

Summary
Spermatogenesis is a complex process that generates haploid germ cells or spores and implements meiosis, a succession of two special cell divisions that are required for homologous chromosome segregation. During prophase to the first meiotic division, homologous recombination (HR) repairs Spo11-dependent DNA double-strand breaks (DSBs) in the presence of telomere movements to allow for chromosome pairing and segregation at the meiosis I division. In contrast to HR, non-homologous end joining (NHEJ), the major DSB repair mechanism during the G1 cell cycle phase, is downregulated during early meiotic prophase. At somatic mammalian telomeres, the NHEJ factor Ku70/80 inhibits HR, as does the Rap1 component of the shelterin complex. Here, we investigated the role of Ku70 and Rap1 in meiotic telomere redistribution and genome protection in spermatogenesis by studying single and double knockout mice. Ku70−/− mice display reduced testis size and compromised spermatogenesis, whereas meiotic telomere dynamics and chromosomal bouquet formation occurred normally in Ku70−/− and Ku70−/−Rap1Δ/Δ knockout spermatocytes. Elevated mid-preleptotene frequencies were associated with significantly increased DNA damage in Ku-deficient B spermatogonia, and in differentiated Sertoli cells. Significantly elevated levels of γH2AX foci in Ku70−/− diplotene spermatocytes suggest compromised progression of DNA repair at a subset of DSBs. This might explain the elevated meiotic metaphase apoptosis that is present in Ku70-deficient stage XII testis tubules, indicating spindle assembly checkpoint activation. In summary, our data indicate that Ku70 is important for repairing DSBs in somatic cells and in late spermatocytes of the testis, thereby assuring the fidelity of spermatogenesis.
Key words: Bouquet formation, DNA damage, Ku70, Meiosis, Mid-preleptotene, Recombination, Rap1, Sertoli cell, NHEJ, Spermatogenesis, Telomere attachment, Shelterin
Introduction
Meiosis is the central process of spermatogenesis by which the genome is bisected to generate haploid gametes. Prior to genome haploidization, homologous chromosomes pair, recombine and separate from each other during the first of two successive divisions that lack an intervening DNA replication (reviewed by Gerton and Hawley, 2005). At the onset of first meiotic prophase, chromosomes receive Spo11-dependent DNA double-strand breaks (DSBs) that are repaired by homologous recombination (HR) to promote crossing over, which ensures homolog separation during the meiosis I division (reviewed by Bannister and Schimenti, 2004; Keeney, 2008). HR in meiosis is facilitated by downregulation of the error-prone non-homologous end joining (NHEJ) DSB repair pathway at the onset and during the early substages of first meiotic prophase I (Goedecke et al., 1999). The NHEJ DSB repair pathway dominates in G0 and G1 phases of the cell cycle when homologous chromatids are not available. NHEJ involves binding of the Ku70/80 heterodimer to broken DNA ends, which are then ligated back together or to other DSB ends in a mutagenic process (for review, see Lieber, 2010).
In mice, Ku70 (also known as Xrcc6) and Ku80 (also known as Xrcc5) disruption leads to deficiency in NHEJ DNA repair, early cellular senescence and compromised growth (Gu et al., 1997; Nussenzweig et al., 1997; Ouyang et al., 1997). In somatic cells, Ku70 is required for full proliferation capacity (Gu et al., 1997; Nussenzweig et al., 1996) and acts as a switch between the HR and NHEJ DSB repair pathways. NHEJ is downregulated in early meiotic cells in which homology-directed repair (HDR) dominates (Goedecke et al., 1999). Ku proteins are again expressed in late pachytene and diplotene spermatocytes, and they mediate repair proficiency in somatic cells of the testis, such as spermatogonia and Sertoli cells (Ahmed et al., 2007).
Besides the well-known function in DSB repair, end-binding Ku proteins are components of the ends of linear chromosomes (Bailey et al., 1999; Celli et al., 2006; Hsu et al., 1999), the telomeres that ensure genomic stability (for review, see Bertuch and Lundblad, 2006; de Lange, 2005). In the mammalian somatic cell nucleus, mammalian telomeres are distributed without an apparent order (Ludérus et al., 1996; Vourc'h et al., 1993), whereas their meiotic counterparts attach to the nuclear envelope early in meiosis (Scherthan et al., 1996) through an interaction with a SUN/KASH protein complex in the meiotic nuclear envelope (Ding et al., 2007; Hiraoka and Dernburg, 2009; Morimoto et al., 2012). Meiotic telomeres move to cluster near the centrosome during the early substages of first meiotic prophase (bouquet formation) (for a review, see Scherthan, 2006). In budding yeast, cells deficient for Ku (Hdf1/2) function display a disturbed three-dimensional telomere location in the vegetative nucleus (Laroche et al., 1998), which leads to altered telomere dynamics at the onset of first meiotic prophase (Scherthan and Trelles-Sticken, 2008). However, the consequences of Ku disruption for mammalian meiosis are largely unexplored. As Ku and the mammalian telomere-interacting protein RAP1 have been shown to inhibit HDR at telomeres (Celli et al., 2006; Sfeir et al., 2010), we made use of Ku70 Rap1 double knockout mice (Sfeir et al., 2010) to determine the combined and single contribution of Ku70 and Rap1 to meiotic telomere dynamics and fidelity of spermatogenesis. Furthermore, we studied the consequences of the absence of these two proteins for the telomere clustering that has been observed in somatic Sertoli cells (Scherthan et al., 2000).
Results
Ku70 mutation reduces spermatogenetic fidelity
Ku knockout mice have been reported to display overall reduced body and organ size and a reduced life span owing to compromised cellular proliferation capacity (Gu et al., 1997; Holcomb et al., 2007; Nussenzweig et al., 1996). Investigation of Ku70−/− and Ku70−/−Rap1Δ/Δ spermatogenesis revealed, in accordance with the literature, a significantly reduced testis size in all Ku70−/− and Ku70−/−Rap1Δ/Δ animals compared with heterozygous, Rap1Δ/Δ and wild-type testes (Fig. 1A,B). Investigating spermatogenesis in more detail using DAPI or Hematoxylin and Eosin (H&E) staining revealed an overall normal tubule histology in wild-type and heterozygous testes (Fig. 1C,D), whereas sperm formation and release was compromised in Ku70−/− and Ku70−/−Rap1Δ/Δ stage VI–VII testis tubules (Fig. 1E,F). In agreement, we noted almost empty epididymides in all Ku70−/− mice (Fig. 1H,I), demonstrating reduced fidelity of spermatogenesis in the absence of Ku70. These observations indicate that Ku70 is epistatic to Rap1, and that Ku deficiency is responsible for reduced testis size and compromised spermatogenesis in the Ku70−/−Rap1Δ/Δ condition.
Fig. 1.

Spermatogenesis in Ku70 and Ku70 Rap1 knockout mice. (A) The testes of Ku70−/− and Ku70−/−Rap1Δ/Δ mice are of reduced size, as shown for wild-type (left) and Ku70−/− (right) testes. Scale bar: 5 mm. (B) Ku70−/− and Ku70−/−Rap1Δ/Δ (dko) mice display a significant (P<0.001) reduction in testis size compared with wild type (WT) and heterozygotes (het). Whiskers represent s.d. (≥3 mice). (C–F) Spermatogenesis appears grossly normal in tubule cross-sections of (C) wild type and (D) a Ku70+/−Rap1+/Δ heterozygote. Sperm production and release is reduced in (E) Ku70−/− and (F) Ku70−/−Rap1Δ/Δ stage VI–VII tubules (asterisks). DAPI staining is shown in inverted gray scale. Scale bar: in F, 50 µm for C–F. (G–I) Epididymides of (G) wild type, (H) Ku70−/− and (I) Ku70−/−Rap1Δ/Δ mice (gray scale images of H&E staining). Ku70 knockout mice produce less sperm, as indicated by reduced diameter of epididymal tubules and their emptiness. Scale bar: in I, 100 µm for G–I.
Ku70 localization in testis cells
Next, we investigated the expression and localization of the Ku protein in testis sections of wild-type and Ku70-deficient mice by immunofluorescence (IF), which revealed that Ku70 is abundant in the nucleus of Sertoli cells, type A spermatogonia, post mid-pachytene (late) spermatocytes I, secondary spermatocytes and round spermatids as well as in the cytoplasm of metaphases I and II cells (Fig. 2A; supplementary material Fig. S1). Wild-type and Rap1-deficient mouse testes showed the same Ku70 localization pattern (not shown), whereas Ku70−/− testes lacked any detectable protein (Fig. 2B).
Fig. 2.
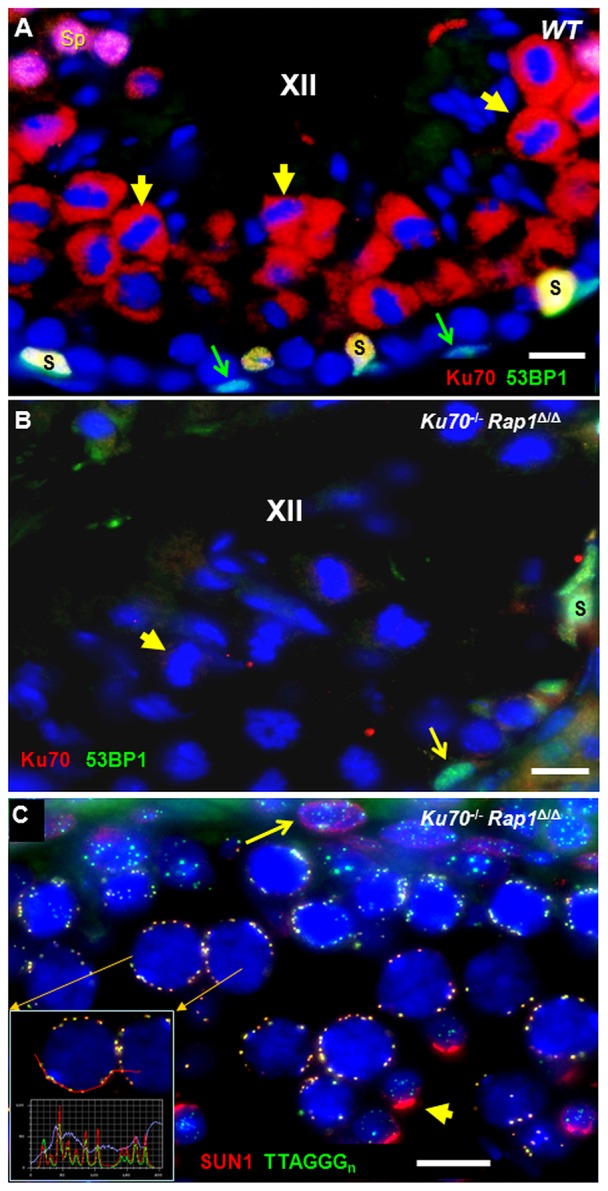
Ku70 (red) and 53BP1 (green) immunofluorescence (IF) staining pattern of wild-type (WT) and Rap1Δ/ΔKu70−/− testes tubules. (A) WT stage XII tubule showing Ku70 protein IF (red) in the cytoplasm of metaphase I and II cells (examples highlighted by short yellow arrows) near the tubule center. Sertoli cells (S) at the tubule periphery appear yellow owing to colocalization of red Ku70 and green 53BP1 fluorescence. A-type spermatogonia show strong greenish 53BP1 expression (green arrows). Spermatids (Sp) appear pink owing to strong fluorescence for Ku70 and 53BP1 at the upper left corner of the image. (B) Ku70−/− stage XII tubule showing absence of red Ku70 protein signals (example indicated by short yellow arrow), whereas an A-type spermatogonium (long arrow) and a Sertoli cell (S) show green 53BP1 fluorescence. (C) IF of SUN1 (red) and (TTAGGG)n telomere FISH (green) revealing the colocalization of SUN1 with meiotic telomeres at the nuclear periphery of pachytene spermatocytes in Rap1Δ/Δ Ku70−/− testis tissue section. The inset shows fluorescence profiles across several telomeres that highlight colocalizing SUN1 (red line) and telomere (green line) fluorescence peaks. This staining is identical to that in wild type (not shown) (cf. Scherthan et al., 2011). A punctate NUP-like somatic distribution pattern of SUN1 (red) is seen at the meiotic NE of a spermatogonium at the tubule periphery (arrow), whereas haploid spermatids (short yellow arrow) display a strong red acrosomal signal. Scale bars: 10 µm.
Rap1 knockout mice, by contrast, lacked the Rap1 telomere protein in testis cells, but spermatogenesis was indistinguishable from that observed in wild type (Scherthan et al., 2011; Sfeir et al., 2010).
Ku70 and Rap1 deficiency leaves meiotic telomere dynamics unchanged
Because Rap1 and Ku70 inhibit homology-directed DNA repair at somatic telomeres (Celli et al., 2006; Sfeir et al., 2010) and are required for normal meiotic telomere behavior in yeasts (Chikashige and Hiraoka, 2001; Scherthan and Trelles-Sticken, 2008), we investigated whether the simultaneous absence of Ku and Rap1 affects meiotic telomere behavior and attachment to the meiotic nuclear envelope (NE). Investigation of (TTAGGG)n fluorescence in situ hybridization (FISH)-tagged telomeres (n>1000) in pachytene nuclei of testis tissue sections revealed normal telomere localization to the nuclear periphery (Fig. 2C). IF for SUN1 (Schmitt et al., 2007) together with telomere repeat FISH on Ku70−/− and Ku70−/−Rap1Δ/Δ spermatocytes showed a wild-type SUN1 distribution at all telomeres in 3D-preserved pachytene nuclei from tissue sections (Fig. 2C). Collectively, these data suggest that meiotic telomeres do not require Rap1 and/or Ku70 for their interaction with SUN1 and attachment to the meiotic nuclear envelope.
At the onset of prophase I, mouse telomeres undergo a dynamic redistribution that involves brief clustering during early zygotene stage (bouquet topology) (Scherthan, 2001). Bouquet spermatocytes, which can be assayed by centromere/telomere FISH (Tel/Cen FISH) in testicular suspensions, are rarely (∼0.3% of spermatocytes) encountered in wild-type mouse meiosis (Liebe et al., 2006), but their significance is underpinned by their enrichment in ATM-deficient meiosis (Pandita et al., 1999). When we determined the frequency of bouquet spermatocytes by Tel/Cen FISH in testicular suspensions of control mice, we noted similar frequencies in Ku70-deficient spermatocytes: Ku70−/− (0.38%, n = 1055), Rap1Δ/+Ku70−/− (0.45%, n = 1103) and Rap1Δ/ΔKu70−/− (0.4%, n = 1004). The difference between the various genetic backgrounds tested was not statistically significant (P>0.05; Fisher's test). Furthermore, bouquet frequencies were similar to those observed in the single Rap1 knockout and in wild type (Liebe et al., 2006; Scherthan et al., 2011).
In contrast, we noted a significant increase (P<0.01, Fisher's test) in the so-called mid-preleptotene spermatocytes in Ku-deficient genotypes, i.e. in Ku70−/− (2.3%, n = 1055), in Ku70−/−Rap1Δ/+ (2.6%, n = 1103) and in Ku70−/−Rap1Δ/Δ (2.6%, n = 1004) spermatocytes, relative to the Rap1 knockout and the wild type, which displayed 0.8% mid-preleptotene spermatocytes (n = 1164). These data corroborate earlier studies that observed a significant increase of mid-preleptotene but not bouquet spermatocytes in the Ku80 knockout mouse (Liebe et al., 2006).
Our data establish that Rap1 and Ku70 are both dispensable for meiotic telomere attachment and clustering in mouse meiosis, whereas passage through the so-called mid-preleptotene stage appears to be prolonged in the absence of Ku70 and NHEJ.
Increased DNA damage in B spermatogonia of the NHEJ-deficient testis
To investigate whether the increased mid-preleptotene levels in Ku70−/− testes indicate elevated dsDNA damage in pre-meiotic cells, we performed immunostaining for the DSB repair markers γH2AX (Rogakou et al., 1998) and 53BP1 (Huyen et al., 2004) in paraffin-embedded tissue sections of Ku70 single and double knockout testes. Surprisingly, we noted a significant elevation of the numbers of DSB-associated foci in B spermatogonia of Ku70−/− and Ku70−/−Rap1Δ/Δ mice (Fig. 3A,B) relative to the frequency in B spermatogonia of Rap1Δ/Δ, heterozygous and control testes (Fig. 3C). These findings indicate that rapidly dividing Ku-deficient B spermatogonia with compromised NHEJ are accumulating replication-dependent DSBs prior to their entry into prophase I.
Fig. 3.

DNA damage in B spermatogonia of Ku70-deficient testes revealed by 53BP1 (red) and γH2AX staining (green). (A) Testicular section of a wild-type mouse showing a few 53BP1 foci (red) in B spermatogonia (indicated by letter B). Wild-type Sertoli cells (S) show no 53BP1 foci. XY denotes the 53BP1- and γH2AX-positive sex body (yellow) of pachytene spermatocytes. (B) Numerous 53BP1 DNA damage foci in B spermatogonia of a Ku70−/−Rap1Δ/Δ testis section. These colocalize with γH2AX foci (green) as shown by the green channel image of one nucleus in the inset. The white arrow denotes a large 53BP1 DNA damage focus in the DAPI-faint chromatin of a double knockout Sertoli cell nucleus. (C) γH2AX and 53BP1 foci numbers are significantly increased (**P<0.001) in Ku70-deficient B spermatogonia of single and double knockout testes relative to the Rap1Δ/Δ, heterozygous and wild-type testes. Error bars indicate s.d.
HR progression in Ku70−/− and Rap1Δ/Δ Ku70−/− spermatocytes
We next investigated the progress of DSB occurrence and DNA repair in Ku- and Rap1-deficient spermatocytes. HR is the dominant repair pathway during much of prophase I and repairs endogenous DSBs that are formed by Spo11 in leptotene chromatin, resulting in crossovers between homologous chromosomes. Leptotene spermatocytes show extensive histone H2AX phosphorylation in their nuclei and H2AX phosphorylation regresses with the progress of HR repair to the sex body of pachytene spermatocytes (Barchi et al., 2005; Mahadevaiah et al., 2001). Delayed repair progression at some DSB sites can lead to a few large, synaptonemal complex-associated γ-H2AX foci during the late pachytene stage of prophase I (Ahmed et al., 2010; Chicheportiche et al., 2007). These foci possibly relate to delayed repair progression as shown by RPA and γH2AX colocalization in mouse and human late pachytene meiocytes (Ahmed et al., 2010; de Vries et al., 2013; Roig et al., 2004). To determine whether carryover of DNA damage from pre-meiotic S phase of B spermatogonia leads to altered DNA repair progression in late Ku70−/− spermatocytes, we examined surface-spread and SYCP3-stained Ku70- and Ku70 Rap1 doubly deficient late pachytene and diplotene spermatocytes for the presence of large γH2AX foci (Fig. 4). It was found that γ-H2AX signals appeared normally during leptotene stage and regressed during zygotene and early pachytene stages to the XY body in all genotypes (data not shown). In late pachytene and diplotene spermatocytes, in which NHEJ repair is reactivated (Ahmed et al., 2010; Goedecke et al., 1999), we noted a moderate but significantly elevated number of spermatocytes displaying on average two γ-H2AX foci/late Ku70−/−Rap1Δ/Δ spermatocyte nucleus (n = 55) (Fig. 4A,B) and in Ku70-deficient diplotene spermatocytes (n = 43) (Fig. 4C,D), as opposed to the one large focus/spermatocyte seen on average in wild-type (n = 49) and Rap1Δ/Δ (n = 66) late pachytene and diplotene spermatocytes.
Fig. 4.
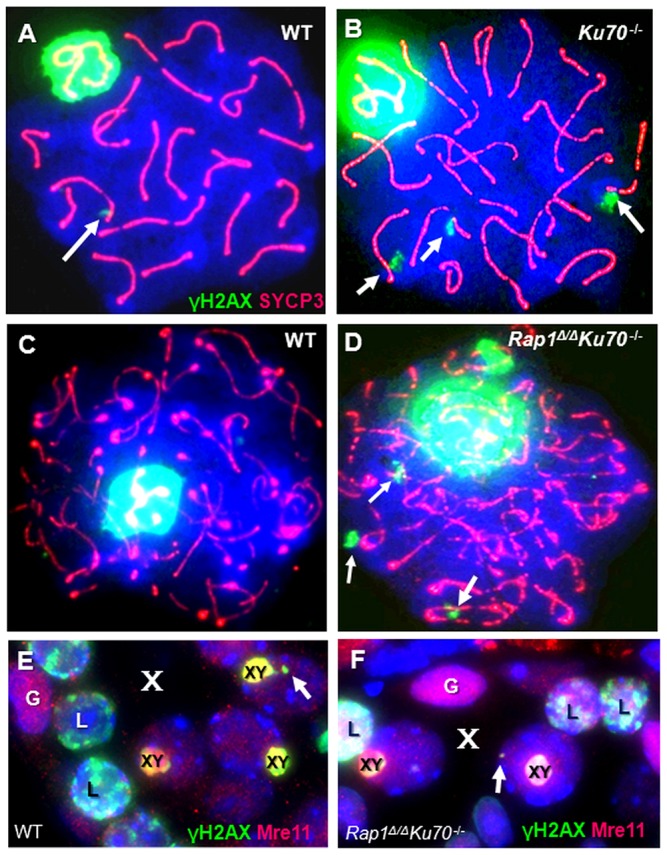
Immunofluorescence of γ-H2AX (green) and SYCP3 (red) in surface-spread late spermatocytes of control, single and double knockout mice reveal normal synapsis. (A,B) Wild-type (A) and Ku70−/− (B) late pachytene spermatocytes. Arrows indicate a few large γH2AX foci. (C,D) Wild-type (C) and Ku70−/−Rap1Δ/Δ (D) diplotene spermatocytes showing a few large γH2AX foci (arrows) on SYCP3 axes (red threads). (E,F) Mre11 (red) and γH2AX (green) staining in details of control (E) and Ku70−/−Rap1Δ/Δ (F) stage IX–X tubules. Large γH2AX foci are seen as dots (arrows) in the chromatin of diplotene spermatocyte nuclei (marked by their XY body; XY) in stage X stage tubules (X). The patchy green-labeled nuclei represent leptotene spermatocytes (L). Spermatogonia (G) display strong Mre11 expression.
To investigate this matter further, we co-stained Mre11 and γH2AX in testis tissue sections of all genotypes and determined the number of γH2AX foci in 3D image projections of late pachytene and diplotene spermatocytes of stage X tubuli (Fig. 4E,F). Comparison of the γH2AX foci frequency per diplotene spermatocyte from five Ku70−/− mice (n = 869 cells) revealed 0.082 (±0.06, s.d.) γH2AX foci/nucleus, whereas there were only 0.061 (±0.03) foci/nucleus in control diplotene spermatocytes (n = 896; four mice), representing a highly significant difference (t-test; P<0.001). These data agree with our γ-H2AX focus analysis in surface-spread spermatocytes and suggest that some DSBs in prophase I might go unrepaired in absence of Ku70/NHEJ. The role of Ku70 in repairing Spo11-dependent DSBs will require further analysis of a Ku70 Spo11 double knockout mouse.
Ku deficiency does not alter DNA repair at meiotic telomeres
To address specifically DSB repair events at meiotic telomeres in the mutants, we also determined the frequency of occurrence of large (L) telomeric γ-H2AX foci at synaptonemal complex (SC) ends in ≥25 surface-spread late spermatocytes (>920 telomeres per genotype tested). The average colocalization between telomeres and L γ-H2AX foci at SC ends per wild-type late pachytene cell was 0.19 (±0.4 s.d.) and 0.15 (±0.36) in Rap1Δ/Δ; 0.33 (±0.48) in Ku70−/− and 0.20 (±0.41) in Ku70−/−Rap1Δ/Δ spermatocytes, indicating insignificant differences (t-test) between the genotypes studied. These results suggest that the single or combined deletion of Ku70 and Rap1 has no effect on HR at meiotic telomeres and agrees with the observations made in the Rap1Δ/Δ mouse (Scherthan et al., 2011).
Homolog pairing is unaffected by absence of NHEJ and Rap1
To test further whether Ku70 deficiency has an impact on chromosome pairing and segregation at meiotic metaphases, we investigated SC formation and X and Y chromosome pairing in pachytene and metaphase I cells (Fig. 5). SC staining with SCP3 antibodies revealed no overt differences in SC formation among the genotypes studied (Fig. 4A–D; data not shown), suggesting normal homolog pairing. X and Y chromosomes share only a small site of homology, the pseudoautosomal region (PAR) (reviewed by Otto et al., 2011), which undergoes late pairing and forms an obligate crossover in nearly all meioses (Kauppi et al., 2011). If there were problems in pairing or recombination we would expect defects in XY segregation. Studying XY distribution in pachytene and MI cells by two color XY chromosome painting in tissue sections revealed a normal association between the X and Y chromosomes in late pachytene and diplotene spermatocytes (Fig. 5A): paired (touching) X and Y chromosome paint signals were observed in 94.1% of wild type (n = 85), in 94.2% of Ku70−/− (n = 154) and in 94.7% of Ku70−/−Rap1Δ/Δ (n = 95) late spermatocytes, the differences being statistically insignificant (P = 1; Fisher's test).
Fig. 5.

XY chromosome pairing as studied by X (green) and Y (red) chromosome painting in tissue sections. (A) Stage VI tubule of a Ku70−/−Rap1Δ/Δ testis showing pairing (touching) of XY signals in pachytene spermatocytes (arrow). (B) Stage XII tubule of a double knockout testis showing XY bivalents with close signal apposition in metaphase I plates. Blue, DNA stain DAPI. Signal distribution in wild type and heterozygotes was essentially the same (not shown). Scale bars: 10 µm.
At metaphase I of meiosis, X and Y chromosome-specific signals were touching each other in 95.6% of wild-type MI cells (n = 45), in 92.7% of Ku70−/− MI cells (n = 41) and in 91.9% of Ku70−/−Rap1Δ/Δ MI cells (n = 49) (Fig. 5B), with the difference between wild type and all Ku70−/− genotypes being insignificant (P = 0.67; Fisher's test). Nonetheless, the slightly more XY univalents in the Ku70−/− condition align with our data showing increased frequency of DSBs in late prophase (diplotene) spermatocytes.
Ku70−/− spermatocytes display elevated metaphase apoptosis
To investigate checkpoint activation in mutant testes, we stained tissue sections of Ku70−/− and Ku70−/−Rap1Δ/Δ testes for apoptosis using the TUNEL assay (Fig. 6A–D). In the testes of Ku70−/− and Ku70−/−Rap1Δ/Δ mice, the frequency of apoptotic cells in stage IV-VI tubules, which are subject to HR checkpoint function (Barchi et al., 2005; de Rooij and de Boer, 2003), was similar for each genotype (Fig. 6A,B,E). This suggests that ectopic DSBs carried over from pre-meiotic cell cycles are processed by HR repair in a manner that allows for stage IV checkpoint bypass.
Fig. 6.

Apoptosis in wild-type and Ku70−/−Rap1Δ/Δ testes sections. (A) Wild-type stage IV tubule showing two TUNEL-positive apoptotic spermatocytes (green). (B) Ku70−/−Rap1Δ/Δ stage IV tubule with three brightly labeled apoptotic cells. (C) Wild-type stage XII tubule showing two apoptotic meiosis I cells (green) and well developed metaphase I plates (blue; arrows). The red fluorescence stems from patchy γH2AX distribution in zygotene spermatocytes. Red arrows indicate examples of metaphase plates. (D) Double knockout stage XII tubule showing high number of TUNEL-positive apoptotic meiosis I cells. The red arrows denote apoptotic metaphase cells without chromosome congression. The long yellow arrow denotes an apoptotic metaphase with two non-aligned chromosomes. (E,F) Quantification of TUNEL-positive apoptotic cells in stages XII-I and stages IV-VI in control, single and double knockout mice. Although apoptosis was similar in stage IV-VI tubules (E), there was a significant increase of metaphase apoptosis in stage XII tubules (F) of Ku70-deficient testes relative to wild type and single Rap1 knockout. Scale bars: 10 µm. Error bars indicate s.d. **P<0.01.
However, apoptosis in stage XII-I tubules that contain meiotic metaphases (Fig. 6C,D) was significantly increased (Fig. 6F), indicating an increased demise of Ku-deficient meiotic metaphase cells. Investigation of 3D image projections of metaphase I cells in stage XII tubules for the presence of γH2AX foci (supplementary material Fig. S2) revealed an average of 0.06 γH2AX foci per Ku70−/− meiotic metaphase (n = 100) and 0.036 foci per meiotic metaphase in the wild type (n = 137). Again, there is a slight (1.4-fold) but insignificant (P = 0.54; Fisher's test) increase of meiotic metaphases with γH2AX foci in the Ku70−/− condition.
To investigate further the cause of an activated spindle assembly checkpoint (SAC) in the Ku70−/− condition, we examined the morphology of apoptotic MI cells for the presence of univalent chromosomes and for signs of congression failure that might activate the SAC, as noted on mouse oocytes (Zhu et al., 2010). To this end, we obtained maximum projection images from three-dimensional image stacks of TUNEL-positive apoptotic MI cells and investigated them for their chromosome distribution by DAPI staining where possible. We observed univalents off the metaphase plate (Fig. 6D, yellow arrow) in 18.8% of apoptotic MI cells in the Ku70−/− background and 16.3% of wild-type and Rap1-deficient meiotic metaphase cells, a non-significant difference (P = 0.86, Fisher's test). However, the Ku70−/− apoptotic MI cells displayed 35.6% of cells (n = 76) with numerous non-aligned chromosomes (Fig. 6D, red arrows), whereas in the controls congression failure was seen in 22.1% of apoptotic MI cells (n = 153), a significant difference (P = 0.005). This suggests that the absence of Ku70 leads to the activation of the SAC in meiotic metaphase cells when chromosome congression is still in process. Ku deficiency might cause metaphase cell death as a result of unrepaired DSBs carried over from late prophase I and/or a defective metaphase structure (Higashiura et al., 1992). Further analysis will be required to understand this in detail.
Sertoli cells of Ku70-deficient testes display DNA damage
Spermatogenesis is under the control of Sertoli cells, which direct meiotic differentiation and are non-proliferative in the adult testes (see Petersen and Soder, 2006). Because of their non-proliferative nature, they depend on NHEJ to repair DSBs. Our analysis of DNA damage surprisingly revealed large 53BP1-positive DNA damage foci in the conspicuous nucleus of Ku70−/− and Rap1Δ/Δ Ku70−/− Sertoli cells (Fig. 7), with >14.7% of Ku−/− Sertoli cells expressing on average 1.22 DNA damage foci (range 1–6) per 53BP1 foci-positive cell (Fig. 7; Table 1). Co-staining of 53BP1 with either γ-H2AX (Fig. 7C,D) or activated (phospho-Ser-1981) ATM revealed a colocalization of ATMp and its substrate γ-H2AX at 53BP1 foci in the Sertoli cell nucleus (Fig. 7E,F), demonstrating an ongoing DNA damage response (DDR). Because dsDNA damage foci were only occasionally observed in Sertoli cells of all other genotypes investigated (Fig. 7A; Table 1), we conclude that the absence of NHEJ leads to the presence of persistent DNA damage also in differentiated Sertoli cells.
Fig. 7.

Persistent DNA damage and active DDR in Ku70-deficient Sertoli cells of testes sections. (A) Wild-type Sertoli cells (arrows) show strong diffuse 53BP1 fluorescence (red) throughout the nucleus except for the nucleolus (dark dot) at its center. Bright red dots in spermatocyte nuclei represent the XY body. (B) Ku70−/−Rap1Δ/Δ Sertoli cells displaying large red 53BP1 DNA damage foci (arrows) in their nuclear chromatin. The two blue dots next to the dark spot (the nucleolus) represent the chromocenters that are specific for this cell type. (C-D′) Ku70-deficient Sertoli cells showing colocalization of strong 53BP1 (red) with weaker γH2AX signals (green) at large DNA damage foci (arrows). (D) Sertoli cell with six 53BP1 foci, four of which contain γ-H2AX fluorescence (green) as indicated by their yellowish color. (D′) Same cell as in D with the blue color of the DAPI stain converted to gray for better display. (E,E′) Colocalization of 53BP1 (red) with activated ATM (ATMp, green) at two large DNA damage foci (arrows) of a Ku70−/− Sertoli cell. One focus (lower arrow) contains a large amount of ATMp. (F) Sertoli cell with three damage foci, two of which contain ATMp (arrows). (G,H) TTAGGGn telomere FISH signals (red) only rarely colocalize with 53BP1 damage foci (green; arrows) in Ku70−/− Sertoli cell nuclei. In such cases, the small telomere signals lie at the border of the large 53BP1 foci (short arrows).
Table 1. Increased DNA damage in Ku70-deficient Sertoli cells.

DNA damage foci in Sertoli cell nuclei do not localize with the telomeres
Sertoli cell nuclei display clustering of proximal telomeres near conspicuous chromocenters of pericentric heterochromatin (Scherthan et al., 2000). Because Rap1 and Ku70 protect telomeres from unwanted HR (Sfeir et al., 2010), deprotection of telomeres can lead to DDR activation and 53BP1 accumulation at dysfunctional telomeres (Takai et al., 2003). Thus, we investigated whether the large DNA damage foci in Rap1Δ/Δ Ku70−/− and Ku70−/− Sertoli cells represent deprotected telomeres. Co-staining for 53BP1 by IF and telomeres by FISH revealed that the large DNA damage foci do not generally colocalize with the smaller telomere signals in Sertoli cell nuclei (Fig. 7G,H), ruling out the possibility that DNA damage foci in NHEJ-deficient Sertoli cell nuclei represent deprotected telomeres.
Discussion
Here we observed that Ku70 disruption leads to reduced testis size, compromised fidelity of spermatogenesis and DNA repair, compromised sperm production and elevated metaphase I apoptosis. In contrast, combined or single deficiency of Ku70 and RAP1 did not affect meiotic telomere attachment and dynamics leading to normal telomere clustering (bouquet formation) in any of the mutants analyzed. These observations match with normal telomere dynamics in Rap1Δ/Δ spermatogenesis (Scherthan et al., 2011) and normal telomere clustering in the Ku80 knockout mouse (Liebe et al., 2006). The observed normal meiotic telomere dynamics in mice contrasts with the situation in single-celled eukaryotes, in which yRap1 and yKu70 deficiency deteriorates meiotic telomere behavior (Chikashige and Hiraoka, 2001; Scherthan and Trelles-Sticken, 2008). Hence, our observations in the mouse establish that the Ku protein complex and Rap1 are dispensable for meiotic telomere dynamics in mammalian meiosis.
As in the Ku80 knockout mouse (Liebe et al., 2006), Ku70−/− spermatogenesis was found to exhibit a significantly elevated level of so-called mid-preleptotene spermatocytes relative to wild-type and Rap1-deficient meiosis. It had been reasoned that the increase in mid-preleptotene spermatocytes is related to carry over of replication-dependent DNA damage into prophase I (Liebe et al., 2006). Along this line, it has been shown that yKu is required for recovery from replication stress (Miyoshi et al., 2009). Here, we observed that Ku70-deficient mice exhibit increased levels of dsDNA damage in somatic testis cells, as indicated by 53BP1, γ-H2AX and pATM foci in B spermatogonia and Sertoli cells. B spermatogonia proliferate and enter first meiotic prophase after a lengthy S phase. Our findings thus highlight the importance of Ku/NHEJ for repairing replication-dependent DNA damage prior to the entry into prophase I.
Sertoli cells also displayed significantly increased DNA damage. They nurse and direct spermatogenic cells (see Petersen and Soder, 2006) and proliferate before puberty after which they arrest (Hayrabedyan et al., 2012). Because they are in cell cycle stage G0/1, Sertoli cells rely on NHEJ for DNA repair. The presence of DNA damage-responsive signaling molecules, such as ATMp and 53BP1, at the large nuclear DNA damage foci of Sertoli nuclei indicates an ongoing DNA damage response. Deprotection of telomeres leads to a DDR involving ATM activation and 53BP1 accumulation at telomeres (Takai et al., 2003). However, Ku70−/− Sertoli cell telomeres did not colocalize with the DNA damage foci, suggesting that the absence of Ku70 or Rap1 does not lead to deprotected telomeres in this cell type. Thus, the formation of large persistent dsDNA damage foci in the absence of NHEJ is likely to relate to the clustering of replication-dependent DNA damage after prepuberal Sertoli cell proliferation. Clustering of DSB-harboring chromatin domains has, for instance, been noted in α-irradiated somatic cells (Aten et al., 2004). Furthermore, it is possible that the action of reactive oxygen species might have induced accumulation of DNA damage in this cell type. However, because other NHEJ-deficient testicular cells did not display such large DNA damage foci, we consider it likely that these damage foci result from replication stress during the prepuberal Sertoli proliferation that remains irreparable in absence of NHEJ, the major repair pathway of cell cycle-arrested Sertoli cells (Ahmed et al., 2009; Ahmed et al., 2007).
In all, it appears that NHEJ is required for DNA repair in somatic cells of the testis, where Ku70 colocalizes with the Mre11 protein and acts as a switch between NHEJ and HR repair pathways. In spermatocytes, Ku is downregulated during the early stages of meiotic prophase, i.e. in leptotene to mid-pachytene spermatocytes, a time frame during which meiotic DSBs are repaired by HR (Goedecke et al., 1999). In Ku70−/− mice, we noted the absence of an activated meiotic (recombination) checkpoint in stage IV tubules, normal synapsis and a regression of the Spo11- and DSB-dependent γH2AX signal to the XY body of pachytene and diplotene spermatocytes. In contrast to the Ku70−/− testis, SCID mice, which carry an inactivating mutation in the DNA-PK catalytic subunit and are compromised in NHEJ, display numerous apoptotic spermatocytes at tubule stage IV, indicating activation of the recombination-dependent meiotic checkpoint (de Rooij and de Boer, 2003; Hamer et al., 2003). The absence of elevated stage IV pachytene apoptosis in our Ku70−/− mice indicates that HR processes the DSBs carried over from pre-meiotic cell cycles to a state that allows stage IV checkpoint bypass. This distinguishes them from SCID mice, which display reduced DNAPKcs activity and elevated stage IV apoptosis (Hamer et al., 2003).
However, there was a significant elevation of large γH2AX foci in diplotene Ku−/− spermatocytes, indicating some unrepaired DSBs in these late prophase cells. The presence of γH2AX foci in low frequency meiotic metaphase cells indicates that HR repairs most, but not all, of the DSBs that are present during prophase I. The observed 1.4-fold increase of meiotic metaphases with γH2AX focus signals (possibly marking unrepaired or not completely repaired DSBs) might explain the ∼1.6-fold increase in meiotic metaphase apoptosis in Ku70−/− stage XII tubules noted by TUNEL staining. A cross talk between the DNA damage response and the spindle assembly checkpoint has been noted previously (Choi and Lee, 2008; Maringele and Lydall, 2002; Yang et al., 2012) and might be instrumental for this situation.
It is noteworthy that Ku70 is highly expressed in the cytoplasm of meiotic metaphase I and II cells and in mitotic metaphase cells where it also localizes to the surface of condensed metaphase chromosomes (Higashiura et al., 1992; Koike et al., 1999). Furthermore, Ku70 seems to play a role in apoptosis regulation, possibly through its interaction with BAX (Amsel et al., 2008; Harrington et al., 1992) and by influencing the cell survival transcription network (Fell and Schild-Poulter, 2012). In all, absence of Ku in metaphase cells could lead to apoptosis by pleiotropic effects influencing DNA damage response, SAC pathways and survival networks. How this is regulated requires further investigation.
In summary, our data indicate that Ku/NHEJ contributes to the repair of replication-dependent and exogenous DNA damage in somatic testis cells, and might also be required for the cleanup repair of some persistent Spo11-dependent DSBs in late meiotic prophase cells and for a normal metaphase structure. By these mechanisms, Ku ensures the fidelity of spermatogenesis.
Materials and Methods
Mice
Rap1 knockout mice were produced by a conditional knockout strategy of exon2 (Rap1Δex2/Δex2) (Sfeir et al., 2010) and denoted Rap1Δ/Δ (Scherthan et al., 2011; Sfeir et al., 2010). Rap1Δex2/Δex2 targeted mice were maintained in a C57BL/6J background. Rap1Δ/Δ and Ku70 knockout mice (Gu et al., 1997) were bred and crossed to give double knockout mice as described elsewhere (Sfeir et al., 2010).
Testicular preparations, detergent spreading and FISH
Testes were recovered from mice immediately after sacrifice by cervical dislocation. Usually one testis was paraffin embedded according to standard procedures and the other was frozen in isopentane (Sigma) until spreading or suspension preparation. Testicular preparations, surface spreading, immunostaining as well as telomere and centromere fluorescence in situ hybridization (FISH) for bouquet stage investigation and chromosome painting were carried out as described in detail elsewhere (Liebe et al., 2006; Scherthan, 2009). Differentially labeled X and Y mouse chromosome paint probes were obtained from MetaSystems (Altlussheim, Germany) and hybridized to tissue sections as described (Scherthan et al., 1996).
TUNEL assay
TUNEL analysis was performed to detect apoptotic nuclei in 5-µm paraffin-embedded sections (Ahmed et al., 2007) according to the manufacturer's protocol (In Situ Cell Death Detection Kit, POD; Roche Diagnostics GmbH, Mannheim, Germany).
Antisera and immunofluorescence
The following affinity-purified antibodies were used in the immunostaining experiments: rabbit anti-SYCP3 (1∶250, Abcam, UK), rabbit anti-γ-H2AX (1∶500, Millipore, Schwalbach), goat anti-Ku70 (sc-1487, 1∶10, Santa Cruz, Heidelberg). Guinea pig anti-SUN1 was a kind gift of M. Alsheimer, University of Würzburg, Germany. Antisera were diluted in PBS, 0.1% Tween 20, 0.2% BSA, 0.1% gelatin (PBTG). All antibodies were tested in individual staining reactions for their specificity and performance. Controls without primary antibodies were all negative (not shown).
Immunostaining was performed as described earlier (Scherthan et al., 2011). Preparations were mounted in antifade solution (Vectashield, Vector Labs) containing 0.5 µg/ml DAPI (Sigma) to reveal nuclear DNA. In mouse testicular suspension preparations, the meiotic prophase stage-specific distribution of SYCP3 proteins and/or DAPI-bright heterochromatin clusters (Liebe et al., 2006; Scherthan et al., 1996) was utilized to identify spermatocytes at various stages of prophase I.
Microscopic evaluation
Microscopy was carried out using a Zeiss Axioplan 2 epifluorescence microscope (Carl Zeiss, Oberkochen) equipped with single band pass filters for excitation of green, red and blue fluorescence (Chroma Technologies, Bellows Falls) and 10×, 40×, 63× and 100× plan-Neofluoar lenses. Three-dimensional evaluation of immunostained nuclei was performed in some experiments by carefully focusing through the nuclei using a 63× lens. Digital black and white images were recorded with a cooled CCD camera and merged with RGB images using the ISIS fluorescence image analysis system (MetaSystems). In some cases, a MetaFer image analysis system was used to automatically record image stacks and to obtain maximum projection images from tissue sections. In this case, image collections were then scanned interactively for scoring.
Supplementary Material
Acknowledgments
We thank Titia de Lange, Rockefeller University, and H.H. Ropers MPI-MG, Berlin, for continuous support; and M. Alsheimer, University of Würzburg, Germany, for providing SUN1 antiserum. H.S. acknowledges technical support from G. Schrock and C. Nowak in the late phase of the project.
Footnotes
Author contributions
H.S. and A.S. designed experiments. E.A.A., H.S., H.T. and A.S. performed experiments and interpreted the data. H.S. wrote the manuscript.
Funding
E.A.A. was supported by a postdoctoral fellowship from the Alexander von Humboldt foundation. A.S. was supported by a postdoctoral fellowship from Susan G. Komen for the Cure. These studies were supported by grants from the National Institutes of Health (NIH) [grant numbers AG016642 and GM049046 to Titia de Lange]; and the Deutsche Forschungsgemeinschaft [grant number SCHE350/10-1, SPP1384 to H.S.]. Deposited in PMC for release after 12 months.
Supplementary material available online at http://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.122788/-/DC1
References
- Ahmed E. A., van der Vaart A., Barten A., Kal H. B., Chen J., Lou Z., Minter-Dykhouse K., Bartkova J., Bartek J., de Boer P. et al. (2007). Differences in DNA double strand breaks repair in male germ cell types: lessons learned from a differential expression of Mdc1 and 53BP1. DNA Repair (Amst.) 6, 1243–1254 10.1016/j.dnarep.2007.02.011 [DOI] [PubMed] [Google Scholar]
- Ahmed E. A., Barten-van Rijbroek A. D., Kal H. B., Sadri-Ardekani H., Mizrak S. C., van Pelt A. M., de Rooij D. G. (2009). Proliferative activity in vitro and DNA repair indicate that adult mouse and human Sertoli cells are not terminally differentiated, quiescent cells. Biol. Reprod. 80, 1084–1091 10.1095/biolreprod.108.071662 [DOI] [PubMed] [Google Scholar]
- Ahmed E. A., Philippens M. E., Kal H. B., de Rooij D. G., de Boer P. (2010). Genetic probing of homologous recombination and non-homologous end joining during meiotic prophase in irradiated mouse spermatocytes. Mutat. Res. 688, 12–18 10.1016/j.mrfmmm.2010.02.004 [DOI] [PubMed] [Google Scholar]
- Amsel A. D., Rathaus M., Kronman N., Cohen H. Y. (2008). Regulation of the proapoptotic factor Bax by Ku70-dependent deubiquitylation. Proc. Natl. Acad. Sci. USA 105, 5117–5122 10.1073/pnas.0706700105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aten J. A., Stap J., Krawczyk P. M., van Oven C. H., Hoebe R. A., Essers J., Kanaar R. (2004). Dynamics of DNA double-strand breaks revealed by clustering of damaged chromosome domains. Science 303, 92–95 10.1126/science.1088845 [DOI] [PubMed] [Google Scholar]
- Bailey S. M., Meyne J., Chen D. J., Kurimasa A., Li G. C., Lehnert B. E., Goodwin E. H. (1999). DNA double-strand break repair proteins are required to cap the ends of mammalian chromosomes. Proc. Natl. Acad. Sci. USA 96, 14899–14904 10.1073/pnas.96.26.14899 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bannister L. A., Schimenti J. C. (2004). Homologous recombinational repair proteins in mouse meiosis. Cytogenet. Genome Res. 107, 191–200 10.1159/000080597 [DOI] [PubMed] [Google Scholar]
- Barchi M., Mahadevaiah S., Di Giacomo M., Baudat F., de Rooij D. G., Burgoyne P. S., Jasin M., Keeney S. (2005). Surveillance of different recombination defects in mouse spermatocytes yields distinct responses despite elimination at an identical developmental stage. Mol. Cell. Biol. 25, 7203–7215 10.1128/MCB.25.16.7203-7215.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertuch A. A., Lundblad V. (2006). The maintenance and masking of chromosome termini. Curr. Opin. Cell Biol. 18, 247–253 10.1016/j.ceb.2006.04.005 [DOI] [PubMed] [Google Scholar]
- Celli G. B., Denchi E. L., de Lange T. (2006). Ku70 stimulates fusion of dysfunctional telomeres yet protects chromosome ends from homologous recombination. Nat. Cell Biol. 8, 885–890 10.1038/ncb1444 [DOI] [PubMed] [Google Scholar]
- Chicheportiche A., Bernardino-Sgherri J., de Massy B., Dutrillaux B. (2007). Characterization of Spo11-dependent and independent phospho-H2AX foci during meiotic prophase I in the male mouse. J. Cell Sci. 120, 1733–1742 10.1242/jcs.004945 [DOI] [PubMed] [Google Scholar]
- Chikashige Y., Hiraoka Y. (2001). Telomere binding of the Rap1 protein is required for meiosis in fission yeast. Curr. Biol. 11, 1618–1623 10.1016/S0960-9822(01)00457-2 [DOI] [PubMed] [Google Scholar]
- Choi E., Lee H. (2008). Chromosome damage in mitosis induces BubR1 activation and prometaphase arrest. FEBS Lett. 582, 1700–1706 10.1016/j.febslet.2008.04.028 [DOI] [PubMed] [Google Scholar]
- de Lange T. (2005). Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev. 19, 2100–2110 10.1101/gad.1346005 [DOI] [PubMed] [Google Scholar]
- de Rooij D. G., de Boer P. (2003). Specific arrests of spermatogenesis in genetically modified and mutant mice. Cytogenet. Genome Res. 103, 267–276 10.1159/000076812 [DOI] [PubMed] [Google Scholar]
- de Vries M., Ramos L., de Boer P. (2013). Immunofluorescent characterization of meiotic recombination in human males with variable spermatogenesis. Andrology 1, 262–273 10.1111/j.2047-2927.2012.00039.x [DOI] [PubMed] [Google Scholar]
- Ding X., Xu R., Yu J., Xu T., Zhuang Y., Han M. (2007). SUN1 is required for telomere attachment to nuclear envelope and gametogenesis in mice. Dev. Cell 12, 863–872 10.1016/j.devcel.2007.03.018 [DOI] [PubMed] [Google Scholar]
- Fell V. L., Schild-Poulter C. (2012). Ku regulates signaling to DNA damage response pathways through the Ku70 von Willebrand A domain. Mol. Cell. Biol. 32, 76–87 10.1128/MCB.05661-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gerton J. L., Hawley R. S. (2005). Homologous chromosome interactions in meiosis: diversity amidst conservation. Nat. Rev. Genet. 6, 477–487 10.1038/nrg1614 [DOI] [PubMed] [Google Scholar]
- Goedecke W., Eijpe M., Offenberg H. H., van Aalderen M., Heyting C. (1999). Mre11 and Ku70 interact in somatic cells, but are differentially expressed in early meiosis. Nat. Genet. 23, 194–198 10.1038/13821 [DOI] [PubMed] [Google Scholar]
- Gu Y., Seidl K. J., Rathbun G. A., Zhu C., Manis J. P., van der Stoep N., Davidson L., Cheng H. L., Sekiguchi J. M., Frank K. et al. (1997). Growth retardation and leaky SCID phenotype of Ku70-deficient mice. Immunity 7, 653–665 10.1016/S1074-7613(00)80386-6 [DOI] [PubMed] [Google Scholar]
- Hamer G., Roepers-Gajadien H. L., van Duyn-Goedhart A., Gademan I. S., Kal H. B., van Buul P. P., Ashley T., de Rooij D. G. (2003). Function of DNA-protein kinase catalytic subunit during the early meiotic prophase without Ku70 and Ku86. Biol. Reprod. 68, 717–721 10.1095/biolreprod.102.008920 [DOI] [PubMed] [Google Scholar]
- Harrington J., Hsieh C. L., Gerton J., Bosma G., Lieber M. R. (1992). Analysis of the defect in DNA end joining in the murine scid mutation. Mol. Cell. Biol. 12, 4758–4768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayrabedyan S., Todorova K., Pashova S., Mollova M., Fernández N. (2012). Sertoli cell quiescence - new insights. Am. J. Reprod. Immunol. 68, 451–455 10.1111/j.1600-0897.2012.01137.x [DOI] [PubMed] [Google Scholar]
- Higashiura M., Shimizu Y., Tanimoto M., Morita T., Yagura T. (1992). Immunolocalization of Ku-proteins (p80/p70): localization of p70 to nucleoli and periphery of both interphase nuclei and metaphase chromosomes. Exp. Cell Res. 201, 444–451 10.1016/0014-4827(92)90293-H [DOI] [PubMed] [Google Scholar]
- Hiraoka Y., Dernburg A. F. (2009). The SUN rises on meiotic chromosome dynamics. Dev. Cell 17, 598–605 10.1016/j.devcel.2009.10.014 [DOI] [PubMed] [Google Scholar]
- Holcomb V. B., Vogel H., Hasty P. (2007). Deletion of Ku80 causes early aging independent of chronic inflammation and Rag-1-induced DSBs. Mech. Ageing Dev. 128, 601–608 10.1016/j.mad.2007.08.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hsu H. L., Gilley D., Blackburn E. H., Chen D. J. (1999). Ku is associated with the telomere in mammals. Proc. Natl. Acad. Sci. USA 96, 12454–12458 10.1073/pnas.96.22.12454 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huyen Y., Zgheib O., Ditullio R. A. Jr., Gorgoulis V. G., Zacharatos P., Petty T. J., Sheston E. A., Mellert H. S., Stavridi E. S., Halazonetis T. D. (2004). Methylated lysine 79 of histone H3 targets 53BP1 to DNA double-strand breaks. Nature 432, 406–411 10.1038/nature03114 [DOI] [PubMed] [Google Scholar]
- Kauppi L., Barchi M., Baudat F., Romanienko P. J., Keeney S., Jasin M. (2011). Distinct properties of the XY pseudoautosomal region crucial for male meiosis. Science 331, 916–920 10.1126/science.1195774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keeney S. (2008). Spo11 and the formation of DNA double-strand breaks in meiosis. Genome. Dyn. Stab. 2, 81–123 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koike M., Awaji T., Kataoka M., Tsujimoto G., Kartasova T., Koike A., Shiomi T. (1999). Differential subcellular localization of DNA-dependent protein kinase components Ku and DNA-PKcs during mitosis. J. Cell Sci. 112, 4031–4039 [DOI] [PubMed] [Google Scholar]
- Laroche T., Martin S. G., Gotta M., Gorham H. C., Pryde F. E., Louis E. J., Gasser S. M. (1998). Mutation of yeast Ku genes disrupts the subnuclear organization of telomeres. Curr. Biol. 8, 653–656 10.1016/S0960-9822(98)70252-0 [DOI] [PubMed] [Google Scholar]
- Liebe B., Petukhova G., Barchi M., Bellani M., Braselmann H., Nakano T., Pandita T. K., Jasin M., Fornace A., Meistrich M. L. et al. (2006). Mutations that affect meiosis in male mice influence the dynamics of the mid-preleptotene and bouquet stages. Exp. Cell Res. 312, 3768–3781 10.1016/j.yexcr.2006.07.019 [DOI] [PubMed] [Google Scholar]
- Lieber M. R. (2010). The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. 79, 181–211 10.1146/annurev.biochem.052308.093131 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludérus M. E., van Steensel B., Chong L., Sibon O. C., Cremers F. F., de Lange T. (1996). Structure, subnuclear distribution, and nuclear matrix association of the mammalian telomeric complex. J. Cell Biol. 135, 867–881 10.1083/jcb.135.4.867 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahadevaiah S. K., Turner J. M., Baudat F., Rogakou E. P., de Boer P., Blanco-Rodríguez J., Jasin M., Keeney S., Bonner W. M., Burgoyne P. S. (2001). Recombinational DNA double-strand breaks in mice precede synapsis. Nat. Genet. 27, 271–276 10.1038/85830 [DOI] [PubMed] [Google Scholar]
- Maringele L., Lydall D. (2002). EXO1-dependent single-stranded DNA at telomeres activates subsets of DNA damage and spindle checkpoint pathways in budding yeast yku70Delta mutants. Genes Dev. 16, 1919–1933 10.1101/gad.225102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyoshi T., Kanoh J., Ishikawa F. (2009). Fission yeast Ku protein is required for recovery from DNA replication stress. Genes Cells 14, 1091–1103 10.1111/j.1365-2443.2009.01337.x [DOI] [PubMed] [Google Scholar]
- Morimoto A., Shibuya H., Zhu X., Kim J., Ishiguro K., Han M., Watanabe Y. (2012). A conserved KASH domain protein associates with telomeres, SUN1, and dynactin during mammalian meiosis. J. Cell Biol. 198, 165–172 10.1083/jcb.201204085 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nussenzweig A., Chen C., da Costa Soares V., Sanchez M., Sokol K., Nussenzweig M. C., Li G. C. (1996). Requirement for Ku80 in growth and immunoglobulin V(D)J recombination. Nature 382, 551–555 10.1038/382551a0 [DOI] [PubMed] [Google Scholar]
- Nussenzweig A., Sokol K., Burgman P., Li L., Li G. C. (1997). Hypersensitivity of Ku80-deficient cell lines and mice to DNA damage: the effects of ionizing radiation on growth, survival, and development. Proc. Natl. Acad. Sci. USA 94, 13588–13593 10.1073/pnas.94.25.13588 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otto S. P., Pannell J. R., Peichel C. L., Ashman T. L., Charlesworth D., Chippindale A. K., Delph L. F., Guerrero R. F., Scarpino S. V., McAllister B. F. (2011). About PAR: the distinct evolutionary dynamics of the pseudoautosomal region. Trends Genet. 27, 358–367 10.1016/j.tig.2011.05.001 [DOI] [PubMed] [Google Scholar]
- Ouyang H., Nussenzweig A., Kurimasa A., Soares V. C., Li X., Cordon-Cardo C., Li W., Cheong N., Nussenzweig M., Iliakis G. et al. (1997). Ku70 is required for DNA repair but not for T cell antigen receptor gene recombination In vivo. J. Exp. Med. 186, 921–929 10.1084/jem.186.6.921 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pandita T. K., Westphal C. H., Anger M., Sawant S. G., Geard C. R., Pandita R. K., Scherthan H. (1999). Atm inactivation results in aberrant telomere clustering during meiotic prophase. Mol. Cell. Biol. 19, 5096–5105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petersen C., Soder O. (2006). The sertoli cell – a hormonal target and ‘super’ nurse for germ cells that determines testicular size. Horm. Res. 66, 153–161 10.1159/000094142 [DOI] [PubMed] [Google Scholar]
- Rogakou E. P., Pilch D. R., Orr A. H., Ivanova V. S., Bonner W. M. (1998). DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 273, 5858–5868 10.1074/jbc.273.10.5858 [DOI] [PubMed] [Google Scholar]
- Roig I., Liebe B., Egozcue J., Cabero L., Garcia M., Scherthan H. (2004). Female-specific features of recombinational double-stranded DNA repair in relation to synapsis and telomere dynamics in human oocytes. Chromosoma 113, 22–33 10.1007/s00412-004-0290-8 [DOI] [PubMed] [Google Scholar]
- Scherthan H. (2001). A bouquet makes ends meet. Nat. Rev. Mol. Cell Biol. 2, 621–627 10.1038/35085086 [DOI] [PubMed] [Google Scholar]
- Scherthan H. (2006). Meiotic telomeres. Telomeres 225–259Cold Spring Harbor, NY: Cold Spring Harbor Press [Google Scholar]
- Scherthan H. (2009). Analysis of telomere dynamics in mouse spermatogenesis. Methods Mol. Biol. 558, 383–399 10.1007/978-1-60761-103-5_22 [DOI] [PubMed] [Google Scholar]
- Scherthan H., Trelles-Sticken E. (2008). Absence of yKu/Hdf1 but not myosin-like proteins alters chromosome dynamics during prophase I in yeast. Differentiation 76, 91–98 [DOI] [PubMed] [Google Scholar]
- Scherthan H., Weich S., Schwegler H., Heyting C., Härle M., Cremer T. (1996). Centromere and telomere movements during early meiotic prophase of mouse and man are associated with the onset of chromosome pairing. J. Cell Biol. 134, 1109–1125 10.1083/jcb.134.5.1109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scherthan H., Jerratsch M., Dhar S., Wang Y. A., Goff S. P., Pandita T. K. (2000). Meiotic telomere distribution and Sertoli cell nuclear architecture are altered in Atm- and Atm-p53-deficient mice. Mol. Cell. Biol. 20, 7773–7783 10.1128/MCB.20.20.7773-7783.2000 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scherthan H., Sfeir A., de Lange T. (2011). Rap1-independent telomere attachment and bouquet formation in mammalian meiosis. Chromosoma 120, 151–157 10.1007/s00412-010-0295-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmitt J., Benavente R., Hodzic D., Höög C., Stewart C. L., Alsheimer M. (2007). Transmembrane protein Sun2 is involved in tethering mammalian meiotic telomeres to the nuclear envelope. Proc. Natl. Acad. Sci. USA 104, 7426–7431 10.1073/pnas.0609198104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sfeir A., Kabir S., van Overbeek M., Celli G. B., de Lange T. (2010). Loss of Rap1 induces telomere recombination in the absence of NHEJ or a DNA damage signal. Science 327, 1657–1661 10.1126/science.1185100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takai H., Smogorzewska A., de Lange T. (2003). DNA damage foci at dysfunctional telomeres. Curr. Biol. 13, 1549–1556 10.1016/S0960-9822(03)00542-6 [DOI] [PubMed] [Google Scholar]
- Vourc'h C., Taruscio D., Boyle A. L., Ward D. C. (1993). Cell cycle-dependent distribution of telomeres, centromeres, and chromosome-specific subsatellite domains in the interphase nucleus of mouse lymphocytes. Exp. Cell Res. 205, 142–151 10.1006/excr.1993.1068 [DOI] [PubMed] [Google Scholar]
- Yang C., Wang H., Xu Y., Brinkman K. L., Ishiyama H., Wong S. T., Xu B. (2012). The kinetochore protein Bub1 participates in the DNA damage response. DNA Repair (Amst.) 11, 185–191 10.1016/j.dnarep.2011.10.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu J. L., Lin S. L., Li M., Ouyang Y. C., Hou Y., Schatten H., Sun Q. Y. (2010). Septin2 is modified by SUMOylation and required for chromosome congression in mouse oocytes. Cell Cycle 9, 1607–1616 10.4161/cc.9.8.11463 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.