

Abstract
Ants are efficient navigators, guided by path integration and visual landmarks. Path integration is the primary strategy in landmark-poor habitats, but landmarks are readily used when available. The landmark panorama provides reliable information about heading direction, routes and specific location. Visual memories for guidance are often acquired along routes or near to significant places. Over what area can such locally acquired memories provide information for reaching a place? This question is unusually approachable in the solitary foraging Australian jack jumper ant, since individual foragers typically travel to one or two nest-specific foraging trees. We find that within 10 m from the nest, ants both with and without home vector information available from path integration return directly to the nest from all compass directions, after briefly scanning the panorama. By reconstructing panoramic views within the successful homing range, we show that in the open woodland habitat of these ants, snapshot memories acquired close to the nest provide sufficient navigational information to determine nest-directed heading direction over a surprisingly large area, including areas that animals may have not visited previously.
Keywords: visual homing, navigation, differential GPS, landmark panorama, image difference function, Myrmecia croslandi
1. Introduction
Ants are central place foragers and use two navigational strategies, path integration and landmark guidance, to move between nest and foraging sites [1]. Whether animals are guided by landmarks or by path integration (PI) depends on the navigational information content of the habitat and on individual experience. In landmark-poor habitats, ants rely predominantly on path integration [2–4] to determine goal directions, whereas in landmark-rich and visually structured environments they are guided by dominant landmarks [5–7] and the landmark panorama [8]. When the information provided by the landmark panorama and the path integrator is made to conflict experimentally, landmark guidance either overrides conflicting path integration information [7,9–12] or ants follow a compromise direction half-way between that indicated by the landmark panorama and that indicated by the path integrator [7,13,14].
Visual landmark information for both heading direction (visual compass) and for location in space needs to be learnt. Landmark memories are thus place- and route-dependent [15–17] and therefore appear to provide animals with navigational knowledge that is limited to areas in the environment they have visited before ([18]; but see [19]).
The effective range of navigational information acquired in a restricted area around a nest or along a route, however, has rarely been mapped. Baerends [15], for instance, found that Ammophila wasps have idiosyncratic knowledge of their nest environment, taking very different routes back to the nest from geographically close locations. This clearly indicated not only that the wasps had been to various places, but also that they had not learnt their true relative positions, as they would be represented in a topographic map. Recently, Wystrach et al. [20] restricted the experience of ant foragers to a narrow corridor and displaced ants to locations outside this familiar territory. They recorded the initial, quite well-directed heading directions of ants at these locations, but unfortunately not their entire paths, which may have indicated the range over which the ants are guided by navigational information acquired in a restricted area. Honeybees find home from displacement distances of up to 10 km away from their hive location [21] and from up to 100 m distance in an unfamiliar environment in which they were allowed to perform one orientation flight [22]. However, because the flight paths of honeybees at that time could not be recorded (see, however, [23]), it remains unclear where animals had been before and to what extent homing success was based on extended search flights. Menzel et al. [19] discovered that bees caught at feeders and released at random locations around the hive, first flew in the home direction indicated by their path integrator, they then engaged in slow search flights and at some point took a straight line back to the hive. Finally, Wehner et al. [12] displaced Cataglyphis fortis foragers that had returned to the nest from a feeder 30 m northeast of the nest, to release locations 30 m northwest, southwest and southeast of the nest. After extended local search, many of these ants were able to home successfully (fig. 6 in [12]).
In two recent studies, homing ants were displaced a few metres away from their foraging routes and released at locations where they had clearly not been before. Yet, many ants were able to correctly identify their location relative to the nest and to head home [7,24]. This raises the question of the range over which the navigational knowledge of individual animals allows them to home from novel locations. We addressed these issues by systematically mapping, in the solitary foraging Australian ant Myrmecia croslandi, the area around the nests from which individuals are able to successfully home.
2. Material and methods
(a). Study animals
Myrmecia croslandi Taylor, commonly known as Jack jumpers, are endemic to Australia. These ants are solitary foragers and exhibit no evidence of recruitment. For ants, they have an unusually well-developed visual system with nearly 2400 facets in each eye [25,26]. They are diurnal ants, active between the end of October to early May [27], feed on sugary extrusions of plant-sucking insects and hunt insects to feed their larvae. We studied ants from five different nests in an urban park in Canberra, Australia (35°15′05.59″ S, 149°09′33.18″ E) over a period of 3 years.
(b). Displacement experiments and tracking
At each nest, we followed individual ants that left for foraging, caught them in foam-stoppered Perspex tubes at the base of their foraging tree and provided them with 10 per cent sugar solution and live insect prey. Animals were transferred in the dark to different release locations. We tracked ants in two ways. We either moved the rover antenna of a differential global positioning system (DGPS; see below) directly behind the walking ant, making sure that the observer who held the antenna was not obstructing the ant's path, or in most cases, we placed coloured flags every 5 s or 20 cm behind a walking ant, again taking care not to disturb the ants’ progress. We later used the DGPS antenna to record the flag-marked path.
Ant paths were recorded with a DGPS (NovAtel Inc., Canada), including a base station antenna (GPS-702-GG L1/L2, GPS plus GLONASS), a base station receiver (FLEXPAK-V2-L1L2-G GPS plus GLONASS RT-2), a rover antenna (ANT-A72GLA-TW-N (532-C)) and a rover receiver (OEMV-2-RT2-G GPS plus GLONASS). In a DGPS, a stationary reference or base station calculates corrections for a mobile rover antenna, the position of which can then be determined with centimetre accuracy at least on a local scale [28], in our case in an area of 100 m radius. The stationary base station electronics and antenna were mounted on a tripod and set to integrate antenna position readings over 30 min. The rover receiver electronics were carried on a backpack and connected to the rover antenna, which was mounted at the end of a long, hand-held stick, so that it could be moved close to the ground behind an ant or along a flag-marked path. The base station and rover communicate through a radio link, allowing the exchange of corrections that provide position accuracy of the rover antennae of better than 10 cm. Northing, Easting and Height coordinates in metres, together with the standard deviations of position error estimates, were recorded and monitored at 1 s intervals with a laptop computer and extracted with a custom-written Matlab program (Matlab, Natick, MA, USA). Tracking was paused whenever the error standard deviations for Northing and Easting became larger than 10 cm. We used GPS Import File Converter and GPS Utility software (GPS Utility Ltd, UK) to register data with aerial photographs (kindly provided by ACTPLA, Canberra, Australia).
(c). Criteria for start of search
To identify the start of search, we measured the cumulative distance travelled from the release point. The location where this distance decreased for more than four consecutive sampling points (corresponding to 40–60 cm path length) was identified as the start of search. We defined the distance from the release point to the start of search as the distance travelled under the guidance of information provided by the path integrator.
(d). Filming initial path directions
Ants were released on a round wooden platform (40 cm diameter) that was raised 15 cm off the ground on aluminium pegs at three release stations around one of the nests. The platform had a circular hole in the centre, into which a black nylon sleeve casing including a foam-stoppered Perspex catching tube fitted tightly. Upon release, the foam stopper of the tube was replaced by a flat piece of cardboard with a central hole of 5 mm diameter, through which the ants could reach the platform. We filmed the ants on the platform, including information about true north and correct homing direction, using a Sony Handycam (HDR-CX550VE, Sony Corp., Japan), a Panasonic Lumix (DMC-FZ200, Panasonic Corp., Osaka, Japan) or an Optronis (CR600x2, Kehl, Germany) mounted on a tripod. Video footage was subjected to a frame-by-frame analysis that determined head position at 40 or 50 ms intervals, using a custom-written Matlab program (Jan Hemmi & Robert Parker, The Australian National University) or GraphClick (Arizona Software, USA). Some, but not all ants were released at all three release stations.
We placed a Sony Bloggie camera (MHS-PM5, Sony Corp., Japan) on the release platform (15 cm off the ground) to record panoramic views at nest sites, at release sites and at defined locations at different distances and directions from the nests.
(e). Calculating rotational image difference functions
Concentric panoramic images were unwarped to rectangular panoramas, measuring 2161 × 338 pixels, corresponding to a field of view of 360° × 56°, with a resolution of 6 pixels per degree, using a custom-written Matlab program. Sun glare and reflection artefacts in the sky were removed by using the colour replacement tool in Corel Photo Paint X5 (Corel Corporation, Ottawa, Canada) to copy adjacent sky patches into the corrupted areas. Eight-bit greyscale images were converted to floating point arrays, low-pass filtered with a 18 × 18 pixel Gaussian filter with σ = 6 pixels, before rotational image difference functions (rotIDFs) were determined using the Matlab circshift function. For each 1 pixel shift, the pixel differences were calculated between the reference image and the shifted image, resulting in 2161 × 338 values that were squared and averaged. For each image shift, we then calculated the root mean squared pixel difference (for details, see [29,30]).
3. Results
Foragers of M. croslandi from a single nest typically head towards a specific tree at a distance of 6–15 m from the nest (figure 1), which they climb to hunt for arthropod prey and to harvest honeydew exuded by plant-sucking insects. We caught such outbound foragers at the base of their foraging trees and fed them. These ants thus were motivated to return to their nest and can be assumed to have access to home vector information computed by their path integration system. We refer to these ants as full-vector ants.
Figure 1.
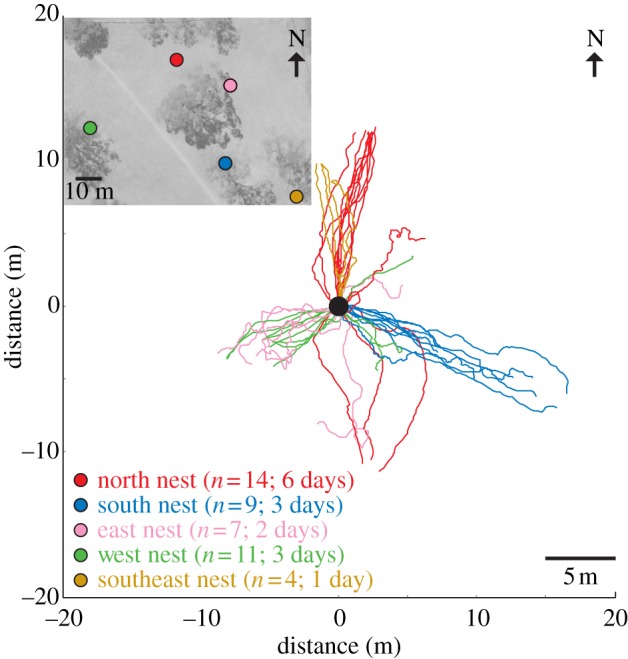
Outbound paths of ants from five nests. Inset: The relative location of the nests in an aerial photograph of the study area. Ant paths were tracked from the nest to the base of foraging trees or until an ant found food. Paths have been aligned such that the nest location is at the centre of origin of the coordinate system (black circle). Colours of ant tracks are nest-specific and numbers refer to the number of tracked animals and the days of observation.
(a). Homing from different compass directions around the nest
We tested the homing abilities of full-vector ants by releasing them at randomly chosen locations in eight compass directions at a distance of 10 m from the nest. Upon release, ants briefly look around and with few exceptions head directly home from all directions, ignoring information from the path integrator (figure 2a,b; red paths). Each ant was captured just before she entered the nest and was released a second time, now as a zero-vector ant, at a point opposite to the first release site (figure 2a,c; blue paths). These ants headed directly home from any location around the nest without the need for search. To identify how quickly ants can determine homing direction, we released 34 ants (12 as full-vector ants) from one nest at 10 m distance from the nest in three compass directions and filmed their initial paths on 40 cm diameter release platforms (figure 3). Typically, ants emerge, look around briefly and in most cases exit the release platform in approximately the nest direction within less than 30 s (see figure 3 and electronic supplementary material, figure S1). This result is surprising since ants with vector information when displaced to unfamiliar locations would be expected to follow their home vector at least for some distance and to eventually search [9], whereas ants without vector information would be expected to be disoriented and to search immediately upon release [12,31,32].
Figure 2.

Mapping the navigational knowledge of Myrmecia croslandi ants. (a) Example homing paths of full-vector (red) and zero-vector (blue) ants from three nests (circles: black, yellow and green) Foragers from each nest were caught at the base of their favourite tree (stars: black, yellow and green) and were transferred in the dark to one of eight randomly selected release stations (open circles) at 10 m distance from the nest. Full-vector ants (red paths) were tracked until they arrived close to the nest and were captured and re-released as zero-vector ants (blue paths) at a release station opposite to the first one. Summary of paths of (b) full-vector (red) and (c) zero-vector ants (blue) from four nests. Paths are rotated to align the nest (black circle) and release location (grey circle).
Figure 3.
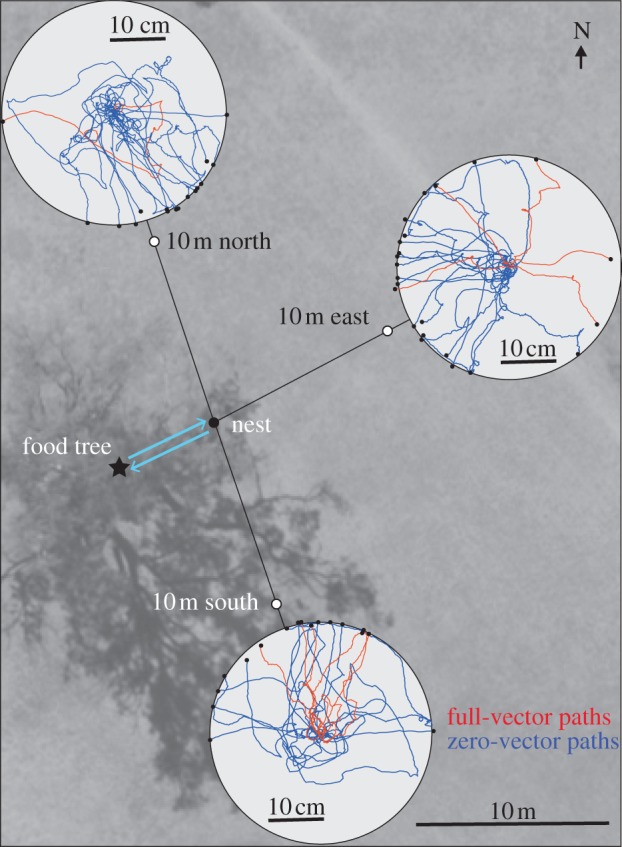
The initial paths of 12 full-vector (red) and 22 zero-vector ants (blue) after release on 40 cm diameter platforms (light grey circles) at three release stations (white circles), 10 m north, south and east from a nest (black dot). Full-vector ants were caught at the base of the nest-specific food tree (black star). Note that some, but not all ants were released at all three stations.
The full-vector ants in our experiments experienced different degrees of mismatch between the information supplied by the path integrator and by the landmark panorama, depending on the release point bearing relative to the nest and relative to the normal foraging corridor towards the food tree. Most ants, however, appear not to be affected by the degree of mismatch (see the electronic supplementary material, figure S2). Of particular interest are ants for which the home vector direction pointed either to the right or to the left of the true home direction (see the electronic supplementary material, figure S2a, second and third panel). We found no difference between the number of ants that chose to travel towards the left or towards the right of a narrow band from the release location to the nest (χ2 = 0.05247, p = 0.81), further indicating the extreme reliance on landmark information in these ants.
(b). Navigational knowledge of individual ants
To map the area around nests from which ants are able to home more extensively, we displaced ants captured at the foraging tree first as full-vector ants to a randomly selected release point and then as zero-vector ants to at least three additional release points. Each ant was clearly able to identify the general home direction from everywhere around the nest, and we document this for four cases in figure 4. It is noteworthy, however, that ants repeatedly correct their heading direction, indicating that they make frequent navigational decisions as they make their way towards the nest. Interestingly, we found that the ants have great difficulty in pinpointing the nest entrance at close range, indicating that the information that accurately directs them over several metres is not sufficient for nest localization. The ants frequently miss the nest entrance by a few centimetres and they eventually find it after an extended search (A. Narendra & J. Zeil 2009–2013, personal observations). This indicates that in contrast to other ant species, nest-odour is unlikely to guide their final approach [33,34].
Figure 4.
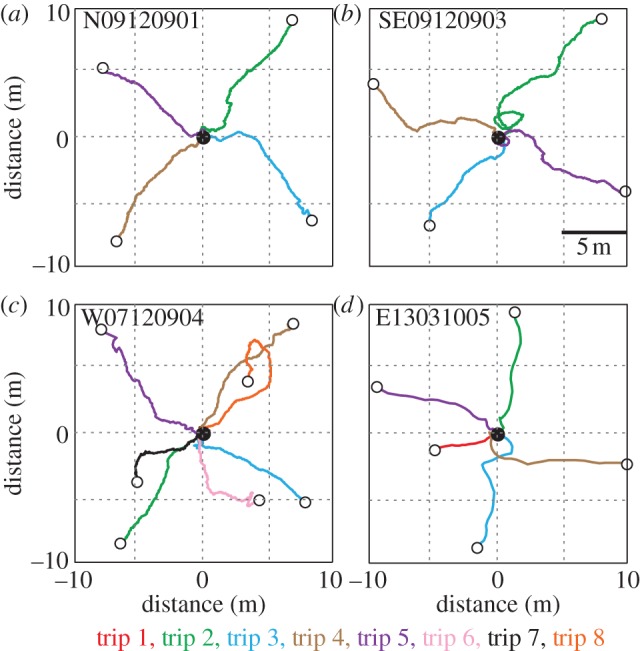
(a–b) Navigational knowledge of four ants from four nests. Ants captured either at their foraging tree (full-vector ants) or close to the nest when returning with prey (zero-vector ants) were displaced in different compass directions around the nest (black circle). Displaced ants were captured up to seven times after they had returned to the nest and re-released at different release stations (open circles) around the nest. For each individual, we show full-vector paths, if recorded, in red and subsequent zero-vector paths in different colours.
These results now raise two questions. First, do M. croslandi foragers path integrate at all? And second, what navigational information allows them to identify nest direction at locations they are very unlikely to have been to before?
(c). Homing from distant locations
To answer the first question, we displaced full-vector ants 15–25 m away from the nest. A majority of the ants (18 of 24) returned directly to the nest (figure 5a,b). At 1 m distance from the release, most of them were heading in the home vector direction (compare circular plots, figure 5a,b). At 10 m, however, the majority were on their way to the true nest location (figure 5b). The examples shown in figure 5c demonstrate that some ants were immediately guided by landmarks (yellow paths), some initially followed a direction intermediate between the home vector and the true nest direction, but subsequently corrected their heading direction (figure 5c, red paths) and some started searching after following the home vector direction (figure 5c, blue paths). We re-captured unsuccessful ants and released them at one of the eight release stations at a distance of 10 m from the nest. All ants returned home directly without searching (data not shown).
Figure 5.

Homing abilities of ants displaced to different compass directions at distances greater than 10 m. Paths of full-vector ants are rotated to align the release location at 0/0 with (a) the path integration direction and (b) with the true nest direction. The green paths are from three ants for which the path integration direction and the true nest direction were congruent. Insets show circular histograms of bearings at 1, 5 and 10 m distance from the release point, together with circular statistics. (c) Example paths of full-vector ants from one nest (blue circle) caught at the base of their food tree (blue star) and displaced to release stations (white circles) more than 10 m from the nest. Ants were immediately oriented to the nest (yellow), or initially relied on vector navigation and subsequently corrected their path to head home (red), or followed the home vector before beginning to search (blue). (d) Paths of full-vector ants released at a location further than 100 m from their nest. Paths are aligned with the home vector direction (arrow points towards the fictive nest direction) at the release point (grey circle).
The behaviour of the ants indicated that there is an area around the nest where the familiar landmark panorama overrides path integration information and that outside this area ants fall back to using that information. To explore this possibility, we displaced full-vector ants to an open field, more than 100 m east of the nest sites (figure 5d). Indeed, we find that most ants released at this remote location walked in directions roughly corresponding to the home vector direction. In most cases, however, ants started searching after having travelled less than the distance indicated by the path integrator (see the electronic supplementary material, figure S3). When re-captured and released at locations 10 m away from the nest, these ants returned to the nest more or less directly (data not shown). Foragers of M. croslandi thus do path integrate, but only use this information outside an area where the landmark panorama fails to provide guidance.
(d). Successful homing from different compass directions
To address the second question of what navigational information would allow ants to identify the direction to the nest, we suggest two hypotheses that could explain the ants’ navigational competence. One is the possibility that they possess detailed acquired knowledge of the nest environment within a radius of 10–15 m by having previously explored the area around the nest in all compass directions. However, we consider this unlikely for two reasons. First, most outbound trips we recorded at each of our nests were directed towards specific foraging trees located in a single compass direction (figure 1). Second, monitoring the movements of individual foragers from the first day of their foraging life reveals that ants head to a single tree during their entire lifetime, while only going on short excursions of up to 2 m distance from the nest in other compass directions (P. Jayatilaka 2011–2013, personal communication).
Our second hypothesis is that ants use panoramic snapshots taken at or near the nest to determine nest-heading direction from distances of up to 10–15 m from the nest. There are a number of proposed homing algorithms that could, in principle, achieve this [20,35–38]. All these algorithms operate on image differences and require, as a first step, the alignment of remembered and currently experienced views. So, at the most basic level, we can ask whether there is sufficient information in a comparison of views close to the nest and at the release sites and, in particular, whether there is a detectable minimum of image differences, by rotating oriented snapshots taken while pointing at the nest against the current scene. Graham et al. [39] (based on Müller & Wehner [40]) have suggested that these rotIDFs [29,30] on their own may provide sufficient information about home direction.
(e). The navigational information content of Myrmecia croslandi habitat
We find that at all release stations around two nests, the rotIDF of an appropriately oriented reference image taken at the nest location shows a detectable minimum, which points towards the nest. The depth of that minimum depends on the distance from the nest (see figure 6 for the nest at which we conducted the experiments shown in figure 3 and the electronic supplementary material, figure S4 for the nest location on top in figure 2a). In principle, then, ants could determine the home direction by detecting the direction of the rotIDF minimum at the release site [39]. They could in addition monitor their distance from the nest by tracking the change in the depth of that minimum, which is equivalent to a gradient descent on translational image differences [30,41].
Figure 6.

The information content of panoramic views as determined by rotIDFs (see §2 for details). (a) Image differences over orientation between a south-oriented panoramic image recorded at the nest (see bottom image on the right) and itself (red), and images recorded to the north of the nest looking south at different distances (0.5 m, green; 1 m, blue; 5 m, yellow; 10 m, black). (b) Same for a north-oriented snapshot at the nest and views recorded to the south of the nest looking north at different distances. Panoramic images are aligned such that nest direction is in the centre. (c) Same for a west-oriented snapshot at the nest and the view at the distant release site further than 100 m east of the nest area (black line and matching frame on right). Blue, rotIDF for a horizontal slice stretching from 22° to 43° above the horizon; green, rotIDF for a horizontal slice stretching from 0° to 22° above the horizon.
The comparison between the nest view and the view at the distant release site, where all full-vector ants walked towards the direction indicated by path integration, is interesting (figure 6c): there is still a broad minimum when the two views are aligned with the same compass direction (west in this case, black lines in figure 6c), however, this is largely due to the brightness distribution of the sky. The rotIDF shows a broad, deep minimum when image differences are calculated for a horizontal slice stretching from 22° to 43° above the horizon (blue lines in figure 6c), but becomes shallow, without a distinct minimum, when selecting an elevation slice from 0° to 22° above the horizon that contains the landmark panorama (green lines in figure 6c). The directional information offered by this remote release site is thus dominated by celestial compass information and therefore explains why all ants initially followed their home vector when released there (figure 5d).
(f). Determining the home direction
However, ants would need to be able to choose the appropriately orientated goal snapshot to detect the correct home direction; in the case of the nest shown in figure 3, for instance, if released to the south of the nest, they would need to recognize the snapshot memory that they stored when facing in that direction across the nest. Otherwise, they would not be able to decide, for instance, whether to move south or north at the north and south release stations, respectively. One possibility for resolving this ambiguity would be for ants to store nest-directed snapshots at some distance and at different bearings from the nest during their learning walks [39,40]. At a release station to the south, they then would detect a more pronounced minimum of the rotIDF when comparing the current view with one learnt at a location south of the nest looking north, in contrast to a view learnt to the north of the nest looking south.
The rotIDF's between the views at 10 m north and south of the nest and nest-directed goal snapshots at distances of 0.5, 1 and 5 m from the nest indicate that appropriate nest directions can indeed be identified. The nest directions can be determined by comparing the depth of minima of all available snapshots, provided, however, the ants had memorized the views between 1 and 5 m distance from the nest (thick dark blue versus thin lines in figure 7; see the electronic supplementary material, figure S5 for second nest).
Figure 7.

Identifying home direction from rotIDFs. RotIDFs computed between nest-directed views 0.5, 1 and 5 m north, south, east and west of the nest (see schematic diagram in (c)) and the views at the release sites 10 m north (a) and south of the nest (b). Thick blue line indicates the true home direction from the north release site (a) and the south release site (b). Note: true home direction at 10 m release sites can be distinguished from all other possible directions, if the views experienced at these locations can be compared with nest-directed views that have been recorded at distances further than 1 m away from the nest.
4. Discussion
We have shown that individually foraging Australian jack jumper ants can quickly and accurately determine their position relative to the nest from all compass directions, when released within a radius of 10–15 m around the nest. Within this area, information from the ants’ path integration system is suppressed. When released beyond this region, the ants fall back on path integration information, but rarely follow the home vector for more than a few metres, a phenomenon described before for ants operating in landmark-rich environments [42,43].
Myrmecia croslandi foragers thus rely heavily on visual landmark information, and we have shown in principle that in their open woodland habitat, the information content is sufficient to explain the ants’ versatile homing abilities. Our experiments with full-vector ants created a conflict between the home directions indicated by the path integrator and by the visual landmark panorama, and it is interesting to note that we found a spatial gradient of how this conflict is resolved. Within what we may call the familiar landmark territory of the ants, path integration information is completely suppressed. Slightly outside this area, we found both instances, where displaced ants followed a compromise direction between those indicated by path integration and by landmark guidance, as has been observed in other studies [7,13,14], and instances where the ants’ path was at least initially exclusively guided by path integration information [2,42].
Our analysis of the navigational information provided by panoramic views in the habitat of M. croslandi has resulted in a clear and testable prediction on the range over which ants would need to acquire nest-directed views during their learning walks. It will therefore be important in the next step to analyse how the learning walks of naive M. croslandi foragers relate to the development of their foraging direction and range. More generally, our results demonstrate the importance of systematically reconstructing the navigational information available in each particular environment in which animals navigate, and warn against concluding that insects operate on map-like representations of their environment, based on the demonstration that they can find home from places they might not have been before [19].
Acknowledgements
We are grateful to many undergraduate students, visiting scholars and research assistants who helped us with fieldwork, in particular Ash Card, Caitlin McDermott, Chloé Raderschall, Eliza Middleton, Emma Birnbaum, Nataniel Yeo, Piyankarie Jayatilaka and Sam Reid. We thank in particular Tom Collett for his critical, constructive and insightful comments and support. We are grateful to Jan Hemmi and Wolfgang Stürzl for their advice and help with Matlab programming and camera calibration. Aerial photography was provided by ACTPLA (the Australian Capital Territory Planning Authority).
Funding statement
We acknowledge funding support from the Australian Research Council's (ARC) Centres of Excellence Scheme (CEO561903), an ARC Discovery Project grant (DP0986606), an ARC Discovery Early Career Award (DE120100019) and the Hermon Slade Foundation. We thank the Defence Science and Technology Organization, Australia, and in particular, Javaan Chahl both for the initial loan of the DGPS system and, with Matt Garratt and Mark Snowball, for designing, constructing and maintaining DGPS electronics.
References
- 1.Wehner R. 2008. The architecture of the desert ant's navigational toolkit (Hymenoptera: Formicidae). Myrmecol. News 12, 85–96 [Google Scholar]
- 2.Buehlmann C, Cheng K, Wehner R. 2011. Vector-based and landmark-guided navigation in desert ants inhabiting landmark-free and landmark-rich environments. J. Exp. Biol. 214, 2845–2853 10.1242/jeb.054601 (doi:10.1242/jeb.054601) [DOI] [PubMed] [Google Scholar]
- 3.Cheung A, Vickerstaff R. 2010. Finding the way with a noisy brain. PLoS Comput. Biol. 6, e1000992. 10.1371/journal.pcbi.1000992 (doi:10.1371/journal.pcbi.1000992) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wehner R, Srinivasan MV. 2003. Path integration in insects. In The neurobiology of spatial behaviour (ed. Jeffery K.), pp. 9–30 Oxford, UK: Oxford University Press [Google Scholar]
- 5.Beugnon G, Lachaud JP, Chagné P. 2005. Use of long-term stored vector information in the neotropical ant Gigantiops destructor. J. Insect Behav. 18, 415–432 10.1007/s10905-005-3700-8 (doi:10.1007/s10905-005-3700-8) [DOI] [Google Scholar]
- 6.Collett M. 2010. How desert ants use a visual landmark for guidance along a habitual route. Proc. Natl Acad. Sci. USA 107, 11 638–11 643 10.1073/pnas.1001401107 (doi:10.1073/pnas.1001401107) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Narendra A. 2007. Homing strategies of the Australian desert ant Melophorus bagoti. II. Interaction of the path integrator with visual cue information. J. Exp. Biol. 210, 1804–1812 10.1242/jeb.02769 (doi:10.1242/jeb.02769) [DOI] [PubMed] [Google Scholar]
- 8.Graham P, Cheng K. 2009. Ants use the panoramic skyline as a visual cue during navigation. Curr. Biol. 19, R935–R937 10.1016/j.cub.2009.08.015 (doi:10.1016/j.cub.2009.08.015) [DOI] [PubMed] [Google Scholar]
- 9.Cheung A, Hiby L, Narendra A. 2012. Ant navigation: fractional use of the home vector. PLoS ONE 7, e50451. 10.1371/journal.pone.0050451 (doi:10.1371/journal.pone.0050451) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Fukushi T, Wehner R. 2004. Navigation in wood ants Formica japonica: context dependent use of landmarks. J. Exp. Biol. 207, 3431–3439 10.1242/jeb.01159 (doi:10.1242/jeb.01159) [DOI] [PubMed] [Google Scholar]
- 11.Kohler M, Wehner R. 2005. Idiosyncratic route-based memories in desert ants, Melophorus bagoti: how do they interact with path-integration vectors? Neurobiol. Learn. Mem. 83, 1–12 10.1016/j.nlm.2004.05.011 (doi:10.1016/j.nlm.2004.05.011) [DOI] [PubMed] [Google Scholar]
- 12.Wehner R, Michel B, Antonsen P. 1996. Visual navigation in insects: coupling of egocentric and geocentric information. J. Exp. Biol. 199, 129–140 [DOI] [PubMed] [Google Scholar]
- 13.Collett M. 2012. How navigational guidance systems are combined in a desert ant. Curr. Biol. 22, 927–932 10.1016/j.cub.2012.03.049 (doi:10.1016/j.cub.2012.03.049) [DOI] [PubMed] [Google Scholar]
- 14.Reid SF, Narendra A, Hemmi JM, Zeil J. 2011. Polarised skylight and the landmark panorama provide night-active bull ants with compass information during route following. J. Exp. Biol. 214, 363–370 10.1242/jeb.049338 (doi:10.1242/jeb.049338) [DOI] [PubMed] [Google Scholar]
- 15.Baerends GP. 1941. Fortpflanzungsverhalten und Orientierung der Grabwespe Ammophilia campestris Jur. Tijdschrift voor Entomologie 84, 68–275 [Google Scholar]
- 16.Collett TS, Collett M. 2002. Memory use in insect visual navigation. Nat. Rev. Neurosci. 3, 542–552 10.1038/nrn872 (doi:10.1038/nrn872) [DOI] [PubMed] [Google Scholar]
- 17.Collett TS, Zeil J. 1998. Places and landmarks: an arthropod perspective. In Spatial representation in animals (ed. Healy S.), pp. 18–53 Oxford, UK: Oxford University Press [Google Scholar]
- 18.Wehner R, Menzel R. 1990. Do insects have cognitive maps? Annu. Rev. Neurosci. 13, 403–414 10.1146/annurev.ne.13.030190.002155 (doi:10.1146/annurev.ne.13.030190.002155) [DOI] [PubMed] [Google Scholar]
- 19.Menzel R, et al. 2005. Honey bees navigate according to a map-like spatial memory. Proc. Natl Acad. Sci. USA 102, 3040–3045 10.1073/pnas.0408550102 (doi:10.1073/pnas.0408550102) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wystrach A, Beugnon G, Cheng K. 2012. Ants might use different view-matching strategies on and off the route. J. Exp. Biol. 215, 44–55 10.1242/jeb.059584 (doi:10.1242/jeb.059584) [DOI] [PubMed] [Google Scholar]
- 21.Pahl M, Zhu H, Tautz J, Zhang SW. 2011. Large scale homing in honeybees. PLoS ONE 6, e19669. 10.1371/journal.pone.0019669 (doi:10.1371/journal.pone.0019669) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Becker L. 1958. Untersuchungen über das Heimfindevermögen der Bienen. Zeitschrift für Vergleichende Physiologie 41, 1–25 [Google Scholar]
- 23.Capaldi EA, et al. 2000. Ontogeny of orientation flight in the honeybee revealed by harmonic radar. Nature 403, 537–540 10.1038/35000564 (doi:10.1038/35000564) [DOI] [PubMed] [Google Scholar]
- 24.Mangan M, Webb B. 2012. Spontaneous formation of multiple routes in individual desert ants (Cataglyphis velox). Behav. Ecol. 23, 944–954 10.1093/beheco/ars051 (doi:10.1093/beheco/ars051) [DOI] [Google Scholar]
- 25.Greiner B, Narendra A, Reid SF, Dacke M, Ribi WA, Zeil J. 2007. Eye structure correlates with distinct foraging-bout timing in primitive ants. Curr. Biol. 17, R879–R880 10.1016/j.cub.2007.08.015 (doi:10.1016/j.cub.2007.08.015) [DOI] [PubMed] [Google Scholar]
- 26.Narendra A, Reid SF, Greiner B, Peters RA, Hemmi JM, Ribi WA, Zeil J. 2011. Caste-specific visual adaptations to distinct daily activity schedules in Australian Myrmecia ants. Proc. R. Soc. B 278, 1141–1149 10.1098/rspb.2010.1378 (doi:10.1098/rspb.2010.1378) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jayatilaka P, Narendra A, Reid SF, Cooper P, Zeil J. 2011. Different effects of temperature on foraging activity schedules in sympatric Myrmecia ants. J. Exp. Biol. 214, 2730–2738 10.1242/jeb.053710 (doi:10.1242/jeb.053710) [DOI] [PubMed] [Google Scholar]
- 28.Monteiro LS, Moore T, Hill C. 2005. What is the accuracy of DGPS? J. Navigation 58, 207–225 10.1017/S037346330500322X (doi:10.1017/S037346330500322X) [DOI] [Google Scholar]
- 29.Stürzl W, Zeil J. 2007. Depth, contrast and view-based homing in outdoor scenes. Biol. Cybern. 96, 519–531 10.1007/s00422-007-0147-3 (doi:10.1007/s00422-007-0147-3) [DOI] [PubMed] [Google Scholar]
- 30.Zeil J, Hofmann MI, Chahl JS. 2003. Catchment areas of panoramic snapshots in outdoor scenes. J. Opt. Soc. Am. A 20, 450–469 10.1364/JOSAA.20.000450 (doi:10.1364/JOSAA.20.000450) [DOI] [PubMed] [Google Scholar]
- 31.Schultheiss P, Cheng K. 2011. Finding the nest: inbound searching behaviour in the Australian desert ant, Melophorus bagoti. Anim. Behav. 81, 1031–1038 10.1016/j.anbehav.2011.02.008 (doi:10.1016/j.anbehav.2011.02.008) [DOI] [Google Scholar]
- 32.Wehner R, Srinivasan MV. 1981. Searching behaviour of desert ants, genus Cataglyphis (Formicidae, Hymenoptera). J. Comp. Physiol. 142, 315–338 10.1007/BF00605445 (doi:10.1007/BF00605445) [DOI] [Google Scholar]
- 33.Buehlmann C, Hansson BS, Knaden M. 2012. Path integration controls nest-plume following in desert ants. Curr. Biol. 22, 645–649 10.1016/j.cub.2012.02.029 (doi:10.1016/j.cub.2012.02.029) [DOI] [PubMed] [Google Scholar]
- 34.Steck K. 2012. Just follow your nose: homing by olfactory cues in ants. Curr. Opinion Neurobiol. 22, 231–235 10.1016/j.conb.2011.10.011 (doi:10.1016/j.conb.2011.10.011) [DOI] [PubMed] [Google Scholar]
- 35.Baddeley B, Graham P, Philippides A, Husbands P. 2011. Holistic visual encoding of ant-like routes: navigation without waypoints. Adapt. Behav. 19, 3–15 10.1177/1059712310395410 (doi:10.1177/1059712310395410) [DOI] [Google Scholar]
- 36.Cartwright BA, Collett TS. 1983. Landmark learning in bees: experiments and models. J. Comp. Physiol. 151, 521–543 10.1007/BF00605469 (doi:10.1007/BF00605469) [DOI] [Google Scholar]
- 37.Cartwright BA, Collett TS. 1987. Landmark maps for honeybees. Biol. Cybern. 57, 85–93 10.1007/BF00318718 (doi:10.1007/BF00318718) [DOI] [Google Scholar]
- 38.Zeil J, Boeddeker N, Stürzl W. 2009. Visual homing in insects and robots. In Flying insects and robots (eds Floreano D, Zufferey J-C, Srinivasan MV, Ellington C.), pp. 87–100 Berlin, Germany: Springer [Google Scholar]
- 39.Graham P, Philippides A, Baddeley B. 2010. Animal cognition: multi-modal interactions in ant learning. Curr. Biol. 20, R639–R640 10.1016/j.cub.2010.06.018 (doi:10.1016/j.cub.2010.06.018) [DOI] [PubMed] [Google Scholar]
- 40.Müller M, Wehner R. 2010. Path integration provides a scaffold for landmark learning in desert ants. Curr. Biol. 20, 1368–1371 10.1016/j.cub.2010.06.035 (doi:10.1016/j.cub.2010.06.035) [DOI] [PubMed] [Google Scholar]
- 41.Vardy A, Möller R. 2005. Biologically plausible visual homing methods based on optical flow techniques. Connect. Sci. 17, 47–89 10.1080/09540090500140958 (doi:10.1080/09540090500140958) [DOI] [Google Scholar]
- 42.Collett M, Collett TS, Wehner R. 1999. Calibration of vector navigation in desert ants. Curr. Biol. 9, 1031–1034 10.1016/S0960-9822(99)80451-5 (doi:10.1016/S0960-9822(99)80451-5) [DOI] [PubMed] [Google Scholar]
- 43.Narendra A. 2007. Homing strategies of the Australian desert ant Melophorus bagoti. I. Proportional path-integration takes the ant half-way home. J. Exp. Biol. 210, 1798–1803 10.1242/jeb.02768 (doi:10.1242/jeb.02768) [DOI] [PubMed] [Google Scholar]