

Abstract
Background/Aims
Many women with polycystic ovary syndrome (PCOS) exhibit insulin resistance. Adipose tissue plays an important role in insulin resistance, and adipokines including tumor necrosis factor (TNF)-α and adiponectin are altered in PCOS. Insulin-like growth factor binding protein-3 (IGFBP-3), alone or in conjunction with other adipokines, is also associated with insulin resistance. We evaluated the effects of TNF-α, adiponectin, and IGFBP-3 on insulin sensitivity and the relationships among these proteins in women with PCOS.
Methods
We recruited 40 women with PCOS and 40 age- and body mass index (BMI)-matched regular cycling women (controls). The women were divided into obese (BMI ≥ 25 kg/m2) and nonobese (BMI < 25 kg/m2) groups. Anthropometric measurements were performed, and serum levels of TNF-α, adiponectin, and IGFBP-3 were determined. Insulin sensitivity was estimated using the metabolic clearance rate (MCR) of glucose calculated from the oral glucose tolerance test.
Results
Serum levels of TNF-α and IGFBP-3 did not differ between the PCOS and control groups, but adiponectin levels in the PCOS group were lower than those in control women in the nonobese group (p < 0.05). TNF-α, adiponectin, and IGFBP-3 levels were not correlated with each other in women with PCOS, but a significant positive correlation was observed between adiponectin levels and MCR (p < 0.05). Multiple regression analysis revealed that adiponectin levels were significantly associated with insulin sensitivity (p < 0.05) in women with PCOS.
Conclusions
IGFBP-3 and TNF-α levels were not associated with insulin sensitivity, but adiponectin levels were related to insulin sensitivity in women with PCOS.
Keywords: Adiponectin, Tumor necrosis factor-alpha, Insulin-like growth factor binding protein 3, Polycystic ovary syndrome
INTRODUCTION
Polycystic ovary syndrome (PCOS) is a heterogeneous condition characterized by hyperandrogenic chronic anovulation and is the most common endocrinopathy in women of reproductive age [1]. The fundamental defect that causes PCOS remains elusive, but the pathogenesis is likely multifactorial in origin. Insulin resistance and compensatory hyperinsulinemia are important in the pathogenesis of PCOS [2,3]. Although the cellular and molecular mechanisms of insulin resistance in patients with PCOS have been investigated extensively, the exact mechanisms remain unknown.
Insulin resistance in patients with PCOS is partially associated with obesity and free fatty acids (FFAs), which reduce the action of insulin in target tissues via serine phosphorylation of the insulin receptor [4]. Serine phosphorylation contributes to androgen over-production in the ovaries. Adipose tissue dysfunction may play a role in insulin resistance, and some adipokines, including adiponectin and tumor necrosis factor (TNF)-α, are associated with insulin resistance in patients with PCOS [5,6]. Adiponectin is down-regulated in obese subjects and may have both anti-inflammatory and insulin-sensitizing effects [7]. A recent systematic review reported that women with PCOS have lower adiponectin levels [6]. Adiponectin has direct insulin-sensitizing effects. Therefore, decreased adiponectin levels could contribute to insulin resistance in women with PCOS. TNF-α has proinflammatory properties, resulting in insulin resistance in adipocytes. TNF-α stimulates lipolysis in adipose tissue, and the resulting increase in FFAs and serine phosphorylation are directly involved in the development of insulin resistance and hyperandrogenism [4]. TNF-α is elevated in women with PCOS [5], but the exact pathogenetic role of TNF-α has not been elucidated.
The insulin-like growth factor binding proteins (IGFBPs) are a family of six binding proteins that bind insulin-like growth factor (IGF) to regulate IGF bioavailability [8]. IGFBPs may also exert intrinsic bioactivity without triggering IGF receptor signaling. IGFBP-1 and IGFBP-3 are likely involved in glucose homeostasis [9]. IGFBP-3 colocalizes with the retinoid X receptor (RXR)-α transcription factor, which is important in glucose homeostasis [10]. Therefore IGFBP-3 may alter glucose homeostasis. RXR-α is an important binding partner for peroxisome proliferator-activated receptor γ (PPARγ), which is a nuclear protein involved in the transcriptional regulation of a variety of glucose and lipid metabolism enzymes. PPARγ also plays several roles in the insulin resistance of women with PCOS [11]. Consequently, insulin resistance in women with PCOS may be associated with IGFBP-3. However, few studies have investigated the relationship between IGFBP-3 and insulin resistance in women with PCOS.
Increased TNF-α levels in adipose tissue inhibit adiponectin gene expression and stimulate IGFBP-3 expression, and IGFBP-3 may inhibit adiponectin expression in the basal state and following PPARγ agonist stimulation [12-15]. These results suggest that insulin resistance in women PCOS is associated with TNF-α, adiponectin, and IGFBP-3. Therefore, we evaluated the effects of TNF-α, adiponectin, and IGFBP-3 on insulin sensitivity and the relationships among these proteins in women with PCOS.
METHODS
Subjects
We recruited 40 women with PCOS from the endocrinology and gynecology clinic at Ewha Womans University Mokdong Hospital. The PCOS diagnosis was based on the National Institute of Child Health and Human Disease criteria [16]. Chronic anovulation was defined as less than eight cycles per year. Hyperandrogenemia was defined as a total testosterone or free testosterone level above the 95th percentile (total testosterone ≥ 67 ng/dL or free testosterone ≥ 0.84 ng/dL) of detected testosterone levels in 1,120 healthy regular cycling women [17]. Other androgen excess disorders (e.g., congenital adrenal hyperplasia) were excluded accordingly. Forty age-matched and, body mass index (BMI)-matched regular cycling women were recruited for the control group, and none of these women had a family history of diabetes or PCOS. Subjects treated with drugs known to influence insulin sensitivity (e.g., steroids, oral contraceptives, metformin, or thiazide diuretics) were excluded. Written informed consent was obtained from all participants, and the Institutional Review Board of Ewha Womans University Mokdong Hospital approved this study.
Methods
The height and weight of all subjects were measured, and BMI was calculated as weight (kg)/height (m)2. A venous blood sample was collected after an overnight fast of at least 8 hours when subjects were in the early follicular phase. The blood was sampled independently of menstrual cycle stage in amenorrheic women.
TNF-α, adiponectin, and IGFBP-3 levels were measured using a radioimmunoassay kit (DPC, Los Angeles, CA, USA). Total cholesterol (TC), high density lipoprotein cholesterol (HDL-C), and triglycerides (TG) were measured using an enzymatic assay and an automated analyzer (Hitachi 7150 Autochemistry Analyzer, Hitachi, Tokyo, Japan). Total testosterone levels were measured using a commercial chemiluminescent immunoassay kit (Siemens, New York, NY, USA), and sex hormone-binding globulin (SHBG) levels were measured by immunoradiometric assay using a commercial kit (DPC). Free testosterone levels were calculated from total testosterone, SHBG, and albumin levels in the same sample from each subject using the International Society for Study of the Aging Male formula (http://www.issam.ch/freetestos.htm) [17].
The 75-g oral glucose tolerance test (OGTT) was performed the morning after an overnight fast. A polyethylene catheter was placed in the antecubital vein prior to the test. Venous blood samples were drawn at baseline after 30 minutes of supine rest and at 30, 60, 90, and 120 minutes after the 75-g glucose load was given. Plasma glucose levels were measured using the glucose oxidase method (Beckman Model Glucose Analyzer 2, Beckman Coulter, Fullerton, CA, USA), and insulin levels were measured using a commercial radioimmunoassay kit (Biosource, Nivelles, Belgium). Insulin sensitivity was estimated using the metabolic clearance rate (MCR) of glucose calculated from the OGTT (MCRest-OGTT, MCR) according to the following formula: MCR = 18.8 - 0.271 × BMI - 0.0052 × I120 - 0.27 × G90 (I120, postload 120 minutes insulin; G90, postload 90 minutes glucose) [18]. The euglycemic-hyperinsulinemic clamp technique is the gold standard for measuring insulin sensitivity, but it is difficult to perform in practice. Therefore, we used MCR as a marker of insulin sensitivity because it is significantly correlated with the M value from the euglycemic clamp [18].
The statistical evaluation was performed using the SPSS 18.0 software package for Windows (IBM Co., Armonk, NY, USA). Quantitative variables are reported as means ± standard deviation. The Kolmogorov-Smirnov test was used to analyze continuous variables for normality, and logarithmic transformations were applied as needed to ensure a normal distribution for the skewed variables. Comparisons of two groups with different parameters were performed using Student unpaired t test. Pearson's correlation test was used to examine the correlation between adipokines and metabolic parameters. A multiple regression analysis was performed to identify variables that independently predicted insulin resistance. Two-tailed p values < 0.05 were considered to indicate significance.
RESULTS
Women with PCOS and the healthy controls were divided into the nonobese (BMI < 25 kg/m2) and obese (BMI ≥ 25 kg/m2) groups based on their BMI. The obesity criteria were based on the Asia-Pacific criteria [19]. Nonobese and obese women with PCOS were of comparable age and BMI to women in the control group.
The clinical and biochemical characteristics of women with PCOS and the controls are shown in Table 1. Total testosterone levels (p < 0.001), free testosterone levels (p < 0.001), 2-hour postload glucose (p < 0.01), and fasting insulin (p < 0.01) were higher, and SHBG levels (p < 0.001) were lower in women with PCOS than those in controls. Women with PCOS exhibited a lower MCR (p < 0.001) and a higher 2-hour postload insulin (p < 0.001) than those in the controls in the nonobese group, and higher TC (p < 0.05) and TG (p < 0.05) levels in the obese group than those in the nonobese group.
Table 1.
Clinical and biochemical characteristics of women with polycystic ovary syndrome and controls
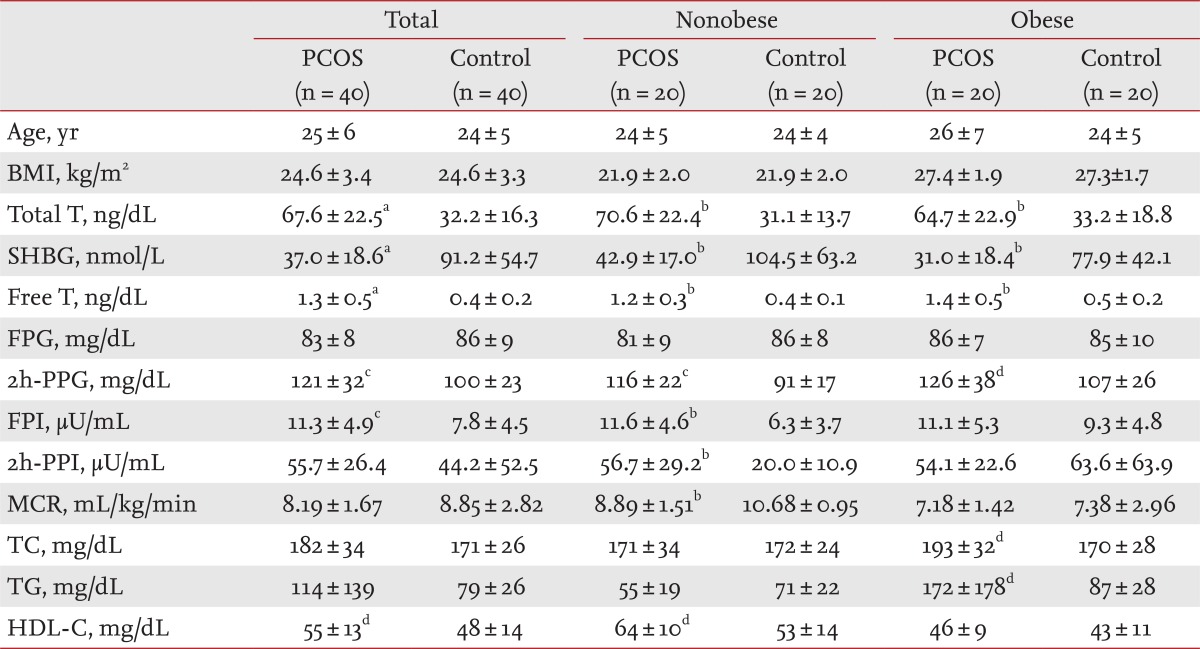
Values are presented as mean ± SD.
PCOS, polycystic ovary syndrome; BMI, body mass index; T, testosterone; SHBG, sex hormone binding globulin; FPG, fasting plasma glucose; PPG, postload plasma glucose; FPI, fasting plasma insulin; PPI, postload plasma insulin; MCR, metabolic clearance rate; TC, total cholesterol; TG, triglycerides; HDL-C, high density lipoprotein cholesterol.
ap < 0.0001 vs. control.
bp < 0.001 vs. control.
cp < 0.01 vs. control.
dp < 0.05 vs. control.
IGFBP-3 and TNF-α levels were comparable between the PCOS and control groups. Serum TNF-α and IGFBP-3 levels did not differ between women with PCOS and the controls after the subjects were divided into obese and nonobese groups (Table 2). Adiponectin levels tended to be lower in women with PCOS than those in the controls (p = 0.079) (Table 2). Adiponectin levels in the nonobese group were significantly lower in women with PCOS than those in the controls (p < 0.05) (Table 2).
Table 2.
Insulin like growth factor binding protein-3, tumor necrosis factor-α, and adiponectin levels in women with polycystic ovary syndrome and controls according to obesity status
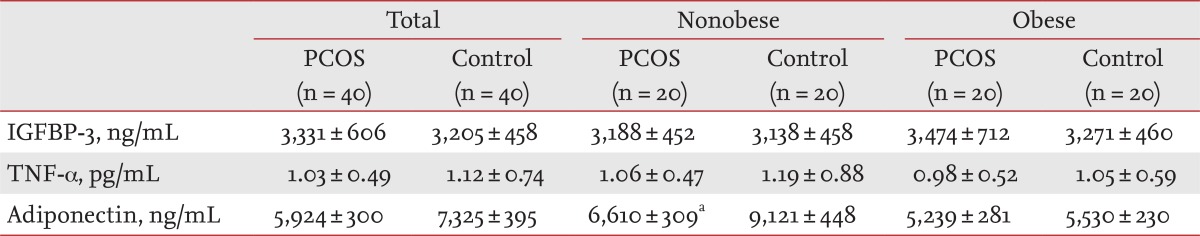
Values are presented as mean ± SD.
PCOS, polycystic ovary syndrome; IGFBP-3, insulin like growth factor binding protein-3; TNF, tumor necrosis factor.
ap < 0.05 vs. nonobese controls.
Adiponectin levels significantly correlated with MCR (r = 0.385, p < 0.05) in women with PCOS, but TNF-α and IGFBP-3 levels were not significantly correlated with MCR or lipid levels (Table 2). Adiponectin levels in controls correlated with MCR (r = 0.475, p < 0.01), HDL-C (r = 0.480, p < 0.01), BMI (r = -0.435, p < 0.01), and TG (r = -0.348, p < 0.05), and IGFBP-3 was positively correlated with TG (r = 0.323, p < 0.05) (Table 3).
Table 3.
Correlation coefficients between serum tumor necrosis factor-α, adiponectin, and insulin-like growth factor binding protein-3 levels with insulin sensitivity and lipid levels in women with polycystic ovary syndrome and controls
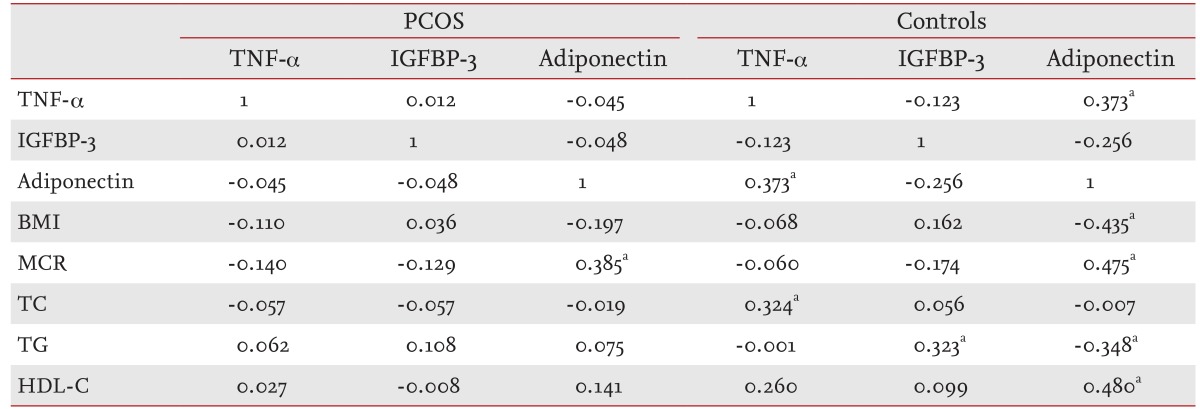
PCOS, polycystic ovary syndrome; TNF, tumor necrosis factor; IGFBP-3, insulin like growth factor binding protein-3; BMI, body mass index; MCR, metabolic clearance rate; TC, total cholesterol; TG, triglycerides; HDL-C, high density lipoprotein cholesterol.
ap < 0.05.
The multiple regression analysis demonstrated that BMI (p < 0.001) and adiponectin levels (p < 0.05) were significantly associated with MCR in women with PCOS, but that adiponectin levels were not associated with insulin sensitivity in controls (Table 4).
Table 4.
Multiple regression analysis of insulin sensitivity in women with polycystic ovary syndrome and controls
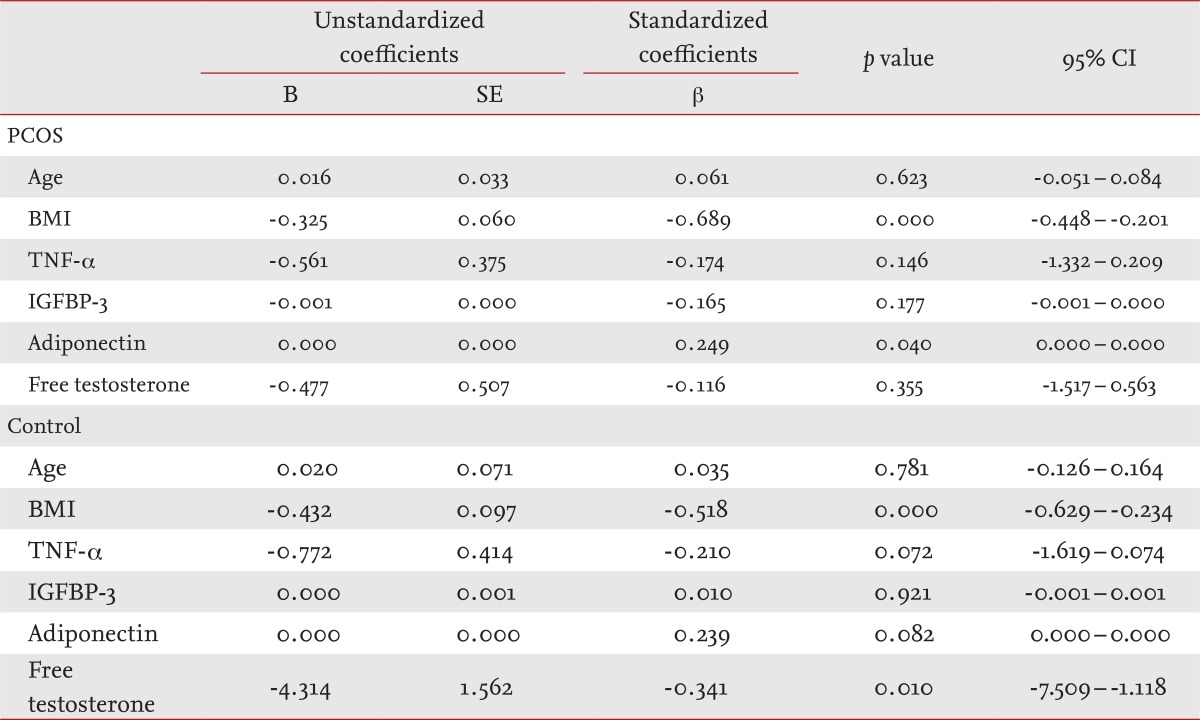
CI, confidence interval; PCOS, polycystic ovary syndrome; BMI, body mass index; TNF, tumor necrosis factor; IGFBP-3, insulin-like growth factor binding protein-3.
DISCUSSION
We determined the associations among serum TNF-α, adiponectin, and IGFBP-3 levels and insulin sensitivity in women with PCOS. TNF-α and IGFBP-3 levels were not associated with insulin sensitivity, but adiponectin levels were significantly associated with insulin sensitivity in women with PCOS.
PCOS is an important metabolic and reproductive disorder and is likely multifactorial in origin. Insulin resistance is important in the pathogenesis of PCOS, but the exact mechanisms of PCOS are unknown. The "central paradox" in PCOS is that the ovary remains sensitive to insulin-induced androgen production despite a systemic insulin-resistant state. The serine phosphorylation theory may explain this paradox. Serine phosphorylation of the insulin receptor inhibits tyrosine kinase activity of the receptor, and this defect in insulin-signaling causes insulin resistance in women with PCOS [3]. Serine phosphorylation of the main regulatory enzyme of androgen biosynthesis, P450c17, may modulate 17, 20 lyase activity and induce androgen production. This hypothesis suggests that a single kinase might phosphorylate the insulin receptor and P450c17 to produce hyperandrogenemia and hyperinsulinemia in women with PCOS [20].
Adipose tissue in women with PCOS is characterized by hypertrophic adipocytes and impaired lipolysis and insulin action. The expression and secretion of a wide variety of adipokines implicated in insulin resistance, including adiponectin and TNF-α, are also altered in women with PCOS [21].
Adiponectin exerts insulin-sensitizing properties by stimulating fatty acid oxidation and suppressing hepatic gluconeogenesis, and a dysregulation of adiponectin has been implicated in the pathogenesis of insulin resistance [22]. Previous studies have reported conflicting results; some groups documented a decrease in adiponectin levels in women with PCOS compared with those in BMI-matched controls, whereas other studies found no differences in adiponectin levels between these groups [23]. Adiponectin levels tended to be lower in women with PCOS than in the controls in our study, but the difference was not significant. However, adiponectin levels in nonobese women with PCOS were significantly lower than those in BMI-matched controls, and MCR was also significantly lower. These results suggest that the production of adiponectin was suppressed in the presence of obesity; however, lower adiponectin per se may be inherent to the pathogenesis of PCOS.
TNF-α is secreted primarily by adipose tissue macrophages, and exhibits proinflammatory properties. TNF-α inhibits tyrosine phosphorylation of the insulin receptor and insulin receptor substrate-1 (IRS-1) by inducing serine phosphorylation. The increased serine phosphorylation of IRS-1 by TNF-α impairs insulin signal transduction and promotes insulin resistance [4]. These mechanisms are similar to the insulin resistance in women with PCOS. Some controversy surrounds serum TNF-α levels in PCOS, but a recent meta-analysis of TNF-α in women with PCOS reported no difference in TNF-α levels between women with PCOS and controls [24], which was consistent with our data. TNF-α levels in our study did not differ between women with PCOS and the controls. Additionally, TNF-α levels were not associated with insulin sensitivity in women with PCOS or the controls. Therefore, TNF-α alone does not significantly contribute to the pathogenesis of insulin resistance.
IGFBP-3 is derived primarily from hepatic Kupffer cells but is produced by a wide variety of tissues, including adipocytes, where it is upregulated during adipocyte differentiation [25,26]. IGFBP-3 inhibits insulin-stimulated glucose uptake and Akt phosphorylation in human adipocytes to induce insulin resistance [27]. The activation of a phosphotyrosine phosphatase and subsequent dephosphorylation of the insulin receptor may contribute to IGFBP-3-related insulin resistance [28]. This mechanism suggests a relationship between PCOS and IGFBP-3. However, IGFBP-3 levels in our study did not differ between women with PCOS and the controls, and obesity did not influence IGFBP-3 levels. IGFBP-3 levels were also not associated with insulin resistance in women with PCOS or the controls.
IGFBP-3 levels are regulated by multiple factors, including several cytokines that have been implicated in insulin resistance such as TNF-α. TNF-α increases IGFBP-3 production in 3T3-L1 adipocytes, and IGFBP-3 antisense partially blocks the insulin-antagonistic effect of TNF-α in cultured adipocytes. These results suggest that the insulin antagonistic activity of TNF-α is partially mediated via induction of IGFBP-3 [12]. IGFBP-3 and TNF-α may inhibit transcription of adiponectin, which contributes to the pathogenesis of insulin resistance. IGFBP-3 also inhibits the thiazolidinedione-induced increase in adiponectin in 3T3-L1 mouse adipocytes [12].
These results suggest that the interaction among TNF-α, adiponectin, and IGFBP-3 contributes to the development of insulin resistance in women with PCOS. We evaluated the correlations among serum TNF-α, adiponectin, and IGFBP-3 levels, but no significant relationships were observed among these proteins in women with PCOS. However, serum TNF-α and adiponectin levels in control women were positively correlated. We measured serum TNF-α and adiponectin levels, which may not reflect TNF-α or adiponectin activity in tissues. Serum TNF-α levels are affected by many factors, and these levels may not reflect TNF-α activity in tissues. Only adiponectin levels were significantly correlated with the insulin sensitivity index in women with PCOS and the controls. The linear regression analysis revealed that adiponectin levels were significantly associated with insulin sensitivity only in women with PCOS but not in the controls. Therefore, decreased adiponectin levels may be closely related to the development of insulin resistance in women with PCOS.
This study is the first evaluation of the association between IGFBP-3 and insulin sensitivity in women with PCOS. IGFBP-3 levels were not associated with insulin sensitivity in women with PCOS or controls, but adiponectin levels were associated with insulin sensitivity in women with PCOS. Future studies to ascertain the relationships among TNF-α, adiponectin, and IGFBP-3 tissue levels and insulin sensitivity are required.
KEY MESSAGE
1. Adipose tissue plays an important role in insulin resistance and adipokines including tumor necrosis factor (TNF)-α and adiponectin are altered in polycystic ovary syndrome (PCOS).
2. Insulin-like growth factor binding protein-3 (IGFBP-3), alone or in conjunction with other adipokines, is also associated with insulin resistance.
3. IGFBP-3 and TNF-α levels were not associated with insulin sensitivity, but adiponectin levels were related to insulin sensitivity in Korean women with PCOS.
Footnotes
No potential conflict of interest relevant to this article is reported.
References
- 1.Dunaif A, Thomas A. Current concepts in the polycystic ovary syndrome. Annu Rev Med. 2001;52:401–419. doi: 10.1146/annurev.med.52.1.401. [DOI] [PubMed] [Google Scholar]
- 2.Dunaif A. Insulin resistance and the polycystic ovary syndrome: mechanism and implications for pathogenesis. Endocr Rev. 1997;18:774–800. doi: 10.1210/edrv.18.6.0318. [DOI] [PubMed] [Google Scholar]
- 3.Oh JY, Lee JA, Lee H, Sung YA, Chung H. Serum C-reactive protein levels in normal-weight polycystic ovary syndrome. Korean J Intern Med. 2009;24:350–355. doi: 10.3904/kjim.2009.24.4.350. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ruan H, Lodish HF. Insulin resistance in adipose tissue: direct and indirect effects of tumor necrosis factor-alpha. Cytokine Growth Factor Rev. 2003;14:447–455. doi: 10.1016/s1359-6101(03)00052-2. [DOI] [PubMed] [Google Scholar]
- 5.Tarkun I, Cetinarslan B, Turemen E, Canturk Z, Biyikli M. Association between circulating tumor necrosis factor-alpha, interleukin-6, and insulin resistance in normal-weight women with polycystic ovary syndrome. Metab Syndr Relat Disord. 2006;4:122–128. doi: 10.1089/met.2006.4.122. [DOI] [PubMed] [Google Scholar]
- 6.Toulis KA, Goulis DG, Farmakiotis D, et al. Adiponectin levels in women with polycystic ovary syndrome: a systematic review and a meta-analysis. Hum Reprod Update. 2009;15:297–307. doi: 10.1093/humupd/dmp006. [DOI] [PubMed] [Google Scholar]
- 7.Weyer C, Funahashi T, Tanaka S, et al. Hypoadiponectinemia in obesity and type 2 diabetes: close association with insulin resistance and hyperinsulinemia. J Clin Endocrinol Metab. 2001;86:1930–1935. doi: 10.1210/jcem.86.5.7463. [DOI] [PubMed] [Google Scholar]
- 8.Firth SM, Baxter RC. Cellular actions of the insulin-like growth factor binding proteins. Endocr Rev. 2002;23:824–854. doi: 10.1210/er.2001-0033. [DOI] [PubMed] [Google Scholar]
- 9.Murphy LJ. The role of the insulin-like growth factors and their binding proteins in glucose homeostasis. Exp Diabesity Res. 2003;4:213–224. doi: 10.1155/EDR.2003.213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Liu B, Lee HY, Weinzimer SA, et al. Direct functional interactions between insulin-like growth factor-binding protein-3 and retinoid X receptor-alpha regulate transcriptional signaling and apoptosis. J Biol Chem. 2000;275:33607–33613. doi: 10.1074/jbc.M002547200. [DOI] [PubMed] [Google Scholar]
- 11.Toth B, Hornung D, Scholz C, Djalali S, Friese K, Jeschke U. Peroxisome proliferator-activated receptors: new players in the field of reproduction. Am J Reprod Immunol. 2007;58:289–310. doi: 10.1111/j.1600-0897.2007.00514.x. [DOI] [PubMed] [Google Scholar]
- 12.Kim HS, Ali O, Shim M, et al. Insulin-like growth factor binding protein-3 induces insulin resistance in adipocytes in vitro and in rats in vivo. Pediatr Res. 2007;61:159–164. doi: 10.1203/pdr.0b013e31802d8a30. [DOI] [PubMed] [Google Scholar]
- 13.Besset V, Le Magueresse-Battistoni B, Collette J, Benahmed M. Tumor necrosis factor alpha stimulates insulin-like growth factor binding protein 3 expression in cultured porcine Sertoli cells. Endocrinology. 1996;137:296–303. doi: 10.1210/endo.137.1.8536626. [DOI] [PubMed] [Google Scholar]
- 14.Rozen F, Zhang J, Pollak M. Antiproliferative action of tumor necrosis factor-alpha on MCF-7 breastcancer cells is associated with increased insulin-like growth factor binding protein-3 accumulation. Int J Oncol. 1998;13:865–869. doi: 10.3892/ijo.13.4.865. [DOI] [PubMed] [Google Scholar]
- 15.Rajah R, Lee KW, Cohen P. Insulin-like growth factor binding protein-3 mediates tumor necrosis factor-alpha-induced apoptosis: role of Bcl-2 phosphorylation. Cell Growth Differ. 2002;13:163–171. [PubMed] [Google Scholar]
- 16.Fauser BC, Tarlatzis BC, Rebar RW, et al. Consensus on women's health aspects of polycystic ovary syndrome (PCOS): the Amsterdam ESHRE/ASRM-Sponsored 3rd PCOS Consensus Workshop Group. Fertil Steril. 2012;97:28–38. doi: 10.1016/j.fertnstert.2011.09.024. [DOI] [PubMed] [Google Scholar]
- 17.Vermeulen A, Verdonck L, Kaufman JM. A critical evaluation of simple methods for the estimation of free testosterone in serum. J Clin Endocrinol Metab. 1999;84:3666–3672. doi: 10.1210/jcem.84.10.6079. [DOI] [PubMed] [Google Scholar]
- 18.Stumvoll M, Mitrakou A, Pimenta W, et al. Use of the oral glucose tolerance test to assess insulin release and insulin sensitivity. Diabetes Care. 2000;23:295–301. doi: 10.2337/diacare.23.3.295. [DOI] [PubMed] [Google Scholar]
- 19.World Health Organization, Regional Office for the Western Pacific; International Association for the Study of Obesity; International Obesity Task Force. The Asia-Pacific Perspective: Redefining Obesity and Its Treatment. Melbourne: Health Communications Australia; 2000. [Google Scholar]
- 20.Bremer AA, Miller WL. The serine phosphorylation hypothesis of polycystic ovary syndrome: a unifying mechanism for hyperandrogenemia and insulin resistance. Fertil Steril. 2008;89:1039–1048. doi: 10.1016/j.fertnstert.2008.02.091. [DOI] [PubMed] [Google Scholar]
- 21.Villa J, Pratley RE. Adipose tissue dysfunction in polycystic ovary syndrome. Curr Diab Rep. 2011;11:179–184. doi: 10.1007/s11892-011-0189-8. [DOI] [PubMed] [Google Scholar]
- 22.Festa A, D'Agostino R, Jr, Howard G, Mykkanen L, Tracy RP, Haffner SM. Chronic subclinical inflammation as part of the insulin resistance syndrome: the Insulin Resistance Atherosclerosis Study (IRAS) Circulation. 2000;102:42–47. doi: 10.1161/01.cir.102.1.42. [DOI] [PubMed] [Google Scholar]
- 23.Wickham EP, 3rd, Cheang KI, Clore JN, Baillargeon JP, Nestler JE. Total and high-molecular weight adiponectin in women with the polycystic ovary syndrome. Metabolism. 2011;60:366–372. doi: 10.1016/j.metabol.2010.02.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Escobar-Morreale HF, Luque-Ramirez M, Gonzalez F. Circulating inflammatory markers in polycystic ovary syndrome: a systematic review and metaanalysis. Fertil Steril. 2011;95:1048–1058. doi: 10.1016/j.fertnstert.2010.11.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Boney CM, Moats-Staats BM, Stiles AD, D'Ercole AJ. Expression of insulin-like growth factor-I (IGF-I) and IGF-binding proteins during adipogenesis. Endocrinology. 1994;135:1863–1868. doi: 10.1210/endo.135.5.7525256. [DOI] [PubMed] [Google Scholar]
- 26.Wabitsch M, Heinze E, Debatin KM, Blum WF. IGF-I- and IGFBP-3-expression in cultured human preadipocytes and adipocytes. Horm Metab Res. 2000;32:555–559. doi: 10.1055/s-2007-978685. [DOI] [PubMed] [Google Scholar]
- 27.Chan SS, Twigg SM, Firth SM, Baxter RC. Insulin-like growth factor binding protein-3 leads to insulin resistance in adipocytes. J Clin Endocrinol Metab. 2005;90:6588–6595. doi: 10.1210/jc.2005-0595. [DOI] [PubMed] [Google Scholar]
- 28.Ricort JM, Binoux M. Insulin-like growth factor-binding protein-3 activates a phosphotyrosine phosphatase: effects on the insulin-like growth factor signaling pathway. J Biol Chem. 2002;277:19448–19454. doi: 10.1074/jbc.M200439200. [DOI] [PubMed] [Google Scholar]