

Abstract
Oligoethylene glycol decorated supramolecular assemblies have been of great interest due to their charge-neutral character and thus the propensity to avoid non-specific interactions. These systems are known to exhibit a macroscopic temperature-sensitive transition, where the assembly phase-separates from the aqueous phase at higher temperatures. While this so-called lower critical solution temperature (LCST) behavior has been well-studied, there have been no studies on the fate of these supramolecular assemblies below this transition temperature. The work here brings to light the presence of a second, sub-LCST transition, observed well below the LCST of oligoethylene glycol (OEG) based dendrons, where the host-guest properties of the assembly are significantly altered. This sub-LCST transition is accompanied by changes in the guest encapsulation stability and dynamics of host exchange.
Introduction
Stimuli sensitive supramolecular assemblies have captured our attention, because of their potential to impact a variety of applications including biosensing, drug delivery and diagnostics.1–9 It is often desirable, in many of these applications, that the surface moieties interfacing with the aqueous milieu do not exhibit any non-specific binding characteristics. Oligo- and poly-ethylene glycol based hydrophilic functional groups have been quite popular in this context.10–13 In addition to the desirable non-specific binding features, these functional groups are also capable of imparting temperature-sensitive character to a supramolecular assembly.14–17 Considering the prevalence of ethylene glycol based functional groups in biomaterials, it is critical that we understand the factors that underlie the thermal sensitivity. It is widely accepted that the thermal sensitivity arises from the fact that the degree of hydrogen bonding between the ethylene glycol moieties and water is inversely proportional to temperature.14,18,19 This feature has been observed in the form of macroscopic phase changes, where a polymer or a supramolecular assembly phase separates from the aqueous phase in response to increased temperature. This phase transition is reported as the molecule’s lower critical solution temperature (LCST) or the cloud point.20,21 Molecules are soluble below their cloud point, but will precipitate above this temperature due to weakened hydrogen bonding with the aqueous medium. In effect, ethylene glycol units become less hydrated (i.e. less hydrophilic) in an elevated thermal environment, resulting in increased intermolecular aggregation and ultimately precipitation. Numerous reports describe cloud point mediated triggers in drug delivery,22–24 sensing,25–27 and catalysis28–30 as precipitation mediated responses. While the cloud point phase transition is an easily observed macroscopic phenomenon, we have found no studies investigating the thermal properties of these molecules below their cloud point. This is especially important in amphiphilic assemblies that feature these temperature-sensitive functional groups, because the change in hydrophilic-lipophilic balance (HLB) above and below the LCST results in dramatically altered supramolecular structures. Since the fidelity of an amphiphilic assembly depends on the HLB,31–33 if there is a temperature-dependent effect prior to the macroscopic phase change in its building blocks, then it will likely alter the assembly itself. Here, we report such a phenomenon in facially amphiphilic dendrons.34,35 Specifically, we have observed a sub-LCST transition in which the molecular organization, host-guest encapsulation properties, and dynamics of the supramolecular assemblies are affected by temperature (Figure 1). In this paper, we explore the previously unreported thermoresponsive behavior as demonstrated by temperature dependent dynamic light scattering (DLS), dynamic FRET-based guest exchange, and host exchange as monitored by an excimer-based fluorescence probe. This work provides insight into the self-assembly and thermoresponsive properties of ethylene glycol containing molecules by experimentally revealing a sub-LCST dynamic to static supramolecular transition.
Figure 1.
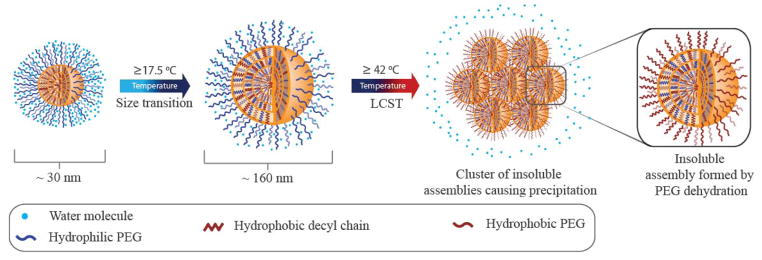
Hypothetical schematic representation of the proposed sub-LCST supramolecular transition at ~ 17.5 °C and LCST at ~ 42 °C respectively.
We chose oligoethylene glycol (OEG) containing facially amphiphilic dendrons (Chart 1) for this study, because: (i) these dendrons are known to exhibit LCST behavior;17 (ii) the monodisperse nature of the dendrons33,36–39 provides the opportunity for systematic variation in the structure of the building blocks of the amphiphilic assembly and investigate its effect upon the temperature-dependent host characteristics; and (iii) the amphiphilic assemblies from these dendrons can act as hosts for non-covalently binding guest molecules.
Chart 1.

Structure of temperature sensitive facially amphiphilic dendrons
Results and Discussion
Aggregation properties
Facially amphiphilic dendrons, with a five repeat OEG unit as the hydrophilic moiety and decyl group as the lipophilic moiety, have been shown to organize into spherical assemblies in the aqueous phase.17 The host-guest studies revealed that these assemblies contain a hydrophobic interior for sequestering lipophilic guest molecules, reminiscent of micelle-like structures. However, size measurements using dynamic light scattering (DLS) and transmission electron microscopy (TEM) reveal that these are about 160 nm in diameter at 25 °C.17 This rather large size, for a micellar assembly, and the facially amphiphilic structure of the dendrons led us to assume that the hydrophobic interior (as probed by the microenvironment of fluorescent guest molecules) is composed of lipophilic alkyl and buried OEG moieties. This is understandable, because unhydrated OEG units are thought to be hydrophobic. Considering these possibilities, we were interested in investigating whether the temperature-dependent hydration of the OEG units would alter the HLB, where a greater number of OEG units are exposed to the solvent at lower temperatures.
To investigate this, we first measured the size of the assembly obtained from our first generation facially amphiphilic dendron 1, using DLS at 10 °C. Indeed, we found that the size of the amphiphilic assembly formed from the dendron is much smaller (~30 nm), compared to that observed at ambient temperature (~160 nm) (Figure 2a). Considering this observation, we carried out a systematic temperature-dependent DLS studies for the dendron 1. The results from these studies highlight a sharp transition in the size of the aggregate between 15 and 17.5 °C (Figure 2b). The PDI of all assemblies were between 0.110–0.306 with correlation functions of >0.82. The correlation functions were slightly lower (0.70 and 0.71) for the smaller sized assemblies at the two lowest temperatures studied, 10 and 12.5 °C (Figure S2). Note that the classical cloud point or the LCST transition for this molecule is 42 °C (Figure S7) 17 and there seems to be a previously unobserved transition at a lower temperature. Interestingly, when the second and third generation dendrons (2 and 3) were investigated, no temperature-dependent size change was observed at lower temperatures within the temperature ranges investigated; the LCST of these dendrons were 32 and 31 °C respectively.17 Thus, following are the noteworthy features of this preliminary finding: (i) dendron 1 exhibits a macrophase separation, resulting in an anisotropic mixture at 40 °C, the so-called LCST; (ii) below the LCST, some higher order aggregation does occur, but the assembly does not exhibit any macrophase separation; (iii) far below the LCST, there exists a second, sub-LCST transition, resulting in smaller aggregates presumably due to greater hydration of the OEG units; (iv) this three phase system is unique to 1, when compared to second and third generation dendrons (2 and 3). We speculate that the higher number of amphiphilic units that are covalently tethered in these dendrons results in a larger energetic penalty for reorganizing the assembly formed at ambient temperature, thus preventing a sub-LCST transition.40,41 It is critical then to understand the dynamic nature of the supramolecular assembly from the first generation dendron, compared to the second and third generation facially amphiphilic dendrons.
Figure 2.
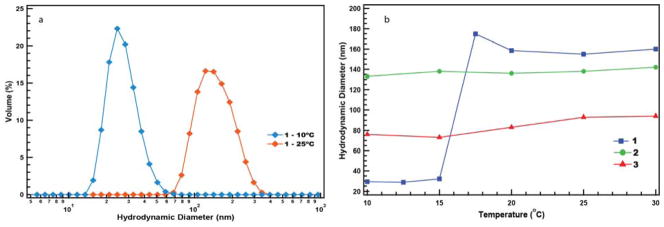
Temperature dependent size variations as observed by dynamic light scattering (DLS). a) A large change in the DH of 1 was observed for 25 °C (160 nm) and 10 °C (30 nm) assemblies. b) The temperature responsiveness of dendrons 1, 2, and 3 were determined. Dendron 1 showed a sharp change in the hydrodynamic radius between 15 and 17.5 °C, while the assemblies from dendrons 2 and 3 were temperature insensitive.
Temperature-dependent host exchange
At first glance, it seems obvious that a size change with decreasing temperature should dictate that the dendron host also exchanges. Note that our previous experiments do suggest that there would be a change in the size upon decrease in temperature. However, once an assembly is at a particular temperature, we do not know whether or not the host molecule rapidly exchanges among the amphiphilic assemblies. Therefore, it is interesting to investigate whether host exchange dynamics is dependent on temperature. For this purpose, we synthesized a G1 dendron (6) with a pyrene moiety “clicked” at its focal point. Dendron 6 forms an assembly, much like 1, with the exception that we now have a covalently bound fluorescent probe at the interior of the aggregate with which we can monitor thermosensitivity of the host exchange. In the amphiphilic supramolecular assembly, pyrene units would be forced into close proximity. This state can be probed through the formation of an excited state dimer (excimer), which spectroscopically reveals itself through a broad emission peak with a large Stokes shift. If we mix the dendron 6 with the pyrene-less dendron 1 and if there were a rapid exchange of the dendron molecules among the aggregates, then the concentration of the pyrene units within an aggregate will decrease (Figure 3a). This decrease should result in reduced excimer emission and corresponding increase in the emission that corresponds to the pyrene monomer. To first investigate the viability of this experiment, we first mixed different concentrations of dendron 1 with a 20 μM solution of 6 and monitored the pyrene fluorescence. Note that the pyrene excimer emission indeed decreases with a concurrent increase in the monomeric emission, when the concentration of 1 is increased in the solution (Figure 3b).
Figure 3.

Dendron exchange via mixed micellar assemblies. Exchange rates are extracted using a covalently linked pyrene probe 6 in a mixed assembly experiment. a) Dendritic supramolecular assemblies are in equilibrium with individual dendrons in solution, mixing 1 and 6 results in a mixed assembly (1*6), where the effective concentration of pyrene units will be reduced. b) The change in the excimer/monomer ratio upon 1*6 formation allows us to directly monitor dendron exchange via time-lapse fluorescence measurements. c) Mixing solutions of 1 and 6 at multiple temperatures while monitoring the pyrene monomer emission shows a distinct change in dendron exchange rates. d) Ramping the temperature by 1 °C increments from 25 °C to 19 °C shows the dramatic change in the equilibrium between supramolecular assembly and individual dendron units in solution. The change in assembly dynamics, coupled with the change in hydrodynamic radius suggests the presence of two organizationally distinct assemblies below the LCST of the material.
To allow for a direct correlation between dendron exchange rates and temperature-sensitivity, we used time-lapse fluorescence where we monitored the increase in monomer emission with time. Solutions of 1 and 6 (25 μM) were brought to the target temperature (±1 °C) for 30 minutes prior to mixing. The solutions were then combined in the fluorescence cuvette holder at a pre-equilibrated temperature. The emission intensity at 379 nm (monomeric pyrene) was monitored with one measurement every 10 seconds (Figure 3c). The peak intensity at 379 nm increased rapidly, when the temperature was 10 °C. On the other hand, there was no change in the emission intensity at 25 °C. In other words, the dynamics of host exchange is faster at lower temperatures. If our hypothesis that the higher generation dendrons pay a larger energetic penalty for dissociating from the amphiphilic aggregate were correct, then those two dendrons should not exhibit host exchange at any temperature (or exhibit this behavior at even lower temperatures). Indeed, dendrons 2 and 3 did not exhibit any exchange over the temperature range of 10 °C – 25 °C, when the experiments were carried out with a mixture of these dendrons with the dendron 6 (Supplementary Figure S5). These results further support the tighter association of the host molecules in the higher generation dendrons.
In analyzing the data in Figure 3c, we noticed not only a strong temperature dependence upon the dendron exchange, but also that the transition from a non-exchanging mixture to an exchanging mixture was rather sharp between 22.5 °C and 20 °C. Assuming first order, we calculated the rate constants at these temperatures using their half-lives and found that there was a significant increase in rate from 22.5 °C to 20 °C, where the rate constants obtained at these temperatures were 8.5468 × 10−4 s−1 and 2.6557 × 10−3 s−1 respectively. The rate further increases with decreasing temperature, and reaches 8.557 × 10−3 s−1 at 10 °C, which is an order of magnitude higher than the rate observed at 22.5 °C (Figure S8). To independently investigate the sharpness of the transition observed in Figure 3c, from a static to a dynamic assembly, the temperature of a single mixed micelle solution was decreased from 25 °C to 19 °C by one degree increments. The 1*6 solution was allowed to equilibrate at each temperature and the exchange coefficients were calculated. The transition from negligible dendron exchange to a very rapidly exchanging assembly occurs over ~2 °C range (Figure 3d). The slight discrepancy in the transition temperature range from the temperature ramping experiments and constant temperature mixing experiments is attributed to the possible difference in pre-equilibration times. Finally to understand if the increase in dynamics of host exchange observed at lower temperatures (Figure 3c) is associated with any sub-LCST size transition, variable temperature DLS was performed with assemblies from 6 and 1*6. Surprisingly assembly from 6 alone did not show a size transition at lower temperatures, presumably due the increased hydrophicity in lipophilic chain due to pyrene. However assembly from 1*6 did show a temperature dependent size transition suggesting that the host 6 is indeed dynamic at lower temperatures (Figure S10).
Guest exchange properties
Considering the temperature-dependent assembly behavior, as well as host exchange, we were interested in investigating the implications of this behavior in guest exchange. We neither anticipated nor observed any significant difference in the extent of encapsulation of guest molecules based on minor temperature variations in the assembly. However, we were interested in identifying the implications of the temperature-dependent changes in the amphiphilic assembly upon the dynamics of guest exchange between the host and the bulk solvent, which is referred to as the encapsulation stability.42 Note that the host-guest interactions in these amphiphilic assemblies are driven by solvophobic interactions. Therefore, if the solvation of the host molecule were to differ with temperatures, then it should follow that the encapsulation stability is also different. That is, the dynamics of guest exchange should be higher at lower temperatures, since the dendron becomes more hydrophilic at lower temperature. In terms of the general relationship between dynamics and temperature, this seems counter-intuitive at first. However, when we account for the solvation of the host, this seems possible. Dynamics of guest exchange and thus the leakage coefficient can be measured using a recently-developed fluorescence resonance energy transfer (FRET) based method.42 Briefly here, two different solutions of the host-guest assembly are generated – one containing a FRET donor dye as the guest molecule and the other containing the corresponding FRET acceptor guest molecule. When these two solutions are mixed, if there is a rapid guest exchange the FRET donor and acceptor guest molecules will result in the same supramolecular assembly; this can be discerned by the decrease in the donor emission with the concurrent increase in the acceptor emission, when exciting the donor molecule. If there is no guest exchange, there should be no evolution of the relative emission intensities with time.
In our experiments, we used 3,3′-dioctadecyloxacarbocyanine perchlorate (DiO – FRET donor) and 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate (DiI – FRET acceptor) as the lipophilic FRET pair. The dye molecules were encapsulated in 1 in separate solutions, referred to as 1-DiO and 1-DiI. The two solutions were then mixed and the evolution of FRET was monitored over time (Figure 4a). Indeed, when selectively exciting the DiO molecule in the solution, the emission intensity from DiO decreased with time, while the emission from DiI increased. The leakage coefficient (Λ), which is the slope of the linear fit of the FRET ratio over time at initial times, was found to be Λ = 0.05 for 1 at 25 °C (Figure 4b). The FRET ratio is defined as Ia/(Ia+Id), where Ia and Id are the intensity of the acceptor and donor emissions at their respective emission maximum.42,43
Figure 4.

a) Excitation of DiO at 450 nm results in FRET, when a mixed assembly is present. b) The leakage coefficient (Λ) is derived from the acceptor – donor ratio as the slope of the FRET ratio (see inset for linear region of dye exchange). c) The temperature sensitivity of 1 was shown to have an inverse effect on the guest exchange dynamics with exchange at 4 °C completing within 10 minutes and virtually no exchange was observed at 37 °C. d) Guest molecules mixing studies were performed with 1–3 to test any generation dependence on the exchange rates. Exchange was only observed for the first generation dendron 1.
Considering our findings with temperature-dependent change in the size of the amphiphilic assembly, we investigated the dynamics of guest exchange in these assemblies at different temperatures. Mixing experiments utilizing 1-DiO and 1-DiI were first performed at 4 °C and 37 °C. Interestingly, an increase in the exchange dynamics of guest molecules was observed at 4 °C as a complete guest exchange occurs in less than 10 minutes with a Λ of >1.897. Conversely, essentially no FRET evolution was observed at 37 °C, which affords a Λ of ≤ 0.001. Although exchange dynamics is typically expected to be faster at higher temperatures, note that the guest exchange is extremely rapid at lower temperatures and non-existent at higher temperatures in this assembly in the aqueous phase. This is taken to indicate that the guest molecule exchange is heavily influenced by the hydration state of OEG for the first generation dendron 1 (Figure 4c).
In order to further test our hypothesis that the temperature-dependent size changes and guest exchange dynamics are likely due to difference in hydration of the OEG units, we tested the temperature-dependent studies with two control dendrons, the carboxylate dendron 4 and the zwitterionic dendon 5. These dendrons are structurally similar to 1, except that the hydrophilic OEG unit is replaced with carboxylate and zwitterionic moieties respectfully. Both 4 and 5 self-assemble into micelle-like aggregates, which are capable of sequestering guest molecules. Dendron 4 was shown to be temperature insensitive with respect to both aggregate size, and encapsulation stability (Supplementary Figure S3). It should be noted that, as a polyelectrolyte, counter-ion effects could have significant effects on the solution properties of 4. Dendron 5 addresses this concern, as it is a neutral zwitterionic molecule that should not be affected by changes in temperature. Theoretically, both dendrons should be insensitive to any hydrogen bonding effects as the interfacial interaction with water is strong enough not to be affected in the temperature range of this study. We therefore expect no size change as a function of temperature or inverse temperature dependence on guest exchange. We were gratified to observe no difference in assembly size with temperature. Similarly, the FRET-based guest exchange studies with 5 revealed that no guest exchange is observed at room temperature or at lower temperatures. Slightly elevated guest exchange was observed for higher temperatures, most likely due to the classical increased diffusion at higher temperature (Figure S4).
Guest exchange experiments for the higher generation dendrons 2 and 3 showed no FRET evolution with time, indicating that the lipophilic guest molecules are stably encapsulated in these amphiphilic assemblies (Figure 4d). This exchange dynamics, or lack thereof, was found to be temperature insensitive, suggesting that the host properties of higher generation dendrons are not affected by the possible increased hydration of OEG. The insensitivity of the larger dendrons is most likely correlated to a larger energetic penalty of rearrangement, which has the effect of providing a stable hydrophobic environment for guest encapsulation. The difference in guest encapsulation stability of 1 can be explained by considering the two limiting scenarios for the dynamics of guest exchange. The first being where the guest molecules are able to diffuse in and out of the assembly through Fickian diffusion, resulting in an unassisted guest exchange among the host molecules present in solution. The second scenario involves a dynamic exchange of individual dendrons among the aggregates, where a dendron dissociates from an aggregate and the monomeric form then reassembles with one of the other aggregates. Here guest molecules, closely associated with an exchanging amphiphilic dendron, could be simultaneously transported until a recombination event occurs with another assembly. This process would also manifest itself as a guest exchange with respect to the FRET-based measurement. It is also important to note that these processes are not mutually exclusive. In addition, the latter scenario would be consistent with our hypothesis that the larger dendrons afford stable encapsulation, because there is a larger energetic penalty for molecular rearrangement.
The dynamic and static nature of dendrons observed from the host exchange as well as the guest exchange experiments below and above a specific temperature could be viewed as a sub-LCST transition, based on a phenomenon that occurs well below the macrophase separation temperature defined by the LCST (Figure 1). The presence of a sub-LCST in this system and the effect it has upon the host-guest capabilities of the dendron assembly illustrate the importance of the temperature-mediated properties of OEG based supramolecular assemblies. Further computational and spectroscopic studies on these systems are warranted for an even greater understanding into the underlying physical properties affecting OEG hydration and supramolecular behavior.
Conclusions
Study of amphiphilic supramolecular assemblies, containing OEG units as hydrophilic functionalities, at temperature below their LCST reveals that there is also a possibility of sub-LCST transitions in these supramolecular assemblies. We found that: (i) the size of the supramolecular assemblies can significantly change in response to temperature variations. This is attributed to the enhanced hydration of the OEG units in the amphiphilic dendron; (ii) the size change and the ensuing temperature-dependent variations in the host-guest properties of the dendrons is unique to the first generation dendron. Structurally similar, but larger, second and third generation dendrons do not exhibit these features. We speculate that the higher number of amphiphilic units that are covalently tethered in these dendrons results in a larger energetic penalty for reorganizing the assembly formed at ambient temperature; (iii) concurrent with the temperature-dependent size change, the guest encapsulation stability of the dendrons also decreases with decreasing temperature. While guest molecules are stably encapsulated at ambient temperature, the dynamics of guest exchange is much faster at lower temperatures; (iv) in addition to the rapid guest exchange at lower temperatures, the dendritic host also rapidly exchanges at low temperatures. When analyzing the temperature at which the dendritic host transitions from being in a static assembly to a dynamic one, we identified that there is a rather sharp transition temperature, which we name as a sub-LCST transition temperature (Figure 1). The fact that this temperature-dependent host exchange dynamics is present in 1, but not in 2 and 3, further supports the hypothesis that there is a larger energetic penalty for reorganizing the assembly in higher generation dendrons. Amphiphilic assemblies have been of interest for a variety of applications. Ethyleneglycol-based amphiphilic systems are often targeted for biological applications, because these systems are known to provide enhanced circulation times and are known as non-fouling hydrophilic surfaces.13,44,45 The commonly anticipated temperature-dependence feature in these assemblies involves the macroscopic phase separation of these assemblies from solution, often described as the LCST. The findings here, that there can be temperature-dependent transitions in these amphiphilic assemblies well below their LCST and that these sub-LCST transitions have significant impact on the host-guest properties of the assemblies, will have important implications in the design and utility of such amphiphilic molecules in these applications.
Supplementary Material
Chart 2.

Structures of the anionic, zwitterionic, and pyrene labeled dendrons 4, 5, and 6 (used in host exchange studies).
Acknowledgments
Support from NIGMS of the National Institutes of Health (GM-065255) is gratefully acknowledged.
Footnotes
Supporting Information Available: Synthetic procedures and data from NMR, MS, fluorescence, and DLS. This material is available free of charge via the internet at http://pubs.acs.org.
References
- 1.Beer PD, Gale PA. Angew Chem Int Ed. 2001;40:486. [PubMed] [Google Scholar]
- 2.Song J, Cheng Q, Zhu SM, Stevens RC. Biomed Microdevices. 2002;4:213. [Google Scholar]
- 3.González DC, Savariar EN, Thayumanavan S. J Am Chem Soc. 2009;131:7708. doi: 10.1021/ja900579g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Gillies ER, Jonsson TB, Fréchet JMJ. J Am Chem Soc. 2004;126:11936. doi: 10.1021/ja0463738. [DOI] [PubMed] [Google Scholar]
- 5.Jeong B, Bae YH, Lee DS, Kim SW. Nature. 1997;388:860. doi: 10.1038/42218. [DOI] [PubMed] [Google Scholar]
- 6.Bae Y, Fukushima S, Harada A, Kataoka K. Angew Chem Int Ed. 2003;42:4640. doi: 10.1002/anie.200250653. [DOI] [PubMed] [Google Scholar]
- 7.Benvin AL, Creeger Y, Fisher GW, Ballou B, Waggoner AS, Armitage BA. J Am Chem Soc. 2007;129:2025. doi: 10.1021/ja066354t. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Cabral H, Nishiyama N, Kataoka K. Acc Chem Res. 2011;44:999. doi: 10.1021/ar200094a. [DOI] [PubMed] [Google Scholar]
- 9.Nystrom AM, Wooley KL. Acc Chem Res. 2011;44:969. doi: 10.1021/ar200097k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Knop K, Hoogenboom R, Fischer D, Schubert US. Angew Chem Int Ed. 2010;49:6288. doi: 10.1002/anie.200902672. [DOI] [PubMed] [Google Scholar]
- 11.Otsuka H, Nagasaki Y, Kataoka K. Adv Drug Delivery Rev. 2003;55:403. doi: 10.1016/s0169-409x(02)00226-0. [DOI] [PubMed] [Google Scholar]
- 12.Alexis F, Pridgen E, Molnar LK, Farokhzad OC. Mol Pharmaceutics. 2008;5:505. doi: 10.1021/mp800051m. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen H, Yuan L, Song W, Wu ZK, Li D. Prog Polym Sci. 2008;33:1059. [Google Scholar]
- 14.Lutz JF, Akdemir O, Hoth A. J Am Chem Soc. 2006;128:13046. doi: 10.1021/ja065324n. [DOI] [PubMed] [Google Scholar]
- 15.Qin S, Geng Y, Discher DE, Yang S. Adv Mater. 2006;18:2905. [Google Scholar]
- 16.Lutz JF. Adv Mater. 2011;23:2237. [Google Scholar]
- 17.Aathimanikandan SV, Savariar EN, Thayumanavan S. J Am Chem Soc. 2005;127:14922. doi: 10.1021/ja054542y. [DOI] [PubMed] [Google Scholar]
- 18.Rackaitis M, Strawhecker K, Manias E. J Polym Sci, Part B: Polym Phys. 2002;40:2339. [Google Scholar]
- 19.Hammouda B, Ho D, Kline S. Macromolecules. 2002;35:8578. [Google Scholar]
- 20.Becer CR, Hahn S, Fijten MWM, Thijs HML, Hoogenboom R, Schubert US. J Polym Sci, Part A: Polym Chem. 2008;46:7138. [Google Scholar]
- 21.Corti M, Minero C, Degiorgio V. J Phys Chem. 1984;88:309. [Google Scholar]
- 22.Chilkoti A, Dreher MR, Meyer DE, Raucher D. Adv Drug Delivery Rev. 2002;54:613. doi: 10.1016/s0169-409x(02)00041-8. [DOI] [PubMed] [Google Scholar]
- 23.Schmaljohann D. Adv Drug Delivery Rev. 2006;58:1655. doi: 10.1016/j.addr.2006.09.020. [DOI] [PubMed] [Google Scholar]
- 24.Chung JE, Yokoyama M, Yamato M, Aoyagi T, Sakurai Y, Okano T. J Controlled Release. 1999;62:115. doi: 10.1016/s0168-3659(99)00029-2. [DOI] [PubMed] [Google Scholar]
- 25.Kataoka K, Miyazaki H, Okano T, Sakurai Y. Macromolecules. 1994;27:1061. [Google Scholar]
- 26.Koopmans C, Ritter H. J Am Chem Soc. 2007;129:3502. doi: 10.1021/ja068959v. [DOI] [PubMed] [Google Scholar]
- 27.Liu T, Liu S. Anal Chem. 2011;83:2775. doi: 10.1021/ac200095f. [DOI] [PubMed] [Google Scholar]
- 28.Bergbreiter DE, Case BL, Liu YS, Caraway JW. Macromolecules. 1998;31:6053. [Google Scholar]
- 29.Bergbreiter DE, Caraway JW. J Am Chem Soc. 1996;118:6092. [Google Scholar]
- 30.Kimura M, Kato M, Muto T, Hanabusa K, Shirai H. Macromolecules. 2000;33:1117. [Google Scholar]
- 31.Mitchell DJ, Ninham BW. J Chem Soc, Faraday Transactions 2. 1981;77:601. [Google Scholar]
- 32.Schott H. J Pharm Sci. 1995;84:1215. doi: 10.1002/jps.2600841014. [DOI] [PubMed] [Google Scholar]
- 33.Ramireddy RR, Raghupathi KR, Torres DA, Thayumanavan S. New J Chem. 2012;36:340. doi: 10.1039/C2NJ20879B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bharathi P, Zhao HD, Thayumanavan S. Org Lett. 2001;3:1961. doi: 10.1021/ol016064b. [DOI] [PubMed] [Google Scholar]
- 35.Vutukuri DR, Basu S, Thayumanavan S. J Am Chem Soc. 2004;126:15636. doi: 10.1021/ja0449628. [DOI] [PubMed] [Google Scholar]
- 36.Grayson SM, Fréchet JMJ. Chem Rev. 2001;101:3819. doi: 10.1021/cr990116h. [DOI] [PubMed] [Google Scholar]
- 37.Fischer M, Vögtle F. Angew Chem Int Ed. 1999;38:884. doi: 10.1002/(SICI)1521-3773(19990401)38:7<884::AID-ANIE884>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 38.George R, Newkome CNM. Dendrimers and Dendrons: Concepts, Syntheses, Applications. Wiley-VCH; 2001. Fritz Vögtle. [Google Scholar]
- 39.Tomalia Donald A, JBC, Boas Ulrik. Dendrimers, Dendrons, and Dendritic Polymers: Discovery, Applications, and the Future. 1. Cambridge University Press; New York, United States of America: 2012. [Google Scholar]
- 40.Choi SH, Lodge TP, Bates FS. Phys Rev Lett. 2010:104. doi: 10.1103/PhysRevLett.104.047802. [DOI] [PubMed] [Google Scholar]
- 41.Waton G, Michels B, Zana R. J Colloid Interface Sci. 1999;212:593. doi: 10.1006/jcis.1998.6020. [DOI] [PubMed] [Google Scholar]
- 42.Jiwpanich S, Ryu JH, Bickerton S, Thayumanavan S. J Am Chem Soc. 2010;132:10683. doi: 10.1021/ja105059g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nir E, Michalet X, Hamadani KM, Laurence TA, Neuhauser D, Kovchegov Y, Weiss S. J Phys Chem B. 2006;110:22103. doi: 10.1021/jp063483n. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Yebra DM, Kiil S, Dam-Johansen K. Prog Org Coat. 2004;50:75. [Google Scholar]
- 45.Banerjee I, Pangule RC, Kane RS. Adv Mater. 2011;23:690. doi: 10.1002/adma.201001215. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.