

Abstract
OBJECTIVE
To describe and make available an interactive, 24-variable homeostasis model assessment (iHOMA2) that extends the HOMA2 model, enabling the modeling of physiology and treatment effects, to present equations of the HOMA2 and iHOMA2 models, and to exemplify iHOMA2 in two widely differing scenarios: changes in insulin sensitivity with thiazolidinediones and changes in renal threshold with sodium glucose transporter 2 (SGLT2) inhibition.
RESEARCH DESIGN AND METHODS
iHOMA2 enables a user of the available software to examine and modify the mathematical functions describing the organs and tissues involved in the glucose and hormonal compartments. We exemplify this with SGLT2 inhibition modeling (by changing the renal threshold parameters) using published data of renal effect, showing that the modeled effect is concordant with the effects on fasting glucose from independent data.
RESULTS
iHOMA2 modeling of thiazolidinediones effect suggested that changes in insulin sensitivity in the fasting state are predominantly hepatic. SGLT2 inhibition modeled by iHOMA2 resulted in a decrease in mean glucose of 1.1 mmol/L. Observed data showed a decrease in glucose of 0.9 mmol/L. There was no significant difference between the model and the independent data. Manipulation of iHOMA2's renal excretion threshold variable suggested that a decrease of 17% was required to obtain a 0.9 mmol/L decrease in mean glucose.
CONCLUSIONS
iHOMA2 is an extended mathematical model for the assessment of insulin resistance and β-cell function. The model can be used to evaluate therapeutic agents and predict effects on fasting glucose and insulin and on β-cell function and insulin sensitivity.
Type 2 diabetes is caused by a combination of progressive β-cell dysfunction, relative insulin deficiency, and variable degrees of insulin resistance that lead to dysregulation of glucose homeostasis. Understanding the biochemistry, phenotypic details, and genetic mechanisms contributing to this can yield important information on pathophysiology. The progressive nature of the disease, as well as measuring the rate of deterioration, has presented an ongoing challenge to clinicians and scientists alike. Tools to track β-cell functional changes and insulin resistance fall into three broad categories: measures of glycemic status (e.g., fasting glucose, HbA1c), physiological investigations (e.g., clamp techniques [1,2], glucose tolerance tests), and mathematical modeling (e.g., minimal model [3], Mari model [4,5], homeostasis model assessment [HOMA] [6–9]).
No single approach proved sufficient, either, for a comprehensive quantitative description of β-cell dysfunction or insulin resistance. Measures of these parameters vary depending on whether measurements are from basal or stimulated or fasting or postprandial subjects and whether pharmaceutical agents are being taken. Physiological techniques, ranging from simple glucose tolerance tests to euglycemic clamps and stable isotope studies, require expertise and are time and resource intensive, limiting their use to relatively small numbers of subjects (10).
Mathematical modeling techniques also vary in their physiological assumptions. Computer-based solutions from clinical interventions (e.g., oral glucose tolerance tests with “minimal model” readout) have limitations because of the high number of samples required from each subject. Simpler modeling methods (e.g., HOMA2) use paired fasting plasma insulin and glucose concentrations to derive data on β-cell function and insulin sensitivity. HOMA2 yields a single readout of β-cell function and insulin resistance for each subject and has the advantage that, since it only requires paired basal insulin and glucose measurements, it can be used in large epidemiological and pharmaceutical studies.
One disadvantage with HOMA2 is that it is not an appropriate model to use when evaluating treatments that have similar functional effects on blood glucose but different modes of action. For example, in HOMA2 β-cell function is characterized internal to the model as a sigmoidal dose response curve relating insulin secretion to the prevailing glucose concentration. The shape of this sigmoidal curve is modeled using two principal variables, one of which describes the rate of insulin secretion (Km) and one that describes the maximal insulin secretion (Vmax). In HOMA2, changes in β-cell function, altering insulin secretion, are entirely attributed to changes in Vmax and Km in the basal state (11). This modeling of β-cell function has been a good approximation, but a comprehensive description of the β-cell dynamics requires more variables (11). The β-cell functional changes caused by sulfonylureas (12) have different dose response changes and characteristics from those induced by incretin hormones (13). Similarly, total body insulin sensitivity, defined as net glucose clearance for any given insulin concentration, cannot be comprehensively described using one fixed function as it is in HOMA2. Hepatic insulin sensitivity may differ from peripheral sensitivity, and change in glucose clearance can be completely unrelated to either hepatic or peripheral sensitivity as in the case of sodium glucose transporter 2 (SGLT2) inhibition (14). The invariant nature of HOMA2 does not allow a sufficient description for other organs and tissues involved in glucose homeostasis, and this can be important where there is knowledge of changed function, e.g., in the liver, which could be used to improve model outputs.
We have developed an interactive, 24-variable model (termed iHOMA2) that addresses problems associated with the fixed assumptions within HOMA2. Here, we describe the model in cartoon, graphical, and mathematical detail; discuss how it can be used and manipulated; present an explicit example of its use to model changes in renal threshold induced by blockade of glucose reabsorption by SGLT2 inhibition; and show how this affects the modeled fasting glucose. The model is available on open access.
RESEARCH DESIGN AND METHODS
Model development
The iHOMA2 model is shown in Fig. 1 as graphical (A), box diagrammatic (B), and mathematical (C), respectively. iHOMA2 is an integrated computer-based mathematical model of glucose and hormonal interaction under homeostatic conditions. The model, now available online at http://www.ihoma.co.uk, runs in real time with 24 operator-controlled variables (Table 1) and graphical output displays. The baseline characteristics were built from those used in the original HOMA2 model, with all of the dose-response variables now explicit. iHOMA2 runs interactively and exactly for each calculation. iHOMA2 in its default start-up setting gives identical readings to HOMA2 and can be used as a direct substitute for HOMA2 in this mode. The operator can modify each of the variables using an interactive sliding control display. The operator can control every aspect of the dose-response curve. For example, the β-cell characteristics are described by P1–P5, each of these being independently adjustable. This allows “what if” scenarios to be explored: “What would be the effect on glucose if Vmax of β-cell function were 50%?” “How might that be modified if the dose response curve were shifted to the left?” “What if autonomous insulin secretion continued at low blood glucose?” Similarly, the functions relating to the other organs and tissues involved in the glucose and hormonal compartments can be modified using sliding control displays. In the HOMA2 model, insulin sensitivity is treated as a whole-body effect, altering the liver and periphery to the same extent. In iHOMA2, this has been uncoupled and the insulin sensitivity of these organs and tissues can be modified independently. The ability to alter the 24 variables of iHOMA2 enables the modeling of known or surmised pathology and physiology and the effect of treatments both alone and in combination. The effects of the treatments on fasting glucose, insulin, β-cell function (%B), and insulin sensitivity (%S) are graphically represented in the model.
Figure 1.
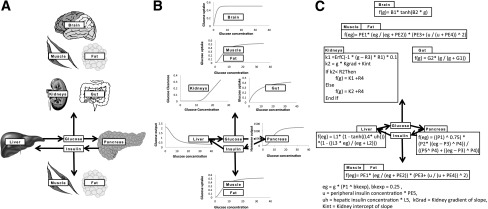
The iHOMA2 model is represented in three different ways: A) as a cartoon displaying the organs and tissues involved; B) showing the dose response curves as graphs modeled for each organ or tissue; and C) providing the equations that govern the organs and tissues functioning in the model. B1 and B2 are modifiable for the brain, PE1–PE5 for the periphery, R1–R4 for the kidneys, G1 and G2 for the gut, L1–L5 for the liver, and P1–P5 for the pancreas (see Table 1).
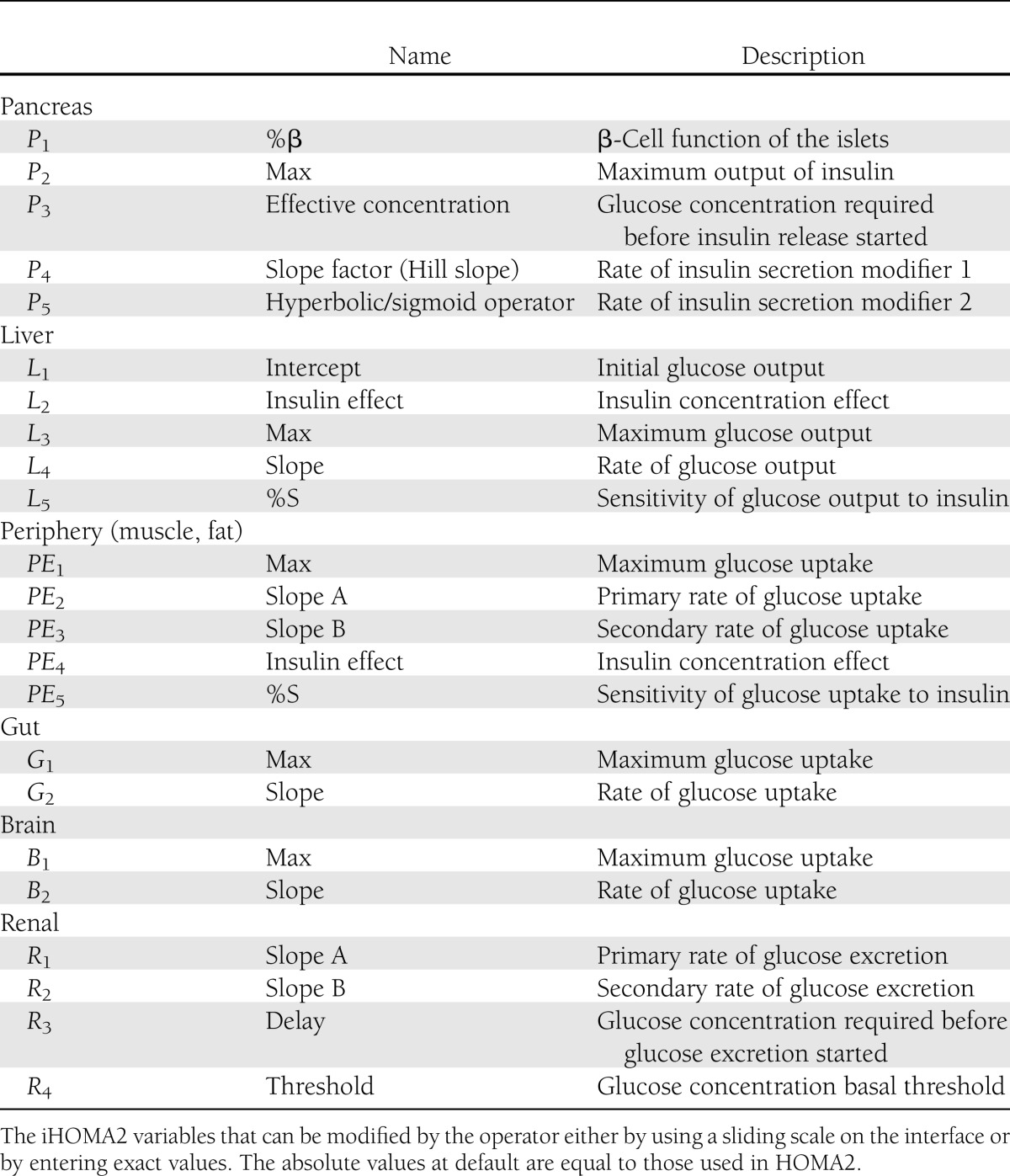
Table 1.
Modifiable variables within iHOMA2
The model allows for analytical and predictive modes of use. The analytical mode allows insulin resistance and β-cell function to be read from the input of insulin and glucose in the basal state, while the predictive function shows the estimated and modeled insulin and glucose concentrations in the basal state when the β-cell function and insulin resistance parameters are set.
This article shows two detailed quantified scenarios to illustrate the interactive modalities. The first example shows that the effect of pioglitazones (thiazolidinediones) on insulin resistance can be partitioned between the liver and periphery. The second example illustrates the model’s use elucidating the effect of an SGLT2 inhibitor on glycemia. All analyses were performed using SPSS, version 19.0 (SPSS, Chicago, IL). Statistical comparisons were made using Z tests for skewness, Student independent samples t test for comparison of means, and F tests for assessment of fit of the model to the observed data (15).
Quantitative model usage: effect of pioglitazone
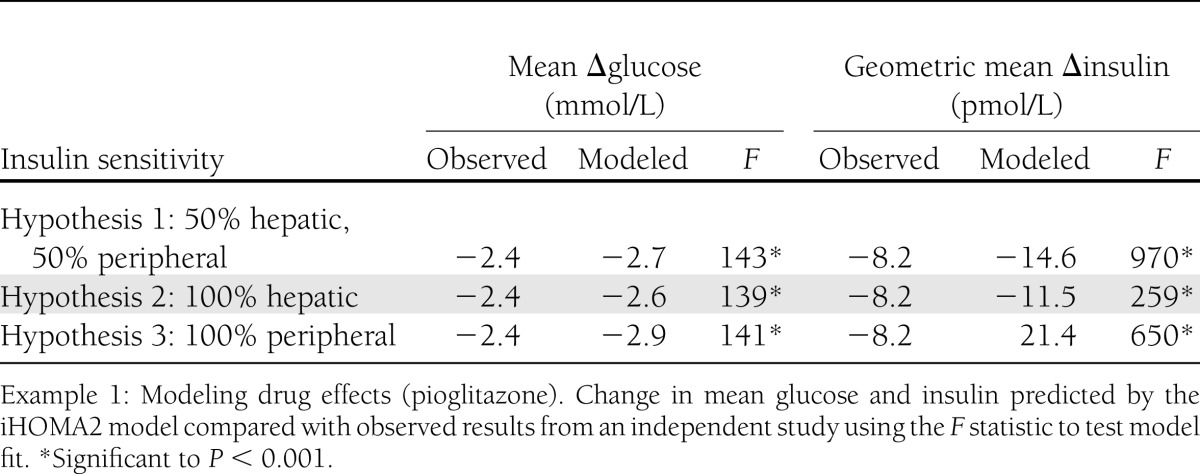
To model the effects of pioglitazone, we examined the outcome when insulin sensitivity was modeled to be in the liver, in the periphery, or at both sites equally—all with a standardized increase in β-cell function. Three possible sites of action on insulin sensitivity for pioglitazone were modeled: hypothesis 1, insulin sensitivity increases in both periphery and hepatic (variables L5 and PE5); hypothesis 2, hepatic insulin sensitivity increases and peripheral insulin sensitivity remains unchanged (variable L5); and hypothesis 3, hepatic insulin sensitivity remains unchanged and peripheral insulin sensitivity increases (variable PE5).
The development dataset comprised insulin and glucose values from a monotherapy study of pioglitazone (16). The changes to β-cell function and insulin sensitivity observed between the baseline visit and end of study (12 months later) in the development group were used as inputs to adjust the variables in the iHOMA2 model for each of the three hypotheses. A separate study of pioglitazone (17) was used for the verification group. The data from the baseline visits of the verification group were submitted to the adjusted iHOMA2, using the model in predictive mode, to determine the effect of pioglitazone for each of the three hypotheses using as output the expected fasting glucose and insulin after therapy. We assessed bias and agreement using a Bland-Altman plot and assessed the fit of the model by examination of the least squares deviation from the line of unity (where the observed values equal the predicted values) using an F statistic to test the model fit.
Quantitative model usage: the effect of glucose reabsorption inhibitors
SGLT2 partially prevent the reabsorption of glucose, thus changing the renal threshold for glycosuria. Patients using an SGLT2 inhibitor demonstrate a marked increase in glucose urinary loss. A recent publication estimated, for a 5-mg dose of dapagliflozin over 2 weeks, a 20% decrease in the renal threshold for glycosuria (18). To model the effect of SGLT2 inhibition, we used a phenotypically similar subject set (previously published [19]) where fasting glucose and insulin measurements were known. SGLT2 inhibition was modeled in iHOMA2 (variable R4 = 120%) based on the published data to predict the effect on the fasting glucose and insulin. The glucose changes predicted were compared with a separate preselected phase 3 trial of dapagliflozin (20) using an independent t test.
Further, the iHOMA2 model was used to examine any change required in renal glucose excretion to achieve an equivalent change in fasting glucose in our cohort. iHOMA2 was manipulated (by change of variable R4) until the change in glucose from the model was equal to the change in glucose from the dapagliflozin study. The value of the variable R4 was then read from the model.
RESULTS
Quantitative model usage: effect of pioglitazone
Data were analyzed for normal distribution before parametric analysis. The z test value for the skewness of the insulin data was 3.9. Log transformation eliminated the skewness as indicated by the z value of 0.5. Geometric mean insulin results are therefore presented.
In the development group, pioglitazone increased β-cell function from 36.9 to 49.2, a relative increase of 33.4%, and increased insulin sensitivity from 61.4 to 79.3, a relative increase of 29.1%. Since we now had a calculated generic change in insulin sensitivity with an associated β-cell functional change, we were able to partition the site of resistance to test three hypotheses (Fig. 2):
Figure 2.
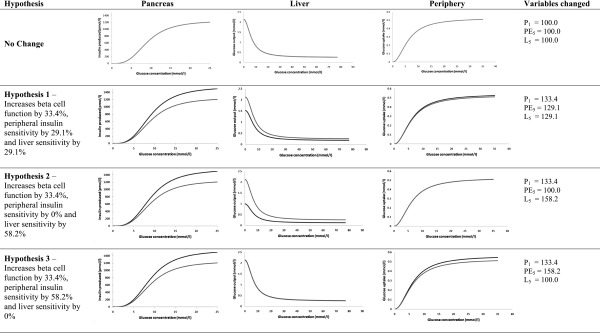
Three hypotheses of action of pioglitazone on β-cell function and insulin sensitivity analyzed by iHOMA2. Gray lines are the unaltered functions in iHOMA2. Black lines are the altered variable functions modeled. Hypothesis 1, increase of whole-body sensitivity and an increase in β-cell function; hypothesis 2, increase in liver insulin sensitivity only with an increase in β-cell function; and hypothesis 3, increase in peripheral insulin sensitivity only and an increase in β-cell function.
That insulin resistance change with pioglitazone is equally partitioned between the liver and the periphery. Whence we set hepatic insulin sensitivity increase by 29.1% and peripheral insulin sensitivity increase by 29.1% with β-cell function increased by 33.4% (variables L5 = 129.1%, PE5 = 129.1%, P1 = 133.4%).
That insulin resistance change with pioglitazone is only sited at the liver. Whence we set hepatic insulin sensitivity increase by 58.2%, peripheral insulin sensitivity unchanged, and β-cell function increase by 33.4% (variables L5 = 158.2%, PE5 = 100.0%, P1 = 133.4%).
That insulin resistance change with pioglitazone is only sited at the periphery. Whence we set peripheral insulin sensitivity increase by 58.2%, hepatic insulin sensitivity unchanged, and β-cell function increase by 33.4% (variables L5 = 100.0%, PE5 = 158.2%, P1 = 133.4%).
The predicted change in glucose and the change in insulin from these hypotheses were contrasted with the observed values from the verification group using Bland-Altman plots and F tests for model fit. The Bland-Altman plots did not show significant differentiation between the models. F test (Table 2) demonstrated that the second hypothesis—maximal hepatic insulin sensitivity—was found to be the model with the best fit to the observed data. The F statistics for linearity were 139 and 259 (with 668 df) (significance P < 0.001) for glucose and insulin, respectively.
Table 2.
Example of hypothesis predictions from the iHOMA2 model
Quantitative model usage: the effect of glucose reabsorption inhibitors
iHOMA2 predicted that a decrease by 20% in the renal excretion threshold using 5 mg dapagliflozin treatment over 2 weeks would result in a decrease in mean (SD) glucose of 1.1 (0.3) mmol/L in the study subjects. Data extracted from published literature on dapagliflozin (5-mg dose) reported a similar change in mean glucose of 0.9 mmol/L in their subjects after 2 weeks. There was no significant difference between the model and the observed data. There are no published data on changes in insulin, so these could not be compared.
Although the model and observed results were broadly concordant, further manipulation of the iHOMA2 models renal excretion threshold variable (R4) suggested that alteration of the renal threshold by 17% (R4 = 117%) gave the closest approximation for the observed mean decrease of 0.9 mmol/L in glucose.
CONCLUSIONS
iHOMA2 is an extension of HOMA2 that enables the interactive modeling of physiology and treatment effects. It is now possible to use more of the potential of the structural basis of the model through the 24 interactive variables. This circumvents a drawback of HOMA2, namely, that this model does not allow individual compartmental or biological variables to be specified even when these are known. iHOMA2 can be used in default mode (for comparison with all published data using HOMA and HOMA2); in analytic mode, which allows %B and %S to be calculated from fasting insulin and glucose; or in predictive mode that allows fasting insulin and glucose to be calculated from %B and %S.
The iHOMA2 model presents all those variables that have some identifiability, so, for example, a renal threshold or glomerular filtration rate (slope of the renal excretion curve) can be entered to give greater specificity to the model. A group that has validated data on the observed glomerular filtration rate could then validate a specific change to the model variable.
We have demonstrated iHOMA2 use in predicting the drug effect of existing compounds with a known method of action, as in the example of pioglitazone. It was shown that of the three hypotheses of the site of insulin sensitivity change in the basal state, the hypothesis that the effect was mainly hepatic was the most robust. This is in line with the published literature (21,22) and accords with clamp data (23) that suggest that a substantial effect is mediated by hepatic gluconeogenesis, although the model, with multiple variables that can be manipulated, cannot be regarded as yielding a definitive answer to the question. As with all models, the conclusion should be taken to be illustrative rather than definitive, which is further exemplified by the inability of the Bland-Altman statistics to discriminate between the hypotheses.
We have exemplified iHOMA2 used in modeling with an SGLT2 inhibitor, where the effect on blood glucose is independent of β-cell function and insulin sensitivity. The model suggested that the renal threshold change produced by dapagliflozin in our study set was best approximated by 17% and not 20% as reported by others. The iHOMA2 model can be used as a method of assessing quantitative identifiable parameters—in this case, renal threshold—from observed data. This has the potential, for example, to quantify SGLT2 dose effects or comparisons between different SGLT2 inhibitors.
The utility of a model depends on its identifiability. A model is structurally identifiable if its parameters can be uniquely estimated by fitting the model to observed data. This is related to the sensitivity of the process outputs to the parameter variations and understanding how the parameter relationships might interact (24–26). The variables in this model all have identifiable functions that are visualized in each panel as changes to the interactive kinetics. This is quite unlike curve-fitting exercises, where the mathematics may mimic the response only because multiple, but physiologically unidentified, terms have been used. For example, a complex reaction curve may be described by a polynomial, but there would be no clear physiological ”sense” in knowing the value of a squared or cubed constant. Model descriptions only give insights where the variables represent some aspect of the known physiology. With a progressive understanding of human physiology, it will be possible to be more specific about the model characteristics; e.g., one can set β-cell functional changes to be Km or Vmax related and insulin resistance to be tissue specific. Additional modules—such as the incretin axis—could be activated and manipulated. The resultant model could then be used to give predictions for combinations of agents, activating specific mathematical subsections describing human pharmacology.
Mathematical models, such as the Mari model (4) or Bergman minimal model (3), are useful for the study of complex phenomena, as they demonstrate how measurements of glucose and insulin can be seen as manifestations of a complex feedback process. iHOMA2 has the functionality to display the individual components of the glucose and insulin interaction and resolve the steady-state mathematics to yield measures of β-cell function and insulin resistance. Using the model interactively, one can predict (and display graphically) many pathological and physiological scenarios. iHOMA2 can also be individualized, by adjustment of the variables, to specific phenotypes, genotypes, populations, or individuals.
The iHOMA2 model does have the disadvantage in that, in common with all mathematical models, it has limitations to its use and is not intended to provide a “complete” metabolic description. Allowing many variables to be internally manipulated means that identifiability of a solution can be at risk (its identifiability is reduced). So, iHOMA2 records a list of the 24 variables currently able to be changed and their settings. For transparency in reporting, we would advise that the values of the settings on the model are explicitly stated when they differ from the default. iHOMA2 is still a homeostatic model; i.e., it calculates a steady-state solution. The HOMA2 model was expanded to include glucose infusion kinetics (27). Further work is underway in iHOMA2 to add dynamic aspects so that glucose tolerance tests or clamp data could be modeled. A drawback of iHOMA2 is that it loses the simple assessment aspect of HOMA2—where there are two outputs from two inputs. iHOMA2 is, instead, a more user-definable model allowing the individual characteristics of organs and tissues to be manipulated. This manipulation allows for hypothesis testing of treatment effects and the modeling of pharmaceutical treatments for clinical care.
In conclusion, the creation of the iHOMA2 model addresses some of the limitations with our previous invariant model. iHOMA2 is an application for extending the range of the existing HOMA2 model and can be used to evaluate therapeutic agents in greater physiological detail and can predict their effects on fasting glucose and insulin and on β-cell function and insulin sensitivity. iHOMA2 can be used for hypothesis testing, for the evaluation of pharmaceutical agents, for the estimation of effect sizes of therapies, and for evaluation of indicative changes to glucose when a combination of agents is used. It can be used in its default mode to generate HOMA2 results directly comparable with those in the literature. The iHOMA2 program is freely available (http://www.ihoma.co.uk) for academic and noncommercial use.
Acknowledgments
The authors thank the Oxford National Institute for Health Research (NIHR) Biomedical Research Centre for the support of N.R.H. D.R.M. acknowledges support from the NIHR, of which he is a Senior Investigator.
The authors thank Takeda PLC and Novo Nordisk for unrestricted educational grants while this work was undertaken. No other potential conflicts of interest relevant to this article were reported.
N.R.H. researched data, wrote the manuscript, and coded the iHOMA2 program. J.C.L. and D.R.M. provided intellectual input into the modeling, wrote parts of the paper and discussion, and reviewed and edited the manuscript. D.R.M. is the guarantor of this work and, as such, had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
The authors acknowledge the help of Dr. R. Newman, University of Oxford, Oxford, U.K., for his assistance is creating the online iHOMA2 software. The authors also acknowledge the work of all the contributors to the previous versions of HOMA (6–8) including but not limited to the following: J. Hosker, J. Levy, D. Matthews, B. Naylor, S. O'Rahilly, A. Rudenski, D. Treacher, and R. Turner, with a special mention of R. Morris, who extended the model considerably to include the proinsulin and renal modules. All were members of the University of Oxford during the development of HOMA.
References
- 1.DeFronzo RA, Tobin JD, Andres R. Glucose clamp technique: a method for quantifying insulin secretion and resistance. Am J Physiol 1979;237:E214–E223 [DOI] [PubMed] [Google Scholar]
- 2.Harano Y, Ohgaku S, Hidaka H, et al. Glucose, insulin and somatostatin infusion for the determination of insulin sensitivity. J Clin Endocrinol Metab 1977;45:1124–1127 [DOI] [PubMed] [Google Scholar]
- 3.Bergman RN, Ider YZ, Bowden CR, Cobelli C. Quantitative estimation of insulin sensitivity. Am J Physiol 1979;236:E667–E677 [DOI] [PubMed] [Google Scholar]
- 4.Mari A, Pacini G, Murphy E, Ludvik B, Nolan JJ. A model-based method for assessing insulin sensitivity from the oral glucose tolerance test. Diabetes Care 2001;24:539–548 [DOI] [PubMed] [Google Scholar]
- 5.Mari A. Mathematical modeling in glucose metabolism and insulin secretion. Curr Opin Clin Nutr Metab Care 2002;5:495–501 [DOI] [PubMed] [Google Scholar]
- 6.Matthews DR, Hosker JP, Rudenski AS, Naylor BA, Treacher DF, Turner RC. Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985;28:412–419 [DOI] [PubMed] [Google Scholar]
- 7.Turner RC, Rudenski AS, Matthews DR, Levy JC, O’Rahilly SP, Hosker JP. Application of structural model of glucose-insulin relations to assess beta-cell function and insulin sensitivity. Horm Metab Res Suppl 1990;24:66–71 [PubMed] [Google Scholar]
- 8.Levy JC, Matthews DR, Hermans MP. Correct homeostasis model assessment (HOMA) evaluation uses the computer program. Diabetes Care 1998;21:2191–2192 [DOI] [PubMed] [Google Scholar]
- 9.Wallace TM, Levy JC, Matthews DR. Use and abuse of HOMA modeling. Diabetes Care 2004;27:1487–1495 [DOI] [PubMed] [Google Scholar]
- 10.McAuley KA, Mann JI, Chase JG, Lotz TF, Shaw GM. Point: HOMA—satisfactory for the time being: HOMA: the best bet for the simple determination of insulin sensitivity, until something better comes along. Diabetes Care 2007;30:2411–2413 [DOI] [PubMed] [Google Scholar]
- 11.Hockaday D, Sayyad M, Yajnik C. Counterpoint: appreciating homeostasis model assessment: more useful earlier rather than later. Diabetes Care 2007;30:2414–2418 [DOI] [PubMed] [Google Scholar]
- 12.Hosker JP, Burnett MA, Davies EG, Harris EA, Turner RC. Sulphonylurea therapy doubles B-cell response to glucose in type 2 diabetic patients. Diabetologia 1985;28:809–814 [DOI] [PubMed] [Google Scholar]
- 13.Degn KB, Juhl CB, Sturis J, et al. One week’s treatment with the long-acting glucagon-like peptide 1 derivative liraglutide (NN2211) markedly improves 24-h glycemia and alpha- and beta-cell function and reduces endogenous glucose release in patients with type 2 diabetes. Diabetes 2004;53:1187–1194 [DOI] [PubMed] [Google Scholar]
- 14.Ehrenkranz JR, Lewis NG, Kahn CR, Roth J. Phlorizin: a review. Diabetes Metab Res Rev 2005;21:31–38 [DOI] [PubMed] [Google Scholar]
- 15.Altman DG. Pratical Statistics for Medical Research. London, Chapman & Hall, 1991 [Google Scholar]
- 16.Charbonnel BH, Matthews DR, Schernthaner G, Hanefeld M, Brunetti P, QUARTET Study Group A long-term comparison of pioglitazone and gliclazide in patients with Type 2 diabetes mellitus: a randomized, double-blind, parallel-group comparison trial. Diabet Med 2005;22:399–405 [DOI] [PubMed] [Google Scholar]
- 17.Schernthaner G, Matthews DR, Charbonnel B, Hanefeld M, Brunetti P, Quartet [corrected] Study Group Efficacy and safety of pioglitazone versus metformin in patients with type 2 diabetes mellitus: a double-blind, randomized trial. J Clin Endocrinol Metab 2004;89:6068–6076 [DOI] [PubMed] [Google Scholar]
- 18.Komoroski B, Vachharajani N, Feng Y, Li L, Kornhauser D, Pfister M. Dapagliflozin, a novel, selective SGLT2 inhibitor, improved glycemic control over 2 weeks in patients with type 2 diabetes mellitus. Clin Pharmacol Ther 2009;85:513–519 [DOI] [PubMed] [Google Scholar]
- 19.Cook JT, Page RC, O’Rahilly S, et al. Availability of type II diabetic families for detection of diabetes susceptibility genes. Diabetes 1993;42:1536–1543 [DOI] [PubMed] [Google Scholar]
- 20.Ferrannini E, Ramos SJ, Salsali A, Tang W, List JF. Dapagliflozin monotherapy in type 2 diabetic patients with inadequate glycemic control by diet and exercise: a randomized, double-blind, placebo-controlled, phase 3 trial. Diabetes Care 2010;33:2217–2224 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sugiyama Y, Shimura Y, Ikeda H. Effects of pioglitazone on hepatic and peripheral insulin resistance in Wistar fatty rats. Arzneimittelforschung 1990;40:436–440 [PubMed] [Google Scholar]
- 22.Smith U. Pioglitazone: mechanism of action. Int J Clin Pract Suppl 2001:13–18 [PubMed] [Google Scholar]
- 23.Gastaldelli A, Miyazaki Y, Mahankali A, et al. The effect of pioglitazone on the liver: role of adiponectin. Diabetes Care 2006;29:2275–2281 [DOI] [PubMed] [Google Scholar]
- 24.Cobelli C, Pacini G, Toffolo G, et al. Estimation of insulin sensitivity and glucose clearance from minimal model: new insights from labeled IVGTT. Am J Physiol 1986;250:E591–E598 [DOI] [PubMed]
- 25.Kahn SE, Prigeon RL, McCulloch DK, et al. Quantification of the relationship between insulin sensitivity and beta-cell function in human subjects. Evidence for a hyperbolic function. Diabetes 1993;42:1663–1672 [DOI] [PubMed] [Google Scholar]
- 26.Jaqaman K, Danuser G. Linking data to models: data regression. Nat Rev Mol Cell Biol 2006;7:813–819 [DOI] [PubMed] [Google Scholar]
- 27.Hosker JP, Matthews DR, Rudenski AS, et al. Continuous infusion of glucose with model assessment: measurement of insulin resistance and beta-cell function in man. Diabetologia 1985;28:401–411 [DOI] [PubMed] [Google Scholar]