

Abstract
The study of the fungal microbiota (‘mycobiome’) is a new and rapidly emerging field that lags behind our understanding of the bacterial microbiome. Every human has fungi as part of their microbiota, but the total number of fungal cells is orders of magnitude smaller than that of the bacterial microbiota. However, the impact of the mycobiome on human health is significant, especially as a reservoir for blooms of pathogenic microbes when the host is compromised and as a potential cofactor in inflammatory diseases and metabolic disorders.
Keywords: fungi, microbiome, health, disease, yeast, mucosal
The human microbiome
Advances in high-throughput sequencing and bioinformatics are just beginning to reveal the breadth, depth, and diversity of the fungal world in humans and throughout the environment. Nobody is fungus-free. Although there is currently no reliable estimate of the fungal load in the human body, the total number of fungal cells in the microbiome (the ‘mycobiome’) is orders of magnitude smaller than that of the bacterial microbiome (which is estimated to be about 100 trillion cells) [1]. Every individual’s microbiome contains thousands of different species of microbes, of which 99.9% of the total number of microbial cells belong to only a few species. The less abundant (<0.1%), but more diverse, component of the microbiome has been termed the ‘rare biosphere’ [2,3]. The impact of this rare biosphere on human health is significant because it can act as a reservoir for blooms of pathogenic microbes when the host is compromised. A recently described example of a low abundance species of the bacterial microbiome altering local microbiome composition, host immune responses, and driving development of inflammatory disease has been reported for Porphyromonas gingivalis in the development of periodontitis [4]. Humans have a fungal microbiome and it is clearly part of the rare biosphere of their entire microbiome (Figure 1). The vast majority of studies have focused on fungal outgrowth when the host is compromised, with little known about the dynamics of the mycobiome during health. This is a new and rapidly emerging field that lags behind our understanding of the bacterial microbiome. This review will discuss what is known about some of the members of the mycobiome, highlight recent culture-independent studies, and discuss implications of changes in the mycobiome as a potential cofactor in inflammatory diseases and metabolic disorders.
Figure 1.

The rare biosphere and the fungal microbiome. (A) Representation of the membership of the microbiome, displayed as a rank abundance curve and illustrating the concept of the ‘rare biosphere’ within the entirety of the microbiome and the relative position of a microbe at 0.01% abundance (asterisk) in this example. In the complex microbial communities of the mucosa, a relatively small number of species dominate, but hundreds to thousands of low abundance species also exist in the communities. At mucosal sites where bacterial abundance can reach >1010/g, a microbe present at 104/g would be <0.0001% of the cellular content of the community. Microbial diversity in the microbiome is extensive and the large number of low abundance species (or operational taxonomic units, OTUs) in mucosal samples have been described as part of a rare biosphere [2,3]. However, the rare biosphere serves as a reservoir for potentially pathogenic microbes, such as members of the fungal microbiome, which grow out (bloom) and cause disease when the mucosal environment is disturbed. The rare biosphere may also harbor species that have a disproportionate effect (positive or negative) on the dominant members of the microbiome, a potential pathway by which the fungal microbiome may promote chronic disease. (B) Estimated relative cellular abundance of fungi to bacteria in human feces, based upon the MetaHIT metagenomic analysis [1].
The fungal microbiome
Disease caused by indigenous fungal organisms such as Candida albicans in immunocompromised and antibiotic-treated individuals is a major cause of morbidity and mortality in critical care settings, as well as playing a major role in other diseases of the mouth, stomach, and vaginal tract. Some disease-causing fungi, such as Histoplasma, Coccidiodes, Paracoccidioides, Blastomyces, and Aspergillus, are considered to be of exogenous origin and, therefore, considered strict pathogens. Others, such as Candida and Malassezia can be found at significant levels on mucosal surfaces and the skin, even in healthy hosts, and it is from within their niche in the microbiome that they can cause disease. Cryptococcus neoformans can exist at low levels for decades in the lungs without causing disease but then bloom to cause a life-threatening illness during immunosuppression. Pneumocystis is an example of a fungus exhibiting species-specific growth and can cause pneumonia in immunocompromised individuals, most likely as an outgrowth from an indigenous niche because it can be found at low levels in healthy individuals. Thus, these latter four types of fungi are potential pathogens, that is, commensals when the host environment is healthy or undisturbed, and pathogenic when the host environment is compromised or disturbed (Figure 2).
Figure 2.

Mechanisms by which the fungal microbiome can grow out and cause disease on mucosal surfaces. The fungal microbiome exists on mucosal surfaces in a symbiotic relationship with the host and the bacterial microbiome. Any of the following will promote the outgrowth of fungi on a mucosal surface: disruption of bacteria-mediated colonization resistance by antibiotics; damage to the mucosa by uncontrolled inflammatory responses, physical damage, or chemical-mediated injury; or defects in host defenses/immunity that affect innate immunity or specific facets of adaptive immunity.
Unlike for some members of the bacterial microbiota, there is no strong evidence for a mutualistic or beneficial relationship with the fungal microbiome. However, there is also little research in this arena. One example of a beneficial fungus in man is Saccharomyces cerevisiae var. boulardii, a well-described probiotic for the relief of gastroenteritis in animal systems and some humans [5]. S. boulardii can be considered a beneficial yeast, although it can cause fungemia in humans when it colonizes indwelling catheters from aerosolization during opening of lyophilized packets [6]. Because many host–microbe mechanisms are generally conserved between human–fungi and plant–fungi interactions, this suggests that there still remains much to be discovered about the spectrum of the ecological relationships between the fungal microbiome and its host. The roles of the fungal microbiome as a cofactor in inflammatory and metabolic disorders and in modulating the bacterial microbiome (thereby indirectly impacting the host) are areas of emerging interest in human health.
Detection of fungi
The methods and approaches to detect, identify, and analyze fungal communities on mucosal surfaces have significantly evolved over the last decade. Historically, the ability to culture fungi was critical for their detection and identification: via microscopy, biochemical assays, and selective media. With the introduction of polymerase chain reaction (PCR) technology to the field in the early 1990s, DNA-based detection began to revolutionize the methodologies for detection of fungi. As genomic sequencing was applied to both environmental and medically important fungi, the database for studying fungi began to expand. Although the fungal databases lag way behind those for bacteria, such as the Ribosomal Database Project, it is now clear that there are many fungi that elude detection by standard culture techniques. The major target for culture-independent analyses and database generation has been the genetic locus containing the 18S, 5.8S, and 28S rRNA genes as well as the internal transcribed spacer regions (ITS1 and ITS2), the DNA that encodes for the nonfunctional RNA transcribed during rRNA synthesis (Figure 3). Next generation sequencing (specifically the use of pyrosequencing of the fungal ITS1 and ITS2 regions) and clone libraries (built from the 18S, 5.8S, 28S regions as well as ITS1 and ITS2) have revealed a previously unrecognized diversity of fungi in the microbiome, both culturable and nonculturable, as members of the rare biosphere [7–11]. The role of the diverse fungal biosphere in host metabolism, immunity, and modulation of the bacterial microbiome remains largely unknown. However, the surprising identification of organisms such as Cryptococcus in 20% of oral samples of healthy individuals [10] raises the possibility that even culturable fungi may exist at low levels in a difficult-to-culture state in healthy individuals, thereby providing a niche for outgrowth and disease when host defenses and/or competitive exclusion become compromised.
Figure 3.
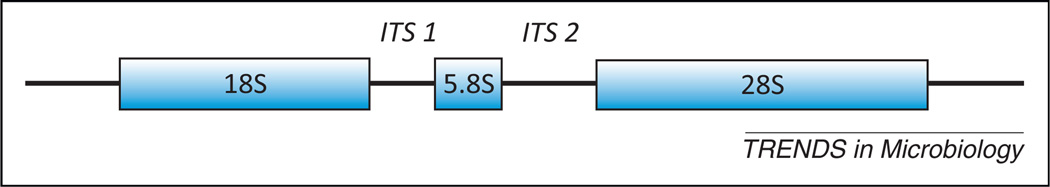
Fungal ribosomal gene locus. The most commonly used genetic region used for culture-independent molecular analysis of fungal diversity. Both clone library/sanger sequencing and pyrosequencing have been used to map and catalogue this region. High-throughput pyrosequencing has been used for amplicon libraries generated through pan-fungal internal transcribed spacer regions (ITS) primers that span the region between the 3′ end of the 18S gene, includes the entire 5S gene, and ends in the 5′ region of the 28S gene [9]. This region has been previously demonstrated to amplify a wide range of medically relevant fungi by PCR and pyrosequencing [7–11].
Fungal microbiome of mucosal sites
Chronic colonization of mucosal surfaces, including the urogenital tract, oral cavity, and gastrointestinal (GI) tract with fungi is common, especially for Candida spp., and carriage rates of Candida among healthy adults ranges from 30 to 70% [12,13]. The genus Candida includes approximately 160 species, most of which are adapted to live in mammalian hosts, causing infections in both immunocompetent and immunocompromised individuals [14]. In humans, C. albicans is the most abundant and significant species associated with disease. Other medically important species include Candida glabrata, Candida rugosa, Candida parapsilosis, Candida tropicalis, Candida dubliniensis, Candida krusei, and Candida lusitaniae. Candida spp. are uniquely adapted to their human hosts and acquisition occurs at birth or with later human contact. Strain types are often the same in the same individual at different tissue sites. In addition, the population of strains associated with infection is statistically the same as that isolated in the absence of infection [15]. Because Candida can colonize a variety of microbiota-containing host niches (both biotic tissues and abiotic indwelling medical devices), its pathogenic potential from within the microbiome is related to its ability to adapt, survive, and grow in constantly changing environments. These include the potential to adhere to and invade host cells, the ability to undergo morphological transitions and form biofilms, and specific metabolic and fitness traits. These latter traits could either allow Candida to synergize with or antagonize other members of the microbiota; likewise, these traits may facilitate asymptomatic colonization or symptomatic infection in the face of host immune defenses [16].
Most individuals are asymptomatically colonized with this organism. However, when environmental conditions permit the outgrowth of C. albicans, colonization can lead to infection and invasion of host tissues. Candida infections may remain localized to mucosal sites (e.g., oral or vaginal candidiasis) or may spread hematogenously leading to candidemia or deep-seated mycoses of other tissues, which can lead to death. Two environmental variables that can lead to increased levels of C. albicans in the host are immunosuppression and antibiotic treatment, the latter of which decreases bacterial colonization resistance against Candida (Figure 2). The risk factors for mucosal overgrowth of Candida and/or inflammation are niche specific [17]. For example, immunosuppression is associated with oropharyngeal Candida disease, but not vaginal candidiasis, whereas antibiotic use leading to Candida outgrowth and host sensitization can lead to the transition from asymptomatic carriage to chronic vaginal inflammation, that is, ‘yeast infection’ [18].
Although carriage is usually asymptomatic, anti-Candida antibodies are often detected in normal individuals, indicating the elaboration of adaptive immune responses to commensal Candida [19]. It is unclear how C. albicans remains at mucosal surfaces in the face of adaptive immunity. However, because Candida can produce immunomodulatory compounds such as oxylipins, it is tempting to speculate that this commensal organism modulates immune responses to favor its persistence [20]. Modulation of host responses during asymptomatic colonization may in itself lead to deleterious consequences.
Several C. albicans factors have been implicated in mediated interactions with the bacterial microbiota. Members of the agglutinin-like sequence family of adhesins mediate C. albicans aggregation with other bacteria and yeasts [21]. Another factor that may influence C. albicans interactions with bacteria is farnesol, a quorum sensing molecule. Farnesol represses hyphal growth and early biofilm formation, is required for virulence during disseminated infection and has similar effects on other bacterial pathogens, inhibiting biofilm growth and formation (reviewed in [22]). This may be a strategy used by C. albicans growing in biofilms to disperse under conditions of stress, which may also lead to dispersal of bacteria in the polymicrobial biofilm. In addition to cross-kingdom adhesion and quorum sensing, metabolic changes in the environment (pH, metabolic substrate production, inhibitor production, nutrient sensing/sequestering, etc.) and indirect activity on the host response are mechanisms by which Candida and other fungi interact with bacteria in the microbiome to regulate fungal levels and host responses to fungal colonization [23].
Fungal microbiome of the skin
The diversity of the microbiome of human skin is an area of intense investigation because changes in the microbiome may be associated with the development or chronicity of many dermatologic conditions or diseases [24]. Cultivation techniques suggested that the fungal microbiome of healthy skin was limited to few genera, mainly Malassezia and Candida. C. albicans can be carried on the skin, with carriage rates ranging from 50 to 65% among hospital workers [25]. Skin sensitization to C. albicans is also strongly correlated with development of atopic dermatitis (AD) [26]. Malassezia are lipophilic yeast, classified in the phylum Basidiomycota, and this genus currently includes 14 species isolated from healthy and diseased human and animal skin [27]. Malassezia spp. are associated with a variety of skin diseases including psoriasis, dandruff, AD, or eczema, seborrheic dermatitis, and pityriasis versicolor [27]. Molecular methods have revealed that a diverse fungal microbiome exists on the skin and the composition differs between healthy and diseased sites. Overall, these methods have revealed that the skin harbors a diverse fungal community comprised colonizers such as Malassezia that increase in numbers in damaged skin, as well as other less numerous colonizers and other transient passengers derived from the environment.
Psoriasis can be triggered and/or exacerbated by a variety of environmental factors, including bacterial and fungal members of the skin microbiome [28]. In studies by Blaser and colleagues, the fungal microbiota of healthy skin and psoriatic lesions was studied through clone library construction and sequencing from different skin sites, using a set of broad-range 18S ribosomal DNA (rDNA) primers and another set specifically targeting the 5.8S rDNA/ITS2 of Malassezia [29]. On the healthy skin of one subject, >98% of the clones could be grouped into two Basidiomycota phylotypes that were related to but distinct from Malassezia furfur while two psoriatic lesion sites from the same subject contained the same phylotypes but at different ratios. The remainder of the study used Malassezia-specific 5.8S/ITS2 primers to analyze a large number of samples from over 20 subjects. Five Malassezia species and four unknown phylotypes were identified. Most notably, the species distribution was largely subject-specific, but conserved throughout different sites of healthy skin. Overall, this study demonstrated the predominance of Malassezia organisms on healthy human skin, host-specific variation in colonization, relative stability over time on healthy skin, and no consistent patterns of Malassezia colonization that differentiated psoriatic skin from healthy skin.
Another study examined the changes in the fungal microbiota of the scalp that accompany dandruff [30]. The investigators used pyrosequencing of 26S rRNA gene libraries derived from healthy and dandruff-afflicted scalp samples. While fungi of the Ascomycota dominated in both healthy and dandruff patients, fungi of the Basidiomycota phyla (which includes Malassezia) were significantly increased in dandruff-afflicted scalps. Specific genera that were increased in patients with severe dandruff included Filobasidium, which jumped to 94% of the total basiomycetes, and Malassezia (5%), which represents a twofold increase over healthy samples. By contrast, healthy scalps were dominated by basiomycete fungi of the genus Cryptococcus. Among the ascomycetes, Acremonium spp. dominated in both healthy individuals and dandruff patients. However, Didymella was significantly increased in healthy samples, whereas Penicillium spp. increased in dandruff patients, particularly in severe cases. This study confirms the previously reported association of Malassezia spp. with dandruff, but also reveals the potential for Filobasidium in this disease.
AD or eczema is a multifactorial allergic disease that is characterized by a variety of symptoms, notably intense itching, edema, and erythema in affected skin sites. The pathophysiology of the disease involves barrier disruption and immune dysregulation, although the cause and effect sequence is debatable. The fungal microbiome of the skin could participate in exacerbating atopic reactions in patients by shedding allergens, secreting compounds that actively modulate host responses, and/or producing enzymes that damage host cells. Malassezia spp. have long been associated with AD, and many studies have focused on this genus in disease [27]. The overall skin fungal microbiota of patients with mild, moderate, or severe AD was compared with healthy subjects using rRNA clone library sequencing [31]. Malassezia spp. were predominant in all groups; however, the diversity of non-Malassezia yeast microbiota was increased in AD patients. Notably, C. albicans, Cryptococcus diffluans, and Cryptococcus liquifaciens were detected in all AD samples, but were seldom detected in healthy samples. In particular, C. albicans was present in only one healthy subject. AD is also more highly associated with sensitization to C. albicans, compared with other fungal species [26]. Increased Candida-specific IgE also correlates with severity of disease [32]. These findings suggest that C. albicans could be an unforeseen contributor to AD pathogenesis and supports a need for further fungal microbiota analysis on a larger scale.
Microbial biofilms formed from microbiota members contribute to the delayed healing of chronic wounds and a survey of clinical specimens taken from chronic wounds revealed 23% were positive for fungal species [33]. The most abundant fungi were yeasts of the genus Candida; however, Malessezia, Curvularia, Aureobasidium, Cladosporium, Ulocladium, Engodontium, and Trichtophyton were also found to be prevalent components of these polymicrobial infections. Quantification of bacteria versus fungi in these chronic wounds demonstrated that fungi contributed to >50% of the microbial burden in the majority of the wounds. In several cases, addition of antifungal drugs to the treatment regimen resulted in gradual healing of the wound site. This clinical survey demonstrates that the incidence of fungal pathogens in wound biofilm infection is more significant than previously reported and may contribute to the extremely recalcitrant nature of many wounds to antibacterial agents.
Fungal microbiome of the mouth and lungs
The human oral microbiome is comprised at least 700 bacterial taxa [34]. These microbial communities are arranged into surface-localized communities that vary considerably in composition according to sites of establishment. Although the mouth clearly harbors fungi, much remains unknown about the oral fungal communities. Ghannoum and colleagues performed the most comprehensive study to date on the diversity of the fungal microbiome at any human body site, analyzing the fungal microbiome of the mouth [10]. In this study, they found that the distribution of fungal species in the mouth varied greatly between different individuals. In about 20% of the study participants, their oral microbiome included at least one of the following four most common genera of pathogenic fungi: Candida, Aspergillus, Fusarium, and Cryptococcus. The most abundant Candida species were C. albicans (in 40% of the subjects), C. parapsilosis (15%), C. tropicalis (15%), C. khmerensis (5%), and C. metapsilosis (5%). Also of note in this study, nonculturable fungi represented almost 40% of the fungi identified in the oral cavity of healthy individuals. One of the most interesting aspects of this study was that 60 nonpathogenic fungal genera were identified in subjects that are ubiquitous environmental microbes. The presence of these microbes in the oral cavities of healthy individuals was not necessarily surprising, because they are most likely acquired from food and mouth breathing, but the observation that transient colonization by environmental fungi may occur in the oral cavity (and upper airways) has potential implications for hypersensitivity diseases.
The lungs, previously believed to be sterile when healthy, clearly harbor a low level bacterial microbiome that changes during disease [35]. To date, little is known about the fungal microbiota of the lungs, with the exception of Pneumocystis spp. (described below). In a recent study by Charlson et al., the fungal microbiome of the mouth and lungs in select healthy and lung transplant recipients was analyzed by ITS-based pyrosequencing [7]. The fungal distribution in the oral wash of healthy subjects was similar to the previously published study described above [10]. In the lung transplant recipients, the fungal microbiome of the oral cavity was dominated by Candida, likely owing to antibiotic and immunosuppressant use [7]. In the bronchoalveolar lavage of healthy volunteers, there was minimal fungal ITS amplification. By contrast, the bronchoalveolar lavage from lung transplant recipients showed a markedly different pattern, with detectable fungi of either Candida spp., Aspergillus spp., or Cryptococcus spp., oftentimes at significant levels. Because all of the transplant recipients had been treated with antibiotics and immunosuppressants, this first study of the lung fungal microbiome supports the notion that host defense, and perhaps some sort of bacterial microbiome-mediated resistance mechanisms, play a major role in keeping fungal colonization extremely low in the lungs.
One notable member of the fungal microbiome in humans and animals are fungi of the Pneumocystis genus [36]. This organism was for a long time considered a unicellular protist. However, the past three decades of research on this organism has firmly established it as a fungus of the Ascomycota phylum. This fungus is highly unusual among medically important fungi in that it appears to have co-evolved with its host and lacks a number of key biosynthetic pathways thereby precluding growth outside its host. Thus, Pneumocystis derived from humans will not grow in mice and vice versa. The preferred growth niche in the host is the lungs and new molecular surveys are revealing that it is carried at low levels, even in healthy individuals. This fungus can be spread from individual to individual through airborne transmission, but it can also cause pneumonia following overgrowth in immunocompromised hosts. Pneumocystis has also been implicated as a cofactor in a number of chronic pulmonary inflammatory diseases. Thus, the fungus Pneumocystis appears to exist as a very low level commensal in the lung microbiome when the host is healthy and becomes pathogenic when the host becomes immunocompromised.
Fungal microbiome of the GI tract
Despite its harsh environment, the stomach harbors a microbiome that can include Lactobacillus, Helicobacter, and Candida spp. [37–40]. The low pH environment of the stomach selects for colonization by microbes with a high degree of acid tolerance, such as Candida. Within the stomach, Lactobacillus growth can antagonize colonization by Candida (Box 1). Some Candida species, such as C. pintolopesii, can exist in the murine microbiome without inducing inflammation [38]. Erosions and ulcerations of the mucosal surfaces in the stomach and intestinal tract can also favor the growth of select microbial populations, including C. albicans [41]. In humans, gastric ulcers associated with C. albicans colonization is a well-documented condition, although generally unappreciated in terms of etiologic agents of gastric ulceration (discussed in [40]). Candida colonization of the GI tract of mice can drive allergic sensitization to food antigens by affecting the mucosal barrier [42].
Box 1. Lactobacillus–Candida antagonism.
The indigenous microbiota of the GI tract is effective at preventing fungi, such as Candida albicans, from high-level colonization and disease [56–63] and germfree mice, lacking the indigenous microbiota, are highly susceptible to Candida colonization [64]. Mouse models of candidiasis demonstrate that disturbance of the microbiota or immunosuppression are necessary to promote Candida colonization. During gastric colonization, Candida spp. can induce inflammation or exist as noninflammatory commensal organisms [38,65,66]. Although the exact mechanisms still remain to be determined, it has been suggested that lactobacilli are critical [59,60]. Results from in vitro studies implicate the bacterial microbiome in blocking yeast adhesion to the epithelium and producing inhibitor substances (such as volatile fatty acids and secondary bile acids) that can reduce C. albicans adhesion, hyphal transformation, and invasion [59,60,66–69]. However, Lactobacillus– Candida antagonism can be a two-way process whereby the presence of Candida can prevent the regrowth of Lactobacillus after antibiotics [40,44]. The effect may be more widespread than effects on Lactobacillus because recent data suggests that yeast, even as numerically inferior microbes in the intestinal microbiome, have the potential to exert marked effects on overall bacterial community reassembly in the intestine after antibiotics without inducing intestinal inflammation [44].
Despite the extensive literature on the bacterial microbiome of the intestinal tract, little is known about the fungal microbiome of the intestinal tract. As discussed earlier in this review, Candida spp. can clearly grow in the intestines, coexist with the bacterial microbiome in the intestines, bloom during antibiotic perturbations of the microbiome, and colonize inflamed mucosa of the intestine. But what about other fungal genera? In one study, using mice from both a specific pathogen-free (SPF) source and from a colony with a restricted bacterial flora (RF mice), a diverse and abundant fungal microbiome was detected by molecular techniques [43]. All four major fungal phyla were detected (Ascomycota, Basidiomycota, Chytridiomycota, and Zygomycota) and there were clear differences in the species distribution within the fungal microbiome between the two types of mice. Of note in this study was the lack of Candida spp. detected, as well as a lack of culture data to compare with the molecular methods. SPF mice routinely lack culturable fungi and even treatment with a broad spectrum antibiotic, such as cefoperazone, rarely results in the culture of fungi from intestinal samples on fungal media (Sabouraud dextrose agar) [40,44,45]. In a recently published study with Dectin-1 knockout and wild type mice, high levels of indigenous fungi were reported in wild type laboratory mice, especially C. tropicalis (a more virulent Candida species for damaged mucosa), which accounted for 65% of the fungi detected by ITS1–2 amplicon pyrosequencing [46]. The total fungal loads were estimated to be 5–7% of the fecal material by staining and were easily visualized by immunohistology on mucosal surfaces, although no culture data was presented for C. tropicalis, which can easily be cultured and identified on Chromagar Candida. Other studies in mice and humans, either healthy or on antibiotics, have not reported anywhere near that level of colonization with C. troplicalis or other fungi [1,43,44,47–49]. The metagenomic analysis performed by the MetaHIT group on fecal specimens collected from 124 healthy, overweight, and obese individuals (including some with inflammatory bowel disease) from Denmark and Spain reported that only 0.1% of the genes in fecal material were of eukaryotic or viral origin, which was consistent with previous reports of fungal 18s rDNA signatures accounting for only 0.03% of the fecal microbiota [1,47]. Thus, as can be summarized from all of these studies, the study of the fungal microbiome in the intestinal tract is in its infancy and much remains to be determined.
Concluding remarks and future perspectives
There is ever-increasing evidence of a significant role for the microbiome in immune regulation, chronic inflammatory diseases, metabolism, and other physiologic processes, including recovery from antibiotics (Box 1). The research to date has focused on the bacterial microbiome; however, clinical and experimental studies are beginning to highlight a role for fungi in these processes. Recent studies have demonstrated that the bacterial GI community reassembly after broad spectrum antibiotics can be altered if the fungal community blooms in the GI tract [40,44] and this can impact the development of allergic airway disease [45,50] (Figure 4). Furthermore, patients with severe asthma with fungal sensitization (SAFS) are often sensitized to C. albicans and benefit from antifungal therapy [51]. Colonization of mice with C. albicans can result in dietary antigen leak from the stomach [42]. Crohn’s disease is associated with a microbiome dysbiosis and the development of antibodies against members of the microbiome [52,53]. This includes anti-Saccharomyces cerevisiae antibodies (ASCA), which have been shown to be reactive to an in vivo-expressed epitope on Candida species, as well as baker’s yeast [54]. A recent study using mice that are missing the C-type lectin receptor Dectin-1 demonstrated that these mice had an increased susceptibility to chemically induced [dextran sulfate sodium (DSS)] colitis and this was abrogated by fluconazole and could be exacerbated by repeated oral delivery of C. tropicalis [46]. This was consistent with the report that C. albicans could exacerbate DSS-induced colitis [55] but demonstrated that an indigenous Candida population could drive disease.
Figure 4.

Hypothetical representation of the effect of the presence and absence of a fungal outgrowth that modulates bacterial community reassembly. Reassembly of the bacterial microbiome after antibiotic therapy is an ordered process (TA→T1→T2→T0). Emerging evidence suggests that reassembly may be impacted by the bloom of specific fungal species when colonization resistance is destroyed. This figure provides an illustration of an ecological modeling (ordination) of such a process, in which the bacterial community structure undergoes a different reassembly process (TA→T1f→T2f→T0) in the presence of a fungal bloom. In this example, the differences in bacterial community structure (membership, richness, and evenness) between the TA→T1→T2→T0 path and the TA→T1f→T2f→T0 path could manifest as secondary changes in microbiome-mediated physiologic processes such as immune regulation or metabolism. Abbreviation: T, timepoint.
Evidence is clearly beginning to accumulate that the fungal members of the rare biosphere of the microbiome may play a greater role than previously ascribed in regulation of mucosal health, especially during blooms when the bacterial microbiome is disturbed. More research is needed to catalog the mycobiome in health and disease and also to view these communities through the lens of microbial ecology in terms of membership, presence/absence, levels, and cross-kingdom microbial interactions. Additionally, once we know about membership, what about function? The metabolic maps and genomic databases for fungi lag significantly behind those for bacteria. Future studies will begin to unravel the questions raised here about the levels and composition of indigenous fungi (the mycobiome) in the mouth, lungs, skin, and GI tract in health and during disease, and the mechanisms whereby they may contribute to (or protect from) disease.
Acknowledgments
G.B.H. is supported in part by National Institutes of Health (NIH) grants R01HL114447, U01HL98961, and U19AI090871. M.C.N. is supported in part by NIH grants R01AI072406 and R01DE022069.
References
- 1.Qin J, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464:59–65. doi: 10.1038/nature08821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Dethlefsen L, et al. The pervasive effects of an antibiotic on the human gut microbiota, as revealed by deep 16S rRNA sequencing. PLoS Biol. 2008;6:e280. doi: 10.1371/journal.pbio.0060280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sogin ML, et al. Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc. Natl. Acad. Sci. U.S.A. 2006;103:12115–12120. doi: 10.1073/pnas.0605127103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hajishengallis G, et al. Low-abundance biofilm species orchestrates inflammatory periodontal disease through the commensal microbiota and complement. Cell Host Microbe. 2011;10:497–506. doi: 10.1016/j.chom.2011.10.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hatoum R, et al. Antimicrobial and probiotic properties of yeasts: from fundamental to novel applications. Front. Microbiol. 2012;3:421. doi: 10.3389/fmicb.2012.00421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cassone M, et al. Outbreak of Saccharomyces cerevisiae subtype boulardii fungemia in patients neighboring those treated with a probiotic preparation of the organism. J. Clin. Microbiol. 2003;41:5340–5343. doi: 10.1128/JCM.41.11.5340-5343.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Charlson ES, et al. Lung-enriched organisms and aberrant bacterial and fungal respiratory microbiota following lung transplant. Am. J. Respir. Crit. Care Med. 2012;186:536–545. doi: 10.1164/rccm.201204-0693OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Delhaes L, et al. The airway microbiota in cystic fibrosis: a complex fungal and bacterial community–implications for therapeutic management. PLoS ONE. 2012;7:e36313. doi: 10.1371/journal.pone.0036313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Gardes M, Bruns TD. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993;2:113–118. doi: 10.1111/j.1365-294x.1993.tb00005.x. [DOI] [PubMed] [Google Scholar]
- 10.Ghannoum MA, et al. Characterization of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog. 2010;6:e1000713. doi: 10.1371/journal.ppat.1000713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lindsley MD, et al. Rapid identification of dimorphic and yeastlike fungal pathogens using specific DNA probes. J. Clin. Microbiol. 2001;39:3505–3511. doi: 10.1128/JCM.39.10.3505-3511.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kleinegger CL, et al. Frequency, intensity, species, and strains of oral Candida vary as a function of host age. J. Clin. Microbiol. 1996;34:2246–2254. doi: 10.1128/jcm.34.9.2246-2254.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Soll DR, et al. Genetic dissimilarity of commensal strains of Candida spp. carried in different anatomical locations of the same healthy women. J. Clin. Microbiol. 1991;29:1702–1710. doi: 10.1128/jcm.29.8.1702-1710.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Blaschke-Hellmessen R. Habitats for Candida in medical and hygienic respects. Mycoses. 1999;42(Suppl. 1):22–29. doi: 10.1111/j.1439-0507.1999.tb04522.x. in German. [DOI] [PubMed] [Google Scholar]
- 15.Odds FC. Ecology and epidemiology of Candida species. Zentralbl. Bakteriol. Mikrobiol. Hyg. 1984;A 257:207–212. [PubMed] [Google Scholar]
- 16.Mayer FL, et al. Candida albicans pathogenicity mechanisms. Virulence. 2013;4:119–128. doi: 10.4161/viru.22913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Fidel PL., Jr Distinct protective host defenses against oral and vaginal candidiasis. Med. Mycol. 2002;40:359–375. [PubMed] [Google Scholar]
- 18.Yano J, et al. Cytokines in the host response to Candida vaginitis: identifying a role for non-classical immune mediators, S100 alarmins. Cytokine. 2012;58:118–128. doi: 10.1016/j.cyto.2011.11.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Faux JA, et al. A comparison of specific IgG antibody levels to the cell wall mannan of Candida albicans in normal individuals and in patients with primary antibody deficiency. J. Immunol. Methods. 1992;153:167–172. doi: 10.1016/0022-1759(92)90319-o. [DOI] [PubMed] [Google Scholar]
- 20.Noverr MC, et al. Pathogenic yeasts Cryptococcus neoformans and Candida albicans produce immunomodulatory prostaglandins. Infect. Immun. 2001;69:2957–2963. doi: 10.1128/IAI.69.5.2957-2963.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Klotz SA, et al. Candida albicans Als proteins mediate aggregation with bacteria and yeasts. Med. Mycol. 2007;45:363–370. doi: 10.1080/13693780701299333. [DOI] [PubMed] [Google Scholar]
- 22.De Sordi L, Muhlschlegel FA. Quorum sensing and fungal-bacterial interactions in Candida albicans: a communicative network regulating microbial coexistence and virulence. FEMS Yeast Res. 2009;9:990–999. doi: 10.1111/j.1567-1364.2009.00573.x. [DOI] [PubMed] [Google Scholar]
- 23.Peleg AY, et al. Medically important bacterial-fungal interactions. Nat. Rev. Microbiol. 2010;8:340–349. doi: 10.1038/nrmicro2313. [DOI] [PubMed] [Google Scholar]
- 24.Kong HH, Segre JA. Skin microbiome: looking back to move forward. J. Invest. Dermatol. 2012;132:933–939. doi: 10.1038/jid.2011.417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Brunetti L, et al. Surveillance of nosocomial infections: a preliminary study on yeast carriage on hands of healthcare workers. J. Prev. Med. Hyg. 2008;49:63–68. [PubMed] [Google Scholar]
- 26.Chang FY, et al. Analysis of the serum levels of fungi-specific immunoglobulin E in patients with allergic diseases. Int. Arch. Allergy Immunol. 2011;154:49–56. doi: 10.1159/000319208. [DOI] [PubMed] [Google Scholar]
- 27.Gaitanis G, et al. The Malassezia genus in skin and systemic diseases. Clin. Microbiol. Rev. 2012;25:106–141. doi: 10.1128/CMR.00021-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fry L, Baker BS. Triggering psoriasis: the role of infections and medications. Clin. Dermatol. 2007;25:606–615. doi: 10.1016/j.clindermatol.2007.08.015. [DOI] [PubMed] [Google Scholar]
- 29.Paulino LC, et al. Molecular analysis of fungal microbiota in samples from healthy human skin and psoriatic lesions. J. Clin. Microbiol. 2006;44:2933–2941. doi: 10.1128/JCM.00785-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Park HK, et al. Characterization of the fungal microbiota (mycobiome) in healthy and dandruff-afflicted human scalps. PLoS ONE. 2012;7:e32847. doi: 10.1371/journal.pone.0032847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhang E, et al. Characterization of the skin fungal microbiota in patients with atopic dermatitis and in healthy subjects. Microbiol. Immunol. 2011;55:625–632. doi: 10.1111/j.1348-0421.2011.00364.x. [DOI] [PubMed] [Google Scholar]
- 32.Sonesson A, et al. Sensitization to skin-associated microorganisms in adult patients with atopic dermatitis is of importance for disease severity. Acta Derm. Venereol. 2012 doi: 10.2340/00015555-1465. http://dx.doi.org/10.2340/00015555-1465. [DOI] [PubMed] [Google Scholar]
- 33.Dowd SE, et al. Survey of fungi and yeast in polymicrobial infections in chronic wounds. J. Wound Care. 2011;20:40–47. doi: 10.12968/jowc.2011.20.1.40. [DOI] [PubMed] [Google Scholar]
- 34.Jenkinson HF. Beyond the oral microbiome. Environ. Microbiol. 2011;13:3077–3087. doi: 10.1111/j.1462-2920.2011.02573.x. [DOI] [PubMed] [Google Scholar]
- 35.Dickson RP, et al. The role of the microbiome in lung disease. Exp. Rev. Respir. Med. 2013;7 doi: 10.1586/ers.13.24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chabe M, et al. Pneumocystis: from a doubtful unique entity to a group of highly diversified fungal species. FEMS Yeast Res. 2011;11:2–17. doi: 10.1111/j.1567-1364.2010.00698.x. [DOI] [PubMed] [Google Scholar]
- 37.Bik EM, et al. Molecular analysis of the bacterial microbiota in the human stomach. Proc. Natl. Acad. Sci. U.S.A. 2006;103:732–737. doi: 10.1073/pnas.0506655103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Savage DC, Dubos RJ. Localization of indigenous yeast in the murine stomach. J. Bacteriol. 1967;94:1811–1816. doi: 10.1128/jb.94.6.1811-1816.1967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Artwohl J, et al. Population changes of indigenous murine Candida pintolopesii under various experimental conditions and routes of inoculation. Appl. Environ. Microbiol. 1988;54:2371–2374. doi: 10.1128/aem.54.10.2371-2374.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mason KL, et al. Interplay between the gastric bacterial microbiota and Candida albicans during postantibiotic recolonization and gastritis. Infect. Immun. 2012;80:150–158. doi: 10.1128/IAI.05162-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kumamoto CA. Inflammation and gastrointestinal Candida colonization. Curr. Opin. Microbiol. 2011;14:386–391. doi: 10.1016/j.mib.2011.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Yamaguchi N, et al. Gastrointestinal Candida colonisation promotes sensitisation against food antigens by affecting the mucosal barrier in mice. Gut. 2006;55:954–960. doi: 10.1136/gut.2005.084954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Scupham AJ, et al. Abundant and diverse fungal microbiota in the murine intestine. Appl. Environ. Microbiol. 2006;72:793–801. doi: 10.1128/AEM.72.1.793-801.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Mason KL, et al. Candida albicans and bacterial microbiota interactions in the cecum during recolonization following broad-spectrum antibiotic therapy. Infect. Immun. 2012;80:3371–3380. doi: 10.1128/IAI.00449-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Noverr MC, et al. Role of antibiotics and fungal microbiota in driving pulmonary allergic responses. Infect. Immun. 2004;72:4996–5003. doi: 10.1128/IAI.72.9.4996-5003.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Iliev ID, et al. Interactions between commensal fungi and the C-type lectin receptor Dectin-1 influence colitis. Science. 2012;336:1314–1317. doi: 10.1126/science.1221789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ott SJ, et al. Fungi and inflammatory bowel diseases: alterations of composition and diversity. Scand. J. Gastroenterol. 2008;43:831–841. doi: 10.1080/00365520801935434. [DOI] [PubMed] [Google Scholar]
- 48.de Repentigny L, et al. Gastrointestinal colonization and systemic dissemination by Candida albicans and Candida tropicalis in intact and immunocompromised mice. Infect. Immun. 1992;60:4907–4914. doi: 10.1128/iai.60.11.4907-4914.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Wingard JR, et al. Pathogenicity of Candida tropicalis and Candida albicans after gastrointestinal inoculation in mice. Infect. Immun. 1980;29:808–813. doi: 10.1128/iai.29.2.808-813.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Noverr MC, et al. Development of allergic airway disease in mice following antibiotic therapy and fungal microbiota increase: role of host genetics, antigen, and interleukin-13. Infect. Immun. 2005;73:30–38. doi: 10.1128/IAI.73.1.30-38.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.O’Driscoll BR, et al. Comparison of skin prick tests with specific serum immunoglobulin E in the diagnosis of fungal sensitization in patients with severe asthma. Clin. Exp. Allergy. 2009;39:1677–1683. doi: 10.1111/j.1365-2222.2009.03339.x. [DOI] [PubMed] [Google Scholar]
- 52.Seow CH, et al. Novel anti-glycan antibodies related to inflammatory bowel disease diagnosis and phenotype. Am. J. Gastroenterol. 2009;104:1426–1434. doi: 10.1038/ajg.2009.79. [DOI] [PubMed] [Google Scholar]
- 53.Vernier G, et al. Relevance of serologic studies in inflammatory bowel disease. Curr. Gastroenterol. Rep. 2004;6:482–487. doi: 10.1007/s11894-004-0070-x. [DOI] [PubMed] [Google Scholar]
- 54.Standaert-Vitse A, et al. Candida albicans is an immunogen for anti-Saccharomyces cerevisiae antibody markers of Crohn’s disease. Gastroenterology. 2006;130:1764–1775. doi: 10.1053/j.gastro.2006.02.009. [DOI] [PubMed] [Google Scholar]
- 55.Jawhara S, et al. Colonization of mice by Candida albicans is promoted by chemically induced colitis and augments inflammatory responses through galectin-3. J. Infect. Dis. 2008;197:972–980. doi: 10.1086/528990. [DOI] [PubMed] [Google Scholar]
- 56.Hummel RP, et al. Inhibition of Candida albicans by Escherichia coli in vitro and in the germfree mouse. J. Surg. Res. 1973;15:53–58. doi: 10.1016/0022-4804(73)90163-7. [DOI] [PubMed] [Google Scholar]
- 57.Kennedy MJ. Inhibition of Candida albicans by the anaerobic oral flora of mice in vitro . Sabouraudia. 1981;19:205–208. [PubMed] [Google Scholar]
- 58.Kennedy MJ, Volz PA. Dissemination of yeasts after gastrointestinal inoculation in antibiotic-treated mice. Sabouraudia. 1983;21:27–33. doi: 10.1080/00362178385380051. [DOI] [PubMed] [Google Scholar]
- 59.Kennedy MJ, Volz PA. Ecology of Candida albicans gut colonization: inhibition of Candida adhesion, colonization, and dissemination from the gastrointestinal tract by bacterial antagonism. Infect. Immun. 1985;49:654–663. doi: 10.1128/iai.49.3.654-663.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Kennedy MJ, Volz PA. Effect of various antibiotics on gastrointestinal colonization and dissemination by Candida albicans . Sabouraudia. 1985;23:265–273. doi: 10.1080/00362178585380391. [DOI] [PubMed] [Google Scholar]
- 61.van der Waaij D. Colonization resistance of the digestive tract–mechanism and clinical consequences. Nahrung. 1987;31:507–517. doi: 10.1002/food.19870310551. [DOI] [PubMed] [Google Scholar]
- 62.van der waaij D, Berghuis JM. Determination of the colonization resistance of the digestive tract of individual mice. J. Hyg. (Lond.) 1974;72:379–387. doi: 10.1017/s0022172400023615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Van der Waaij D, Van der Waaij BD. The colonization resistance of the digestive tract in different animal species and in man; a comparative study. Epidemiol. Infect. 1990;105:237–243. doi: 10.1017/s0950268800047841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Naglik JR, et al. Animals models of mucosal Candida infection. FEMS Microbiol. Lett. 2008;283:129–139. doi: 10.1111/j.1574-6968.2008.01160.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Helstrom PB, Balish E. Effect of oral tetracycline, the microbial flora, and the athymic state on gastrointestinal colonization and infection of BALB/c mice with Candida albicans . Infect. Immun. 1979;23:764–774. doi: 10.1128/iai.23.3.764-774.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Savage DC. Microbial interference between indigenous yeast and lactobacilli in the rodent stomach. J. Bacteriol. 1969;98:1278–1283. doi: 10.1128/jb.98.3.1278-1283.1969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Noverr MC, Huffnagle GB. Regulation of Candida albicans morphogenesis by fatty acid metabolites. Infect. Immun. 2004;72:6206–6210. doi: 10.1128/IAI.72.11.6206-6210.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Wagner RD, et al. Probiotic effects of feeding heat-killed Lactobacillus acidophilus and Lactobacillus casei to Candida albicans-colonized immunodeficient mice. J. Food Prot. 2000;63:638–644. doi: 10.4315/0362-028x-63.5.638. [DOI] [PubMed] [Google Scholar]
- 69.Yamaguchi N, et al. Gastric colonization of Candida albicans differs in mice fed commercial and purified diets. J. Nutr. 2005;135:109–115. doi: 10.1093/jn/135.1.109. [DOI] [PubMed] [Google Scholar]