

Life on earth has evolved within a complex mixture of organic and inorganic compounds. While organic molecules such as amino acids, carbohydrates and nucleotides form the backbone of proteins and genetic material, these fundamental components of macromolecules are enzymatically synthesized and ultimately degraded. Inorganic elements such as copper (Cu), iron (Fe) and zinc (Zn), once solubilized from the earth’s crust, are neither created nor destroyed and therefore their homeostatic regulation is under strict control. In the fascinating field of “metals in biology”, by virtue of direct interactions with amino acid side-chains within polypeptide chains, metals play unique and critical roles in biology, promoting structures and chemistries that would not otherwise be available to proteins alone.
After the rise of photosynthetic organisms such as the cyanobacteria, oxygen accumulated in the atmosphere and oxygenated the oceans. This led to a decrease in the solubility of Fe and an expansion of the biological role of Cu, suggesting there was a shift from exclusively Fe in biology to embrace similar, though not identical roles for Cu. One property of Cu that drives its diverse roles in structure and catalysis, is its existence in either a reduced, Cu+, or oxidized, Cu2+, state. Since Cu+ has an affinity for thiol and thioether groups (examples found in cysteine or methionine), and Cu2+ exhibits preferred coordination to oxygen or imidazole nitrogen groups (found in aspartic and glutamic acid, or histidine, respectively), these metal ions can participate in a wide spectrum of interactions with proteins to drive diverse structures and biochemical reactions (Table 1). Moreover, in the process of moving between Cu+ and Cu2+, free intracellular Cu can generate hydroxyl radical, which can damage proteins, nucleic acids, and lipids, and can interfere with the synthesis of Fe-sulfur clusters that are essential for the activity of a number of important cellular enzymes. Additionally, the Irving-Williams series predicts that Cu can displace other metals such as Zn from their cognate ligands in metalloproteins, resulting in inappropriate protein structures or inhibition of enzymatic activity. Here we provide a brief overview of some roles for Cu in biology in bacterial, fungal, plant and mammalian systems, and we outline key ways that Nature balances the acquisition, distribution and regulation of Cu. It will become clear that as life evolved, more complex roles for Cu arose, concurrent with the elaboration of mechanisms to tightly regulate acquisition, distribution and the protection against Cu toxicity.
Table 1.
Examples of Cu Dependent Proteins and Cu Homeostasis Proteins
| Protein | Function | Bacteria | Fungi | Animals | Plants |
|---|---|---|---|---|---|
| Transcriptional Regulators | |||||
| Ace1 | Transcriptional activation in high Cu conditions | X | |||
| CopY | Bacterial Cu metalloregulatory repressor | X | |||
| CsoR | Bacterial Cu metalloregulatory repressor | X | |||
| Mac1 | Transcriptional activator in low Cu conditions | X | |||
| CueR | Bacterial Cu metalloregulatory repressor | X | |||
| Mtf1 | Metal-regulatory transcription factor | X | |||
| Spl7 | Transcriptional activator responding to Cu deficiency | X | |||
| Chaperones/Storage | |||||
| Atox1 | Metallochaperonedlivering Cu to P-type ATPases | X | X | X | |
| Ccs | Delivers Cu to the Cu/Zn SOD1 | X | X | X | |
| CopZ | Bacterial Cu chaperone | X | |||
| Metallothionein | Low molecular weight, cysteine rich metal binding and detoxification | X | X | X | X |
| Cell Surface/Secretory Compartment Transporters and Receptors | |||||
| P1B-Type ATPases | Cu+ exporting proteins | X | X | X | X |
| Ctr | Cu+ importing proteins | X | X | X | |
| Ethylene receptor | Uses Cu as a cofactor for ethylene signalling | X | |||
| Oxidoreductases | |||||
| Ascorbate oxidase | Reduction of L-ascorbate | X | |||
| Dopamine-monooxygenase | Tyrosine metabolism | X | |||
| Galactose oxidase | Reduction of Galactose | X | |||
| Amine Oxidase | Oxidation of diamines | X | X | X | X |
| Electron transfer/Energy production/Blue Cu protiens | |||||
| Cytochrome c oxidase | Necessary for the last step of respiration | X | X | X | X |
| Plastocyanin | Electron transfer during photosynthesis | X | X | ||
| NADH Dehydrogenase | Electron transfer from NADH to coenzyme Q | X | X | X | X |
| Nitrite Reductase | Reduces Nitrite to Nitric Oxide | X | |||
| Amicyanin | Electron accepting intermediate in the conversion of methylamine to formaldehyde and ammonia | X | |||
| Free Radical Scavenging | |||||
| Cu/Zn SOD | Free radical scavenging | X | X | X | X |
| Oxidase | |||||
| Laccase | Melanine production | X | X | X | X |
| Lysyl oxidase | Catalyzes the formation of collagen and elastin precursors, extracellular | X | |||
| Ceruloplasmin | MultiCu oxidase | X | |||
| Hephaestin | Transmembrane ferroxidase, transports iron from the intestine to the circulatory system | X | |||
| Multicopperferroxidase | Cu dependent iron uptake | X | X | X | |
| Monooxygenase | |||||
| Methane monooxygenase | Oxidizes C-H bond in methane | X | |||
| Phenylalanine hydrolase | Hydroxylation of the aromatic side chain of phenylalanine to generate tyrosine | X | |||
| Tyrosinase | Monophenolmonooxygenase, catalyses the oxidation of phenols, melanin synthesis | X | X | X | X |
Copper in prokaryotes
In general, prokaryotic organisms have adopted a limited role for Cu in their biochemistry and physiology. Almost all anaerobic bacteria and all anaerobic Archaea are limited Cu users, likely reflective of the limited availability of the metal in these conditions, and may be representative of the majority of life in primordial, anaerobic Earth. Yet, while Cu is not widely used by these prokaryotes, many of their genomes encode Cu exporters and Cu delivery proteins, known as chaperones, to protect against the toxic effects of Cu. A majority of prokayotes that use Cu express cytochrome c oxidase (Cox), the last enzyme in the respiratory electron transport chain. Typical Cu containing enzymes such as Cox, NADH dehydrogenase-2, and tyrosinases reside in the cytoplasmic membrane or periplasm where they are loaded with Cu, thereby reducing risk to cytoplasmic components. It is currently unclear how Cu is imported into bacterial cells. Current hypotheses include the presence of a broad-spectrum cation transporter and, in the case of the heavily Cu-dependent methanotrophs which use Cu as a catalytic co-factor in methane mono-oxygenase, the use of the Cu-binding peptide methanobactin.
A general model for Cu homeostasis in bacterial cells can be considered with respect to three basic functions that are focused on protection from Cu more than Cu utilization: sensing, intracellular mobilization, and export (Figure 1A). Many bacteria are primed to sense and respond to elevated Cu levels by one of three families of metalloregulatory repressors (CopY, CsoR, and CueR) that repress transcription of genes encoding the Cu export machinery under low Cu conditions. Cu is exported from bacterial cells by polytopic integral membrane Cu+ transporters, P1B-type ATPases, characterized by Cys-X2-Cys motifs or histidine-rich domains in their amino-termini and conserved His-Pro and Cys-Pro-Cys/His motifs within specific transmembrane domains. Through such ATPases, Cu is exported out of the cytoplasm into the periplasm or out of the cell. The periplasm of some Gram-negative species contains additional Cu detoxification and export mechanisms, such as Cu+ oxidases, Cu chaperones, and non-specific outer membrane metal cation transporters. Once Cu is exported from the cytosol, Cu-responsive transcriptional repressors rheostatically dampen transcription of the genes encoding the Cu transporting ATPases. Archaea appear to only encode Cu exporters, with no clear homology to known Cu importers or the chaperones identified in bacterial cells or in eukaryotes.
Figure 1. Copper homeostasis in Bacteria.

The themes of Cu homeostasis are conserved among Kingdoms. Import (orange), chaperones (blue), export (yellow), transcriptional control (grey). The general bacterial Cu response activates an operon consisting of a transcriptional repressor, a chaperone and a P-type exporter. In the absence of Cu, transcriptional repressors such as CueR, CsoR and CopY maintain repression of the Cu response machinery. In the presence of Cu, repressor proteins bind Cu and release from their promoter DNA bindings sites, resulting in derepression of genes encoding chaperones and P-type ATPases to facilitate protection against Cu toxicity by sequestration and export. Cytochrome c oxidase, located in the bacterial membrane, is the most common cuproenzyme in bacteria. The mechanism of Cu import in bacterial cells is currently poorly defined but may include a relatively non-metal-specific cation transporter. Alternatively, methanotrophic bacteria may use a non-translationally synthesized Cu binding peptide, methanobactin, to acquire Cu.
Copper in eukaryotes
With the evolution of single-cell eukaryotes came the new challenge of deliverying Cu to an expanding array of metalloproteins located within organelles such as mitochondria, chloroplasts, and the secretory compartments. Furthermore, with the subsequent appearance of multicellular organisms came the requirement to regulate Cu allocations to specific tissues according to varying metabolic needs. Thus, Cu homeostasis became a more complex and tightly controlled process at both an intra- and intercellular level.
Fungi
As the model single cell eukaryote, much of the basic nuts and bolts that mediate Cu homeostasis were first described in Saccharomyces cerevisiae, and the enhanced complexity of Cu balance in yeast, as compared to that found in prokaryotes, can be appreciated (Figure 1 B). In S. cerevisiae Cu is taken up by two apparently functionally redundant high affinity Cu transporters, Ctr1 and Ctr3, or a low affinity transporter, Fet4, after Cu2+ is reduced at the plasma membrane to Cu+ by cell surface metalloreductases such as Fre1. The Ctr family of importers have characteristic methionine rich amino termini that are thought to bind extracellular Cu+, potentially enhancing the local concentration of the metal. Additionally, there is an MxxxM motif within the second of its three transmembrane domains (where × is any amino acid), with the methionines likely functioning in Cu+ binding during translocation across the membrane, perhaps to carefully control the Cu+ ion as it traverses the lipid bilayer to prevent damage and to provide vectorial movement. Mac1 is a nuclear Cu-sensing transcriptional regulator that is responsible for activating the CTR1, CTR3 and FRE1 genes under low Cu conditions. Endocytosis and degradation of the Ctr1 protein is promoted under high Cu conditions. If external sources of Cu are vanishingly low, S. cerevisiae responds by mobilizing Cu stores from the vacuole, a process carried out by Ctr2, and a vacuolar-localized metalloreductase, Fre6. Branching out to other yeast, Schizosaccharomyces pombe encodes a major facilitator superfamily-type transporter, called Mfc1, that is responsible for Cu acquisition during meiosis, opening up the possibility for cell cycle specific and cell-type specific Cu transporters and Cu-dependent processes in other organisms.
Given that eukaryotic cells have elaborate compartments and organelles in which Cu-dependent proteins reside, or traverse on their journey to be secreted, tightly controlled intracellular Cu delivery mechanisms have evolved. Genetic studies in yeast identified two such small Cu-chaperone proteins, Atx1 and Ccs, which directly bind Cu near the surface on their three dimensional structure so that it is protected, yet exchangeable to other biological ligands. Atx1 interacts directly with the Ccc2 protein, a P-type Cu-transporting ATPase similar to that found in bacterial cells, that pumps Cu into the lumen of the secretory compartment where it is incorporated onto ligands of secreted Cu metalloproteins. Atx1 is structurally similar to the cytoplasmic Cu binding domains of Ccc2, providing similar protein surfaces to mediate Cu transfer between the two proteins. Cu, Zn superoxide dismutase (Sod1), which disproportionates the potentially toxic superoxide anion, is localized both to the cytoplasm and mitochondrial intermembrane space. The Cu chaperone for Sod1, Ccs, has a domain that structurally resembles Sod1, directly interacts with Sod1 in the cytoplasm and intermembrane space, delivering Cu to Sod1 in a complex series of reactions. While mitochondrial cytochrome oxidase has a critical requirement for Cu, the mitochondrial matrix harbors a significant pool of Cu that has been calculated to be in far excess of cytochrome oxidase needs, suggesting that mitochondria may harbor a storage pool of Cu. However, the molecular mechanisms by which Cu is imported into and mobilized from mitochondria are not yet clear. Several mitochondrial-associated proteins including Cox17, Sco1 and Sco2, conserved from yeast to humans, are involved in the delivery of Cu to cytochrome oxidase and mutations in human Sco genes are associated with severe infantile cardiomyopathy, underscoring the importance of deciphering how cytosolic Cu is routed to and from mitochondria.
While Cu uptake and distribution mechanisms in yeast are able to cope with a broad range of Cu availability, as cells encounter increases in environmental Cu, the expression of genes encoding the Cu detoxification proteins called metallothioneins (MTs) is dialed up. The S. cerevisiae Cup1 and Crs5 metallothioneins scavenge excess Cu by tight coordination to cysteine thiolates, buried from solvent, so that it is poorly exchangeable and therefore not highly reactive. MTs from different species rarely share sequence similarity, and are usually identified based solely on their high number of cysteines. In fact, while MTs were originally thought to be restricted to eukaryotes, their presence in Mycobacterium tuberculosis (Mtb) and Synechococcus species suggests they are also widely distributed in prokaryotes. The CUP1, CRS5 and SOD1 genes are transcriptionally activated by a second Cu metalloregulatory transcription factor, Ace1, under conditions of high Cu. Therefore, baker’s yeast activate distinct genes in response to low and high Cu using distinct Cu sensing transcription factors.
Mammals
Much of the cellular Cu acquisition and trafficking machinery in mammals is similar to that found in yeast. As yeast cells use Ctr family members to transport extracellular Cu+ across the plasma membrane, several studies indicate that Ctr1 is the primary transporter responsible for dietary Cu uptake into the intestinal epithelium. Unlike the transcriptional activation of yeast Ctr1 and Ctr3 in response to Cu deficiency, mammalian Ctr1 is regulated at the level of elevated Cu-induced endocytosis and degradation as a means to rheostatically regulate the steady state levels of Ctr1 and its abundance at the plasma membrane. Mammals also express the Atox1 (Atx1) and Ccs Cu chaperones that are functionally analogous to those in yeast. Copper specific transcriptional control has not been characterized in mammalian cells, although Cu activates MT gene transcription, in an apparently indirect manner, via the MTF-1 transcription factor.
Metazoans need not only acquire Cu and mediate its intracellular distribution, they must also parse Cu out to peripheral tissues where it drives cellular processes such as the high demand for mitochondrial oxidative phosphorylation in brain and heart tissue. In mammals Cu transported across the apical membrane by Ctr1 in intestinal enterocytes is shuttled to the ATP7A Cu-transporting P-type ATPase at the basolateral membrane by the Atox1 Cu chaperone and pumped into the portal circulation, where it makes its way to the liver, the major Cu storage organ. ATP7A plays crucial roles in moving Cu across other polarized cell layers including the placenta and the blood-brain barrier to ensure adequate copper in the developing fetus, and to meet the high demands for brain development and function. Similar to Ccc2 in yeast, ATP7A is also important for the delivery of copper to nascent proteins in the Golgi apparatus. In mammals, ATP7A is expressed in many tissues except the liver, where its expression is high in neonatal mice and diminishes during maturation. In the liver and other tissues, a homologous Cu-transporting ATPase, ATP7B, is important for loading Cu on ceruloplasmin, a secreted Cu containing ferroxidase in the plasma that has a key function in peripheral iron distribution. ATP7B also mobilizes excess Cu into the bile, preventing tissue Cu overload. Recent work suggests that mice lacking Ctr1 specifically in the cardiomyocytes of the heart develop a myopathy secondary to Cu deficiency, and in the process, release an unknown signal into the bloodstream that stimulates the activation of hepatic and intestinal ATP7A expression and the mobilization of Cu into the bloodstream. How mammals sense tissue Cu deficiencies, and elaborate signals that communicate with Cu acquisition and storage organs, will be an important area for further investigation.
Plants
The Cu homeostasis machinery in plants has undergone a dramatic expansion in complexity over the other forms of life explored to date, perhaps due to the presence of both mitochondria and chloroplasts, their relatively sessile lifestyle, and a collection of Cu-dependent proteins that are unique to plants (Figure 1D). For example, Arabidopsis thaliana expresses six Ctr-family transporters, COPT1-6, each with a distinct tissue-specific expression and subcellular localization patterns. COPT1, the most extensively studied transporter, is thought to be largely responsible for root Cu acquisition, but COPT2 also localizes to the plasma membrane in root tissues. Similar to Ctr2 in mammals, COPT4 does not appear to play a prominent role in Cu import. COPT5 may play a role in mobilization of vacuolar Cu stores, and COPT3 and COPT6 have less defined functions.
The P-type ATPases, known as heavy metal P-type ATPases (HMA) in plants, are composed of at least eight identified members in Arabidopsis. HMA1 to HMA4 are divalent cation transporters implicated in the export of not only Cu2+, but also Zn2+ and Cd2+. In contrast, HMA5 to HMA8 are thought to serve as monovalent Cu+ ion transporters. HMA7 (RAN1) is, to a large extent, responsible for the biogenesis of ethylene receptors, ETR, (ethylene is a key hormone for plant signalling) by delivering Cu to ETR1 en route through the endoplasmic reticulum, and it is not unreasonable that other secreted cuproproteins might also receive Cu in this manner. HMA5, primarily expressed in roots, is likely responsible for root Cu detoxification. HMA6 (PAA1) and HMA8 (PAA2) are targeted to the chloroplast and needed to deliver Cu to the thylakoidal protein plastocyanin, while only HMA6 is responsible for delivery of Cu to stromal SOD. Plastocyanin is required for photosynthesis and like cytochrome c oxidase is involved in electron flow.
The Arabidopsis CCH protein harbors an ATX1-like metallochaperone domain in its amino terminus, but has a plant-specific coiled-coil carboxyl terminal domain. This domain forms amyloid-like fibrils and may play a role in CCH-mediated Cu transport through the plasmodesmata, representing a novel method of inter-cellular Cu movement and, perhaps, signaling. It is unclear how Cu is loaded into the xylem for transport around the plant, although organic acids such as citrate, malate and oxalate, as well as phytochelatins (oligomers of glutathione, also found in some fungi) or nicotianimide may function as Cu-ligands in the vacuoles or xylem.
Copper in infectious disease
A striking example of how Nature has evolved multiple uses for elements is the observation that, while Cu is essential for many life processes, Cu is a potent anti-microbial weapon against invading pathogens. Indeed, Cu has been used as a bacteriocidal and fungicidal agent for over a century, with a notable use as the active ingredient in Bordeaux mixture that protects grapes from fungal infection. This anti-microbial activity of Cu seems to have been harnessed by eukaryotes, as there is mounting evidence that innate immune cells use Cu as an anti-microbial weapon. Studies have shown that activated macrophages accumulate Cu within the phagosome, the intracellular compartment that captures and disables invading microbes. Moreover, ATP7A protein levels are elevated in activated macrophages and the protein re-localizes from the secretory compartment to the phagosome, where there are parallel increases in phagosomal Cu levels. Taken together, these and other observations suggest that ATP7A may be responsible for the increased phagosomal Cu concentrations observed during infection. This increased pool of potentially toxic Cu in the lumen of the phagosome is not without impact on invading pathogens. Several reports demonstrate that bacterial cells up-regulate expression of the Cu export ATPases during infection in a manner dependent on Cu metalloregulatory transcription factors, suggesting that they mount a defense mechanism by enhancing their ability to export Cu. In fact, Mtb has at least two Cu resistance operons, governed by two distinct Cu-responsive transcription factors, underscoring the potential importance of Cu resistance mechanisms for a productive infection by Mtb. This Cu increase in the phagosome would add to the armamentarium of the phagosome that includes low pH, generation of reactive oxygen and nitrogen species, the elaboration of proteases and iron starvation to cope with invading pathogens.
While evidence is mounting that pathogenic microbes and their mammalian hosts do battle over Cu, this same dynamic may also occur in plant-pathogen interactions. Copper-containing amine oxidases, located around plant cell walls, utilize diamines and polyamines as substrates to produce hydrogen peroxide which has microbiocidal activity. Perhaps in an attempt to counteract this and other host defense mechanisms, the rice bacterial pathogen Xanthomonas oryzae expresses and secretes a protein effector, Xa13, that is proposed to directly interact with the COPT1 and COPT5 Cu transporters, leading to the redistribution of Cu out of the xylem where X. oryzae replicates, allowing pathogen dissemination. The observations that bacterial pathogens actively avoid intracellular Cu accumulation or alter Cu distribution in mammalian and plants hosts suggests that Cu status is a key battleground during infection.
Cu and human disease
Defects in Cu homeostasis lead to human disease. Wilson’s Disease describes the symptoms resulting from mutations in ATP7B, leading to hepatic and neuronal Cu accumulation. Wilson’s disease frequently leads to liver malfunction, neurological defects including movement disorders, seizures, and depression that is managed by low Cu diets, chelation therapy or in extreme cases by liver transplant. There is a potential link between Cu and Parkinson’s disease as Cu is associated with the accelerated aggregation of the alpha-synuclein protein in the formation of Lewy bodies. Similarly, Cu has been shown to promote the aggregation of mutant Huntington disease poly glutamine repeat proteins, as well as the neurotoxic oligomerization of the amyloid-beta peptide in the brains of Alzheimer’s disease patients. Whether Cu dysregulation is a cause or a consequence of these neurodegenerative diseases is under considerable investigation.
Menkes disease, caused by mutations in the ATP7A gene, results in peripheral Cu deficiency secondary to a failure in the mobilization of dietary Cu from intestinal enterocytes into the circulation. As an X-chromosome linked recessive trait, Menkes patients are typically male infants who present with seizure and profound neurological defects, abnormal thermoregulation, connective tissue disorders, immune cell dysfunction and other symptoms that typically lead to mortality prior to three years of age. While some cases can be partially ameliorated by intravenous Cu administration, the blockade in Cu delivery across the blood brain barrier renders this lethal neurodegenerative disease largely refractile to parenteral Cu supplementation. However, recent exciting studies with a mouse model of Menkes disease suggest that long-term rescue therapies might be developed by the delivery of a wild type version of the ATP7A into the ventricles of the brain.
Concluding remarks
While iron and zinc have long been known as metal ions that are important for life, it is clear that Cu is a critical metal in biology. Given that Cu is also a potentially dangerous toxin that is used by immune cells and its dysregulation causes human disease, the homeostasis of this metal ion must be under exquisite regulatory control. Many questions remain to be deciphered with respect to Cu in biology, including an articulation of the entire constellation of Cu-dependent processes, how hosts and microbes interact with respect to Cu, how cells and organs distribute and communicate their Cu status and precisely how Cu dysregulation factors into human disease.
Figure 2. Copper Homeostasis in Yeast.
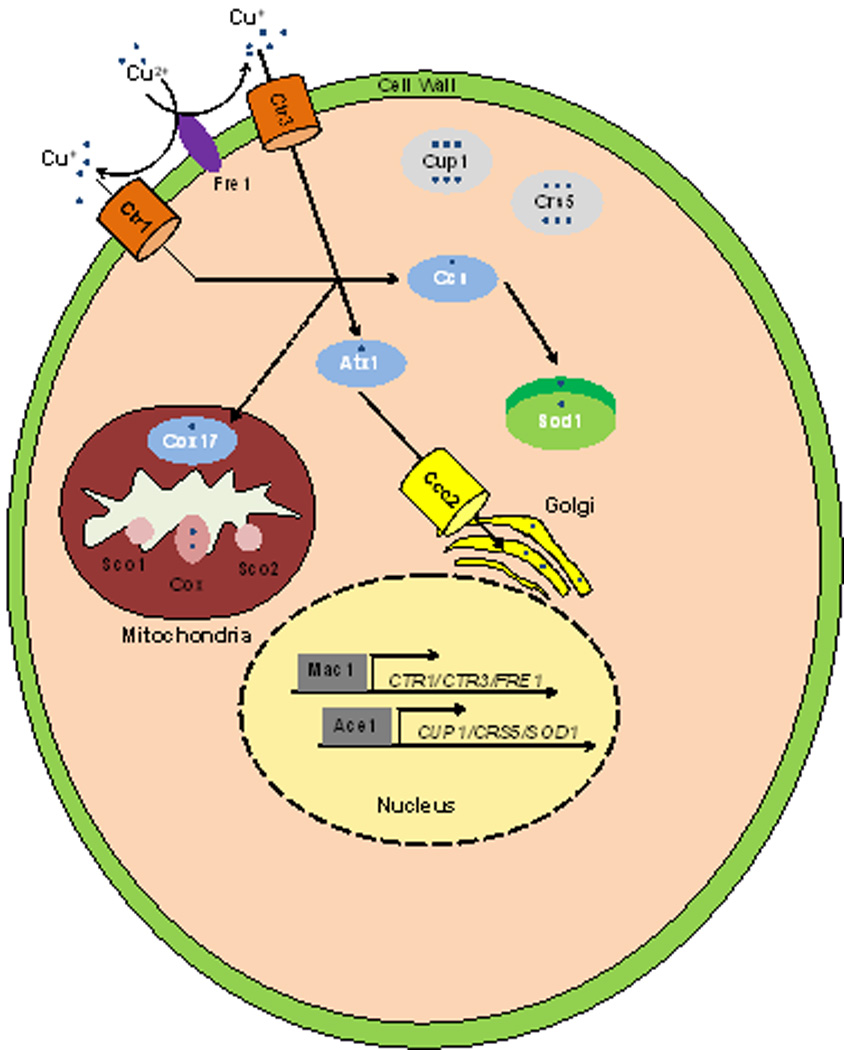
Unicellular organisms such as yeast have increased complexity to distribute Cu to intracellular proteins and compartments. Most notably, chaperones are utilized to safeguard the cell from free Cu and to expedite delivery, as the metal is delivered to intracellular Cu containing proteins. Ccs delivers Cu after reduction from Cu2+ to Cu+ by the Fre1 metalloreductase, and imported by the Ctr1 and Ctr3 Cu transporters, to Sod1 to protect against oxidative stress. Atx1 is a chaperone that delivers Cu to Ccc2 where Cu is translocated into the Golgi apparatus to be loaded onto cuproenzyems. Cup1 and Crs5 are metallothioneins that protect yeast cells from Cu toxicity. Mac1 and Ace1 are transcriptional activators that respond to Cu starvation and excess respectively. The precise mechanisms whereby cytosolic Cu is delivered to the mitochondria, for loading onto cytochrome oxidase (Cox) by Cox17, Sco1/2 and other proteins, is currently unknown.
Figure 3. Copper Homeostasis in Mammals.
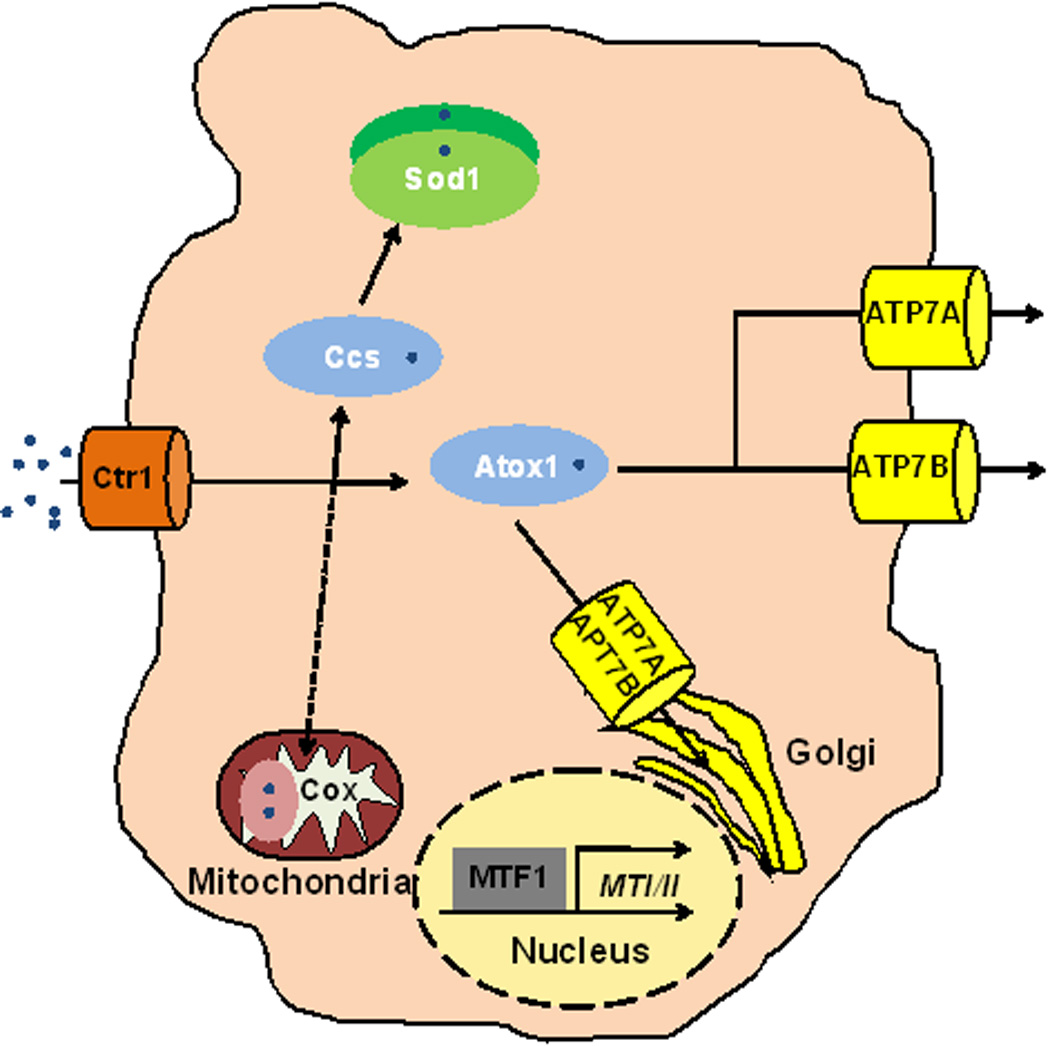
A generalized mammalian cell retains similar Cu homeostasis machinery to yeast, with increased complexity of components. Ctr1 is the predominant importer of Cu into mammalian cells. The Ccs and Sod1 chaperones are conserved from single cell eukaryotes. Atox1, the Atx1 homologue in mammals is responsible for delivery of Cu to the two P-type ATPases in mammals, ATP7A and ATP7B. These two P-type ATPases are also important for delivery of Cu to the Golgi to load onto proteins and in distinct cell types such as intestinal epithelial cells or hepatocytes, to efflux Cu. MTF1 transcriptionally activates metallothionein genes (MTI/II) and other targets. How Cu status is communicated between tissues must be explored to understand Cu homeostasis throughout metazoan organisms.
Figure 4. Copper Homeostasis in Plants.
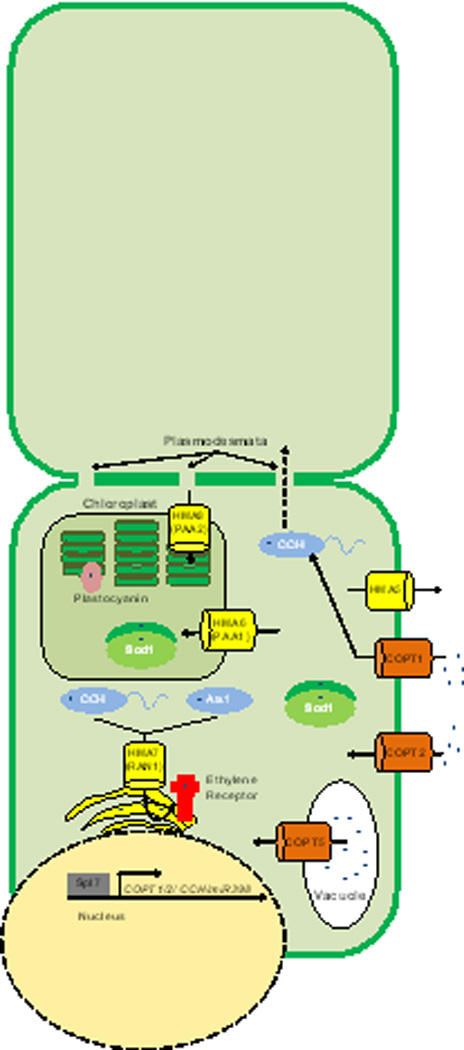
Plants have expanded the number of encoded Cu homeostasis proteins. The COPT proteins are similar to the Ctr transporters found in mammals and yeasts. COPT1 imports Cu and delivers it to CCH. CCH, while similar to Atox1, has a carboxy-terminal extension that may be important for cell-to-cell transport of Cu between plasmodesmata. Plants do encode another Atox1 homologue, ATX1. Copt5 has been associated with Cu export from the vacuole during times of Cu starvation. Chloroplast specific importers (PAA1 and PAA2) have also been identified and help to deliver Cu to important chloroplast specific proteins such as plastocyanin. RAN1 plays a similar role to Ccc2 by transporting Cu to the Golgi to be loaded onto nascent proteins such as the ethylene receptor. Spl7 is a Cu-responsive transcriptional regulator that is responsible for activation and expression of CCH, COPT1, and COPT2 among other genes, as well as micro RNAs that impact the expression of other Cu related genes such as that encoding Cu, Zn SOD1.
Further Reading
- Beaudoin J, Ioannoni R, Lopez-Maury L, Bahler J, Ait-Mohand S, et al. Mfc1 is a novel forespore membrane copper transporter in meiotic and sporulating cells. The Journal of biological chemistry. 2011 doi: 10.1074/jbc.M111.280396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boal AK, Rosenzweig AC. Structural biology of copper trafficking. Chem Rev. 2009;109:4760–4779. doi: 10.1021/cr900104z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Donsante A, Yi L, Zerfas PM, Brinster LR, Sullivan P, et al. ATP7A Gene Addition to the Choroid Plexus Results in Long-term Rescue of the Lethal Copper Transport Defect in a Menkes Disease Mouse Model. Molecular therapy : the journal of the American Society of Gene Therapy. 2011 doi: 10.1038/mt.2011.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim BE, Nevitt T, Thiele DJ. Mechanisms for copper acquisition, distribution and regulation. Nat Chem Biol. 2008;4:176–185. doi: 10.1038/nchembio.72. [DOI] [PubMed] [Google Scholar]
- Lutsenko S. Human copper homeostasis: a network of interconnected pathways. Curr Opin Chem Biol. 2010;14:211–217. doi: 10.1016/j.cbpa.2010.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nose Y, Kim BE, Thiele DJ. Ctr1 drives intestinal copper absorption and is essential for growth, iron metabolism, and neonatal cardiac function. Cell Metab. 2006;4:235–244. doi: 10.1016/j.cmet.2006.08.009. [DOI] [PubMed] [Google Scholar]
- Puig S, Andres-Colas N, Garcia-Molina A, Penarrubia L. Copper and iron homeostasis in Arabidopsis: responses to metal deficiencies, interactions and biotechnological applications. Plant Cell Environ. 2007;30:271–290. doi: 10.1111/j.1365-3040.2007.01642.x. [DOI] [PubMed] [Google Scholar]
- Ridge PG, Zhang Y, Gladyshev VN. Comparative genomic analyses of copper transporters and cuproproteomes reveal evolutionary dynamics of copper utilization and its link to oxygen. PLoS One. 2008;3:e1378. doi: 10.1371/journal.pone.0001378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tottey S, Harvie DR, Robinson NJ. Understanding how cells allocate metals using metal sensors and metallochaperones. Accounts of chemical research. 2005;38:775–783. doi: 10.1021/ar0300118. [DOI] [PubMed] [Google Scholar]
- White C, Lee J, Kambe T, Fritsche K, Petris MJ. A role for the ATP7A copper-transporting ATPase in macrophage bactericidal activity. J Biol Chem. 2009;284:33949–33956. doi: 10.1074/jbc.M109.070201. [DOI] [PMC free article] [PubMed] [Google Scholar]