

Abstract
The APOBEC family of proteins comprises deaminase enzymes that edit DNA and/or RNA sequences. The APOBEC3 subgroup plays an important role on the innate immune system, acting on host defense against exogenous viruses and endogenous retroelements. The role of APOBEC3 proteins in the inhibition of viral infection was firstly described for HIV-1. However, in the past few years many studies have also shown evidence of APOBEC3 action on other viruses associated with human diseases, including HTLV, HCV, HBV, HPV, HSV-1, and EBV. APOBEC3 inhibits these viruses through a series of editing-dependent and independent mechanisms. Many viruses have evolved mechanisms to counteract APOBEC effects, and strategies that enhance APOBEC3 activity constitute a new approach for antiviral drug development. On the other hand, novel evidence that editing by APOBEC3 constitutes a source for viral genetic diversification and evolution has emerged. Furthermore, a possible role in cancer development has been shown for these host enzymes. Therefore, understanding the role of deaminases on the immune response against infectious agents, as well as their role in human disease, has become pivotal. This review summarizes the state-of-the-art knowledge of the impact of APOBEC enzymes on human viruses of distinct families and harboring disparate replication strategies.
1. Introduction
The human immune system is constantly challenged by invading pathogens, against which it acts by eliminating them or reducing their impact once infection is established. Current and emerging viruses constitute an important fraction of these pathogens that are able to develop short to life-long persistent infections, to some of which no protective vaccines are yet available. In this regard, a better understanding of the mechanisms by which innate and adaptive immunity restricts viral infections and/or modulate viral pathogenesis is urged.
The innate immune system constitutes the first line of defense against viruses, initiating an antiviral response. Viruses are recognized by this system primarily through detection of their nucleic acids, either their packaged genome or viral replication intermediates within the infected cell [1]. Toll-like receptors are good examples of the former viral sensing mechanisms, while the latter are represented by RIG-I-like or DAI and AIM2 receptors [2, 3]. These types of recognition induce the transcription of proinflammatory cytokines and type I interferons (IFNs). These, on their hand, activate the expression of hundreds of IFN-stimulated genes (ISGs) which will engage in counteracting virus replication and spread [4].
Among the ISGs, the genes encoding the family of apolipoprotein B mRNA-editing catalytic polypeptide (APOBEC) cytidine deaminases have been largely studied in the last years. Mounting evidence suggest that these enzymes are key players in restricting infections by different viruses. In this review, we summarize the state-of-the-art knowledge of the impact of APOBEC enzymes on human viruses of distinct families and harboring disparate replication strategies.
2. The APOBEC Family of Deaminases
The APOBEC family of proteins comprises a group of cytidine deaminases that are able to edit DNA and/or RNA sequences. Although it belongs to a larger superfamily of deaminases, APOBECs are restricted to vertebrates [5]. In humans, the family comprises eleven members with distinct functions: activation-induced deaminase (AID) and APOBEC1, whose genes are located in chromosome 12; APOBEC2, whose gene is in chromosome 6; seven APOBEC3 genes, located in chromosome 22; and APOBEC4, whose gene is located in chromosome 1 [6–10]. The members of this family are distinguished by the presence of one or two catalytic domains containing a zinc-binding deaminase motif, characterized by the conserved amino acid sequences H-X-E-X(23–28)-P-C-X(2–4)-C (X is any amino acid; Figure 1) [9]. The deamination mediated by these enzymes involves the hydrolytic removal of the amino group at the C4 position of a cytidine (C) or deoxycytidine (dC), generating a uridine (U) or deoxyuridine (dU) (Figure 1) [11].
Figure 1.
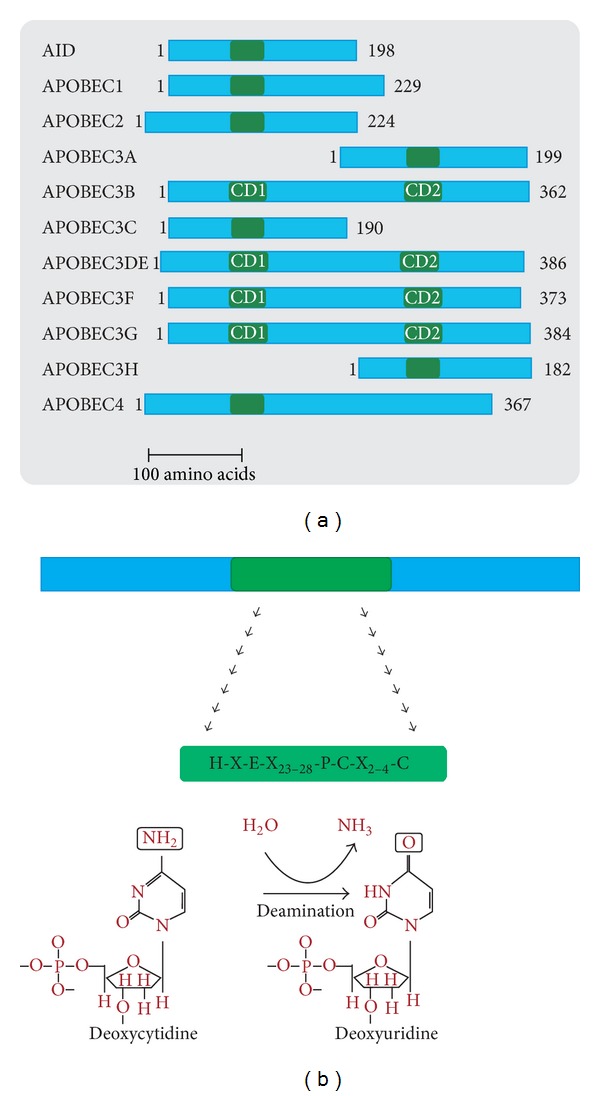
Structure and function of human APOBEC proteins. (a) Schematic representation of APOBEC proteins. The zinc-binding motifs, represented by the catalytic domains (CD) and present in single or double copies, are depicted in green. In the proteins that harbor two CD copies, the N- and C-terminal domains are named CD1 and CD2, respectively. APOBEC proteins are drawn to scale, and the total number of amino acids is shown to the right of each version. The scale bar represents the length of 100 amino acids. (b) The conserved amino acid sequence of the zinc-binding motif is shown; the hydrolytic deamination reaction mediated by these enzymes is shown at the bottom of the figure.
APOBEC1 (A1), the first member of the family to be described, is an RNA-editing enzyme [12] but also presents the ability to edit DNA in bacterial assays [13]. It is primarily expressed in the gastrointestinal compartment and catalyzes the posttranscriptional editing of the apolipoprotein B (apoB) mRNA. A1 is a nucleocytoplasmic protein [14], and the apoB mRNA editing occurs in the nucleus in the presence of the APOBEC-1 complementation factor (ACF) [15, 16]. A1 edits this mRNA at a single base, resulting in the formation of a premature stop codon and leading to the synthesis of a truncated protein (ApoB48) [12]. As a consequence, the human gut produces two forms of ApoB, a longer (ApoB100) and a shorter (ApoB48), involved in the transport of endogenously produced cholesterol and triglycerides and the absorption and transport of exogenous dietary lipids, respectively [17]. Recently, additional mRNA targets of APOBEC1 have been described; the interaction occurs at AU-rich segments of their 3′ untranslated regions. A1 may thus play a role regulating the stability of such specific mRNAs [18, 19].
AID, which deaminates single stranded DNA [20, 21], is predominantly cytoplasmic and shuttles between the nucleus and the cytoplasm [22–25]. It is expressed in germinal center B cells and is essential for the events of class switch recombination and somatic hypermutation during the process of antibody diversification [26–28]. AID has also been reported as involved in DNA demethylation [29–31].
APOBEC2 is expressed in heart and skeletal muscles [8], and although its precise physiologic role is not established, its expression appears to be essential to muscle development [32].
The APOBEC3 group comprises seven proteins in humans: APOBEC3A, APOBEC3B, APOBEC3C, APOBEC3DE, APOBEC3F, APOBEC3G, and APOBEC3H. A3A, A3C, and A3H present one copy of the zinc-binding domain, while the remaining harbor two copies; the N- and C-terminal domains are named CD1 and CD2, respectively (Figure 1). In those three enzymes, only CD2 is catalytically active [13, 33, 34]. A3 enzymes are capable of editing single-stranded DNA and recognize specific target sequences. A3G and A3F, for example, edit C's preferentially at CC and TC dinucleotide contexts (GG and AG in the complementary DNA strand), respectively [35–38].
A3 enzymes play an important role on the innate immune system, acting on host defense against exogenous viruses and endogenous retroelements [39–42]. Viral restriction occurs mainly by their DNA editing mechanism, but A3s also display editing-independent phenotypes [38, 43–45], as will be discussed later. They are further possibly implicated into a specific pathway of exogenous DNA clearance in human cells [46]. More recently, lines of mounting evidence have also shown that A3 enzymes insert mutations in human nuclear and mitochondrial DNA, suggesting roles in DNA catabolism [47]. On the other hand, this phenomenon may represent a possible source of mutations towards the development of cancer [47, 48].
A3 enzymes localize to the cell cytoplasm and/or nucleus, enabling the protection of both compartments through restriction of nuclear (such as the human papillomavirus, the herpes simplex virus, and non-LTR retrotransposons) or cytoplasmic (like the hepatitis B virus, retroviruses, and LTR retrotransposons) replicating elements. A3D, A3F, and A3G are known to be cytoplasmic [40, 49]; A3B localizes to the nucleus [50], while A3A, A3C, and A3H are found both in the nucleus and in the cytoplasm [40, 51]. Noteworthy, different A3H haplotypes present distinct localizations; the protein encoded by haplotype I is mainly nuclear, while the one encoded by haplotype II is predominantly cytoplasmic [51]. With respect to A3A, it has been recently reported that its endogenous version in primary CD14+ monocytes and in the monocytic cell line THP-1 localizes to the cytoplasm, contrasting with the broad nucleocytoplasmic distribution observed upon A3A transfection, an observation likely explained by artificial overexpression of the enzyme [52].
In addition to presenting distinct subcellular localization, some APOBEC3 proteins also display an intracellular mode of regulation by localization into specific subcellular structures. It is known that A3G is present in two distinct molecular forms within the cell: a form of low molecular mass (LMM) and another in ribonucleoproteic complexes of high molecular mass (HMM) [53–55]. The HMM complex is enzymatically inactive and can be converted into LMM complexes, enzymatically active, through RNase digestion [53]. Besides A3G, other APOBECs like A3C, A3F, and A3H also show the ability of assembling into HMM complexes [56–58].
It has been shown that the switch between HMM and LMM can be stimulated by different cytokines [59, 60], and the predominant form varies among different cell types or distinct cell type subsets. The presence of LMM A3G has been related to a reduced susceptibility to HIV-1 infection, as is suggested as postentry restriction factor for this virus [53, 60, 61]. For example, unstimulated peripheral blood CD4+ T-cell lymphocytes and monocytes, which are nonpermissive to HIV-1 infection, present LMM A3G. However, when CD4+ T-cell lymphocytes are activated or the monocytes stimulated to differentiate into macrophages, they shift their A3G profile to HMM [53]. Noteworthy, the knockdown of A3G in unstimulated CD4+ T cells does not turn them permissive to infection, suggesting that the presence of LMM A3G in the cells is not the unique determinant for their resistance to HIV-1 [62, 63]. Moreover, LMM A3G is preferentially packaged into HIV-1 particles [56, 64–66]. Finally, HMM A3G is also able to interact with and sequester Alu RNA elements, inhibiting their transposition and evidencing the role of different A3G molecular forms in the restriction of retroelements [67].
A3G and A3F are also able to accumulate in processing bodies (P-bodies) and stress granules, where they interact with RNAs and several proteins that regulate their metabolism [68–71]. However, the functional consequences of the occurrence of A3 proteins in those structures are not yet clear [72].
APOBEC4 is expressed in testicles, and its function is still unknown [10]. Like A2, A4 does not present mutagenic activity in bacterial or yeast assays [73].
APOBEC proteins are found throughout vertebrates, with AID and APOBEC2 being ancestral members of the family and APOBEC1 and APOBEC3 being more recent, while the origins of APOBEC4 are not clear [74–77]. The APOBEC3 enzymes are exclusively found in mammals [5, 78], and their gene copy number is species-specific. While mice have only a single APOBEC3 gene, pigs have two, sheep and cattle have three, cats have four, horses have six, and primates have at least seven APOBEC3 genes [5, 9, 79, 80].
The evolutionary history of the APOBEC3 genes involves expansion, divergence, selection and extinction of specific A3 copies [80]. It is believed that the genome of the mammalian ancestor encoded for at least one ancestral APOBEC3 gene and that this gene family expanded in the different lineages as a response to changes in viral, retroviral, and retrotransposon pressure [78, 79]. Interestingly, the rapid expansion of the APOBEC3 locus in primates is correlated with a marked reduction in retrotransposon activity, suggesting an important role in the host genome defense against retroelements [81, 82].
There is evidence that APOBEC3 proteins are able to restrict non-LTR and LTR retrotransposons, including both long interspersed nuclear elements (LINEs) and short interspersed nuclear elements (SINEs) [40, 57, 83–87]. While for some murine LTR retrotransposons, like IAP e MusD, DNA deamination was observed as part of the restriction mechanism [39, 85], the exact mechanism and the retrotransposition step targeted by APOBEC3 is unknown for non-LTR elements (reviewed in [82]).
It is interesting to note that AID and APOBEC1 from multiple species have been shown to possess activity against retroelements [86, 88–90] and exogenous viruses [89, 91–93], suggesting that these proteins may also have a role in innate immunity of some vertebrates [82, 89, 92].
3. Role of APOBEC Enzymes on Different Human Viral Infections
3.1. Human Immunodeficiency Virus (HIV)
The human immunodeficiency virus (HIV) is a member of the Retroviridae family, and belongs to the Lentivirus genus, which characterizes viruses of slow symptomatology. As a retrovirus, HIV harbors a genome consisting of two single-stranded RNA molecules of positive polarity that undergoes a reverse transcription step (through a complementary DNA-cDNA-molecule) carried out by its encoded reverse transcriptase (reviewed in [111]). The cDNA is then integrated into the host cell genome, from where the viral genes are transcribed by the host RNA polymerase II. In addition to the essential genes, HIV also encodes several accessory and regulatory proteins that enhance virus replication and burden, being the viral infectivity factor (Vif) among them (reviewed in [111]).
The hypermutation mediated by APOBEC enzymes in HIV type 1 (HIV-1) is well described (Figure 2). It is known that in HIV-1-infected cells, in the absence of a functional Vif, A3G molecules are incorporated into incoming virus particles. This packaging is mediated by the interaction of A3G with the nucleocapsid (NC) domain of the Gag protein [112–116] and occurs in an RNA-dependent manner [114, 115, 117–120]. After a new infection, the editing process occurs during viral reverse transcription. A3G deaminates dC residues in the negative strand of the complementary DNA (cDNA), originating dU. These nucleotides serve as templates for the incorporation of dA in the positive strand and are evidenced as G-to-A changes in the proviral DNA. The frequency of edited dC's in the viral genome can exceed 10% of the sites [38, 121]. The excessive number of changes results in loss of genetic information and production of largely defective virions in the subsequent replication cycle.
Figure 2.
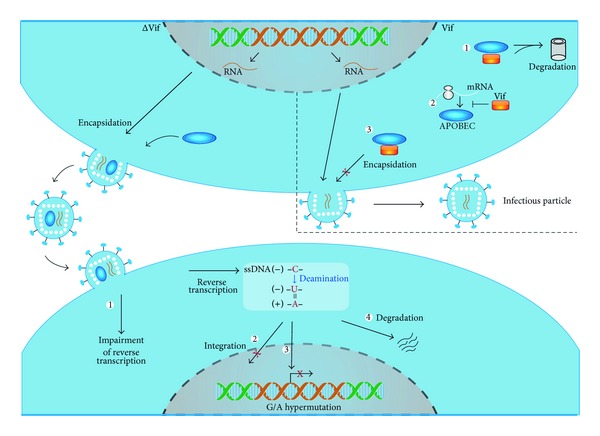
Mechanism of action of APOBEC3 (A3) and Vif in the HIV-1 life cycle. At the top panel, a virus-producing cell in the absence (left) or in the presence (right) of a functional Vif protein is shown. In the presence of Vif (orange rectangle), A3 (blue ellipse) is primarily targeted to proteasomal degradation (1); Vif can also block A3 mRNA translation (2) and prevent A3 packaging into the virion in a degradation-independent manner (3). In the absence of Vif, A3 molecules are packaged into incoming virus particles. After a new infection (bottom panel), A3 exerts its antiviral activity in multiple ways. A3 can interfere with reverse transcription in a deamination-independent manner (1). A3 can also interfere with proviral integration through the formation of abnormal viral DNA ends (2). In the hypermutation process, A3 mainly deaminates dC residues in the negative strand of the complementary viral DNA, originating dU, that serves as template for the incorporation of dA in the positive strand. If able to integrate, hypermutated proviruses are normally largely defective (3). Alternatively, viral DNA containing multiple dU can also be degraded before integration (4).
A reduction of viral reverse transcription products is also observed in the presence of A3G. It has been hypothesized that the presence of dU's in the retroviral DNA could be recognized as anomalous, leading to its degradation even before its integration into the host cell genome (Figure 2). This would occur by removal of the uracil residues by uracil-DNA glycosylases (UDGs), followed by apurinic/apyrimidinic (AP) endonuclease-mediated degradation [122, 123]. Noteworthy, it has been shown that APOBEC-mediated restriction occurs even in the absence of UDGs such as UNG2 and SMUG1 [124, 125], leaving the requirement of retroviral DNA degradation for virus restriction as an open question.
In addition to restricting viral infection through hypermutation, A3G also exert editing-independent mechanisms of restriction. These involve the interference with reverse transcription and with proviral integration by disturbing tRNA primer annealing and removal, DNA synthesis initiation and elongation and strand transfer reaction [43–45, 126–135].
Further to the direct mechanisms of viral inhibition, A3G appears to play a pivotal role in the activation of the host immune system. It has been shown that the generated pool of defective viruses encoding truncated or misfolded proteins represents an important source of viral antigens, associated with a strong activation of HIV-1-specific CD8+ cytotoxic T-cells [136]. APOBEC3 may also enhance the recognition of HIV-infected cells by natural killer cells through the activation of the DNA-damage repair response by viruses harboring uridines in their genomes [137].
HIV-1, like the majority of lentiviruses, counteracts the restriction mediated by the A3 enzymes through expression of the Vif protein [138, 139]. The main mechanism of action assigned to Vif is the induction of A3G protein degradation via proteasome. Vif simultaneously binds to A3 and to an E3 ubiquitin-ligase complex, leading to polyubiquitination of A3G and its consequent degradation [138–141]. Vif is also able to prevent A3G packaging into the virion in a degradation-independent manner and to interfere with A3G mRNA translation, thus reducing the intracellular levels of the protein (Figure 2) [142–147].
APOBEC3F (A3F) has also been associated with HIV restriction in a consistent manner. However, A3G seems to have a more important role in viral restriction of cells targeted by HIV, whereas the role of A3F seems to be dispensable for virus restriction in these cells [148, 149]. A3A, A3B, A3C, A3DE, and some A3H haplotypes have also been implicated in restriction [148, 150–157], although controversial results have been observed (reviewed in [158]).
HIV hypermutated proviral DNA sequences have been reported in several in vivo studies. Yet some reports have shown a correlation between hypermutation and a favorable clinical outcome, this relationship is not consensual. In a population level analysis of HIV-1 subtype B near full-length proviral sequences, hypermutation levels were associated with reduced pretreatment viremia [101]. In agreement with this, higher hypermutation levels were observed in patients with low HIV viral loads (<10,000 copies/mL for at least 3 years) in another study [159]. The presence of hypermutation was also correlated with higher CD4+ T-cell counts [160]. More recently, Kourteva et al. [161] found more A3G-hypermutated sites in proviral sequences derived from HIV long-term nonprogressors (LNTP) compared to noncontrollers (NC). On the other hand, some studies could not find such associations, either in adults or in children [162–164].
A more consistent association has been observed between the APOBEC3 mRNA levels and clinical outcomes. Several studies showed a positive correlation of A3 levels with CD4+ T-cell counts and a negative correlation with HIV-1 viral load [159, 161, 165, 166] or viral set point [167]. However, two studies could not find an association between A3G levels and CD4+ T-cell counts or viral load [104, 168]. APOBEC3G expression has also been inversely correlated with provirus burden [161] and positively correlated with the level of G-to-A changes [159, 161, 162]. A3G has also been found significantly increased in HIV-exposed uninfected (EU) compared to healthy controls [159, 169] or infected individuals [169]. The level of expression in EU significantly decreased after one year from HIV diagnosis and subsequent treatment of their partners. This suggests that, in these individuals, exposure to HIV can trigger APOBEC3G expression in the absence of infection and that the expression decreases with cessation of exposure [159]. Interestingly, the higher expression of APOBEC3G in EU was seen not only in PBMCs but also in cervical tissues, and may be important for the susceptibility to sexually transmitted HIV infection [169].
Higher levels of APOBEC3G mRNA were observed in LTNPs when compared to HIV-uninfected subjects and progressors [165]. Accordingly, higher levels of A3G and A3F were found in LTNP when compared to noncontrollers [161], and higher levels of A3G and A3B were also found in slow progressing patients (SP) when compared to AIDS patients [166]. However, in a group of perinatally HIV-infected children, no correlation was observed between A3G/A3F expression and disease progression [164]. Paradoxically, A3G levels were higher in HIV-negative when compared with HIV positive individuals [104, 166, 168], including matched pre- and postinfection samples from the same subjects. This may suggest that APOBEC3G transcription is rapidly downregulated upon HIV-1 infection [104]. In view of these confounding evidence, additional work is necessary to robustly define the role of A3 expression in the control of HIV infection and disease progression.
Due to the potential antiviral role of the A3 enzymes, therapeutic interventions have been idealized to enhance their action and lead to viral inhibition through hypermutation [170–174]. In this sense, molecules have been identified that are able to interact with A3G and counteract Vif-mediated degradation [171] or to induce Vif degradation in the presence of A3G [172]. On the other hand, it is possible that the editing mediated by the APOBEC enzymes, upon sublethal conditions (which do not drive virus extinction), acts in the diversification of the viral genome, providing source for the selection and evolution of highly fit variants. This can favor, for example, the acquisition of immune system escape mutations or drug resistance mutations [75, 175–182]. In this regard, novel therapeutic strategies based on complete A3G inhibition, eliminating this additional source of virus diversification, have also been proposed [174, 183, 184].
3.2. Human T-Cell Lymphotropic Virus (HTLV)
HTLV is a complex retrovirus belonging to the Deltaretrovirus genus. In most cases infection by HTLV-1 is asymptomatic; however, up to 5% of the infected subjects develop adult T-cell leukemia (ATL) and another 1-2% develop HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) [185, 186].
Like HIV-1, HTLV targets mainly CD4+ T lymphocytes [187], and therefore, it would be exposed to several A3 enzymes that are expressed in this cell type [188]. HTLV does not encode any product with Vif-like, A3 antagonist function, and it is apparently incapable of inducing A3 degradation in cell culture [34, 189]. Unexpectedly, however, hyperedited HTLV sequences appear to be rare. Yet HTLV hypermutated sequences with estimated frequencies between 0.1 and 5% have been observed in vitro, no instance of hypermutation was reported in PBMC from HTLV-infected patients [34]. Moreover, a retrospective analysis of previously published HTLV sequences has identified a single hypermutated sequence recovered from an HTLV-1 infection of an animal model [190].
Although rare in vivo, HTLV hypermutated sequences were recovered from cell lines derived from ATL and HAM/TSP patients. The main context of the observed changes was GG, followed by GA and GC, suggesting the involvement of A3G and also of other A3 such as A3A and A3B in HTLV hypermutation [189]. Despite the fact that A3G is able to edit the viral genome [191, 192], some studies have shown that HTLV-1 is resistant or poorly susceptible to this enzyme [34, 189, 192, 193], in agreement with the low frequency of editing observed in vivo.
Two possible reasons for the low frequency of hypermutated HTLV sequences in vivo, when compared to HIV hypermutation, have been discussed. One of the possibilities resides in the differences of replicative strategies of these two viruses. After primary infection, HTLV presents a low level of productive replication, and the proviral genome is mainly replicated through oligoclonal expansion of infected cells, a fact that contrasts with the high rate of de novo cell infection seen during chronic HIV disease [185, 194–196]. Therefore, it has been suggested that the infrequent replication via reverse transcription seen in HTLV infection represents a reduction of opportunities for APOBEC3-mediated edition to occur. Moreover, a direct resistance mechanism to A3G has been described in HTLV-1. Through a cis-acting exclusion mechanism, elements in the C-terminal region of the HTLV-1 nucleocapsid inhibit A3G packaging to the particle, resulting in reduced efficiency of its packaging in HTLV-1 particles when compared to HIV Δvif virus-like particles [197].
Recently, an analysis of proviral genomes of 60 ATL patients and asymptomatic carriers showed that G-to-A changes are the most frequent nucleotide substitutions. These changes occurred preferentially in the target context of A3G and were involved in the generation of multiple missense substitutions. It was then suggested that HTLV-1-infected cells can take advantage of A3G activity to escape the host immune system by abrogating the expression of viral proteins [191].
3.3. Hepatitis B Virus (HBV)
HBV belongs to the Hepadnaviridae family and presents a circular, partially double stranded DNA genome [198]. It replicates through reverse transcription of an intermediate pregenomic RNA. It is estimated that two billion people have been infected and more than 240 million have chronic liver infections worldwide [199]. HBV infection can cause acute and chronic liver disease, including cirrhosis and hepatocellular carcinoma [198].
A number of studies have shown that the HBV genome is susceptible to human APOBEC enzymes. In addition to A1 and AID, all A3 enzymes except for A3DE were able to edit the viral genome in vitro [200–206] with editing levels estimated between 10−2 and 10−5 [205]. Viral genome editing occurs preferentially in the negative DNA strand resulting in G-to-A changes in the positive strand; C-to-T changes have also been observed in the positive strand, evidencing editing of both viral DNA strands [202].
In addition to the identification of HBV sequences extensively edited by APOBEC enzymes, several studies have shown evidence of restriction to virus replication in vitro, by both editing-dependent and -independent mechanisms [201, 207, 208]. In this sense, a strong inhibitory effect of A3G has been described [201, 207, 208], and a reduction of approximately 30-fold in the levels of viral DNA in the presence of A3G expression has been shown [204]. Turelli et al. [207] showed that A3G leads to a reduction of viral DNA and also of core-associated RNA; this effect was sustained when a catalytically-inactive A3G mutant was used. Among the possible deamination-independent mechanisms of action are the inhibition of pregenomic RNA packaging and the interference of reverse transcription [201, 207, 208]. Additional unrelated factors may account for the suppression of HBV replication in hepatocytes, including the inhibition of HBV transcription through the interaction of A3B with the heterogeneous nuclear ribonucleoprotein K (hnRNP-K), a positive regulator of HBV expression [209].
In the healthy human liver, low to moderate APOBEC expression levels are observed [9, 188, 210]. However, some of these enzymes, particularly A3G, have their expression significantly increased in primary hepatocytes and in hepatoma cell lines in response to stimulation by interferon alpha and gamma [210–212]. In agreement with those data, low hypermutation levels (from 10−4 to 0.6%, depending on the method used) have been described in patients with acute and chronic HBV infection [202, 213, 214]. This has led to the hypothesis that hypermutation is intrinsic to the natural response to HBV infection and that it may contribute to the noncytolytic clearance of HBV [202, 207, 210]. However, there are no current in vitro evidence supporting a major role of A3 on the IFN-mediated HBV inactivation in the liver [215, 216].
In contrast to the moderate levels of APOBEC expression in the healthy liver, overexpression of these enzymes is observed in cirrhotic tissues, likely resultant from the high production of cytokines associated with the chronic inflammatory response against the infection. Consequent to this increase in APOBEC expression, high levels of HBV hypermutation have been observed, reaching 35% of the sequences in some cases [206].
A potential role of APOBEC3 has been suggested on the oncogenesis of the hepatocellular carcinoma (HCC). It has been shown that some APOBEC3 enzymes are able to generate truncation mutants of the HBx viral protein, leading to a selective advantage to preneoplastic and neoplastic hepatocytes [217]. Moreover, A3B was found to be overexpressed in HCC tissues. In HepG2 cells, A3B overexpression promoted their growth and led to an upregulation of the heat shock transcription factor-1 (HSF-1), which was also found to be upregulated in HCC [217]. HSF-1 is a regulator of the heat-shock response and is known to facilitate malignant transformation, cancer cell survival, and proliferation in model systems. Recently, it has been shown to coordinate a transcriptional program in malignancy, which differs from that induced by thermal stress. This program activates cancer-specific genes that support oncogenic events and was found to be active in breast, colon, and lung tumors [180].
3.4. Hepatitis C Virus (HCV)
HCV belongs to the Flaviviridae family and presents a positive sense, single-stranded RNA genome [218]. HCV is a causative agent of acute and chronic liver diseases. Chronic HCV infection, along with chronic HBV infection and its associated liver cirrhosis, constitutes major risk factors for HCC development [219]. It has been recently shown that A3G is able to inhibit HCV replication in vitro. However, viral hypermutated sequences have not been found, suggesting a role of deaminase-independent mechanisms of viral inhibition. Considering that A3G targets ssDNA, such lack of hypermutation is expected for HCV, which presents exclusively RNA as genomic material during all phases of its replication. Interestingly, it has also been shown that the presence of exogenous HIV-1 Vif led to an intracellular decrease of A3G and consequently to an increase of HCV replication, suggesting the involvement of Vif in the HIV-1/HCV coinfection [220].
As previously mentioned, the APOBEC enzymes are expressed at moderate levels in the normal liver. However, HCV infection is also associated to an increase in expression of those enzymes. In patients chronically infected by HCV, a significant increase in A3G expression was seen in hepatocytes and in lymphocytes [221]. Overexpression of APOBEC has also been observed in HCV/HBV coinfection [206].
APOBEC3 also appears to play an important role on treatment with exogenous interferon alpha (IFNα) in vivo. Jiménez-Sousa et al. [222] analyzed the profile of gene expression in HCV chronically infected patients after 12 weeks of treatment with IFNα/ribavirin (RBV), and A3A was among the IFN-induced genes that was upregulated in early responders but not in nonresponders. In another study, a significant increase in the expression of A3G/3F was observed in CD4 T-cell lymphocytes of HIV/HCV coinfected patients during treatment with pegIFN/RBV. In that study, APOBEC3 induction was correlated with the levels of HIV hypermutation [223].
In addition to the HCV restriction phenotype during the natural course of infection and during treatment, APOBEC3 enzymes have also been suggested as a putative target in anti-HCV drug development. Treatment of HCV-infected Huh7.5 cells with two stabilizing components of APOBEC3G, which increased its intracellular levels, inhibited HCV replication [220].
3.5. Human Papillomavirus (HPV)
HPV belongs to the Papillomaviridae family and presents a circular, double-stranded DNA genome. Infection by HPV is a necessary condition for the development of cervical cancer, but the evolution to invasive carcinoma only occurs in a fraction of the infected women [224].
Vartanian et al. [225] showed that both strands of HPV DNA are susceptible to APOBEC editing. In that study, nine HPV16-positive precancerous cervical and six HPV1a-positive plantar wart samples were analyzed for the presence of hypermutation. Of the samples, two HPV-16 and one HPV-1a presented edited sequences. In vitro A3A, A3C and A3H were shown to be able to hyperedit HPV DNA. The preferred in vitro dinucleotide context for these three A3 enzymes correlated with the editing contexts observed in vivo, suggesting that APOBEC3A, APOBEC3C, and APOBEC3H may be involved in edition of HPV in vivo.
3.6. Human Herpesviruses (HHV)
Human APOBEC enzymes have also been shown to restrict DNA genome viruses belonging to the Herpesviridae family [226, 227]. Herpesviruses are enveloped viruses harboring a double-stranded DNA genome. They are associated with a range of different diseases and are able to establish latent infection and persist in the infected host for life [228–231].
Herpes simplex virus type 1 (HSV-1) can cause from mild infections of mucous membranes, including herpes labialis and genital infections, to life threatening infections, such as HSV encephalitis [228, 229]. Suspène et al. [226] have identified the presence of HSV hypermutated genomes in four out of eight oral lesions. Overexpression of A3C in vitro led to a fourfold reduction in viral titers and to a 10-fold reduction in viral infectivity. Moreover, it has been shown that not only A3C but also AID, A3A, and A3G are able to edit the HSV-1 genome in vitro, although the last three had no significant impact on virus replication.
In addition to APOBEC3, APOBEC1 has also been shown to restrict HSV-1 replication in vitro in a significant fashion, in both deamination-dependent and -independent ways. Upregulation of A1 has been observed in rat brain tissues upon HSV-1 infection, suggesting that A1 induction during encephalitis can promote restriction to HSV-1 infection [227].
Epstein-Barr virus (EBV) can cause mucocutaneous manifestations in infectious mononucleosis and is also associated with other benign and malignant conditions, including plasmablastic lymphoma, oral hairy leukoplakia, posttransplant lymphoproliferative disorders, Burkitt's lymphoma, and Hodgkin's lymphoma [231]. In order to know whether EBV genomes were also susceptible to A3 editing, Suspène et al. [226] analyzed EBV from transformed peripheral blood mononuclear cell lines, which carry EBV in a latent form. Edited EBV DNA was found in four out of five EBV cell lines studied. A3C was found to be the most abundantly expressed A3 in these cell lines.
4. Polymorphisms in APOBEC3 Genes and Susceptibility to Viral Infections
Polymorphisms in the genes encoding APOBEC proteins have been associated with the modulation in the course of some human viral infections. A deletion of approximately 29.5 kb located between exon 5 of APOBEC3A and exon 8 of APOBEC3B, which results in the complete removal of the APOBEC3B coding region, has been found in different human populations. Its frequency varies among ethnic groups, being more prevalent in East Asians, Amerindians, and Oceanic populations (36.9%, 57.7%, and 92.9%, resp.) but rare among Africans and Europeans (0.9% and 6%, resp.) [232]. This polymorphism has been associated with increased risk for persistent HBV infection and for the development of HBV-associated hepatocellular carcinoma. Due to the A3B ability of restricting HBV, it has been suggested that this gene deletion can result in reduced viral clearance, culminating with persistent infection [97]. In other studies, the homozygosity status for the deletion was associated with mild liver fibrosis, but not with a chronic carrier status [95], and with faster progression of liver disease [98]. Homozygous individuals for this deletion have also been reported with an increased risk for HIV acquisition, for a higher viral set point and for progression to AIDS [94], yet no effect was found on susceptibility to HIV infection and AIDS in Japanese or Indian populations [96].
Several polymorphisms in the APOBEC3G gene have also been described. In a study with 3,073 HIV patients from 6 different cohorts, a variant at APOBEC3G exon 4, H186R, was frequently found in African Americans (37%) but rare in European American (<3%) and in Europeans (5%). This polymorphism was associated with CD4+ T-cell decline and with accelerated progression to AIDS-defining conditions in African Americans [99]. In another study with South African infected women, H186R was associated with higher HIV viral loads, and an extragenic mutation (rs35228531) was associated with decreased CD4+ T-cell levels [104]. In a study with HIV-1-infected children from Pediatric AIDS Clinical Trials Group (PACTG) protocols P152 and P300, the H186R and F119F variants were associated with altered HIV-1-related disease progression and central nervous system impairment [108]. However, no correlation was found between H186R and disease progression in a French cohort [100]. This polymorphism has not been found in Indians [103]. Another APOBEC3G gene polymorphism, C40693T, was associated with an increased risk of HIV infection in a cohort of 122 Caucasian individuals highly exposed to HIV-1 [102]. Finally, the APOBEC3G SNP-571 (rs5757463) was associated with lower CD4+ T-cell counts in homo- and heterozygosis in a group of HIV-1-infected individuals naive to drug therapy from Brazil [106].
APOBEC3H gene is also polymorphic, with seven haplotypes identified in human populations so far (named from I to VII). Of them, only three (II, V, and VII) seem to originate a stable A3H protein, and have a higher anti-HIV-1 activity [156, 157, 233]. APOBEC3H haplotype II has also been reported to potently restrict HTLV-1 [189]. Some of these haplotypes have a variable frequency. Haplotype 2 is present in high frequency in African populations [233]; haplotype V is more frequently detected in African-Americans, Caribbeans, and Chinese, while haplotype VII was rarer and found only in European Caucasians [157].
A comprehensive list of studies that described the association of particular APOBEC gene variants with viral infections of disease outcomes can be seen in Table 1. Despite several of these so-called candidate gene analysis studies identified definite associations, particularly in the context of HIV and HBV infections, they were not confirmed in genome-wide association studies (GWAS). The latter studies point mainly to a significant role of human leukocyte antigen (HLA) alleles [234–239], suggesting that the impact of APOBEC variations on viral diseases is less robust and only moderate [234].
Table 1.
Studies that investigated the association of APOBEC gene variants and the course of human viral infections.
| A3 family member | Series description | Main findings | Reference |
|---|---|---|---|
| A3B | 4,216 individuals from five HIV-1 natural history cohorts based in the United States of America | Homozygous deletion associated with increased risk for HIV-1 infection (P = 0.24), progression to AIDS (P = 0.03), and viral set point (P = 0.04) | [94] |
|
| |||
| A3B | 724 HBV carriers and 469 healthy control subjects |
APOBEC3B deletion homozygosity was associated with mild liver fibrosis (P = 0.0019) No significant association between deletion and chronic HBV infection |
[95] |
|
| |||
| A3B | 361 Japanese subjects: 95 HIV-1-infected patients (48 nonprogressors and 47 slow progressors) and 266 controls 453 Indian subjects: 251 HIV-1-infected patients and 202 controls |
No evidence of association between the APOBEC3B deletion and susceptibility to HIV infection and AIDS | [96] |
|
| |||
| A3B | 1,124 individuals with HCC, 510 individuals with persistent HBV infection, and 826 healthy controls. All subjects were of Han Chinese ethnicity | Higher frequency of the APOBEC3B deletion allele in persistent HBV carriers (P = 0.0015) and HCC patients (P = 1.28 × 10−11) compared to controls Presence of at least one deletion allele was associated with an increased risk for persistent HBV infection (P = 0.0272) and HCC development (P = 1.28 × 10−11) |
[97] |
|
| |||
| A3B | 179 HBV chronic carriers and 216 healthy control subjects from the Moroccan population | No significant difference in the frequency of deleted APOBEC3B alleles between patients with chronic hepatitis B and control subjects Subjects carrying the Del/Del genotype displayed a trend for increased susceptibility to HBV infection compared to the wild type genotype (P = 0.07) Carriers of the APOBEC3B deletion had significantly lower viral loads than patients with the wild type genotype (P = 0.0023) |
[98] |
|
| |||
| A3G | 3,073 participants enrolled in six HIV/AIDS prospective cohorts: 1,481 European Americans, 949 African Americans from five US-based cohorts, and 643 patients enrolled in the Swiss HIV cohort | For African Americans, the variant allele 186R was strongly associated with a decline of CD4+ T cells (P = 0.009) | [99] |
|
| |||
| A3G | 773 white French individuals: 327 HIV-1+ (245 slow progressors; 82 rapid progressors) and 446 healthy control subjects of similar ethnic origin | 29 polymorphisms with allele frequencies >1% were identified No significant associations were found between the polymorphisms or haplotypes and disease progression |
[100] |
|
| |||
| A3G | 136 adult HIV-infected patients from the Western Australian HIV cohort | 22 single nucleotide polymorphisms were identified No significant association of these APOBEC3G genetic variants and the presence of HIV-1 hypermutation was found (although an intronic allele 6892C was marginally associated with HIV-1 hypermutation) |
[101] |
|
| |||
| A3G | 122 Caucasian individuals exposed to HIV enrolled in prospective cohort studies in Montreal | The C40693T variant was significantly associated with an increased risk of infection (P = 0.03) | [102] |
|
| |||
| A3G | 560 North Indians: 50 HIV-1 exposed seronegative individuals, 190 HIV-1+ patients, and 320 healthy controls | No H186R polymorphism of APOBEC3G was found among North Indians | [103] |
|
| |||
| A3G | 250 South African women at high risk for HIV-1 subtype C infection |
The H186R mutation and a 3′ extragenic mutation (rs35228531) were associated with high HIV viral loads (P = 0.0097 and P < 0.0001) and decreased CD4+ T-cell counts (P = 0.0081 and P < 0.0001) | [104] |
|
| |||
| A3G | 534 children perinatally exposed to HIV-1 (109 exposed uninfected and 425 HIV-1-infected), from a pediatric cohort of white-Hispanic ethnicity from Argentina | HIV-1 perinatal transmission and progression to AIDS were not affected by APOBEC3G H186R or APOBEC3G C40693T APOBEC3G C40693T was correlated with substitutions in Vif motifs involved in the interaction with APOBEC3G (P = 0.004) |
[105] |
|
| |||
| A3G | 400 HIV-1-infected individuals naive to drug therapy from the Brazilian population | Seven loci were analyzed: SNP −571 (rs5757463); −199 (rs34550797); −90 (rs5750743); 119 (rs5757465); 186 (rs8177832); 197 (rs3736685); 199 (rs2294367) For the SNP −571, heterozygous (C/G) and homozygous (G/G) individuals had lower CD4+ T-cell counts compared to homozygous (C/C) individuals (P = 0.0076) |
[106] |
|
| |||
| A3G | 93 perinatally infected children with white-Hispanic ethnicity, from an Argentinian pediatric cohort | The APOBEC3G H186R and APOBEC3G C40693T variants were not associated with different levels of HIV-1 editing | [107] |
|
| |||
| A3G | 1,049 HIV-1-infected children from the Pediatric AIDS Clinical Trials Group (PACTG) protocols P152 and P300 (60% non-Hispanic black, 26% Hispanic, 13% non-Hispanic white, and 1% other or unknown race/ethnicity) |
APOBEC3G H186R homozygous G/G genotype was associated with faster HIV-1 disease progression (P = 0.01) and central nervous system (CNS) impairment (P = 0.02) APOBEC3G F119F-C allele was associated with protection against disease progression and CNS impairment in both additive and dominant models (P = 0.002 and P = 0.001, resp.) and CNS impairment (P = 0.02 and P = 0.007, resp.) |
[108] |
|
| |||
| A3G | 179 HBV chronic carriers and 216 healthy control subjects from the Moroccan population | No significant difference in the frequencies of APOBEC3G H186R genotype between patients with chronic hepatitis B and control subjects | [98] |
|
| |||
| A3H | 70 Italian HIV-exposed seronegative individuals and their HIV-1-infected sexual partners | The APOBEC3H haplotype I was found in a higher frequency in the exposed seronegative compared to the HIV+ individuals (P = 0.0056), suggesting a protection from sexually transmitted HIV-1 infection | [109] |
|
| |||
| A3H | 96 recently HIV-1-infected treatment-naïve adults | 68 SNPs were analyzed Homozygous carriers of an APOBEC3H risk haplotype (A3Hrh) had lower GA→AA (A3F) sequence editing on proviral HIV-1 vif sequence (P = 0.01) and lower HIV-1 RNA levels (P = 0.015) |
[110] |
5. Concluding Remarks
Innate immunity mechanisms are the first line against invading viruses and are promising targets for preventing viral infections. Some infections are life-long once established, for example, by HIV and HSV-1, and therefore innate immunity is pivotal to avoiding the unfavorable consequences of infection of the host. The APOBEC family of cytidine deaminases plays an important role within innate immunity by deteriorating the genetic information of many viruses through hypermutation-dependent and -independent mechanisms. Polymorphisms in APOBEC genes that render differences in their expression and enzymatic activity also affect viral infection outcomes, and yet currently these effects appear modest and are not evidenced by GWAS. Viruses, on their hand, evolved molecular mechanisms to counteract APOBEC effects. Diminished APOBEC activity and sublethal levels of hypermutation may favor virus evolution, by generating viable variants carrying immune escape or drug resistance mutations. Conversely, the unfavorable consequences of APOBEC upregulation, particularly its recently described carcinogenic and genotoxic potential, are important caveats that will require further assessment and will pose a challenge to strategies aiming at increasing APOBEC expression to counteract viral infections. These issues will certainly warrant continuing investigation on the role and effects of cytidine deamination in infectious diseases and cancer.
Acknowledgments
V. C. Vieira is a recipient of a Ph.D. scholarship from the Brazilian Ministry of Health. This work is part of her requirements for obtaining a Ph.D. degree at the Graduate Program in Oncology of the Brazilian Cancer Institute (INCA). M. A. Soares is sponsored by grants of the Brazilian Ministry of Health, Brazilian Research Council (CNPq) and the Rio de Janeiro State Science Foundation (FAPERJ).
References
- 1.Iwasaki A. A virological view of innate immune recognition. Annual Review of Microbiology. 2012;66:177–196. doi: 10.1146/annurev-micro-092611-150203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Takeuchi O, Akira S. Innate immunity to virus infection. Immunological Reviews. 2009;227(1):75–86. doi: 10.1111/j.1600-065X.2008.00737.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Thompson MR, Kaminski JJ, Kurt-Jones EA, Fitzgerald KA. Pattern recognition receptors and the innate immune response to viral infection. Viruses. 2011;3(6):920–940. doi: 10.3390/v3060920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stetson DB, Medzhitov R. Type I interferons in host defense. Immunity. 2006;25(3):373–381. doi: 10.1016/j.immuni.2006.08.007. [DOI] [PubMed] [Google Scholar]
- 5.Conticello SG, Thomas CJF, Petersen-Mahrt SK, Neuberger MS. Evolution of the AID/APOBEC family of polynucleotide (deoxy)cytidine deaminases. Molecular Biology and Evolution. 2005;22(2):367–377. doi: 10.1093/molbev/msi026. [DOI] [PubMed] [Google Scholar]
- 6.Muto T, Muramatsu M, Taniwaki M, Kinoshita K, Honjo T. Isolation, tissue distribution, and chromosomal localization of the human activation-induced cytidine deaminase (AID) gene. Genomics. 2000;68(1):85–88. doi: 10.1006/geno.2000.6268. [DOI] [PubMed] [Google Scholar]
- 7.Espinosa R, III, Funahashi T, Hadjiagapiou C, le Beau MM, Davidson NO. Assignment of the gene encoding the human apolipoprotein B mRNA editing enzyme (APOBEC1) to chromosome 12p13.1. Genomics. 1994;24(2):414–415. doi: 10.1006/geno.1994.1645. [DOI] [PubMed] [Google Scholar]
- 8.Liao W, Hong S, Chann BH, Rudolph FB, Clark S, Chan L. APOBEC-2, a cardiac- and skeletal muscle-specific member of the cytidine deaminase supergene family. Biochemical and Biophysical Research Communications. 1999;260:398–404. doi: 10.1006/bbrc.1999.0925. [DOI] [PubMed] [Google Scholar]
- 9.Jarmuz A, Chester A, Bayliss J, et al. An anthropoid-specific locus of orphan C to U RNA-editing enzymes on chromosome 22. Genomics. 2002;79(3):285–296. doi: 10.1006/geno.2002.6718. [DOI] [PubMed] [Google Scholar]
- 10.Rogozin IB, Basu MK, Jordan IK, Pavlov YI, Koonin EV. APOBEC4, a new member of the AID/APOBEC family of polynucleotide (deoxy)cytidine deaminases predicted by computational analysis. Cell Cycle. 2005;4(9):1281–1285. doi: 10.4161/cc.4.9.1994. [DOI] [PubMed] [Google Scholar]
- 11.Wedekind JE, Dance GSC, Sowden MP, Smith HC. Messenger RNA editing in mammals: new members of the APOBEC family seeking roles in the family business. Trends in Genetics. 2003;19(4):207–216. doi: 10.1016/S0168-9525(03)00054-4. [DOI] [PubMed] [Google Scholar]
- 12.Teng B, Burant CF, Davidson NO. Molecular cloning of an apolipoprotein B messenger RNA editing protein. Science. 1993;260(5115):1816–1818. doi: 10.1126/science.8511591. [DOI] [PubMed] [Google Scholar]
- 13.Harris RS, Petersen-Mahrt SK, Neuberger MS. RNA editing enzyme APOBEC1 and some of its homologs can act as DNA mutators. Molecular Cell. 2002;10(5):1247–1253. doi: 10.1016/s1097-2765(02)00742-6. [DOI] [PubMed] [Google Scholar]
- 14.Yang Y, Yang Y, Smith HC. Multiple protein domains determine the cell type-specific nuclear distribution of the catalytic subunit required for apolipoprotein B mRNA editing. Proceedings of the National Academy of Sciences of the United States of America. 1997;94(24):13075–13080. doi: 10.1073/pnas.94.24.13075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lau PP, Xiong W, Zhu H-J, Chen S-H, Chan L. Apolipoprotein B mRNA editing is an intranuclear event that occurs posttranscriptionally coincident with splicing and polyadenylation. The Journal of Biological Chemistry. 1991;266(30):20550–20554. [PubMed] [Google Scholar]
- 16.Sowden MP, Ballatori N, de Mesy Jensen KL, Hamilton Reed L, Smith HC. The editosome for cytidine to uridine mRNA editing has a native complexity of 27S: identification of intracellular domains containing active and inactive editing factors. Journal of Cell Science. 2002;115(5):1027–1039. doi: 10.1242/jcs.115.5.1027. [DOI] [PubMed] [Google Scholar]
- 17.Anant S, Davidson NO. Molecular mechanisms of apolipoprotein B mRNA editing. Current Opinion in Lipidology. 2001;12(2):159–165. doi: 10.1097/00041433-200104000-00009. [DOI] [PubMed] [Google Scholar]
- 18.Anant S, Davidson NO. An AU-rich sequence element (UUUN[A/U]U) downstream of the edited C in apolipoprotein B mRNA is a high-affinity binding site for Apobec-1: binding of Apobec-1 to this motif in the 3’ untranslated region of c-myc increases mRNA stability. Molecular and Cellular Biology. 2000;20(6):1982–1992. doi: 10.1128/mcb.20.6.1982-1992.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Rosenberg BR, Hamilton CE, Mwangi MM, Dewell S, Papavasiliou FN. Transcriptome-wide sequencing reveals numerous APOBEC1 mRNA-editing targets in transcript 3’ UTRs. Nature Structural & Molecular Biology. 2011;18(2):230–238. doi: 10.1038/nsmb.1975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dickerson SK, Market E, Besmer E, Papavasiliou FN. AID mediates hypermutation by deaminating single stranded DNA. The Journal of Experimental Medicine. 2003;197(10):1291–1296. doi: 10.1084/jem.20030481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Bransteitter R, Pham P, Scharfft MD, Goodman MF. Activation-induced cytidine deaminase deaminates deoxycytidine on single-stranded DNA but requires the action of RNase. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(7):4102–4107. doi: 10.1073/pnas.0730835100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rada C, Jarvis JM, Milstein C. AID-GFP chemirec protein increases hypermutation og Ig genes with no evidence of nuclear localization. Proceedings of the National Academy of Sciences of the United States of America. 2002;99(10):7003–7008. doi: 10.1073/pnas.092160999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Brar SS, Watson M, Diaz M. Activation-induced cytosine deaminase (AID) is actively exported out of the nucleus but retained by the induction of DNA breaks. The Journal of Biological Chemistry. 2004;279(25):26395–26401. doi: 10.1074/jbc.M403503200. [DOI] [PubMed] [Google Scholar]
- 24.Ito S, Nagaoka H, Shinkura R, et al. Activation-induced cytidine deaminase shuttles between nucleus and cytoplasm like apolipoprotein B mRNA editing catalytic polypeptide 1. Proceedings of the National Academy of Sciences of the United States of America. 2004;101(7):1975–1980. doi: 10.1073/pnas.0307335101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McBride KM, Barreto V, Ramiro AR, Stavropoulos P, Nussenzweig MC. Somatic hypermutation is limited by CRM1-dependent nuclear export of activation-induced deaminase. The Journal of Experimental Medicine. 2004;199(9):1235–1244. doi: 10.1084/jem.20040373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Muramatsu M, Kinoshita K, Fagarasan S, Yamada S, Shinkai Y, Honjo T. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 2000;102(5):553–563. doi: 10.1016/s0092-8674(00)00078-7. [DOI] [PubMed] [Google Scholar]
- 27.Di Noia JM, Neuberger MS. Molecular mechanisms of antibody somatic hypermutation. Annual Review of Biochemistry. 2007;76:1–22. doi: 10.1146/annurev.biochem.76.061705.090740. [DOI] [PubMed] [Google Scholar]
- 28.Muramatsu M, Sankaranand VS, Anant S, et al. Specific expression of activation-induced cytidine deaminase (AID), a novel member of the RNA-editing deaminase family in germinal center B cells. The Journal of Biological Chemistry. 1999;274(26):18470–18476. doi: 10.1074/jbc.274.26.18470. [DOI] [PubMed] [Google Scholar]
- 29.Popp C, Dean W, Feng S, et al. Genome-wide erasure of DNA methylation in mouse primordial germ cells is affected by AID deficiency. Nature. 2010;463(7284):1101–1105. doi: 10.1038/nature08829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bhutani N, Brady JJ, Damian M, Sacco A, Corbel SY, Blau HM. Reprogramming towards pluripotency requires AID-dependent DNA demethylation. Nature. 2010;463(7284):1042–1047. doi: 10.1038/nature08752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Guo JU, Su Y, Zhong C, Ming G-L, Song H. Hydroxylation of 5-methylcytosine by TET1 promotes active DNA demethylation in the adult brain. Cell. 2011;145(3):423–434. doi: 10.1016/j.cell.2011.03.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Sato Y, Probst HC, Tatsumi R, Ikeuchi Y, Neuberger MS, Rada C. Deficiency in APOBEC2 leads to a shift in muscle fiber type, diminished body mass, and myopathy. The Journal of Biological Chemistry. 2010;285(10):7111–7118. doi: 10.1074/jbc.M109.052977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Wedekind JE, Gillilan R, Janda A, et al. Nanostructures of APOBEC3G support a hierarchical assembly model of high molecular mass ribonucleoprotein particles from dimeric subunits. The Journal of Biological Chemistry. 2006;281(50):38122–38126. doi: 10.1074/jbc.C600253200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Navarro F, Bollman B, Chen H, et al. Complementary function of the two catalytic domains of APOBEC3G. Virology. 2005;333(2):374–386. doi: 10.1016/j.virol.2005.01.011. [DOI] [PubMed] [Google Scholar]
- 35.Beale RCL, Petersen-Mahrt SK, Watt IN, Harris RS, Rada C, Neuberger MS. Comparison of the differential context-dependence of DNA deamination by APOBEC enzymes: correlation with mutation spectra in vivo . Journal of Molecular Biology. 2004;337(3):585–596. doi: 10.1016/j.jmb.2004.01.046. [DOI] [PubMed] [Google Scholar]
- 36.Armitage AE, Katzourakis A, de Oliveira T, et al. Conserved footprints of APOBEC3G on hypermutated human immunodeficiency virus type 1 and human endogenous retrovirus HERV-K(HML2) sequences. Journal of Virology. 2008;82(17):8743–8761. doi: 10.1128/JVI.00584-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Bishop KN, Holmes RK, Sheehy AM, Davidson NO, Cho S-J, Malim MH. Cytidine deamination of retroviral DNA by diverse APOBEC proteins. Current Biology. 2004;14(15):1392–1396. doi: 10.1016/j.cub.2004.06.057. [DOI] [PubMed] [Google Scholar]
- 38.Harris RS, Bishop KN, Sheehy AM, et al. DNA deamination mediates innate immunity to retroviral infection. Cell. 2003;113(6):803–809. doi: 10.1016/s0092-8674(03)00423-9. [DOI] [PubMed] [Google Scholar]
- 39.Bogerd HP, Wiegand HL, Doehle BP, Lueders KK, Cullen BR. APOBEC3A and APOBEC3B are potent inhibitors of LTR-retrotransposon function in human cells. Nucleic Acids Research. 2006;34(1):89–95. doi: 10.1093/nar/gkj416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kinomoto M, Kanno T, Shimura M, et al. All APOBEC3 family proteins differentially inhibit LINE-1 retrotransposition. Nucleic Acids Research. 2007;35(9):2955–2964. doi: 10.1093/nar/gkm181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Delebecque F, Suspène R, Calattini S, et al. Restriction of foamy viruses by APOBEC cytidine deaminases. Journal of Virology. 2006;80(2):605–614. doi: 10.1128/JVI.80.2.605-614.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yu Q, Chen D, König R, Mariani R, Unutmaz D, Landau NR. APOBEC3B and APOBEC3C are potent inhibitors of simian immunodeficiency virus replication. The Journal of Biological Chemistry. 2004;279(51):53379–53386. doi: 10.1074/jbc.M408802200. [DOI] [PubMed] [Google Scholar]
- 43.Anderson JL, Hope TJ. APOBEC3G restricts early HIV-1 replication in the cytoplasm of target cells. Virology. 2008;375(1):1–12. doi: 10.1016/j.virol.2008.01.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bishop KN, Holmes RK, Malim MH. Antiviral potency of APOBEC proteins does not correlate with cytidine deamination. Journal of Virology. 2006;80(17):8450–8458. doi: 10.1128/JVI.00839-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Bishop KN, Verma M, Kim E-Y, Wolinsky SM, Malim MH. APOBEC3G inhibits elongation of HIV-1 reverse transcripts. PLoS Pathogens. 2008;4(12) doi: 10.1371/journal.ppat.1000231.e1000231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Stenglein MD, Burns MB, Li M, Lengyel J, Harris RS. APOBEC3 proteins mediate the clearance of foreign DNA from human cells. Nature Structural & Molecular Biology. 2010;17(2):222–229. doi: 10.1038/nsmb.1744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Suspène R, Aynaud M-M, Guétard D, et al. Somatic hypermutation of human mitochondrial and nuclear DNA by APOBEC3 cytidine deaminases, a pathway for DNA catabolism. Proceedings of the National Academy of Sciences of the United States of America. 2011;108(12):4858–4863. doi: 10.1073/pnas.1009687108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Burns MB, Lackey L, Carpenter MA, et al. APOBEC3B is an enzymatic source of mutation in breast cancer. Nature. 2013;494:366–370. doi: 10.1038/nature11881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Bennett RP, Presnyak V, Wedekind JE, Smith HC. Nuclear exclusion of the HIV-1 host defense factor APOBEC3G requires a novel cytoplasmic retention signal and is not dependent on RNA binding. The Journal of Biological Chemistry. 2008;283(12):7320–7327. doi: 10.1074/jbc.M708567200. [DOI] [PubMed] [Google Scholar]
- 50.Lackey L, Demorest ZL, Land AM, Hultquist JF, Brown WL, Harris RS. APOBEC3B and AID have similar nuclear import mechanisms. Journal of Molecular Biology. 2012;419(5):301–314. doi: 10.1016/j.jmb.2012.03.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Li MMH, Emerman M. Polymorphism in human APOBEC3H affects a phenotype dominant for subcellular localization and antiviral activity. Journal of Virology. 2011;85(16):8197–8207. doi: 10.1128/JVI.00624-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Land AM, Law EK, Carpenter MA, Lackey L, Brown WL, Harris RS. Endogenous APOBEC3A is cytoplasmic and non-genotoxic. The Journal of Biological Chemistry. 2013 doi: 10.1074/jbc.M113.458661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chiu Y-L, Soros VB, Kreisberg JF, Stopak K, Yonemoto W, Greene WC. Cellular APOBEC3G restricts HIV-1 infection in resting CD4+ T cells. Nature. 2005;435(7038):108–114. doi: 10.1038/nature03493. [DOI] [PubMed] [Google Scholar]
- 54.Chiu Y-L, Greene WC. The APOBEC3 cytidine deaminases: an innate defensive network opposing exogenous retroviruses and endogenous retroelements. Annual Review of Immunology. 2008;26:317–353. doi: 10.1146/annurev.immunol.26.021607.090350. [DOI] [PubMed] [Google Scholar]
- 55.Smith HC, Bennett RP, Kizilyer A, McDougall WM, Prohaska KM. Functions and regulation of the APOBEC family of proteins. Seminars in Cell and Developmental Biology. 2012;23(3):258–268. doi: 10.1016/j.semcdb.2011.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Wang X, Dolan PT, Dang Y, Zheng Y-H. Biochemical differentiation of APOBEC3F and APOBEC3G proteins associated with HIV-1 life cycle. The Journal of Biological Chemistry. 2007;282(3):1585–1594. doi: 10.1074/jbc.M610150200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Tan L, Sarkis PTN, Wang T, Tian C, Yu X-F. Sole copy of Z2-type human cytidine deaminase APOBEC3H has inhibitory activity against retrotransposons and HIV-1. The FASEB Journal. 2009;23(1):279–287. doi: 10.1096/fj.07-088781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Niewiadomska AM, Tian C, Tan L, Wang T, Sarkis PTN, Yu X-F. Differential inhibition of long interspersed element 1 by APOBEC3 does not correlate with high-molecular-mass-complex formation or P-body association. Journal of Virology. 2007;81(17):9577–9583. doi: 10.1128/JVI.02800-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kreisberg JF, Yonemoto W, Greene WC. Endogenous factors enhance HIV infection of tissue naive CD4 T cells by stimulating high molecular mass APOBEC3G complex formation. The Journal of Experimental Medicine. 2006;203(4):865–870. doi: 10.1084/jem.20051856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Stopak KS, Chiu Y-L, Kropp J, Grant RM, Greene WC. Distinct patterns of cytokine regulation of APOBEC3G expression and activity in primary lymphocytes, macrophages, and dendritic cells. The Journal of Biological Chemistry. 2007;282(6):3539–3546. doi: 10.1074/jbc.M610138200. [DOI] [PubMed] [Google Scholar]
- 61.Ellery PJ, Tippett E, Chiu Y-L, et al. The CD16+ monocyte subset is more permissive to infection and preferentially harbors HIV-1 in vivo . Journal of Immunology. 2007;178(10):6581–6589. doi: 10.4049/jimmunol.178.10.6581. [DOI] [PubMed] [Google Scholar]
- 62.Kamata M, Nagaoka Y, Chen ISY. Reassessing the role of APOBEC3G in human immunodeficiency virus type 1 infection of quiescent CD4+ T-cells. PLoS Pathogens. 2009;5(3) doi: 10.1371/journal.ppat.1000342.e1000342 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Santoni de Sio FR, Trono D. APOBEC3G-depleted resting CD4+ T cells remain refractory to HIV1 infection. PLoS ONE. 2009;4(8, article e6571) doi: 10.1371/journal.pone.0006571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Soros VB, Yonemoto W, Greene WC. Newly synthesized APOBEC3G is incorporated into HIV virions, inhibited by HIV RNA, and subsequently activated by RNase H. PLoS Pathogens. 2007;3(2, article e15) doi: 10.1371/journal.ppat.0030015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Khan MA, Goila-Gaur R, Kao S, Miyagi E, Walker RC, Jr., Strebel K. Encapsidation of APOBEC3G into HIV-1 virions involves lipid raft association and does not correlate with APOBEC3G oligomerization. Retrovirology. 2009;6, article 1742 doi: 10.1186/1742-4690-6-99. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Ma J, Li X, Xu J, et al. The cellular source for APOBEC3G’s incorporation into HIV-1. Retrovirology. 2011;8(article 2) doi: 10.1186/1742-4690-8-2. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 67.Chiu Y-L, Witkowska HE, Hall SC, et al. High-molecular-mass APOBEC3G complexes restrict Alu retrotransposition. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(42):15588–15593. doi: 10.1073/pnas.0604524103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Gallois-Montbrun S, Kramer B, Swanson CM, et al. Antiviral protein APOBEC3G localizes to ribonucleoprotein complexes found in P bodies and stress granules. Journal of Virology. 2007;81(5):2165–2178. doi: 10.1128/JVI.02287-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Gallois-Montbrun S, Holmes RK, Swanson CM, et al. Comparison of cellular ribonucleoprotein complexes associated with the APOBEC3F and APOBEC3G antiviral proteins. Journal of Virology. 2008;82(11):5636–5642. doi: 10.1128/JVI.00287-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Kozak SL, Marin M, Rose KM, Bystrom C, Kabat D. The anti-HIV-1 editing enzyme APOBEC3G binds HIV-1 RNA and messenger RNAs that shuttle between polysomes and stress granules. The Journal of Biological Chemistry. 2006;281(39):29105–29119. doi: 10.1074/jbc.M601901200. [DOI] [PubMed] [Google Scholar]
- 71.Wichroski MJ, Robb GB, Rana TM. Human retroviral host restriction factors APOBEC3G and APOBEC3F localize to mRNA processing bodies. PLoS Pathogens. 2006;2(5, article e41) doi: 10.1371/journal.ppat.0020041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Phalora PK, Sherer NM, Wolinsky SM, Swanson CM, Malim MH. HIV-1 replication and APOBEC3 antiviral activity are not regulated by P bodies. Journal of Virology. 2012;86(21):11712–11724. doi: 10.1128/JVI.00595-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Lada AG, Frahm Krick C, Kozmin SG, et al. Mutator effects and mutation signatures of editing deaminases produced in bacteria and yeast. Biochemistry. 2011;76(1):131–146. doi: 10.1134/s0006297911010135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Conticello SG. Creative deaminases, self-inflicted damage, and genome evolution. Annals of the New York Academy of Sciences. 2012;1267:79–85. doi: 10.1111/j.1749-6632.2012.06614.x. [DOI] [PubMed] [Google Scholar]
- 75.Münk C, Jensen BEO, Zielonka J, Häussinger D, Kamp C. Running loose or getting lost: how HIV-1 counters and capitalizes on APOBEC3-induced mutagenesis through its Vif protein. Viruses. 2012;4:3132–3161. doi: 10.3390/v4113132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Severi F, Chicca A, Conticello SG. Analysis of reptilian APOBEC1 suggests that RNA Editing may not be its ancestral function. Molecular Biology and Evolution. 2011;28(3):1125–1129. doi: 10.1093/molbev/msq338. [DOI] [PubMed] [Google Scholar]
- 77.Harris RS, Liddament MT. Retroviral restriction by APOBEC proteins. Nature Reviews Immunology. 2004;4(11):868–877. doi: 10.1038/nri1489. [DOI] [PubMed] [Google Scholar]
- 78.Münk C, Willemsen A, Bravo I. An ancient history of gene duplications, fusions and losses in the evolution of APOBEC3 mutators in mammals. BMC Evolutionary Biology. 2012;12(article 71) doi: 10.1186/1471-2148-12-71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.LaRue RS, Jónsson SR, Silverstein KAT, et al. The artiodactyl APOBEC3 innate immune repertoire shows evidence for a multi-functional domain organization that existed in the ancestor of placental mammals. BMC Molecular Biology. 2008;9(104):1–20. doi: 10.1186/1471-2199-9-104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Münk C, Beck T, Zielonka J, et al. Functions, structure, and read-through alternative splicing of feline APOBEC3 genes. Genome Biology. 2008;9(3, article R48) doi: 10.1186/gb-2008-9-3-r48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Schumann GG. APOBEC3 proteins: major players in intracellular defence against LINE-1-mediated retrotransposition. Biochemical Society Transactions. 2007;35(3):637–642. doi: 10.1042/BST0350637. [DOI] [PubMed] [Google Scholar]
- 82.Koito A, Ikeda T. Intrinsic immunity against retrotransposons by APOBEC cytidine deaminases. Frontiers in Microbiology. 2013;4:1–9. doi: 10.3389/fmicb.2013.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Bogerd HP, Wiegand HL, Hulme AE, et al. Cellular inhibitors of long interspersed element 1 and Alu retrotransposition. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(23):8780–8785. doi: 10.1073/pnas.0603313103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Hulme AE, Bogerd HP, Cullen BR, Moran JV. Selective inhibition of Alu retrotransposition by APOBEC3G. Gene. 2007;390(1-2):199–205. doi: 10.1016/j.gene.2006.08.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Esnault C, Heidmann O, Delebecque F, et al. APOBEC3G cytidine deaminase inhibits retrotransposition of endogenous retroviruses. Nature. 2005;433(7024):430–433. doi: 10.1038/nature03238. [DOI] [PubMed] [Google Scholar]
- 86.Esnault C, Millet J, Schwartz O, Heidmann T. Dual inhibitory effects of APOBEC family proteins on retrotransposition of mammalian endogenous retroviruses. Nucleic Acids Research. 2006;34(5):1522–1531. doi: 10.1093/nar/gkl054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Schumacher AJ, Nissley DV, Harris RS. APOBEC3G hypermutates genomic DNA and inhibits Ty1 retrotransposition in yeast. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(28):9854–9859. doi: 10.1073/pnas.0501694102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Macduff DA, Demorest ZL, Harris RS. AID can restrict L1 retrotransposition suggesting a dual role in innate and adaptive immunity. Nucleic Acids Research. 2009;37(6):1854–1867. doi: 10.1093/nar/gkp030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Ikeda T, Abd El Galil KH, Tokunaga K, et al. Intrinsic restriction activity by apolipoprotein B mRNA editing enzyme APOBEC1 against the mobility of autonomous retrotransposons. Nucleic Acids Research. 2011;39(13):5538–5554. doi: 10.1093/nar/gkr124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Metzner M, Jäck H-M, Wabl M. LINE-1 retroelements complexed and inhibited by activation induced cytidine deaminase. PloS One. 2012;7(11, article e49358) doi: 10.1371/journal.pone.0049358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Bishop KN, Holmes RK, Sheehy AM, Malim MH. APOBEC-mediated editing of viral RNA. Science. 2004;305(5684, article 645) doi: 10.1126/science.1100658. [DOI] [PubMed] [Google Scholar]
- 92.Ikeda T, Ohsugi T, Kimura T, et al. The antiretroviral potency of APOBEC1 deaminase from small animal species. Nucleic Acids Research. 2008;36(21):6859–6871. doi: 10.1093/nar/gkn802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Petit V, Guétard D, Renard M, et al. Murine APOBEC1 is a powerful mutator of retroviral and cellular RNA in vitro and in vivo . Journal of Molecular Biology. 2009;385(1):65–78. doi: 10.1016/j.jmb.2008.10.043. [DOI] [PubMed] [Google Scholar]
- 94.An P, Johnson R, Phair J, et al. APOBEC3B deletion and risk of HIV-1 acquisition. The Journal of Infectious Diseases. 2009;200(7):1054–1058. doi: 10.1086/605644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Abe H, Ochi H, Maekawa T, et al. Effects of structural variations of APOBEC3A and APOBEC3B genes in chronic hepatitis B virus infection. Hepatology Research. 2009;39(12):1159–1168. doi: 10.1111/j.1872-034X.2009.00566.x. [DOI] [PubMed] [Google Scholar]
- 96.Itaya S, Nakajima T, Kaur G, et al. No evidence of an association between the APOBEC3B deletion polymorphism and susceptibility to HIV infection and AIDS in Japanese and indian populations. The Journal of Infectious Diseases. 2010;202(5):815–816. doi: 10.1086/655227. [DOI] [PubMed] [Google Scholar]
- 97.Zhang T, Cai J, Chang J, et al. Evidence of associations of APOBEC3B gene deletion with susceptibility to persistent HBV infection and hepatocellular carcinoma. Human Molecular Genetics. 2012;22(6):1262–1269. doi: 10.1093/hmg/dds513. [DOI] [PubMed] [Google Scholar]
- 98.Ezzikouri S, Kitab B, Rebbani K, et al. Polymorphic APOBEC3 modulates chronic hepatitis B in Moroccan population. Journal of Viral Hepatitis. 2012:1–9. doi: 10.1111/jvh.12042. [DOI] [PubMed] [Google Scholar]
- 99.An P, Bleiber G, Duggal P, et al. APOBEC3G genetic variants and their influence on the progression to AIDS. Journal of Virology. 2004;78(20):11070–11076. doi: 10.1128/JVI.78.20.11070-11076.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Do H, Vasilescu A, Diop G, et al. Exhaustive genotyping of the CEM15 (APOBEC3G) gene and absence of association with AIDS progression in a French cohort. The Journal of Infectious Diseases. 2005;191(2):159–163. doi: 10.1086/426826. [DOI] [PubMed] [Google Scholar]
- 101.Pace C, Keller J, Nolan D, et al. Population level analysis of human immunodeficiency virus type 1 hypermutation and its relationship with APOBEC3G and vif genetic variation. Journal of Virology. 2006;80(18):9259–9269. doi: 10.1128/JVI.00888-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Valcke HS, Bernard NF, Bruneau J, Alary M, Tsoukas CM, Roger M. APOBEC3G genetic variants and their association with risk of HIV infection in highly exposed Caucasians. AIDS. 2006;20(15):1984–1986. doi: 10.1097/01.aids.0000247124.35129.e1. [DOI] [PubMed] [Google Scholar]
- 103.Rathore A, Chatterjee A, Yamamoto N, Dhole TN. Absence of H186R polymorphism in exon 4 of the APOBEC3G gene among north Indian individuals. Genetic Testing. 2008;12(3):453–456. doi: 10.1089/gte.2008.0017. [DOI] [PubMed] [Google Scholar]
- 104.Reddy K, Winkler CA, Werner L, Mlisana K, Abdool Karim SS, Ndung’U T. Apobec3g expression is dysregulated in primary hiv-1 infection and polymorphic variants influence cd4+ t-cell counts and plasma viral load. AIDS. 2010;24(2):195–204. doi: 10.1097/QAD.0b013e3283353bba. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.de Maio FA, Rocco CA, Aulicino PC, Bologna R, Mangano A, Sen L. Effect of HIV-1 Vif variability on progression to pediatric AIDS and its association with APOBEC3G and CUL5 polymorphisms. Infection, Genetics and Evolution. 2011;11(6):1256–1262. doi: 10.1016/j.meegid.2011.04.020. [DOI] [PubMed] [Google Scholar]
- 106.Bizinoto MC, Leal É, Diaz RS, Janini LM. Loci polymorphisms of the APOBEC3G gene in HIV type 1-infected Brazilians. AIDS Research and Human Retroviruses. 2011;27(2):137–141. doi: 10.1089/aid.2010.0146. [DOI] [PubMed] [Google Scholar]
- 107.de Maio FA, Rocco CA, Aulicino PC, Bologna R, Mangano A, Sen L. APOBEC3-mediated editing in HIV type 1 from pediatric patients and its association with APOBEC3G/CUL5 polymorphisms and Vif variability. AIDS Research and Human Retroviruses. 2012;28(6):619–627. doi: 10.1089/AID.2011.0291. [DOI] [PubMed] [Google Scholar]
- 108.Singh KK, Wang Y, Gray KP, et al. Genetic variants in the host restriction factor APOBEC3G are associated with HIV-1-related disease progression and central nervous system impairment in children. Journal of Acquired Immune Deficiency Syndromes. 2013;62(2):197–203. doi: 10.1097/QAI.0b013e31827ab612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Cagliani R, Riva S, Fumagalli M, et al. A positively selected APOBEC3H haplotype is associated with natural resistance to HIV-1 infection. Evolution. 2011;65(11):3311–3322. doi: 10.1111/j.1558-5646.2011.01368.x. [DOI] [PubMed] [Google Scholar]
- 110.Gourraud PA, Karaouni A, Woo JM, et al. APOBEC3H haplotypes and HIV-1 pro-viral vif DNA sequence diversity in early untreated human immunodeficiency virus-1 infection. Human Immunology. 2011;72(3):207–212. doi: 10.1016/j.humimm.2010.12.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Levy JA. HIV pathogenesis: 25 years of progress and persistent challenges. AIDS. 2009;23(2):147–160. doi: 10.1097/QAD.0b013e3283217f9f. [DOI] [PubMed] [Google Scholar]
- 112.Cen S, Guo F, Niu M, Saadatmand J, Deflassieux J, Kleiman L. The interaction between HIV-1 gag and APOBEC3G. The Journal of Biological Chemistry. 2004;279(32):33177–33184. doi: 10.1074/jbc.M402062200. [DOI] [PubMed] [Google Scholar]
- 113.Alce TM, Popik W. APOBEC3G is incorporated into virus-like particles by a direct interaction with HIV-1 gag nucleocapsid protein. The Journal of Biological Chemistry. 2004;279(33):34083–34086. doi: 10.1074/jbc.C400235200. [DOI] [PubMed] [Google Scholar]
- 114.Luo K, Liu B, Xiao Z, et al. Amino-terminal region of the human immunodeficiency virus type 1 nucleocapsid is required for human APOBEC3G packaging. Journal of Virology. 2004;78(21):11841–11852. doi: 10.1128/JVI.78.21.11841-11852.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Zennou V, Perez-Caballero D, Göttlinger H, Bieniasz PD. APOBEC3G incorporation into human immunodeficiency virus type 1 particles. Journal of Virology. 2004;78(21):12058–12061. doi: 10.1128/JVI.78.21.12058-12061.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Douaisi M, Dussart S, Courcoul M, Bessou G, Vigne R, Decroly E. HIV-1 and MLV Gag proteins are sufficient to recruit APOBEC3G into virus-like particles. Biochemical and Biophysical Research Communications. 2004;321(3):566–573. doi: 10.1016/j.bbrc.2004.07.005. [DOI] [PubMed] [Google Scholar]
- 117.Schäfer A, Bogerd HP, Cullen BR. Specific packaging of APOBEC3G into HIV-1 virions is mediated by the nucleocapsid domain of the gag polyprotein precursor. Virology. 2004;328(2):163–168. doi: 10.1016/j.virol.2004.08.006. [DOI] [PubMed] [Google Scholar]
- 118.Svarovskaia ES, Xu H, Mbisa JL, et al. Human apolipoprotein B mRNA-editing enzyme-catalytic polypeptide-like 3G (APOBEC3G) is incorporated into HIV-1 virions through interactions with viral and nonviral RNAs. The Journal of Biological Chemistry. 2004;279(34):35822–35828. doi: 10.1074/jbc.M405761200. [DOI] [PubMed] [Google Scholar]
- 119.Burnett A, Spearman P. APOBEC3G multimers are recruited to the plasma membrane for packaging into human immunodeficiency virus type 1 virus-like particles in an RNA-dependent process requiring the NC basic linker. Journal of Virology. 2007;81(10):5000–5013. doi: 10.1128/JVI.02237-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Wang T, Zhang W, Tian C, et al. Distinct viral determinants for the packaging of human cytidine deaminases APOBEC3G and APOBEC3C. Virology. 2008;377(1):71–79. doi: 10.1016/j.virol.2008.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Lecossier D, Bouchonnet F, Clavel F, Hance AJ. Hypermutation of HIV-1 DNA in the absence of the Vif protein. Science. 2003;300(5622):p. 1112. doi: 10.1126/science.1083338. [DOI] [PubMed] [Google Scholar]
- 122.Schröfelbauer B, Yu Q, Zeitlin SG, Landau NR. Human immunodeficiency virus type 1 Vpr induces the degradation of the UNG and SMUG Uracil-DNA glycosylases. Journal of Virology. 2005;79(17):10978–10987. doi: 10.1128/JVI.79.17.10978-10987.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Yang B, Chen K, Zhang C, Huang S, Zhang H. Virion-associated uracil DNA glycosylase-2 and apurinic/apyrimidinic endonuclease are involved in the degradation of APOBEC3G-edited nascent HIV-1 DNA. The Journal of Biological Chemistry. 2007;282(16):11667–11675. doi: 10.1074/jbc.M606864200. [DOI] [PubMed] [Google Scholar]
- 124.Kaiser SM, Emerman M. Uracil DNA glycosylase is dispensable for human immunodeficiency virus type 1 replication and does not contribute to the antiviral effects of the cytidine deaminase Apobec3G. Journal of Virology. 2006;80(2):875–882. doi: 10.1128/JVI.80.2.875-882.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Langlois M-A, Neuberger MS. Human APOBEC3G can restrict retroviral infection in avian cells and acts independently of both UNG and SMUG1. Journal of Virology. 2008;82(9):4660–4664. doi: 10.1128/JVI.02469-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Holmes RK, Koning FA, Bishop KN, Malim MH. APOBEC3F can inhibit the accumulation of HIV-1 reverse transcription products in the absence of hypermutation: comparisons with APOBEC3G. The Journal of Biological Chemistry. 2007;282(4):2587–2595. doi: 10.1074/jbc.M607298200. [DOI] [PubMed] [Google Scholar]
- 127.Iwatani Y, Chan DSB, Wang F, et al. Deaminase-independent inhibition of HIV-1 reverse transcription by APOBEC3G. Nucleic Acids Research. 2007;35(21):7096–7108. doi: 10.1093/nar/gkm750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Newman ENC, Holmes RK, Craig HM, et al. Antiviral function of APOBEC3G can be dissociated from cytidine deaminase activity. Current Biology. 2005;15(2):166–170. doi: 10.1016/j.cub.2004.12.068. [DOI] [PubMed] [Google Scholar]
- 129.Mbisa JL, Barr R, Thomas JA, et al. Human immunodeficiency virus type 1 cDNAs produced in the presence of APOBEC3G exhibit defects in plus-strand DNA transfer and integration. Journal of Virology. 2007;81(13):7099–7110. doi: 10.1128/JVI.00272-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Gillick K, Pollpeter D, Phalora P, Kim EY, Wolinsky SM, Malim MH. Suppression of HIV-1 infection by APOBEC3 proteins in primary human CD4+ T cells is associated with inhibition of processive reverse transcription as well as excessive cytidine deamination. Journal of Virology. 2013;87(3):1508–1517. doi: 10.1128/JVI.02587-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Guo F, Cen S, Niu M, Saadatmand J, Kleiman L. Inhibition of tRNA3Lys-primed reverse transcription by human APOBEC3G during human immunodeficiency virus type 1 replication. Journal of Virology. 2006;80(23):11710–11722. doi: 10.1128/JVI.01038-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Li X-Y, Guo F, Zhang L, Kleiman L, Cen S. APOBEC3G inhibits DNA strand transfer during HIV-1 reverse transcription. The Journal of Biological Chemistry. 2007;282(44):32065–32074. doi: 10.1074/jbc.M703423200. [DOI] [PubMed] [Google Scholar]
- 133.Wang X, Ao Z, Chen L, Kobinger G, Peng J, Yao X. The cellular antiviral protein APOBEC3G interacts with HIV-1 reverse transcriptase and inhibits its function during viral replication. Journal of Virology. 2012;86(7):3777–3786. doi: 10.1128/JVI.06594-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Luo K, Wang T, Liu B, et al. Cytidine deaminases APOBEC3G and APOBEC3F interact with human immunodeficiency virus type 1 integrase and inhibit proviral DNA formation. Journal of Virology. 2007;81(13):7238–7248. doi: 10.1128/JVI.02584-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Mbisa JL, Bu W, Pathak VK. APOBEC3F and APOBEC3G inhibit HIV-1 DNA integration by different mechanisms. Journal of Virology. 2010;84(10):5250–5259. doi: 10.1128/JVI.02358-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Casartelli N, Guivel-Benhassine F, Bouziat R, Brandler S, Schwartz O, Moris A. The antiviral factor APOBEC3G improves CTL recognition of cultured HIV-infected T cells. The Journal of Experimental Medicine. 2010;207(1):39–49. doi: 10.1084/jem.20091933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Norman JM, Mashiba M, McNamara LA, et al. The antiviral factor APOBEC3G enhances the recognition of HIV-infected primary T cells by natural killer cells. Nature Immunology. 2011;12(10):975–983. doi: 10.1038/ni.2087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Sheehy AM, Gaddis NC, Malim MH. The antiretroviral enzyme APOBEC3G is degraded by the proteasome in response to HIV-1 Vif. Nature Medicine. 2003;9(11):1404–1407. doi: 10.1038/nm945. [DOI] [PubMed] [Google Scholar]
- 139.Yu X, Yu Y, Liu B, et al. Induction of APOBEC3G ubiquitination and degradation by an HIV-1 Vif-Cul5-SCF complex. Science. 2003;302(5647):1056–1060. doi: 10.1126/science.1089591. [DOI] [PubMed] [Google Scholar]
- 140.Conticello SG, Harris RS, Neuberger MS. The Vif protein of HIV triggers degradation of the human antiretroviral DNA deaminase APOBEC3G. Current Biology. 2003;13(22):2009–2013. doi: 10.1016/j.cub.2003.10.034. [DOI] [PubMed] [Google Scholar]
- 141.Shirakawa K, Takaori-Kondo A, Kobayashi M, et al. Ubiquitination of APOBEC3 proteins by the Vif-Cullin5-ElonginB-ElonginC complex. Virology. 2006;344(2):263–266. doi: 10.1016/j.virol.2005.10.028. [DOI] [PubMed] [Google Scholar]
- 142.Mariani R, Chen D, Schröfelbauer B, et al. Species-specific exclusion of APOBEC3G from HIV-1 virions by Vif. Cell. 2003;114(1):21–31. doi: 10.1016/s0092-8674(03)00515-4. [DOI] [PubMed] [Google Scholar]
- 143.Mehle A, Strack B, Ancuta P, Zhang C, McPike M, Gabuzda D. Vif overcomes the innate antiviral activity of APOBEC3G by promoting its degradation in the ubiquitin-proteasome pathway. The Journal of Biological Chemistry. 2004;279(9):7792–7798. doi: 10.1074/jbc.M313093200. [DOI] [PubMed] [Google Scholar]
- 144.Opi S, Kao S, Goila-Gaur R, et al. Human immunodeficiency virus type 1 Vif inhibits packaging and antiviral activity of a degradation-resistant APOBEC3G variant. Journal of Virology. 2007;81(15):8236–8246. doi: 10.1128/JVI.02694-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Kao S, Khan MA, Miyagi E, Plishka R, Buckler-White A, Strebel K. The human immunodeficiency virus type 1 Vif protein reduces intracellular expression and inhibits packaging of APOBEC3G (CEM15), a cellular inhibitor of virus infectivity. Journal of Virology. 2003;77(21):11398–11407. doi: 10.1128/JVI.77.21.11398-11407.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Stopak K, de Noronha C, Yonemoto W, Greene WC. HIV-1 Vif blocks the antiviral activity of APOBEC3G by impairing both its translation and intracellular stability. Molecular Cell. 2003;12(3):591–601. doi: 10.1016/s1097-2765(03)00353-8. [DOI] [PubMed] [Google Scholar]
- 147.Mercenne G, Bernacchi S, Richer D, et al. HIV-1 Vif binds to APOBEC3G mRNA and inhibits its translation. Nucleic Acids Research. 2009;38(2):633–646. doi: 10.1093/nar/gkp1009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 148.Chaipan C, Smith JL, Hu WS, Pathak VK. APOBEC3G restricts HIV-1 to a greater extent than APOBEC3F and APOBEC3DE in human primary CD4+ T cells and macrophages. Journal of Virology. 2013;87(1):444–453. doi: 10.1128/JVI.00676-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Mulder LCF, Ooms M, Majdak S, et al. Moderate influence of human APOBEC3F on HIV-1 replication in primary lymphocytes. Journal of Virology. 2010;84(18):9613–9617. doi: 10.1128/JVI.02630-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Liddament MT, Brown WL, Schumacher AJ, Harris RS. APOBEC3F properties and hypermutation preferences indicate activity against HIV-1 in vivo . Current Biology. 2004;14(15):1385–1391. doi: 10.1016/j.cub.2004.06.050. [DOI] [PubMed] [Google Scholar]
- 151.Wiegand HL, Doehle BP, Bogerd HP, Cullen BR. A second human antiretroviral factor, APOBEC3F, is suppressed by the HIV-1 and HIV-2 Vif proteins. The EMBO Journal. 2004;23(12):2451–2458. doi: 10.1038/sj.emboj.7600246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Bourara K, Liegler TJ, Grant RM. Target cell APOBEC3C can induce limited G-to-A mutation in HIV-1. PLoS Pathogens. 2007;3(10):1477–1485. doi: 10.1371/journal.ppat.0030153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Dang Y, Lai MS, Wang X, Han Y, Lampen R, Zheng Y-H. Human cytidine deaminase APOBEC3H restricts HIV-1 replication. The Journal of Biological Chemistry. 2008;283(17):11606–11614. doi: 10.1074/jbc.M707586200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Dang Y, Wang X, Esselman WJ, Zheng Y-H. Identification of APOBEC3DE as another antiretroviral factor from the human APOBEC family. Journal of Virology. 2006;80(21):10522–10533. doi: 10.1128/JVI.01123-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Doehle BP, Schäfer A, Cullen BR. Human APOBEC3B is a potent inhibitor of HIV-1 infectivity and is resistant to HIV-1 Vif. Virology. 2005;339(2):281–288. doi: 10.1016/j.virol.2005.06.005. [DOI] [PubMed] [Google Scholar]
- 156.Harari A, Ooms M, Mulder LCF, Simon V. Polymorphisms and splice variants influence the antiretroviral activity of Human APOBEC3H. Journal of Virology. 2009;83(1):295–303. doi: 10.1128/JVI.01665-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Wang X, Abudu A, Son S, Dang Y, Venta PJ, Zheng Y-H. Analysis of human APOBEC3H haplotypes and anti-human immunodeficiency virus type 1 activity. Journal of Virology. 2011;85(7):3142–3152. doi: 10.1128/JVI.02049-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Albin JS, Harris RS. Interactions of host APOBEC3 restriction factors with HIV-1 in vivo: implications for therapeutics. Expert Reviews in Molecular Medicine. 2010;12(e4):1–26. doi: 10.1017/S1462399409001343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Vázquez-Pérez JA, Ormsby CE, Hernández-Juan R, Torres KJ, Reyes-Terán G. APOBEC3G mRNA expression in exposed seronegative and early stage HIV infected individuals decreases with removal of exposure and with disease progression. Retrovirology. 2009;6, article 23 doi: 10.1186/1742-4690-6-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Land AM, Ball TB, Luo M, et al. Human immunodeficiency virus (HIV) type 1 proviral hypermutation correlates with CD4 count in HIV-infected women from Kenya. Journal of Virology. 2008;82(16):8172–8182. doi: 10.1128/JVI.01115-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Kourteva Y, de Pasquale M, Allos T, McMunn C, D’Aquila RT. APOBEC3G expression and hypermutation are inversely associated with human immunodeficiency virus type 1 (HIV-1) burden in vivo . Virology. 2012;430:1–9. doi: 10.1016/j.virol.2012.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Ulenga NK, Sarr AD, Hamel D, Sankale J-L, Mboup S, Kanki PJ. The level of APOBEC3G (hA3G)-related G-to-A mutations does not correlate with viral load in HIV type 1-infected individuals. AIDS Research and Human Retroviruses. 2008;24(10):1285–1290. doi: 10.1089/aid.2008.0072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Piantadosi A, Humes D, Chohan B, McClelland RS, Overbaugh J. Analysis of the percentage of human immunodeficiency virus type 1 sequences that are hypermutated and markers of disease progression in a longitudinal cohort, including one individual with a partially defective vif. Journal of Virology. 2009;83(16):7805–7814. doi: 10.1128/JVI.00280-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Amoêdo ND, Afonso AO, Cunha SM, Oliveira RH, Machado ES, Soares MA. Expression of APOBEC3G/3F and G-to-A hypermutation levels in HIV-1-infected children with different profiles of disease progression. PloS One. 2011;6(8, article e24118) doi: 10.1371/journal.pone.0024118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Jin X, Brooks A, Chen H, Bennett R, Reichman R, Smith H. APOBEC3G/CEM15 (hA3G) mRNA levels associate inversely with human immunodeficiency virus viremia. Journal of Virology. 2005;79(17):11513–11516. doi: 10.1128/JVI.79.17.11513-11516.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Zhao M, Geng W, Jiang Y, et al. The associations of hA3G and hA3B mRNA levels with HIV disease progression among HIV-infected individuals of China. Journal of Acquired Immune Deficiency Syndromes. 2010;53(supplement 1):S4–S9. doi: 10.1097/QAI.0b013e3181c7d349. [DOI] [PubMed] [Google Scholar]
- 167.Ulenga NK, Sarr AD, Thakore-Meloni S, Sankalé J-L, Eisen G, Kanki PJ. Relationship between human immunodeficiency type 1 infection and expression of human APOBEC3G and APOBEC3F. The Journal of Infectious Diseases. 2008;198(4):486–492. doi: 10.1086/590212. [DOI] [PubMed] [Google Scholar]
- 168.Cho S-J, Drechsler H, Burke RC, Arens MQ, Powderly W, Davidson NO. APOBEC3F and APOBEC3G mRNA levels do not correlate with human immunodeficiency virus type 1 plasma viremia or CD4+ T-cell count. Journal of Virology. 2006;80(4):2069–2072. doi: 10.1128/JVI.80.4.2069-2072.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Biasin M, Piacentini L, Lo Caputo S, et al. Apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3G: a possible role in the resistance to HIV of HIV-exposed seronegative individuals. The Journal of Infectious Diseases. 2007;195(7):960–964. doi: 10.1086/511988. [DOI] [PubMed] [Google Scholar]
- 170.Ejima T, Hirota M, Mizukami T, Otsuka M, Fujita M. An anti-HIV-1 compound that increases steady-state expression of apoplipoprotein B mRNA-editing enzyme-catalytic polypeptide-like 3G. International Journal of Molecular Medicine. 2011;28(4):613–616. doi: 10.3892/ijmm.2011.737. [DOI] [PubMed] [Google Scholar]
- 171.Cen S, Peng Z-G, Li X-Y, et al. Small molecular compounds inhibit HIV-1 replication through specifically stabilizing APOBEC3G. The Journal of Biological Chemistry. 2010;285(22):16546–16552. doi: 10.1074/jbc.M109.085308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Nathans R, Cao H, Sharova N, et al. Small-molecule inhibition of HIV-1 Vif. Nature Biotechnology. 2008;26(10):1187–1192. doi: 10.1038/nbt.1496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 173.Xiao Z, Ehrlich E, Luo K, Xiong Y, Yu X-F. Zinc chelation inhibits HIV Vif activity and liberates antiviral function of the cytidine deaminase APOBEC3G. The FASEB Journal. 2007;21(1):217–222. doi: 10.1096/fj.06-6773com. [DOI] [PubMed] [Google Scholar]
- 174.Hultquist JF, Harris RS. Leveraging APOBEC3 proteins to alter the HIV mutation rate and combat AIDS. Future Virology. 2009;4(6):605–619. doi: 10.2217/fvl.09.59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175.Mulder LCF, Harari A, Simon V. Cytidine deamination induced HIV-1 drug resistance. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(14):5501–5506. doi: 10.1073/pnas.0710190105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176.Jern P, Russell RA, Pathak VK, Coffin JM. Likely role of APOBEC3G-mediated G-to-A mutations in HIV-1 evolution and drug resistance. PLoS Pathogens. 2009;5(4) doi: 10.1371/journal.ppat.1000367.e1000367 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Wood N, Bhattacharya T, Keele BF, et al. HIV evolution in early infection: selection pressures, patterns of insertion and deletion, and the impact of APOBEC. PLoS Pathogens. 2009;5(5) doi: 10.1371/journal.ppat.1000414.e1000414 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Haché G, Shindo K, Albin JS, Harris RS. Evolution of HIV-1 isolates that use a novel Vif-independent mechanism to resist restriction by human APOBEC3G. Current Biology. 2008;18(11):819–824. doi: 10.1016/j.cub.2008.04.073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Pillai SK, Wong JK, Barbour JD. Turning up the volume on mutational pressure: is more of a good thing always better? (A case study of HIV-1 Vif and APOBEC3) Retrovirology. 2008;5, article 26 doi: 10.1186/1742-4690-5-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Kim E-Y, Bhattacharya T, Kunstman K, et al. Human APOBEC3G-mediated editing can promote HIV-1 sequence diversification and accelerate adaptation to selective pressure. Journal of Virology. 2010;84(19):10402–10405. doi: 10.1128/JVI.01223-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Sadler HA, Stenglein MD, Harris RS, Mansky LM. APOBEC3G contributes to HIV-1 variation through sublethal mutagenesis. Journal of Virology. 2010;84(14):7396–7404. doi: 10.1128/JVI.00056-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Fourati S, Malet I, Lambert S, et al. E138K and M184I mutations in HIV-1 reverse transcriptase coemerge as a result of APOBEC3 editing in the absence of drug exposure. AIDS. 2012;26(13):1619–1624. doi: 10.1097/QAD.0b013e3283560703. [DOI] [PubMed] [Google Scholar]
- 183.Olson ME, Li M, Harris RS, Harki DA. Small-molecule APOBEC3G DNA cytosine deaminase inhibitors based on a 4-amino-1, 2, 4-triazole-3-thiol scaffold. ChemMedChem. 2013;8:112–117. doi: 10.1002/cmdc.201200411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Li M, Shandilya SMD, Carpenter MA, et al. First-in-class small molecule inhibitors of the single-strand DNA cytosine deaminase APOBEC3G. ACS Chemical Biology. 2012;7(3):506–517. doi: 10.1021/cb200440y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Matsuoka M, Jeang K-T. Human T-cell leukaemia virus type 1 (HTLV-1) infectivity and cellular transformation. Nature Reviews Cancer. 2007;7(4):270–280. doi: 10.1038/nrc2111. [DOI] [PubMed] [Google Scholar]
- 186.Kannian P, Green PL. Human T lymphotropic virus type 1 (HTLV-1): molecular biology and oncogenesis. Viruses. 2010;2(9):2037–2077. doi: 10.3390/v2092037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Richardson JH, Edwards AJ, Cruickshank JK, Rudge P, Dalgleish AG. In vivo cellular tropism of human T-cell leukemia virus type 1. Journal of Virology. 1990;64(11):5682–5687. doi: 10.1128/jvi.64.11.5682-5687.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Refsland EW, Stenglein MD, Shindo K, Albin JS, Brown WL, Harris RS. Quantitative profiling of the full APOBEC3 mRNA repertoire in lymphocytes and tissues: implications for HIV-1 restriction. Nucleic Acids Research. 2010;38(13):4274–4284. doi: 10.1093/nar/gkq174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Ooms M, Krikoni A, Kress AK, Simon V, Münk C. APOBEC3A, APOBEC3B, and APOBEC3H haplotype 2 restrict human T-lymphotropic virus type 1. Journal of Virology. 2012;86(11):6097–6108. doi: 10.1128/JVI.06570-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Vartanian J-P, Plikat U, Henry M, et al. HIV genetic variation is directed and restricted by DNA precursor availability. Journal of Molecular Biology. 1997;270(2):139–151. doi: 10.1006/jmbi.1997.1104. [DOI] [PubMed] [Google Scholar]
- 191.Fan J, Guangyong M, Nosaka K, et al. APOBEC3G generates nonsense mutations in human T-cell leukemia virus type 1 proviral genomes in vivo . Journal of Virology. 2010;84(14):7278–7287. doi: 10.1128/JVI.02239-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Mahieux R, Suspène R, Delebecque F, et al. Extensive editing of a small fraction of human T-cell leukemia virus type 1 genomes by four APOBEC3 cytidine deaminases. Journal of General Virology. 2005;86(9):2489–2494. doi: 10.1099/vir.0.80973-0. [DOI] [PubMed] [Google Scholar]
- 193.Sasada A, Takaori-Kondo A, Shirakawa K, et al. APOBEC3G targets human T-cell leukemia virus type 1. Retrovirology. 2005;2:1–10. doi: 10.1186/1742-4690-2-32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Wattel E, Vartanian J-P, Pannetier C, Wain-Hobson S. Clonal expansion of human T-cell leukemia virus type I-infected cells in asymptomatic and symptomatic carriers without malignancy. Journal of Virology. 1995;69(5):2863–2868. doi: 10.1128/jvi.69.5.2863-2868.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Cavrois M, Wain-Hobson S, Gessain A, Plumelle Y, Wattel E. Adult T-cell leukemia/lymphoma on a background of clonally expanding human T-cell leukemia virus type-1-positive cells. Blood. 1996;88(12):4646–4650. [PubMed] [Google Scholar]
- 196.Cavrois M, Leclercq I, Gout O, Gessain A, Wain-Hobson S, Wattel E. Persistent oligoclonal expansion of human T-cell leukemia virus type 1-infected circulating cells in patients with Tropical spastic paraparesis/HTLV-1 associated myelopathy. Oncogene. 1998;17(1):77–82. doi: 10.1038/sj.onc.1201906. [DOI] [PubMed] [Google Scholar]
- 197.Derse D, Hill SA, Princler G, Lloyd P, Heidecker G. Resistance of human T cell leukemia virus type 1 to APOBEC3G restriction is mediated by elements in nucleocapsid. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(8):2915–2920. doi: 10.1073/pnas.0609444104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Kao J-H, Chen D-S. Global control of hepatitis B virus infection. Lancet Infectious Diseases. 2002;2(7):395–403. doi: 10.1016/s1473-3099(02)00315-8. [DOI] [PubMed] [Google Scholar]
- 199.WHO. HBV Fact Sheet no. 204. http://www.who.int/mediacentre/factsheets/fs204/en/
- 200.Gonzalez MC, Suspène R, Henry M, Guétard D, Wain-Hobson S, Vartanian J-P. Human APOBEC1 cytidine deaminase edits HBV DNA. Retrovirology. 2009;6, article 96 doi: 10.1186/1742-4690-6-96. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201.Rösler C, Köck J, Kann M, et al. APOBEC-mediated interference with hepadnavirus production. Hepatology. 2005;42(2):301–309. doi: 10.1002/hep.20801. [DOI] [PubMed] [Google Scholar]
- 202.Suspène R, Guétard D, Henry M, Sommer P, Wain-Hobson S, Vartanian J-P. Extensive editing of both hepatitis B virus DNA strands by APOBEC3 cytidine deaminases in vitro and in vivo . Proceedings of the National Academy of Sciences of the United States of America. 2005;102(23):8321–8326. doi: 10.1073/pnas.0408223102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Baumert TF, Rösler C, Malim MH, von Weizsäcker F. Hepatitis B virus DNA is subject to extensive editing by the human deaminase AP0BEC3C. Hepatology. 2007;46(3):682–689. doi: 10.1002/hep.21733. [DOI] [PubMed] [Google Scholar]
- 204.Köck J, Blum HE. Hypermutation of hepatitis B virus genomes by APOBEC3G, APOBEC3C and APOBEC3H. Journal of General Virology. 2008;89(5):1184–1191. doi: 10.1099/vir.0.83507-0. [DOI] [PubMed] [Google Scholar]
- 205.Henry M, Guétard D, Suspène R, Rusniok C, Wain-Hobson S, Vartanian J-P. Genetic editing of HBV DNA by monodomain human APOBEC3 cytidine deaminases and the recombinant nature of APOBEC3G. PLoS ONE. 2009;4(1, article e4277) doi: 10.1371/journal.pone.0004277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Vartanian J-P, Henry M, Marchio A, et al. Massive APOBEC3 editing of hepatitis B viral DNA in cirrhosis. PLoS Pathogens. 2010;6(5) doi: 10.1371/journal.ppat.1000928.e1000928 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Turelli P, Mangeat B, Jost S, Vianin S, Trono D. Inhibition of Hepatitis B virus replication by APOBEC3G. Science. 2004;303(5665, article 1829) doi: 10.1126/science.1092066. [DOI] [PubMed] [Google Scholar]
- 208.Nguyen DH, Gummuluru S, Hu J. Deamination-independent inhibition of hepatitis B virus reverse transcription by APOBEC3G. Journal of Virology. 2007;81(9):4465–4472. doi: 10.1128/JVI.02510-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Zhang W, Zhang X, Tian C, et al. Cytidine deaminase APOBEC3B interacts with heterogeneous nuclear ribonucleoprotein K and suppresses hepatitis B virus expression. Cellular Microbiology. 2008;10(1):112–121. doi: 10.1111/j.1462-5822.2007.01020.x. [DOI] [PubMed] [Google Scholar]
- 210.Bonvin M, Achermann F, Greeve I, et al. Interferon-inducible expression of APOBEC3 editing enzymes in human hepatocytes and inhibition of hepatitis B virus replication. Hepatology. 2006;43(6):1364–1374. doi: 10.1002/hep.21187. [DOI] [PubMed] [Google Scholar]
- 211.Tanaka Y, Marusawa H, Seno H, et al. Anti-viral protein APOBEC3G is induced by interferon-α stimulation in human hepatocytes. Biochemical and Biophysical Research Communications. 2006;341(2):314–319. doi: 10.1016/j.bbrc.2005.12.192. [DOI] [PubMed] [Google Scholar]
- 212.Noguchi C, Hiraga N, Mori N, et al. Dual effect of APOBEC3G on Hepatitis B virus. Journal of General Virology. 2007;88(2):432–440. doi: 10.1099/vir.0.82319-0. [DOI] [PubMed] [Google Scholar]
- 213.Günther S, Sommer G, Plikat U, et al. Naturally occurring hepatitis B virus genomes bearing the hallmarks of retroviral G → A hypermutation. Virology. 1997;235(1):104–108. doi: 10.1006/viro.1997.8676. [DOI] [PubMed] [Google Scholar]
- 214.Margeridon-Thermet S, Shulman NS, Ahmed A, et al. Ultra-deep pyrosequencing of hepatitis b virus quasispecies from nucleoside and nucleotide reverse-transcriptase inhibitor (NRTI)-treated patients and NRTI-naive patients. The Journal of Infectious Diseases. 2009;199(9):1275–1285. doi: 10.1086/597808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Proto S, Taylor JA, Chokshi S, Navaratnam N, Naoumov NV. APOBEC and iNOS are not the main intracellular effectors of IFN-γ-mediated inactivation of Hepatitis B virus replication. Antiviral Research. 2008;78(3):260–267. doi: 10.1016/j.antiviral.2008.01.006. [DOI] [PubMed] [Google Scholar]
- 216.Jost S, Turelli P, Mangeat B, Protzer U, Trono D. Induction of antiviral cytidine deaminases does not explain the inhibition of hepatitis B virus replication by interferons. Journal of Virology. 2007;81(19):10588–10596. doi: 10.1128/JVI.02489-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Xu R, Zhang X, Zhang W, Fang Y, Zheng S, Yu X-F. Association of human APOBEC3 cytidine deaminases with the generation of hepatitis virus B x antigen mutants and hepatocellular carcinoma. Hepatology. 2007;46(6):1810–1820. doi: 10.1002/hep.21893. [DOI] [PubMed] [Google Scholar]
- 218.Moradpour D, Penin F, Rice CM. Replication of hepatitis C virus. Nature Reviews Microbiology. 2007;5(6):453–463. doi: 10.1038/nrmicro1645. [DOI] [PubMed] [Google Scholar]
- 219.Levrero M. Viral hepatitis and liver cancer: the case of hepatitis C. Oncogene. 2006;25(27):3834–3847. doi: 10.1038/sj.onc.1209562. [DOI] [PubMed] [Google Scholar]
- 220.Peng Z-G, Zhao Z-Y, Li Y-P, et al. Host apolipoprotein b messenger RNA-editing enzyme catalytic polypeptide-like 3G is an innate defensive factor and drug target against hepatitis C virus. Hepatology. 2011;53(4):1080–1089. doi: 10.1002/hep.24160. [DOI] [PubMed] [Google Scholar]
- 221.Komohara Y, Yano H, Shichijo S, Shimotohno K, Itoh K, Yamada A. High expression of APOBEC3G in patients infected with hepatitis C virus. Journal of Molecular Histology. 2006;37(8-9):327–332. doi: 10.1007/s10735-006-9059-0. [DOI] [PubMed] [Google Scholar]
- 222.Jiménez-Sousa MÁ, Almansa R, de la Fuente C, et al. Gene expression profiling in the first twelve weeks of treatment in chronic hepatitis C patients. Enfermedades Infecciosas y Microbiologia Clinica. 2011;29(8):573–580. doi: 10.1016/j.eimc.2011.04.006. [DOI] [PubMed] [Google Scholar]
- 223.Pillai SK, Abdel-Mohsen M, Guatelli J, et al. Role of retroviral restriction factors in the interferon-α-mediated suppression of HIV-1 in vivo . Proceedings of the National Academy of Sciences of the United States of America. 2012;109(8):3035–3040. doi: 10.1073/pnas.1111573109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 224.Muñoz N, Castellsagué X, de González AB, Gissmann L. HPV in the etiology of human cancer. Vaccine. 2006;24(supplement 3):S3/1–S3/10. doi: 10.1016/j.vaccine.2006.05.115. [DOI] [PubMed] [Google Scholar]
- 225.Vartanian J-P, Guétard D, Henry M, Wain-Hobson S. Evidence for editing of human papillomavirus DNA by APOBEC3 in benign and precancerous lesions. Science. 2008;320(5873):230–233. doi: 10.1126/science.1153201. [DOI] [PubMed] [Google Scholar]
- 226.Suspène R, Aynaud M-M, Koch S, et al. Genetic editing of herpes simplex virus 1 and epstein-barr herpesvirus genomes by Human APOBEC3 cytidine deaminases in culture and in vivo . Journal of Virology. 2011;85(15):7594–7602. doi: 10.1128/JVI.00290-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Gee P, Ando Y, Kitayama H, et al. APOBEC1-mediated editing and attenuation of herpes simplex virus 1 DNA indicate that neurons have an antiviral role during herpes simplex encephalitis. Journal of Virology. 2011;85(19):9726–9736. doi: 10.1128/JVI.05288-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228.Whitley RJ, Kimberlin DW, Roizman B. Herpes simplex viruses. Clinical Infectious Diseases. 1998;26(3):541–555. doi: 10.1086/514600. [DOI] [PubMed] [Google Scholar]
- 229.Whitley RJ, Roizman B. Herpes simplex virus infections. The Lancet. 2001;357(9267):1513–1518. doi: 10.1016/S0140-6736(00)04638-9. [DOI] [PubMed] [Google Scholar]
- 230.Chisholm C, Lopez L. Cutaneous infections caused by herpesviridae: a review. Archives of Pathology and Laboratory Medicine. 2011;135(10):1357–1362. doi: 10.5858/arpa.2010-0156-RS. [DOI] [PubMed] [Google Scholar]
- 231.Mendoza N, Diamantis M, Arora A, et al. Mucocutaneous manifestations of Epstein-Barr virus infection. American Journal of Clinical Dermatology. 2008;9(5):295–305. doi: 10.2165/00128071-200809050-00003. [DOI] [PubMed] [Google Scholar]
- 232.Kidd JM, Newman TL, Tuzun E, Kaul R, Eichler EE. Population stratification of a common APOBEC gene deletion polymorphism. PLoS Genetics. 2007;3(4, article e63) doi: 10.1371/journal.pgen.0030063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.OhAinle M, Kerns JA, Li MMH, Malik HS, Emerman M. Antiretroelement activity of APOBEC3H was lost twice in recent human evolution. Cell Host & Microbe. 2008;4(3):249–259. doi: 10.1016/j.chom.2008.07.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.An P, Winkler CA. Host genes associated with HIV/AIDS: advances in gene discovery. Trends in Genetics. 2010;26(3):119–131. doi: 10.1016/j.tig.2010.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 235.Lingappa JR, Petrovski S, Kahle E, et al. Genomewide association study for determinants of HIV-1 acquisition and viral set point in HIV-1 serodiscordant couples with quantified virus exposure. PLoS ONE. 2011;6(12, article e28632) doi: 10.1371/journal.pone.0028632. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.van Manen D, van Wout AB, Schuitemaker H. Genome-wide association studies on HIV susceptibility, pathogenesis and pharmacogenomics. Retrovirology. 2012;9(70):1–8. doi: 10.1186/1742-4690-9-70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 237.Kamatani Y, Wattanapokayakit S, Ochi H, et al. A genome-wide association study identifies variants in the HLA-DP locus associated with chronic hepatitis B in Asians. Nature Genetics. 2009;41(5):591–595. doi: 10.1038/ng.348. [DOI] [PubMed] [Google Scholar]
- 238.Zhang H, Zhai Y, Hu Z, et al. Genome-wide association study identifies 1p36.22 as a new susceptibility locus for hepatocellular carcinoma in chronic hepatitis B virus carriers. Nature Genetics. 2010;42(9):755–758. doi: 10.1038/ng.638. [DOI] [PubMed] [Google Scholar]
- 239.Li S, Qian J, Yang Y, et al. GWAS identifies novel susceptibility loci on 6p21. 32 and 21q21. 3 for hepatocellular carcinoma in chronic hepatitis B virus carriers. PLoS Genetics. 2012;8(7):1–8. doi: 10.1371/journal.pgen.1002791. [DOI] [PMC free article] [PubMed] [Google Scholar]