 30 mg/kg during 24 hours did not alter the antioxidant competence and yielded in higher levels of lipid peroxidation. The lack in the antioxidant response to a pro-oxidant situation could explain the augmented lipid peroxidation found in zebrafish brain. In addition, Totsuka and colleagues [43] reported increased micronuclei frequencies, induced DNA damage, and increased mutant frequencies after C60 nano/microparticles suspension exposure.
AChE is indispensable for terminating acetylcholine-mediated neurotransmission at cholinergic synapses [46]. In this context, AChE is inhibited by organophosphorus and carbamate insecticides and by neurotoxins, which are structural analogues of acetylcholine [47]. In addition, there are lines of evidence to suggest that AChE contributes to diverse physiological processes through its involvement in the regulation of cell proliferation, differentiation, and survival. As a consequence, more recently AChE has been redefined as an important regulator of apoptosis, because it can be induced by a variety of apoptotic stimuli [48, 49]. It is well known that apoptosis underlies the neurotoxic effects of various compounds. Moreover, zebrafish brain AChE activation has also been demonstrated as a consequence of exposure to known neurotoxic compounds, including aluminum [20] and ethanol [21] and to the cyanobacterial toxin microcystin-LR [50].
The antioxidant or pro-oxidant effects induced by C60 exposure are still a debatable issue [40, 51]. C60 is photoexcited under UV or visible light [42], a condition, for example, that elicited lipid peroxidation in brains of common carp (C. carpio) [11]. On the other hand, the absence of light did not completely inhibit oxidative stress generation in embryonic zebrafish after C60 exposure [52]. Moreover, cell membrane lipid peroxidation was suggested as the main mechanism of toxicity caused by fullerenes' exposure [10, 11]. In addition, the study by Sayes and colleagues [9] reported cytotoxicity (cell apoptosis) in three human cell cultures including astrocytes caused by cell membrane lipid peroxidation due to exposure to nano-C60 (a water-soluble fullerene species).
The results presented in this paper provide further experimental evidence that C60 exposure can be neurotoxic. Adult zebrafish exposed to nonphotoexcited C60 nano/microparticles (30 mg/kg for 24 hours) have shown enhanced AChE activity and augmented lipid peroxidation in brain. The study by Melo and colleagues [53] proposed a mechanism to explain enhanced AChE activity mediated by oxidative stress generation in cultured retinal cells exposed to amyloidogenic peptide Aβ 25–35. The incubation with Aβ 25–35 led to an increment of ROS formation and increased significantly lipid peroxidation levels which decreased cell membrane order and ultimately led to the exposure of more active sites of the AChE. As a consequence, AChE activity was increased. It is possible to speculate that similar events had occurred in zebrafish brain. However, further studies must be performed in order to evaluate this hypothesis.
Conflict of Interests
The authors do not have any conflict of interests.
Acknowledgments
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (“Nanotoxicologia ocupacional e ambiental: subsídios científicos para estabelecer marcos regulatórios e avaliação de riscos”-Proc. 552131/2011-3) and DECIT/SCTIEMS through CNPq and Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS) (Proc. FAPERGS 10/0036-5-PRONEX). G. O. Dal Forno, M. B. de Azevedo and R. S. Britto were recipients of fellowships from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). L. W. Kist and R. S. Fritsch were recipient of fellowships from CNPq. S. S. Guterres, C. D. Bonan, J. M. Monserrat, and M. R. Bogo are productivity research fellows from CNPq. The logistic and material support from the Instituto Nacional de Ciência e Tecnologia de Nanomateriais de Carbono (CNPq) was essential for the execution of the present study.
Abbreviations
- ChAT:
Choline acetyltransferase
- ACh:
Acetyl choline
- AChE:
Acetylcholinesterase
- BuChE:
Butyrylcholinesterase
- UV:
Ultraviolet light
- ACAP:
Antioxidant capacity
- TBARS:
Thiobarbituric acid reactive substances
- EDTA:
Ethylenedioxy-diethylene-dinitrilo-tetraacetic acid
- EGTA:
Ethylene glycol bis(beta amino ethylether)-N,N,N′,N′-tetraacetic acid
- DTNB:
5,5′-Dithiobis-2-nitrobenzoic acid
- HEPES:
4-(2-Hydroxyethyl)-1-piperazineethanesulfonic acid
- BHT:
Butylated hydroxytoluene
- ABAP:
2,2′-Azobis(2-methylpropionamidine) dihydrochloride
- KCl:
Potassium chloride
- SDS:
Sodium dodecyl sulfate
- TMP:
Tetramethoxypropane
- DMSO:
Dimethyl sulfoxide
- MeOH:
Methanol
- ACSCh:
Acetylthiocholinex hydrolysis
- SCh:
Thiocholine
- ROS:
Reactive oxygen species.
References
- 1.ISO/TC 229. http://www.iso.org/iso/iso_technical_committee?commid=381983.
- 2.Project on Emerging Nanotechnologies. http://www.nanotechproject.org/inventories/consumer.
- 3.Oberdörster G, Oberdörster E, Oberdörster J. Nanotoxicology: an emerging discipline evolving from studies of ultrafine particles. Environmental Health Perspectives. 2005;113(7):823–839. doi: 10.1289/ehp.7339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kahru A, Dubourguier H. From ecotoxicology to nanoecotoxicology. Toxicology. 2010;269(2-3):105–119. doi: 10.1016/j.tox.2009.08.016. [DOI] [PubMed] [Google Scholar]
- 5.Kroto HW, Heath JR, O’Brien SC, Curl RF, Smalley RE. C60: buckminsterfullerene. Nature. 1985;318(6042):162–163. [Google Scholar]
- 6.Prato M. [60]Fullerene chemistry for materials science applications. Journal of Materials Chemistry. 1997;7(7):1097–1109. [Google Scholar]
- 7.Montellano A, Da Ros T, Bianco A, Prato M. Fullerene C60 as a multifunctional system for drug and gene delivery. Nanoscale. 2011;3(10):4035–4041. doi: 10.1039/c1nr10783f. [DOI] [PubMed] [Google Scholar]
- 8.Kolosnjaj J, Szwarc H, Moussa F. Toxicity studies of fullerenes and derivatives. Advances in Experimental Medicine and Biology. 2007;620:168–180. doi: 10.1007/978-0-387-76713-0_13. [DOI] [PubMed] [Google Scholar]
- 9.Sayes CM, Gobin AM, Ausman KD, Mendez J, West JL, Colvin VL. Nano-C60 cytotoxicity is due to lipid peroxidation. Biomaterials. 2005;26(36):7587–7595. doi: 10.1016/j.biomaterials.2005.05.027. [DOI] [PubMed] [Google Scholar]
- 10.Oberdörster E. Manufactured nanomaterials (fullerenes, C60) induce oxidative stress in the brain of juvenile largemouth bass. Environmental Health Perspectives. 2004;112(10):1058–1062. doi: 10.1289/ehp.7021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Shinohara N, Matsumoio T, Gamo M, et al. Is lipid peroxidation induced by the aqueous suspension of fullerene C60 nanoparticles in the brains of Cyprinus carpio? Environmental Science and Technology. 2009;43(3):948–953. doi: 10.1021/es802182f. [DOI] [PubMed] [Google Scholar]
- 12.Soreq H, Seidman S. Acetylcholinesterase—new roles for an old actor. Nature Reviews Neuroscience. 2001;2(4):294–302. doi: 10.1038/35067589. [DOI] [PubMed] [Google Scholar]
- 13.Hernández PP, Allende ML. Zebrafish (Danio rerio) as a model for studying the genetic basis of copper toxicity, deficiency, and metabolism. The American Journal of Clinical Nutrition. 2008;88(3):835S–839S. doi: 10.1093/ajcn/88.3.835S. [DOI] [PubMed] [Google Scholar]
- 14.Bertrand C, Chatonnet A, Takke C, et al. Zebrafish acetylcholinesterase is encoded by a single gene localized on linkage group 7. Gene structure and polymorphism; molecular forms and expression pattern during development. The Journal of Biological Chemistry. 2001;276(1):464–474. doi: 10.1074/jbc.M006308200. [DOI] [PubMed] [Google Scholar]
- 15.van Dyk JS, Pletschke B. Review on the use of enzymes for the detection of organochlorine, organophosphate and carbamate pesticides in the environment. Chemosphere. 2011;82(3):291–307. doi: 10.1016/j.chemosphere.2010.10.033. [DOI] [PubMed] [Google Scholar]
- 16.Rico EP, Rosemberg DB, Senger MR, et al. Methanol alters ecto-nucleotidases and acetylcholinesterase in zebrafish brain. Neurotoxicology and Teratology. 2006;28(4):489–496. doi: 10.1016/j.ntt.2006.05.001. [DOI] [PubMed] [Google Scholar]
- 17.Richetti SK, Rosemberg DB, Ventura-Lima J, Monserrat JM, Bogo MR, Bonan CD. Acetylcholinesterase activity and antioxidant capacity of zebrafish brain is altered by heavy metal exposure. NeuroToxicology. 2011;32(1):116–122. doi: 10.1016/j.neuro.2010.11.001. [DOI] [PubMed] [Google Scholar]
- 18.Monserrat JM, Yunes JS, Bianchini A. Effects of Anabaena spiroides (cyanobacteria) aqueous extracts on the acetylcholinesterase activity of aquatic species. Environmental Toxicology and Chemistry. 2001;20(6):1228–1235. doi: 10.1897/1551-5028(2001)020<1228:eoasca>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 19.Pereira VM, Bortolotto JW, Kist LW, et al. Endosulfan exposure inhibits brain AChE activity and impairs swimming performance in adult zebrafish (Danio rerio) NeuroToxicology. 2012;33(3):469–475. doi: 10.1016/j.neuro.2012.03.005. [DOI] [PubMed] [Google Scholar]
- 20.Senger MR, Seibt KJ, Ghisleni GC, Dias RD, Bogo MR, Bonan CD. Aluminum exposure alters behavioral parameters and increases acetylcholinesterase activity in zebrafish (Danio rerio) brain. Cell Biology and Toxicology. 2011;27(3):199–205. doi: 10.1007/s10565-011-9181-y. [DOI] [PubMed] [Google Scholar]
- 21.Rico EP, Rosemberg DB, Dias RD, Bogo MR, Bonan CD. Ethanol alters acetylcholinesterase activity and gene expression in zebrafish brain. Toxicology Letters. 2007;174(1–3):25–30. doi: 10.1016/j.toxlet.2007.08.005. [DOI] [PubMed] [Google Scholar]
- 22.Tang R, Dodd A, Lai D, McNabb WC, Love DR. Validation of zebrafish (Danio rerio) reference genes for quantitative real-time RT-PCR normalization. Acta Biochimica et Biophysica Sinica. 2007;39(5):384–390. doi: 10.1111/j.1745-7270.2007.00283.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Isaacson CW, Usenko CY, Tanguay RL, Field JA. Quantification of fullerenes by LC/ESI-MS and its application to in vivo toxicity assays. Analytical Chemistry. 2007;79(23):9091–9097. doi: 10.1021/ac0712289. [DOI] [PubMed] [Google Scholar]
- 24.Kim KT, Jang MH, Kim JY, Kim SD. Effect of preparation methods on toxicity of fullerene water suspensions to Japanese medaka embryos. Science of the Total Environment. 2010;408(22):5606–5612. doi: 10.1016/j.scitotenv.2010.07.055. [DOI] [PubMed] [Google Scholar]
- 25.Ji Z, Zhang D, Li L, et al. The hepatotoxicity of multi-walled carbon nanotubes in mice. Nanotechnology. 2009;20(44) doi: 10.1088/0957-4484/20/44/445101.445101 [DOI] [PubMed] [Google Scholar]
- 26.Zhang D, Deng X, Ji Z, et al. Long-term hepatotoxicity of polyethylene-glycol functionalized multi-walled carbon nanotubes in mice. Nanotechnology. 2010;21(17) doi: 10.1088/0957-4484/21/17/175101.175101 [DOI] [PubMed] [Google Scholar]
- 27.Clichici S, Mocan T, Filip A, et al. Blood oxidative stress generation after intraperitoneal administration of functionalized single-walled carbon nanotubes in rats. Acta Physiologica Hungarica. 2011;98(2):231–241. doi: 10.1556/APhysiol.98.2011.2.15. [DOI] [PubMed] [Google Scholar]
- 28.Kusters KA, Wijers JG, Thoenes D. Particle sizing by laser diffraction spectrometry in the anomalous regime. Applied Optics. 1991;30(33):4839–4847. doi: 10.1364/AO.30.004839. [DOI] [PubMed] [Google Scholar]
- 29.Phelps HA, Runft DL, Neely MN. Adult zebrafish model of streptococcal infection. Current Protocols in Microbiology. 2009;9:p. 9D.1. doi: 10.1002/9780471729259.mc09d01s13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.da Rocha AM, Ferreira JR, Barros DM, et al. Gene expression and biochemical responses in brain of zebrafish Danio rerio exposed to organic nanomaterials: carbon nanotubes (SWCNT) and fullerenol (C60(OH)18-22(OK4)) Comparative Biochemistry and Physiology A. 2013;165(4):460–467. doi: 10.1016/j.cbpa.2013.03.025. [DOI] [PubMed] [Google Scholar]
- 31.Seibt KJ, Oliveira RDL, Rico EP, Dias RD, Bogo MR, Bonan CD. Antipsychotic drugs inhibit nucleotide hydrolysis in zebrafish (Danio rerio) brain membranes. Toxicology In Vitro. 2009;23(1):78–82. doi: 10.1016/j.tiv.2008.10.003. [DOI] [PubMed] [Google Scholar]
- 32.Siebel AM, Rico EP, Capiotti KM, et al. In vitro effects of antiepileptic drugs on acetylcholinesterase and ectonucleotidase activities in zebrafish (Danio rerio) brain. Toxicology In Vitro. 2010;24(4):1279–1284. doi: 10.1016/j.tiv.2010.03.018. [DOI] [PubMed] [Google Scholar]
- 33.Ellman GL, Courtney KD, Andres V, Jr., Featherstone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochemical Pharmacology. 1961;7(2):88–95. doi: 10.1016/0006-2952(61)90145-9. [DOI] [PubMed] [Google Scholar]
- 34.Amado LL, Garcia ML, Ramos PB, et al. A method to measure total antioxidant capacity against peroxyl radicals in aquatic organisms: application to evaluate microcystins toxicity. Science of the Total Environment. 2009;407(6):2115–2123. doi: 10.1016/j.scitotenv.2008.11.038. [DOI] [PubMed] [Google Scholar]
- 35.Oakes KD, van der Kraak GJ. Utility of the TBARS assay in detecting oxidative stress in white sucker (Catostomus commersoni) populations exposed to pulp mill effluent. Aquatic Toxicology. 2003;63(4):447–463. doi: 10.1016/s0166-445x(02)00204-7. [DOI] [PubMed] [Google Scholar]
- 36.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Analytical Biochemistry. 1976;72(1-2):248–254. doi: 10.1006/abio.1976.9999. [DOI] [PubMed] [Google Scholar]
- 37.Rubin LF. Toxicologic update of dimethyl sulfoxide. Annals of the New York Academy of Sciences. 1983;411:6–10. doi: 10.1111/j.1749-6632.1983.tb47278.x. [DOI] [PubMed] [Google Scholar]
- 38.Yamada T, Jung D, Sawada R, Matsuoka A, Nakaoka R, Tsuchiya T. Effects intracerebral microinjection and intraperitoneal injection of [60]Fullerene on brain functions differ in rats. Journal of Nanoscience and Nanotechnology. 2008;8(8):3973–3980. doi: 10.1166/jnn.2008.414. [DOI] [PubMed] [Google Scholar]
- 39.Mokrushin AA. Neurotoxic effects of fullerenes on the electrical activity of surviving sections of the rat brain olfactory cortex. Doklady Biological Sciences. 2001;377(1–6):122–124. [Google Scholar]
- 40.Henry TB, Menn F, Fleming JT, Wilgus J, Compton RN, Sayler GS. Attributing effects of aqueous C60 nano-aggregates to tetrahydrofuran decomposition products in larval zebrafish by assessment of gene expression. Environmental Health Perspectives. 2007;115(7):1059–1065. doi: 10.1289/ehp.9757. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Zhu X, Zhu L, Lang Y, Chen Y. Oxidative stress and growth inhibition in the freshwater fish Carassius auratus induced by chronic exposure to sublethal fullerene aggregates. Environmental Toxicology and Chemistry. 2008;27(9):1979–1985. doi: 10.1897/07-573.1. [DOI] [PubMed] [Google Scholar]
- 42.Kamat JP, Devasagayam TPA, Priyadarsini KI, Mohan H. Reactive oxygen species mediated membrane damage induced by fullerene derivatives and its possible biological implications. Toxicology. 2000;155(1–3):55–61. doi: 10.1016/s0300-483x(00)00277-8. [DOI] [PubMed] [Google Scholar]
- 43.Totsuka Y, Higuchi T, Imai T, et al. Genotoxicity of nano/microparticles in in vitro micronuclei, in vivo comet and mutation assay systems. Particle and Fibre Toxicology. 2009;6(article 23) doi: 10.1186/1743-8977-6-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Jani P, Halbert GW, Langridge J, Florence AT. Nanoparticle uptake by the rat gastrointestinal mucosa: quantitation and particle size dependency. Journal of Pharmacy and Pharmacology. 1990;42(12):821–826. doi: 10.1111/j.2042-7158.1990.tb07033.x. [DOI] [PubMed] [Google Scholar]
- 45.Sarlo K, Blackburn KL, Clark ED, et al. Tissue distribution of 20 nm, 100 nm and 1000 nm fluorescent polystyrene latex nanospheres following acute systemic or acute and repeat airway exposure in the rat. Toxicology. 2009;263(2-3):117–126. doi: 10.1016/j.tox.2009.07.002. [DOI] [PubMed] [Google Scholar]
- 46.Taylor P, Radić Z. The cholinesterases: from genes to proteins. Annual Review of Pharmacology and Toxicology. 1994;34:281–320. doi: 10.1146/annurev.pa.34.040194.001433. [DOI] [PubMed] [Google Scholar]
- 47.Matsumura F. Toxicology of Insecticides. 2nd edition. New York, NY, USA: Plenum Press; 1985. [Google Scholar]
- 48.Zhang XJ, Yang L, Zhao Q, et al. Induction of acetylcholinesterase expression during apoptosis in various cell types. Cell Death and Differentiation. 2002;9(8):790–800. doi: 10.1038/sj.cdd.4401034. [DOI] [PubMed] [Google Scholar]
- 49.Jiang H, Zhang XJ. Acetylcholinesterase and apoptosis: a novel perspective for an old enzyme. FEBS Journal. 2008;275(4):612–617. doi: 10.1111/j.1742-4658.2007.06236.x. [DOI] [PubMed] [Google Scholar]
- 50.Kist LW, Rosemberg DB, Pereira TCB, et al. Microcystin-LR acute exposure increases AChE activity via transcriptional ache activation in zebrafish (Danio rerio) brain. Comparative Biochemistry and Physiology C. 2012;155(2):247–252. doi: 10.1016/j.cbpc.2011.09.002. [DOI] [PubMed] [Google Scholar]
- 51.Henry TB, Petersen EJ, Compton RN. Aqueous fullerene aggregates (nC60) generate minimal reactive oxygen species and are of low toxicity in fish: a revision of previous reports. Current Opinion in Biotechnology. 2011;22(4):533–537. doi: 10.1016/j.copbio.2011.05.511. [DOI] [PubMed] [Google Scholar]
- 52.Usenko CY, Harper SL, Tanguay RL. Fullerene C60 exposure elicits an oxidative stress response in embryonic zebrafish. Toxicology and Applied Pharmacology. 2008;229(1):44–55. doi: 10.1016/j.taap.2007.12.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Melo JB, Agostinho P, Oliveira CR. Involvement of oxidative stress in the enhancement of acetylcholinesterase activity induced by amyloid beta-peptide. Neuroscience Research. 2003;45(1):117–127. doi: 10.1016/s0168-0102(02)00201-8. [DOI] [PubMed] [Google Scholar]