

Abstract
Both studies on animals and humans suggest the presence of dopamine (DA) receptors in the anterior segment of the eye. Their role in the dynamics of intraocular pressure (IOP) is not yet clear. DA2 and DA3 receptors are mainly located on postganglionic sympathetic nerve endings. Their stimulation reduces the release of norepinephrine and suppresses the production of aqueous humor. DA1 receptors seem to be more expressed by the ciliary body and the outflow pathway of aqueous humor. The administration of DA1-selective agonists stimulates the production of aqueous humor, increasing IOP, whereas DA2- and DA3-selective agonists could reduce IOP and, therefore, the risk to develop a glaucoma (GL). GL is a broad spectrum of eye diseases which have in common the damage to the optic nerve and the progressive loss of the visual field. Further studies are desirable to clarify the role of the dopaminergic system and the usefulness of DA2 and DA3 agonists in reducing IOP.
1. Introduction
Biological basis of glaucoma (GL) are not yet fully understood, and the factors involved in its progression are not yet fully identified. The most important risk factor is high intraocular pressure (IOP), due to the reduction in the outflow of aqueous humor through the iridocorneal angle [1, 2]. Other risk factors are aging (in elderly the prevalence of the disease is higher), family history, African-Caribbean race, high myopia (>4D), thinner cornea, lens pseudoexfoliation, low blood pressure (diastolic one in particular), and local and systemic vascular risk factors (optic disc hemorrhage or atrophy, vasospasm, etc.) [1–4].
Physiologically, IOP is regulated by a proper balance between secretion and drainage of aqueous humor. This is produced by ciliary body's epithelium into the posterior chamber from which passively diffuses into the anterior one through pupil [3–5]. Once in the anterior chamber, about 80% of aqueous humor outflows through the trabecular meshwork and then in a small vessel called Schlemm's canal. Finally, through aqueous veins, it comes out from ocular bulb by episcleral veins and reaches the bloodstream. The remaining 20% of aqueous humor flows, independently from IOP, through the uveoscleral outflow or along nerves and small ocular vessels [4–7].
High IOP is one of the most important risk factors involved in GL and is the only one against whom actual therapies are really effective. Epidemiological studies showed that the risk of GL increases by 12% for each 1 mmHg increase of IOP [1, 2, 8]. To date, GL is the second leading cause of blindness in the world [8]. GL can be congenital or acquired. The further classification in primary open-angle glaucoma (POAG) and primary angle-closure glaucoma (PACG) concerns the mechanisms that hinders aqueous humor outflow. In primary GL, the increase of IOP is not associated with other ocular disorders, while in secondary GL, a recognizable factor, ocular or nonocular one, alters the outflow of aqueous humor. In western countries, POAG is more common than PACG [1, 2, 8].
Molecular events responsible for GL are still for the most part unknown. Several molecules are able to regulate IOP: adrenergic, cholinergic, serotonergic, and dopaminergic systems are all involved.
Dopamine (DA) is a simple organic chemical in the catecholamine family released from postganglionic nerve fibers of the superior cervical dopaminergic ganglion in the aqueous humor [9–12] and exerts its action by binding to 5 different types of DA receptors (DA1–5) which belong to the superclass of G-proteins coupled receptors.
DA receptors are grouped in two distinct families: DA1-like receptors (which include DA1 and DA5 receptors) and DA2-like receptors, like DA2, DA3, and DA4 [11–14]. The main difference between those families of receptors concerns the action of G-proteins: DA1-like receptor has a Gs-protein whose activation results in an increase of cyclic AMP (cAMP) mediated by adenylate cyclase [9, 10, 12]. DA2-like receptors are coupled to a Gi-protein which determines, instead, the inhibition of adenylate cyclase and, consequentially, a decrease of cAMP (Figure 1).
Figure 1.
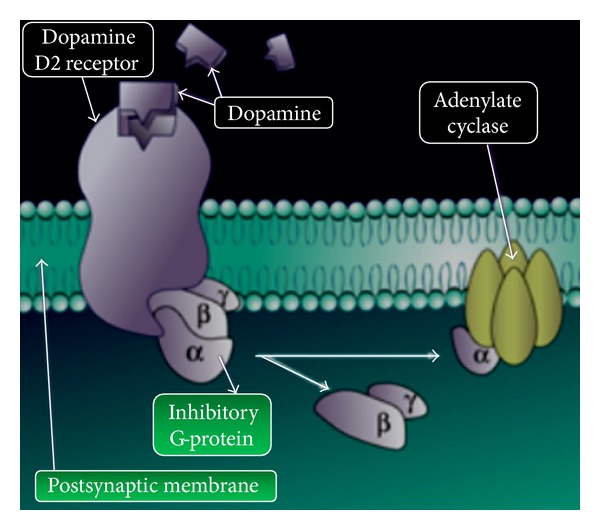
Structures of dopamine receptors (DA2/DA3) and G-protein coupled to receptors.
The aim of this review is to investigate, specifically, the dopaminergic influence on IOP to evaluate the role of new hypotensive drugs.
2. Studies on Animals
Early studies of the role of DA in GL on animal models led to contrasting results. Shannon et al. [15] showed that DA, administered topically, increases IOP in rabbit eyes, and haloperidol (DA inverse agonist) can contrast that action. Green and Elijah [16] noticed, by contrast, that IOP decreased during parenteral administration of DA, indicating that a different route of administration of DA produces different effects on IOP. In 1984, Potter et al. [17] studied the sequel of topical administration of DA and its methylated analogs: N-methyl-dopamine (NMDA), N,N-dimethyl dopamine (DMDA), and N,N-Di-n-propyl dopamine (DPDA) in eyes of healthy rabbits and of sympathectomized ones. Thus, in normal rabbits, DA, DMDA, and NMDA induced an increase of IOP unilaterally, while DPDA induced a bilateral hypotension after unilateral topical administration.
Furthermore, in sympathectomized rabbits they noted the following results:
the administration of NMDA and DMDA produced an exaggerated ocular hypertension, followed by hypotension;
the hypotensive effects of DPDA registered in normal rabbits eyes were absent in sympathectomized ones.
In summary, they assessed that DA and its analogues produce two different effects:
a direct and postjunctional effect, mediated by DA agonists that bind to DA1, α, and β receptors;
an indirect and prejunctional (or “neuronal”) effect, mediated by DA agonists that bind α 2 and DA2 receptors.
Green et al. [9, 16], studying rabbit's ciliary epithelium in vitro, observed that DA increases passive permeability and active secretion of aqueous humor. Both effects could be inhibited by α and β antagonists, phentolamine, and sotalol, respectively, but not by a relatively selective DA antagonist (butaclamol) [18]. The secretory function of DA was subsequently confirmed by studies that identified, on nonpigmented epithelium of the ciliary body of animals and humans, the phosphoprotein 3′–5′ monophosphate (DARPP-32) which regulates adenosine and DA systems [19].
Potter et al., however, showed that topical administration of some ergoline (lergotrile, pergolide) and an ergopeptide (bromocriptine), both in rabbits and monkeys, decreased IOP by acting on DA2 receptors [17, 20, 21]. The action of DA2 receptors was further confirmed by a subsequent study by Potter [12]. He noted a decrease hypotensive effect of ergoline in eyes of rabbits pretreated with domperidone (a DA2 antagonist) and surgical sympathectomy.
In summary:
the decrease of IOP mediated by ergot derivatives is partly dependent to sympathetic neuronal activity;
probably DA2 receptors are expressed in the sympathetic ganglion and/or sympathetic postganglionic nerveendings;
ocular hypotension is just partially due to the inhibition of formation of aqueous humor. To further confirm that indirect effects are responsible for lowering IOP, Savolainen et al. [22] reported that selective DA2/α 2 agonist like CHF1035 and its metabolite CHF1024, if instilled in rabbit's eyes, have similar effects to brimonidine in lowering IOP, but CHF1035 has a long lasting action too.
These data suggest the presence of DA receptors in the anterior segment, but in order to better understand and investigate their role, it was important to precisely identify where the structures were present. Many biochemical, immunohistochemical and functional studies have been performed on animal tissue samples to find DA1 receptors localization, but not DA2 ones. DA1 receptors are present on ciliary body epithelium of bovine [11, 19] and on ciliary processes epithelium of rabbit [9, 15, 18] confirming that their stimulation causes from one side an increased production of aqueous humor and, on the other hand, an altered outflow of it affecting the tone of ciliary muscle. DA2 receptors main localization seems to be the postganglionic presynaptic nerve ending [22].
In addition to the previously mentioned receptors, Chu et al. showed the involvement of a specific DA receptor, the DA3, responsible for IOP hypotonization in an animal model of rabbit [23]. Recently, some authors studied the hypotensive effect of 7-hydroxy-2-dipropilaminotetralina (7OH-DPAT), the main agonist of DA3 receptor on rabbit eyes [23], showing that
IOP lowering effect of 7-OH-DPAT was due in part to the reduction of the production of aqueous humor;
the nervous site of action of 7-OH-DPAT has been identified immunohistochemically as the DA3 receptor of ciliary body and has been associated with the decrease of hypotensive effects in rabbits deprived of sympathetic nerve endings;
the inhibition of the hypotensive effect of 7-OH-DPAT by DA3 receptor antagonists (U99194A and UH232) and the deprivation of postganglionic sympathetic endings, together with the immunohistochemical data, showed that the primary site of action of 7-OH-DPAT is located in the presynaptic postganglionic nerve endings of the ciliary body of rabbit eyes.
A similar study, conducted on mice, was performed by Bucolo et al. [14]. The authors observed that topical application of 7-OH-DPAT, α DA3 preferring receptor against, significantly decreased in wild-type (WT) mice both in an ocular normotensive group and an ocular hypertensive steroid-induced group. Instead, there were no changes of IOP in DA3 receptor knockout mice. Moreover, genetic analysis conducted on ocular tissues of WT mice with PCR showed several genes encoding for all DA receptors but the absence of the receptor gene in DA3 KO mice [14].
3. Studies on Humans
The expression of DA2 receptors in the anterior segment of the eye in humans has been confirmed by functional data, but there are no studies which revealed their localization.
Unlike DA2 receptors, DA1 have also been identified in human eyes [19] with biochemical and autoradiographic techniques using [3]-SCH-23390, a selective antagonist of DA1 receptor. DA1 receptors were present on uveoscleral tissue, trabecular meshwork, and ciliary processes [7]. Eleven eyes, enucleated because of trauma but with no involvement of the iridocorneal angle, were analyzed. Seven eyes were treated for GL, while four of them had a normal IOP. Although the sample size was very small, a higher response, due to DA1 receptors, was noticed in eyes with normal IOP [7].
Mekki et al. demonstrated, in a double-blind study, that in 8 healthy volunteers without IOP alteration treated with oral bromocriptine (1.25 mg), IOP decreased (compared to placebo) at 3 and 4 hours after the administration, with no effects on systemic arterial pressure and on pupil's diameter [24, 25]. These results were later confirmed by a study involving patients suffering from Parkinson's disease and POAG. In fact, a decrease in IOP occurred when levodopa was replaced by bromocriptine per os (2.5 mg) [26]. This drug has effects on the production of prolactin, and Mekki et al. showed that the eye drops containing bromocriptine can reduce IOP compared to placebo [24, 25].
In a double-blind, randomized, and prospective study, IOP was measured in 24 healthy subjects at baseline and 2, 4, and 6 hours after the instillation of topical timolol (0.25%) and bromocriptine (0.05%). Both bromocriptine and timolol decreased IOP significantly from baseline with no difference between them at 2 and 4 h, but, at 6 h, bromocriptine was more effective than timolol [27].
Al-Sereiti et al. showed that the hypotensive effect of pergolide (25 micrograms orally) was antagonized by a pretreatment with oral domperidone (15 mg), a DA2 receptor low-selective peripheral antagonist [28].
DA1 receptor agonists were topically administered (DA, ibopamine, fenoldopam, and 3B90) in a study performed on 20 patients with POAG [29]. The authors demonstrated that the increase of IOP after the administration of those molecules was not inhibited neither by the administration of haloperidol nor by already known hypotensive drugs. Just the administration of a selective DA1 agonist (SCH-23390) was capable to reduce IOP, confirming that those receptors are involved in determining IOP.
Karnezis et al. showed that intravenous administration of angiotensin II, norepinephrine, and fenoldopam increased IOP in human subjects by the stimulation of DA1 receptors [30]. Subsequently, it was confirmed that the stimulation by fenoldopam could alter the outflow of the aqueous humor increasing IOP [31].
Among DA1 receptor agonists, we can also include the 3,4-di-isobutyrrylester of N-methyldopamine, better known as ibopamine [32, 33]. This widely studied molecule has the following pharmacological properties:
it increases the production of aqueous humor stimulating DA1 receptors both in healthy eyes and in glaucomatous ones;
it is a valid provocative test to study the efficiency of aqueous humor outflow pathways; in fact, in close relatives of patients with POAG, the instillation of ibopamine 2% causes an increase in IOP which would not occur in subjects without familiarity for POAG.
Those data explain that high IOP due to the administration of ibopamine is not caused by the increased production of aqueous humor but by an alteration of the outflow pathways [13, 32–34]. Ibopamine induces a cycloplegic mydriasis not acting on α receptors. Moreover, it can be used in hypotensive syndromes like anterior uveitis or damage of the ciliary body or after GL filtration surgery [33, 34].
Recently, Virno et al. [35] assessed the effect of ibopamine, a D1-dopamine agonist, on IOP in offsprings of parents with POAG as a consequence of outflow structures impairment. D1-dopaminergic stimulation due to ibopamine increases IOP as a result of increased production of the aqueous humor in participants with an impaired outflow. The study showed that after 2% ibopamine administration, there was a significant increase in aqueous humor production, both in glaucomatous and normal eyes. The intraocular hypertensive response due to ibopamine in normotensive eyes is a sign of initial outflow impairment and, therefore, a predisposition to intraocular hypertension and possible GL.
After this overview, we can conclude that the effects of DA and its analogs on IOP are complex: they can have both direct effects (postjunctional) and indirect (prejunctional) ones. The postjunctional effects of DA agonists can stimulate α, β, and DA1 receptors. Similarly, indirect effects neuronal may be mediated by α 2, DA2 and DA3 receptors (Figure 2). In particular, DA1 agonists seems to increase the production of aqueous humor with a consequent increase of IOP, while, DA2 agonists are responsible of a decrease of IOP (Figure 3).
Figure 2.

Prejunctional, indirect effects and postjunctional direct effects of dopamine (DA) and its analogues.
Figure 3.
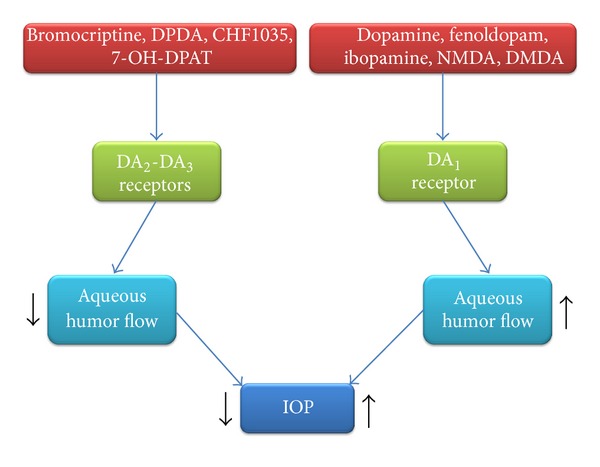
Effects of dopamine (DA) receptors on aqueous humor flow. DMDA: N,N-dimethyldopamine; DPDA: N,N-di-n-propyldopamine; NMDA: N-methyldopamine; 7-OH-DPAT: 7-hydroxy-2-dipropylaminotetralin.
4. Conclusions
Although several classes of drugs are known to reduce IOP, just 5 of them are used in clinical practice. It is still necessary to look for the most powerful (and safer) therapeutic agent in order to reduce the serious consequences of GL. From the studies carried out on animals and humans, discussed previously, we can conclude that DA2, and DA3 agonists have an important influence on the modulation of IOP, with significant implications for the management of patients affected by GL.
Further studies are needed to clarify the role of the dopaminergic system and the usefulness of DA2 and DA3 agonists in lowering IOP.
Conflict of Interests
There are no potential conflict of interests or any financial or personal relationships with other people or organizations that could inappropriately influence conduct and findings of this study.
References
- 1.Quigley HA, Broman AT. The number of people with glaucoma worldwide in 2010 and 2020. British Journal of Ophthalmology. 2006;90(3):262–267. doi: 10.1136/bjo.2005.081224. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Quigley HA. Glaucoma. The Lancet. 2011;377(9774):1367–1377. doi: 10.1016/S0140-6736(10)61423-7. [DOI] [PubMed] [Google Scholar]
- 3.Zhong Y, Wang J, Luo X. Integrins in trabecular meshwork and optic nerve head: possible association with the pathogenesis of glaucoma. Journal of Biomedicine and Biotechnology. 2013;2013:8 pages. doi: 10.1155/2013/202905.202905 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Pescosolido N, Cavallotti C, Rusciano D, Nebbioso M. Trabecular meshwork in normal and pathological eyes. Ultrastructural Pathology. 2012;36(2):102–107. doi: 10.3109/01913123.2011.634090. [DOI] [PubMed] [Google Scholar]
- 5.Scarsella G, Nebbioso M, Stefanini S, Librando A, Pescosolido N. Degenerative effects in rat eyes after experimental ocular hypertension. European Journal of Histochemistry. 2012;56(4, article e42):265–271. doi: 10.4081/ejh.2012.e42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Brubaker RF. Measurement of uveoscleral outflow in humans. Journal of Glaucoma. 2001;10(5):S45–S48. doi: 10.1097/00061198-200110001-00017. [DOI] [PubMed] [Google Scholar]
- 7.Cavallotti C, Pescosolido N, Pescosolido V, Iannetti G. Determination of dopamine D1 receptors in the human uveo scleral tissue by light microscope autoradiography. International Ophthalmology. 2001;23(3):171–179. doi: 10.1023/a:1010611419602. [DOI] [PubMed] [Google Scholar]
- 8.Kingman S. Glaucoma is second leading cause of blindness globally. Bulletin of the World Health Organization. 2004;82(11):887–888. [PMC free article] [PubMed] [Google Scholar]
- 9.Green K, Hensley A, Lollis G. Dopamine stimulation of passive permeability and secretion in the isolated rabbit ciliary epithelium. Experimental Eye Research. 1979;29(4):423–427. doi: 10.1016/0014-4835(79)90059-9. [DOI] [PubMed] [Google Scholar]
- 10.Libet B. Which postsynaptic action of dopamine is mediated by cyclic AMP? Life Sciences. 1979;24(12):1043–1057. doi: 10.1016/0024-3205(79)90037-7. [DOI] [PubMed] [Google Scholar]
- 11.Lograno MD, Daniele E, Govoni S. Biochemical and functional evidence for the presence of dopamine D1 receptors in the bovine ciliary body. Experimental Eye Research. 1990;51(5):495–501. doi: 10.1016/0014-4835(90)90079-a. [DOI] [PubMed] [Google Scholar]
- 12.Potter DE. Do dopamine and dopamine receptors have roles in modulating function in the anterior segment? The evidence. Progress in Retinal and Eye Research. 1996;15(1):103–111. [Google Scholar]
- 13.Prünte C, Nuttli I, Markstein R, Kohler C. Effects of dopamine D-1 and D-2 receptors on intraocular pressure in conscious rabbits. Journal of Neural Transmission. 1997;104(2-3):111–123. doi: 10.1007/BF01273174. [DOI] [PubMed] [Google Scholar]
- 14.Bucolo C, Leggio GM, Maltese A, Castorina A, D’Agata V, Drago F. Dopamine-3 receptor modulates intraocular pressure: implications for glaucoma. Biochemical Pharmacology. 2012;83(5):680–686. doi: 10.1016/j.bcp.2011.11.031. [DOI] [PubMed] [Google Scholar]
- 15.Shannon RP, Mead A, Sears ML. The effect of dopamine on the intraocular pressure and pupil of the rabbit eye. Investigative Ophthalmology. 1976;15(5):371–380. [PubMed] [Google Scholar]
- 16.Green K, Elijah D. Drug effects on aqueous humor formation and pseudofacility in normal rabbit eyes. Experimental Eye Research. 1981;33(3):239–245. doi: 10.1016/s0014-4835(81)80047-4. [DOI] [PubMed] [Google Scholar]
- 17.Potter DE, Burke JA, Chang FW. Alteration in ocular function induced by phenylethylamine analogs of dopamine. Current Eye Research. 1984;3(6):851–859. doi: 10.3109/02713688409000798. [DOI] [PubMed] [Google Scholar]
- 18.Mancino R, Cerulli L, Ricci A, Amenta F. Direct demonstration of dopamine D1-like receptor sites in the ciliary body of the rabbit eye by light microscope autoradiography. Naunyn-Schmiedeberg’s Archives of Pharmacology. 1992;346(6):644–648. doi: 10.1007/BF00168737. [DOI] [PubMed] [Google Scholar]
- 19.Stone RA, Laties AM, Hemmings HC, Jr., Ouimet CC, Greengard P. DARPP-32 in the ciliary epithelium of the eye: a neurotransmitter-regulated phosphoprotein of brain localizes to secretory cells. The Journal of Histochemistry and Cytochemistry. 1986;34(11):1465–1468. doi: 10.1177/34.11.2877023. [DOI] [PubMed] [Google Scholar]
- 20.Potter DE, Burke JA. Effects of ergoline derivatives on intraocular pressure and iris function in rabbits and monkeys. Current Eye Research. 1983;2(5):281–288. doi: 10.3109/02713688209000771. [DOI] [PubMed] [Google Scholar]
- 21.Potter DE, Burke JA. An in vivo model for dissociating alpha 2-and DA2-adrenoceptor activity in an ocular adnexa: utility of the cat nictitating membrane preparation. Current Eye Research. 1984;3(11):1289–1298. doi: 10.3109/02713688409007415. [DOI] [PubMed] [Google Scholar]
- 22.Savolainen J, Rautio J, Razzetti R, Järvinen T. A novel D2-dopaminergic and alpha2-adrenoceptor receptor agonist induces substantial and prolonged IOP decrease in normotensive rabbits. The Journal of Pharmacy and Pharmacology. 2003;55(6):789–794. doi: 10.1211/002235703765951393. [DOI] [PubMed] [Google Scholar]
- 23.Chu E, Chu T-C, Potter DE. Mechanisms and sites of ocular action of 7-hydroxy-2- dipropylaminotetralin: a dopamine3 receptor agonist. Journal of Pharmacology and Experimental Therapeutics. 2000;293(3):710–716. [PubMed] [Google Scholar]
- 24.Mekki QA, Hassan SM, Turner P. Bromocriptine lowers intraocular pressure without affecting blood pressure. The Lancet. 1983;1(8336):1250–1251. doi: 10.1016/s0140-6736(83)92699-5. [DOI] [PubMed] [Google Scholar]
- 25.Mekki QA, Warrington SJ, Turner P. Bromocriptine eyedrops lower intraocular pressure without affecting prolactin levels. The Lancet. 1984;1(8371):287–288. doi: 10.1016/s0140-6736(84)90162-4. [DOI] [PubMed] [Google Scholar]
- 26.Lustig A. Does bromocriptine enhance the effect of drugs that reduce intraocular pressure? Deutsche Medizinische Wochenschrift. 1983;108(43):1656–1657. [PubMed] [Google Scholar]
- 27.Elibol O, Güler C, Arici K, Topalkara A, Demircan S. The determination of additive effect and intraocular pressure lowering effects of 0.05% bromocriptine and 0.25% timolol. International Ophthalmology. 1997;20(1–3):53–55. doi: 10.1007/BF00212946. [DOI] [PubMed] [Google Scholar]
- 28.Al-Sereiti MR, Quik RFP, Hedges A, Turner P. Antagonism by domperidone of the ocular hypotensive effect of pergolide. European Journal of Clinical Pharmacology. 1990;38(5):461–463. doi: 10.1007/BF02336684. [DOI] [PubMed] [Google Scholar]
- 29.Virno M, Gazzaniga A, Taverniti L, Pecori Giraldi J, de Gregorio F. Dopamine, dopaminergic drugs and ocular hypertension. International Ophthalmology. 1992;16(4-5):349–353. doi: 10.1007/BF00917989. [DOI] [PubMed] [Google Scholar]
- 30.Karnezis TA, Murphy MB, Weber RR, Nelson KS, Tripathi BJ, Tripathi RC. Effects of selective dopamine-1 receptor activation on intraocular pressure in man. Experimental Eye Research. 1988;47(5):689–697. doi: 10.1016/0014-4835(88)90036-x. [DOI] [PubMed] [Google Scholar]
- 31.Piltz JR, Stone RA, Boike S, et al. Fenoldopam, a selective dopamine-1 receptor agonist, raises intraocular pressure in males with normal intraocular pressure. Journal of Ocular Pharmacology and Therapeutics. 1998;14(3):203–216. doi: 10.1089/jop.1998.14.203. [DOI] [PubMed] [Google Scholar]
- 32.Prünte C, Flammer J. The novel dopamine D-1 antagonist and D-2 agonist, SDZ GLC-756, lowers intraocular pressure in healthy human volunteers and in patients with glaucoma. Ophthalmology. 1995;102(9):1291–1297. doi: 10.1016/s0161-6420(95)30872-x. [DOI] [PubMed] [Google Scholar]
- 33.Magacho L, Lima FE, Costa ML, Fayad FA, Guimarães NL, Avila MP. Ibopamine provocative test and glaucoma: consideration of factors that may influence the examination. Current Eye Research. 2004;28(3):189–193. doi: 10.1076/ceyr.28.3.189.26252. [DOI] [PubMed] [Google Scholar]
- 34.Hepsen IF, Yilmaz H, Keskin UC. The comparison of topical ibopamine 2% with tonography to identify the outflow resistance in eyes with ocular hypertension. Ophthalmic Research. 2005;37(1):17–22. doi: 10.1159/000082779. [DOI] [PubMed] [Google Scholar]
- 35.Virno M, Sampaolesi R, Pecori Giraldi J, et al. Ibopamine: D1-dopaminergic agonist in the diagnosis of glaucoma. Journal of Glaucoma. 2011 doi: 10.1097/IJG.0b013e3182311db4. [DOI] [PubMed] [Google Scholar]