

Abstract
Introduction
Ischemia/reperfusion (I/R) of the liver contributes to the pathobiology of liver injury in transplantation, liver surgery, and hemorrhagic shock. I/R induces an inflammatory response that is driven, in part, by TLR4 signaling. CD14 is known to participate in the function of TLR4. We hypothesized that CD14 would be involved in the pathobiology of warm hepatic I/R.
Methods
Using a 70% liver inflow inclusion model, CD14 knockout and wild type mice were subjected to 1h warm ischemia followed by reperfusion. CD14 mRNA levels, circulating transaminase, Interleukin-6, soluble CD14 and HMGB1 levels were measured. CD14 neutralizing antibody or isotype control antibody was given before ischemia or reperfusion for CD14 blockade in wild type mice. Recombinant HMGB1 was given before reperfusion in some experiments to test if liver injury worsens.
Results
There was an upregulation of CD14 mRNA in reperfused livers together with increased sCD14 levels in the circulation. Compared with wild type control mice, CD14 knockout mice had much lower ALT and IL-6 levels at 6h and 24h following I/R, and much less liver necrosis by histology. Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining displayed less apoptosis at 24h in the absence of CD14. CD14 blockage by neutralizing antibody also attenuated liver injury and the inflammatory response in C57BL/6 mice following I/R, but did not provide additional protection to toll-like receptor 4 mutant C3H/Hej mice. CD14 deficiency did not change circulating HMGB1 levels following I/R (6h). A dose of recombinant HMGB1 which worsened hepatic injury when given before reperfusion in wild type mice did not increase liver damage in CD14 deficient mice.
Conclusions
CD14 is actively involved in hepatic I/R injury. Its deficiency or blockade ischemia attenuates liver injury and inflammatory response. CD14 mediates liver damage and inflammatory responses in the setting of warm hepatic I/R in mice.
Keywords: CD14, sCD14, High mobility group box 1, hepatic ischemia reperfusion injury, toll-like receptor 4
Introduction
Hepatic ischemia reperfusion injury (I/R) plays an important role following liver transplantation, liver surgery, and hemorrhagic shock (1). Evidence has accumulated that TLR4 plays a central role in liver injury and the associated inflammation in warm (2, 3, 4) and cold (5) liver I/R. Endogenous TLR4 activators such as HMGB1 account for a significant part of TLR4 signaling in liver I/R (4). During LPS signaling, CD14 plays an important role in recognition of LPS by the TLR4/MD2 complex (6). We have recently shown that CD14 is also required for the recognition of HMGB1 by macrophages (7). However, the contribution of CD14 to I/R –induced liver injury or inflammation induced by warm I/R is not known. Here, we show CD14 deficiency or neutralization of CD14 suppressed both injury and inflammation in warm liver I/R. Furthermore, CD14 is required for the recognition of exogenous HMGB1 in the setting of I/R. Thus, CD14 is part of the host response to I/R in the liver.
Materials and methods
Reagents
Purified NA/LE rat anti-mouse CD14 antibody were purchased from BD Biosciences (San Jose, CA) and its Isotype control antibody was bought from Biolegend (San Diego, CA). Mouse CD14 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) primers were from Qiagen (Qiagen Sciences, MD). Mouse Interleukin-6 ELISA kit was from R&D System (Minneapolis, MN). Mouse sCD14 ELISA kit was purchased from Cellsciences (Canton, MA). HMGB1 ELISA kit was ordered from IBL International (Toronto, Canada). Recombinant mouse HMGB1 was kindly provided by Dr. Kevin Tracey. Anti-mouse CD14 and GADPH primary antibody were purchased from R&D Systems (Minneapolis, MN) and abcam (Cambridge, MA) respectively.
Animals
CD14 knock-out mice were provided by Dr Mason Freeman and were bred and maintained in the animal facility of the University of Pittsburgh. Wild type control CD57BL/6 mice, C3H/Hej and C3H/HeOuj mice were purchased from Jackson Laboratory (Bar Harbor, Maine). All mice were male, 8 to 12 weeks old and weighed 20 to 30g at the time of the experimental procedures. Animal handling and care complied with the regulations regarding the care and use of experimental animals published by the National Institutes of Health and was approved by the Institutional Animal Use and Care Committee of the University of Pittsburgh. The animals were maintained in the animal facility with a 12-hour light-dark cycle and free access to standard laboratory chow and water.
Warm hepatic ischemia reperfusion injury
Mice 8–12 weeks old were anesthetized with ketamine (100mg/kg) and Xylazine (10mg/kg) by intraperitoneal injection. After a midline laparotomy, the hepatic lobes were freed by dividing the surrounding ligaments. The vessels supplying to the left side 70% of liver were disrupted by a micro atraumatic vascular clamp, while the blood supply to the remaining liver lobes still remained patent to avoid intestinal venous congestion. The incision was then covered with a piece of saline-soaked gauze, together with a piece of plastic film, to reduce dehydration and mice were placed on a heating pad to maintain a rectal temperature of 37°C. The clamp was subsequently removed after 60 minutes to initiate hepatic reperfusion, and the abdominal incision was closed with running 4-0 Vicryl-plus (Ethicon) sutures. Sham treated mice underwent the same manipulation except the vascular clamp was not applied.
Sample collection
At the conclusion of the experiment, animals were anesthetized with isoflurane, and blood was harvested by cardiac puncture. Serum was collected by centrifuging the blood at 5,000 revolutions per minute for 10min at 4°C. Serum was aliquoted and stored at −80°C until further analysis. The same left hepatic lobe was harvested and snap-frozen in liquid nitrogen and then stored at −80°C degrees. The liver was also removed and fixed in 10% neutral formalin for Hematoxylin and Eosin (HE) staining. For immunofluoresence staining, liver was flushed with Phosphate-Buffered Saline (PBS) and then perfused with 2% Para formaldehyde. After fixation for 2h, the tissues were switched to 30% sucrose for 24h with sucrose change a total of three times. The samples were cryoperserved in 2-Methylbutane and stored at −80°C until ready for sectioning.
Alanine aminotransferase measurement
Serum alanine aminotransferase was measured with HESKA Dri-Chem 4000 (HESKA; slides from Fujifilm Japan).
RT-PCR
The harvested frozen liver lobes were pulverized and an aliquot of 20–30ug from each mouse was used to extract total RNA with Qiagen RNeasy kit (Qiagen) following the standard protocol. A total of 1μg RNA was used for cDNA synthesis using iScript™ Reverse Transcription Supermix (Bio-Rad) according to the manufacturer’s instructions. Pre-designed CD14 and GAPDH primers from Qiagen were used. PCR was performed on a conventional thermocycler and cycle conditions were as the following: 95°C for 30s, 55°C for 30s, and 72°C for 35s. The PCR products were run on a 1.2% agarose gel to compare the relative intensity semi-quantitatively with Adobe Photoshop CS5 (Adobe Systems).
Western blot
Western blot analysis was used to assess CD14 protein levels in whole liver. Fifty micrograms of liver homogenate was separated by SDS-PAGE and transferred onto a nitrocellulose membrane. The membrane was blocked for 1hr in PBS-Tween (0.1%) with 5% milk, followed by immunostaining with optimized dilutions of a polyclonal rabbit anti-mouse CD14 antibody (1:2,000) in 1% milk in PBS-Tween overnight at 4°C. Horseradish peroxidase-conjugated secondary antibodies were given, and membranes were developed with the Super Signal West Pico chemiluminescent kit (Thermo Fisher Scientific) and exposed to film. GADPH was used as the loading controls for each group.
Enzyme-linked immune sorbent assay (ELISA) for sCD14 and IL-6
sCD14 and IL-6 levels in serum was detected with ELISA kits according to their standard protocols. To measure IL-6 level in liver tissue, liver homogenates were lysed with cell lysis buffer with protease inhibitors added. After centrifugation at full speed for 15min at 4 degree, supernatants were harvested and protein levels were measured with BCA kit. Then the IL-6 level of each ischemic liver was measured by ELISA kit and standardized by its protein concentration.
Serum HMGB1 level measurement
Serum HMGB1 levels were measured with the HMGB1 ELISA kit developed by Shino-test Corporation according to its standard protocol.
TdT-mediated dUTP digoxigenin nick end labeling assay
Livers were fixed in 2% paraformaldehyde followed by standardized protocol for cryopreservation. Apoptosis was measured with terminal deoxynucleotidyl-transferase dUTP nick end-labeling (TUNEL) assay using a kit purchased from Promega following manufacturer’s protocol and counterstained with Hoechst nuclear stain. TUNEL positive cells were quantitated using a Metamorph image acquisition and analysis system (Chester, PA) incorporating a Nikon microscope (Nikon, Melville, NY). The number of positive stained cells per randomized high power field were calculated and compared.
Histopathology
Liver samples were fixed in 10% formalin and embedded in paraffin, then cut to 6-μm thick sections. Tissues were stained with H&E, and slides were assessed for inflammation and tissue damage.
Statistical analysis
Results are expressed as the mean ± standard error of mean (SEM). We used SigmaPlot 11.0(Systat Software, Inc, Point Richmond, CA) for the statistical analysis. One-way ANOVA was used for multiple groups and comparisons of two groups under the same treatment were performed using the Student’s t-test. Significance was established at the 95% confidence (P < 0.05).
Results
1. CD14 expression increases following warm hepatic I/R
We first assessed the level of CD14 expression in the liver and circulation following liver ischemia and reperfusion. Ischemia alone did not change CD14 mRNA expression from baseline (Fig. 1a and Fig. 1b). However, reperfusion induced a marked up-regulation of CD14 mRNA in the liver that was first detected at 1 hour of reperfusion. CD14 protein levels in the ischemic lobe of the liver decreased with the start of ischemia but then increased again between 3 and 6 hours following reperfusion (Fig. 1c). Control mice showed detectable but low sCD14 levels (Fig.2). Ischemia alone did not increase serum sCD14 levels. However, serum sCD14 were significant higher than baseline at 6h and 24h following reperfusion (P<0.01, comparison with baseline levels). These observations show that CD14 gene expression is increased by hepatic I/R and that liver I/R is associated with release of sCD14 into the circulation. It is noteworthy that several cell types in the liver, including hepatocytes express CD14. Total CD14 levels reflect intracellular and cell surface expression of CD14.
Fig. 1.


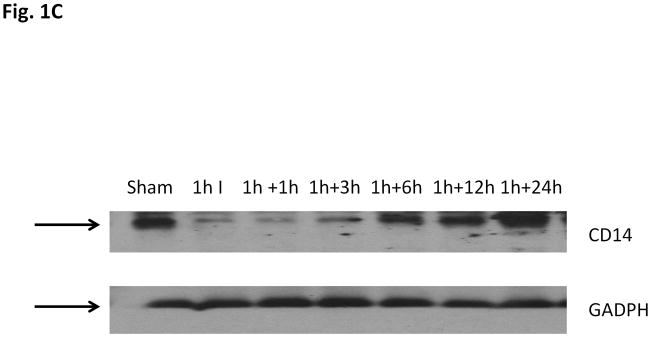
Fig. 1a Upregulation of CD14 mRNA after hepatic I/R.
CD14 mRNA levels measured by RT-PCR in the livers of control mice and mice subjected to ischemia (I, 60 minutes) and reperfusion (R) for the indicated time periods. A representative gel of three similar experiments showed upregulation of CD14 mRNA with the start of reperfusion.
Fig. 1b Quantification of CD14 mRNA expression after hepatic I/R. Compared with CD14 mRNA level in control livers, there is no difference with 1h ischemia only, but with the start of reperfusion, its expression was significantly increased (**P<0.01, n=3/group).
Fig. 1c CD14 protein levels in hepatic I/R livers. A representative western blot analysis result of CD14 protein levels in homogenates of the liver lobes subjected to I/R. The blot shown is representative of five experiments yielding similar results.
Fig. 2. Time-course of sCD14 levels in the circulation after hepatic I/R.

Wild type mice were subjected to I/R and serum assessed for sCD14 levels at the indicated time points. sCD14 levels at 6hr and 24hr of reperfusion were significantly higher than the control group (**both P<0.01, Values are expressed as mean ± SEM, n=6–8/ group).
2. CD14 contributes to liver injury following I/R
We next carried out experiments to determine if CD14 contributed to I/R-induced liver injury. CD14 knockout mice and their wild type counterparts were subjected to 1hr of ischemia followed by 6 or 24hr of reperfusion. There was no difference in ALT levels between wild type and CD14 Knockout sham groups. However, circulating ALT levels were significantly higher in wild type mice subjected to I/R at both 6 and 24hr than CD14 knockout mice (Fig. 3). Histological analysis confirmed that CD14 knockout mice were protected from the extensive central lobular necrosis observed at 6 and 24hr in the wild type mice following I/R (Fig. 4). I/R was also associated with apoptosis assessed by TUNEL staining at 24hr. Here again, CD14 deficiency was associated with the near absence of apoptosis (Fig. 5a and Fig. 5b). To confirm the importance of CD14 to I/R –induced injury we treated wild type mice with a neutralizing anti-CD14 antibody either prior to the onset of ischemia or just prior to reperfusion. As shown in Fig. 6, pre-treatment with anti-CD14 antibody significantly attenuated the injury as measured by circulating ALT levels. Administration of antibody just prior to reperfusion had minimal protective effects (Data not shown). Thus, CD14 participates in the pathobiology of I/R-induced liver injury and most likely is engaged early in the ischemia insult.
Fig. 3. ALT levels in wild type and CD14 knockout mice after hepatic I/R.

Wild-type and CD14 knockout mice were subjected to I/R and ALT levels measured a 6 and 24 hours. Values are expressed as mean ± SEM. There was no difference between the sham groups. CD14 knockout mice had significantly lower ALT levels after 6hr (** P<0.01, n=8–9) and 24hr of reperfusion (* P<0.05, n=5–7).
Fig. 4. Histological findings after hepatic I/R.
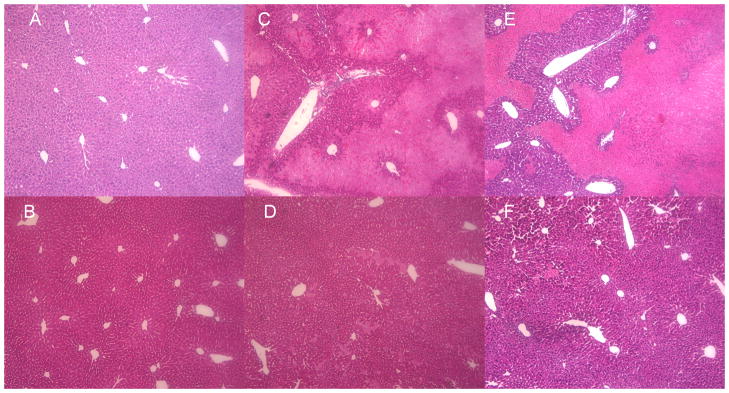
Representative histological liver images of WT and CD14 knockout mice subjected to I/R.
A. Shows representative normal liver structures in WT control mice (× 100)
B. Shows representative normal liver structures in CD14KO control mice (× 100)
C. Shows representative WT liver histological changes after 1h ischemia+6h reperfusion (× 100).
D. Shows representative CD14KO mice liver structure after 1h ischemia+ 6h reperfusion (× 100).
E. Shows representative WT liver after 1h ischemia+24h reperfusion (× 100).
F. Shows representative CD14KO mice after 1h ischemia+ 24h reperfusion (× 100).
Fig. 5.


Fig. 5a Apoptosis in WT and CD14KO mice after hepatic I/R by TUNEL staining and morphology
Representative TUNEL staining images of WT and CD14KO mouse livers.
A. Negative staining in WT control liver (× 100)
B. CD14 knockout control liver (× 100)
C. WT liver after 1h ischemia+6h reperfusion (× 100)
D. CD14 knockout liver after 1h ischemia+ 6h reperfusion (× 100)
E. WT liver after 1h ischemia+24h reperfusion (× 100)
F. CD14 knockout liver after 1h ischemia+ 24h reperfusion (× 100)
Fig. 5b Quantitative analysis of positive TUNEL-staining cells. Positive stained cell numbers per high power field were compared between WT, CD14KO control groups and I/R groups. Results are expressed as mean±SEM, ** P<0.01, 3–4 mice/group.
Fig. 6. CD14 Neutralizing antibody treatment reduced liver I/R injury.

Anti-CD14 or isotype control antibody was given to wild type mice 30 minutes prior to I/R. Liver injury was assessed by measuring circulating ALT levels at 6 hrs. Values are expressed as mean±SEM (n=5–6/group) *P<0.05.
3. CD14 contributes to the inflammatory response after hepatic I/R
I/R –induced liver injury is the result of both ischemic cell death and the inflammatory response that is induced by I/R. To determine if CD14 is involved in I/R-induced inflammation, we measured IL-6 in the liver and circulation as a marker of the inflammatory response. CD14 knockout mice showed much lower elevations in IL-6 in the liver (Fig. 7a) and circulation (Fig. 7b) than seen in the wild type mice subjected to I/R. Wild type mice pretreated with anti-CD14 antibody also had lower circulating IL-6 levels following I/R (Fig. 7c). Thus, CD14 is involved in the regulation of I/R –induced inflammation.
Fig. 7. IL-6 levels after hepatic I/R following treatment with anti-CD14 antibody.



Fig. 7a: Comparison of IL-6 levels in ischemic liver hemogenates after hepatic I/R between WT and CD14 Knockout mice. Values are expressed as mean ± SEM. n=4–6/ group. * P<0.05; Fig.7b: IL-6 levels in the circulation after hepatic I/R in WT and CD14 knockout mice. Values are expressed as mean ± SEM. * P<0.05, n=4–6; Fig.7c : Serum IL-6 levels after hepatic I/R in wild type pretreated with control or anti-CD14 antibody. Values are expressed as mean ± SEM. n=5/ group. * P<0.05
4. Neutralizing CD14 does not reduce I/R-induced injury in TLR4 deficient mice
We (4) and others (2, 3) have shown that TLR4 is involved in I/R-induced liver damage. Because CD14 is part of the TLR4 receptor complex, we sought evidence that CD14 might participate with TLR4 in liver I/R injury. Anti-CD14 antibody was administered to both wild type and TLR4 mutant mice. As expected, TLR4 mutant mice were partially protected from I/R-induced damage (Fig. 8a). No further reduction in liver injury was seen by neutralizing CD14 in these mice (Fig. 8b).
Fig. 8. Liver injuries of CD14 blockade in TLR4 mutant mice and HMGB1 treatment in CD14 deficient mice.




Fig. 8a: Liver injury after hepatic I/R in C3H/Hej and C3H/HeOuj mice was assessed by measuring circulating ALT levels. Values are expressed as mean ± SEM n=7/ group. ** P<0.01; Fig.8b: ALT levels after hepatic I/R in C3H/Hej mice treated with anti-CD14 antibody. Values are expressed as mean ± SEM. n=4–7/ group. P=0.75; Fig.8c: HMGB1 levels after hepatic I/R. Values are expressed as mean ± SEM. n=6–8, P=0.23; Fig. 8d: Liver I/R injury measured as serum ALT levels in WT and CD14 knockout mice treated with recombinant HMGB1 (55 ug/mouse) just prior to reperfusion. Values are expressed as mean ± SEM. n=8 - 9 per group, * P<0.05, control I/R compared to wild type mice treated with rHMGB1; P=0.795, control I/R mice compared to CD14 knockout mice treated with rHMGB1.
5. Recombinant HMGB1 given before reperfusion does not increase liver injury in CD14KO mice
HMGB1 is a prototypical DAMP previously shown to contribute to liver damage in I/R through TLR4. Furthermore, rHMGB1 exacerbates I/R-induced liver damage via a TLR4-dependent mechanism (4). We found no difference in circulating HMGB1 levels between wild type and CD14 KO mice at 6hrs suggesting that CD14 is not required for early HMGB1 release (Fig. 8c). We administered 55ug/mouse rHMGB1 (dose based on a pilot study, not shown), which did not result in any detectable liver injury by itself, to both WT and CD14KO mice intraperitoneally. This dose of HMGB1 did not induce liver injury in control mice. Whereas rHMGB1 worsened liver damage when administered at reperfusion to wild type mice, no increase in damage was seen in CD14 KO mice receiving rHMGB1(Fig. 8d). Taken together, these data suggest that CD14 is involved in sensing extracellular HMGB1 in liver I/R.
Discussion
This study was undertaken to determine if CD14 is involved in I/R-induced liver injury. This question is important because CD14 is known to participate in ligand recognition by the TLR4 receptor complex (6) and TLR4 is involved in the injury and inflammation induced by warm I/R in liver (4). Moreover, recently Li and colleagues reported that I/R could up-regulate CD14 expression in Kupffer cells in rats (8). We show here that CD14 is involved in both the injury and the inflammation induced by liver I/R. Whereas the release of HMGB1 in liver I/R did not require CD14, the recognition of HMGB1 does. These findings suggest that DAMP recognition in liver I/R involves CD14.
CD14, a 55 kDa cell surface glycoprotein that together with Toll-like receptor 4 and MD-2, act as a receptor for endotoxin (6). CD14 exists in two forms: one is anchored to the cellular membrane by a glycosylphosphatidyl inositol tail, i.e. mCD14, and the other a soluble form (sCD14). mCD14 is mainly expressed on macrophages, neutrophils, and dendritic cells, but epithelial cells(9), endothelial cells(10) and fibroblasts(11) also express low levels of mCD14. Although mononuclear phagocytes are known to shed their membrane-expressed CD14 (12) and liver can secret CD14 (13), the origin of sCD14 in plasma is not yet totally clear. sCD14 may confer LPS-responsiveness to cells that do not express CD14 (14). Blood and other body fluids contain sCD14, and levels increase during inflammation and infection. Therefore, sCD14 can be regarded as an acute phase protein (15). Here we show that CD14 mRNA levels in the liver increase after the hepatic I/R and that this corresponds to an increase in circulating sCD14 levels. It is unclear if the source of this sCD14 is from the liver. These results do indicate that I/R is a potent stimulus for CD14 upregulation and release.
CD14 knockout mice resist 10 times the LD100 dose of LPS (16). CD14 neutralizing antibody prevented lethality after in vivo exposure to endotoxin in rabbits and primates (17, 18). Neutralizing antibody to mouse CD14 even protects mice sensitized with galactosamine against LPS challenge (19). Our group found that the essential components for LPS uptake by hepatocytes were CD14, TLR4, MD2, and the beta2-integrin CD11b/CD18 of hepatocytes (20). Although LPS is considered as its main ligand, previous studies have shown that CD14 can also recognize other pathogen-associated molecular patterns, including peptidoglycan, lipoteichoic acid (LTA) and lipoarabinomannan (LAM)(21). Recent findings show that CD14 can promote TLR4 endocytosis and interferon expression, and CD14-induced endocytosis occurs independent of TLR4 signaling (22). Moreover, CD14 also enhances TLR2 (23) or TLR3-dependent cellular activation (24). In this study, we have shown that CD14 is actively involved in this sterile model of hepatic I/R injury, and CD14 deficiency or blockade before ischemia attenuates liver injury and inflammatory response in hepatic I/R. Attempts to reduce liver damage through delayed anti-CD14 antibody administration were less successful, suggesting that CD14 is involved in the early events in hepatic I/R.
The proinflammatory properties of HMGB1 were first identified in sepsis models where HMGB1 was show to be a late mediator of lethality (25). In contrast to sepsis, HMGB1 acts as an early mediator in models of sterile inflammation. For example, we have previously shown that HMGB1 contributes to TLR4 dependent signaling in liver I/R and that rHMGB1 exacerbates I/R induced injury in the liver in a TLR4-dependent manner(4). Here, we show that CD14 is also required for the exacerbation in I/R injury induced by rHMGB1. We have also reported that CD14 is involved in the injury and inflammation in cold cardiac I/R, a model that would involve no PAMP exposure (26). Therefore, CD14 is likely to be involved in the detection of DAMPs, such as HMGB1 released in I/R injury.
Whether I/R responses depend on sCD14 or mCD14 or both deserves further exploration. Since HMGB1 has been shown to bind to LPS and facilitate transfer of LPS to CD14 and enhance human monocyte activation (27), it is possible that HMGB1 can also bind other DAMPs. One potential candidate is HSP 70. Similar to HMGB1, it can be released actively from stressed cells (28) or passively from necrotic cells (29). Asea et al (30) reported CD14 was a co-receptor for HSP70-mediated signaling in human monocytes. Exogenous HSP70 bound with high affinity to the plasma membrane of monocytes, caused monocyte activation in both CD14 dependent and CD14 independent pathways.
Taken together, our findings provide new insights into a novel role for CD14 in I/R. A better understanding of its mechanisms may ultimately lead to the development of new methods to prevent warm hepatic I/R injury.
Acknowledgments
The authors acknowledge Qingde Wang, Pei Zhou and Rick Shapiro’s technical supports in this study.
This work was supported grants from National Institute of General Medical Sciences Grant P50-GM-53789.
Abbreviations
- I/R
ischemia/reperfusion
- HMGB1
high mobility group box 1
- ALT
alanine aminotransferase
- IL-6
interleukin-6
- TLR
toll-like receptor
- CD14
cluster of differentiation 14
- sCD14
soluble cluster of differentiation 14
- TNF-α
tumor necrosis factor alpha
- ELISA
enzyme-linked immune sorbent assay
- WT
wild type
- Ctrl
control
- Ab
antibody
- mCD14
membrane cluster of differentiation 14
- LD100
lethal dose 100 percent
- PAMP
pathogen-associated molecular pattern
- DAMP
danger-associated molecular pattern
- HSP
heat shock protein
References
- 1.Clemens MG, Bauer M, Pannen BH, Bauer I, Zhang JX. Remodeling of hepatic microvascular responsiveness after ischemia/reperfusion. Shock. 1997;8 (2):80–85. doi: 10.1097/00024382-199708000-00002. [DOI] [PubMed] [Google Scholar]
- 2.Wu HS, Zhang JX, Wang L, Tian Y, Wang H, Rotstein O. Toll-like receptor 4 involvement in hepatic ischemia/reperfusion injury in mice. Hepatobiliary Pancreat Dis Int. 2004;3 (2):250–253. [PubMed] [Google Scholar]
- 3.Zhai Y, Shen XD, O’Connell R, Gao F, Lassman C, Busuttil RW, Cheng G, Kupiec-Weglinski JW. Cutting edge: TLR4 activation mediates liver ischemia/reperfusion inflammatory response via IFN regulatory factor 3-dependent MyD88-independent pathway. J Immunol. 2004;173 (12):7115–7119. doi: 10.4049/jimmunol.173.12.7115. [DOI] [PubMed] [Google Scholar]
- 4.Tsung A, Sahai R, Tanaka H, Nakao A, Fink MP, Lotze MT, Yang H, Li J, Tracey KJ, Geller DA, Billiar TR. The nuclear factor HMGB1 mediates hepatic injury after murine liver ischemia-reperfusion. J Exp Med. 2005;201 (7):1135–1143. doi: 10.1084/jem.20042614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Shen XD, Ke B, Zhai Y, Gao F, Tsuchihashi S, Lassman CR, Busuttil RW, Kupiec-Weglinski JW. Absence of toll-like receptor 4 (TLR4) signaling in the donor organ reduces ischemia and reperfusion injury in a murine liver transplantation model. Liver Transpl. 2007;13 (10):1435–1443. doi: 10.1002/lt.21251. [DOI] [PubMed] [Google Scholar]
- 6.Miller SI, Ernst RK, Bader MW. LPS, TLR4 and infectious disease diversity. Nat Rev Microbiol. 2005;3(1):36–46. doi: 10.1038/nrmicro1068. [DOI] [PubMed] [Google Scholar]
- 7.Kim Sodam, Kim Sun Young, Pribis John P, Lotze Michael, Mollen Kevin P, Shapiro Richard, Loughran Patricia, Scott Melanie J, Billiar Timothy R. Signaling of High Mobility Group Box 1 (HMGB1) through Toll-like Receptor 4 in Macrophages Requires CD14. Mol Med. doi: 10.2119/molmed.2012.00306. (in press) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Luan X, Liu Y, Li M. The role of CD14 and Toll-like receptor 4 of Kupffer cells in hepatic ischemia- reperfusion injury in rats. Transplant Proc. 2012;44 (4):937–941. doi: 10.1016/j.transproceed.2011.11.001. [DOI] [PubMed] [Google Scholar]
- 9.Funda DP, Tuckova L, Farre MA, Iwase T, Moro I, Tlaskalova-Hogenova H. CD14 is expressed and released as soluble CD14 by human intestinal epithelial cells in vitro: lipopolysaccharide activation of epithelial cells revisited. Infect Immun. 2001;69 (6):3772–3781. doi: 10.1128/IAI.69.6.3772-3781.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jersmann HP, Hii CS, Hodge GL, Ferrante A. Synthesis and surface expression of CD14 by human endothelial cells. Infect Immun. 2001;69 (1):479–485. doi: 10.1128/IAI.69.1.479-485.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Stoll LL, Denning GM, Li WG, Rice JB, Harrelson AL, Romig SA, Gunnlaugsson ST, Miller FJ, Jr, Weintraub NL. Regulation of endotoxin-induced proinflammatory activation in human coronary artery cells: expression of functional membrane-bound CD14 by human coronary artery smooth muscle cells. J Immunol. 2004;173 (2):1336–1343. doi: 10.4049/jimmunol.173.2.1336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bufler P, Stiegler G, Schuchmann M, Hess S, Krüger C, Stelter F, Eckerskorn C, Schütt C, Engelmann H. Soluble lipopolysaccharide receptor (CD14) is released via two different mechanisms from human monocytes and CD14 transfectants. Eur J Immunol. 1995;25 (2):604–610. doi: 10.1002/eji.1830250244. [DOI] [PubMed] [Google Scholar]
- 13.Pan Z, Zhou L, Hetherington CJ, Zhang DE. Hepatocytes contribute to soluble CD14 production, and CD14 expression is differentially regulated in hepatocytes and monocytes. J Biol Chem. 2000;275 (46):36430–36435. doi: 10.1074/jbc.M003192200. [DOI] [PubMed] [Google Scholar]
- 14.Pugin J, Schürer-Maly CC, Leturcq D, Moriarty A, Ulevitch RJ, Tobias PS. Lipopolysaccharide activation of human endothelial and epithelial cells is mediated by lipopolysaccharide-binding protein and soluble CD14. Proc Natl Acad Sci USA. 1993;90 (7):2744–2748. doi: 10.1073/pnas.90.7.2744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bas S, Gauthier BR, Spenato U, Stingelin S, Gabay C. CD14 is an acute-phase protein. J Immunol. 2004;172 (7):4470–4479. doi: 10.4049/jimmunol.172.7.4470. [DOI] [PubMed] [Google Scholar]
- 16.Haziot A, Ferrero E, Köntgen F, Hijiya N, Yamamoto S, Silver J, Stewart CL, Goyert SM. Resistance to endotoxin shock and reduced dissemination of Gram-negative bacteria in CD14-deficient mice. Immunity. 1996;4 (4):407–414. doi: 10.1016/s1074-7613(00)80254-x. [DOI] [PubMed] [Google Scholar]
- 17.Schimke J, Mathison J, Morgiewicz J, Ulevitch RJ. Anti-CD14 mAb treatment provides therapeutic benefit after exposure to endotoxin. Proc Natl Acad Sci USA. 1998;95 (23):13875–13880. doi: 10.1073/pnas.95.23.13875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Leturcq D, Moriarty AM, Talbott G, Winn RK, Martin TR, Ulevitch RJ. Antibodies against CD14 protect primates from endotoxin-induced shock. J Clin Invest. 1996;98 (7):1533–1538. doi: 10.1172/JCI118945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Le Roy D, Di Padoba F, Adachi Y, Glauser MP, Calandra T, Heumann D. Critical role of lipopolysaccharide-binding protein and CD14 in immune responses against Gram-negative bacteria. J Immunol. 2001;167 (5):2759–2765. doi: 10.4049/jimmunol.167.5.2759. [DOI] [PubMed] [Google Scholar]
- 20.Scott MJ, Billiar TR. Beta2-integrin-induced p38 MAPK activation is a key mediator in the CD14/TLR4/MD2-dependent uptake of lipopolysaccharide by hepatocytes. J Biol Chem. 2008;283 (43):29433–29446. doi: 10.1074/jbc.M803905200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Anas A, Poll Tv, de Vos AF. Role of CD14 in lung inflammation and infection. Crit Care. 2010;14 (2):209–217. doi: 10.1186/cc8850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Zanoni I, Ostuni R, Marek LR, Barresi S, Barbalat R, Barton GM, Granucci F, Kagan JC. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Cell. 2011;147 (4):868–880. doi: 10.1016/j.cell.2011.09.051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Schroder NW, Morath S, Alexander C. Lipoteichoic acid (LTA) of Streptococcus pneumoniae and Staphylococcus aureus activates immune cells via Toll-like receptor (TLR)-2, lipopolysaccharide-binding protein (LBP), and CD14, whereas TLR-4 and MD-2 are not involved. J Biol Chem. 2003;278 (18):15587–15594. doi: 10.1074/jbc.M212829200. [DOI] [PubMed] [Google Scholar]
- 24.Lee HK, Dunzendorfer S, Soldau K, Tobias PS. Double-stranded RNA-mediated TLR3 activation is enhanced by CD14. Immunity. 2006;24 (2):153–163. doi: 10.1016/j.immuni.2005.12.012. [DOI] [PubMed] [Google Scholar]
- 25.Wang H, Ward MF, Sama AE. Novel HMGB1-inhibiting therapeutic agents for experimental sepsis. Shock. 2009;32 (4):348–357. doi: 10.1097/SHK.0b013e3181a551bd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kaczorowski DJ, Nakao A, Mollen KP, Vallabhaneni R, Sugimoto R, Kohmoto J, Tobita K, Zuckerbraun BS, McCurry KR, Murase N, Billiar TR. Mechanisms of Toll-Like receptor 4 (TLR4)-mediated inflammation after cold ischemia/reperfusion in the heart. Transplantation. 2009;87 (10):1455–1463. doi: 10.1097/TP.0b013e3181a36e5e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Youn JH, Oh YJ, Kim ES, Choi JE, Shin JS. High mobility group box 1 protein binding to lipopolysaccharide facilitates transfer of lipopolysaccharide to CD14 and enhances lipopolysaccharide-mediated TNF-alpha production in human monocytes. J Immunol. 2008;180 (7):5067–5074. doi: 10.4049/jimmunol.180.7.5067. [DOI] [PubMed] [Google Scholar]
- 28.Vega VL, Rodríguez-Silva M, Frey T, Gehrmann M, Diaz JC, Steinem C, Multhoff G, Arispe N, De Maio A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. J Immunol. 2008;180 (6):4299–4307. doi: 10.4049/jimmunol.180.6.4299. [DOI] [PubMed] [Google Scholar]
- 29.Zong WX, Thompson CB. Necrotic death as a cell fate. Genes Dev. 2006;20 (1):1–15. doi: 10.1101/gad.1376506. [DOI] [PubMed] [Google Scholar]
- 30.Asea A, Kraeft SK, Kurt-Jones EA, Stevenson MA, Chen LB, Finberg RW, Koo GC, Calderwood SK. HSP70 stimulates cytokine production through a CD14-dependant pathway, demonstrating its dual role as a chaperone and cytokine. Nat Med. 2000;6 (4):435–442. doi: 10.1038/74697. [DOI] [PubMed] [Google Scholar]