

Abstract
Background
Spermatogonial stem cells are subpopulation of spermatogonial cells in testis tissue that support beginning and maintenance of spermatogenesis. Ubiquitin carboxy-terminal hydrolase L1 (UCHL1) could be a specific marker for identification of spermatogonial stem cells including spermatogonial sperm cells (SSCs) in testis tissue and during the culture; therefore we undertook this study to culture these human testicular stem cells (hTSCs) in vitro and approved the presence of human testicular stem cells (hTSCs) by UCHL1, also known as PGP9.5.
Methods
Enzymatic digestion of human testicular biopsies was done by collagenase IV (4 mg/ml) and trypsin (0.25%). Differential plating of testicular cells in DMEM/F12 and 10% FBS was applied for 16 hr. Floating cells were collected and transferred onto laminin-coated plates with Stem-Pro 34 media supplemented with growth factors of GDNF, bFGF, EGF and LIF to support self-renewal divisions; testicular stem cell clusters were passaged every 14 days for two months. Spermatogonial cells propagation was studied through Expression of UCHL1 in testis tissue and the entire testicular stem cell culture.
Results
Testicular stem cell clusters from 10 patients with obstructive azoospermia were cultured on laminin-coated plates and subsequently propagated for two months. The average of harvested viable cells was approximately 89.6%. UCHL1 was expressed as specific marker in testicular stem cells entire the culture.
Conclusion
Human testicular stem cells could be obtained from human testicular tissue by a simple digestion, culturing and propagation method for long-term in vitro conditions. Propagation of these cells approved by specific marker UCHL1, during the culture period.
Keywords: Human testicular stem cells, Laminin, Long term culture, UCHL1
Introduction
Spermatogonial stem cells (SSCs) provide the foundation for spermatogenesis that begins with self-renewal and differentiation of spermatogonial stem cells (SSCs) (1). SSCs reside within a specialized microenvironment called niche that regulates testicular homeostasis by balancing SSC self-renewal and differentiation. A stem cell niche is comprised of cells, extracellular matrix components, and local soluble factors present in the vicinity of the stem cell that regulates cell fate. The structural basis for the SSC niche in the mammalian testis is the basal compartment of the seminiferous tubules and omposed of sertoli cells and peritubular myoid cells (2).
The establishment of human SSC culture in vitro would provide a model for studying SSC self-renewal and differentiation in vivo (3). However, there are very few advances in human spermatogonia and SSC research due to the restrictions on respective experiments, such as limitations to gain sufficient amounts of normal human testis for research intents (4). Another reason is the lack of specific markers for the identification of human SSCs. However, after more than forty years from the time Clermont constituted his findings, very little new progress is available on the identification of human spermatogonial stem cells, their renewal or differentiation (5).
Self-renewal division of SSCs is regulated by sertoli cells in the SSC niche. Growth factors such as Glial cell line-derived neurotrophic factor (GD NF) that support SSCs self-renewal are secreted by Sertoli cells (6). According to this observation, a long-term SSC culture was created which SSCs self-renewal and proliferation was promoted by the support of GDNF, Epidermal growth factor (E GF) or basic fibroblast growth factor (FGF2) and presence of fetal bovine serum (FBS) (7). Kanatsu shinohara et al. established serum-free and feeder-free culture of SSCs in vitro in 2005 (5). They suggested that presence of serum or feeder cells complicates the culture conditions making them uncontrollable (6).
In addition, to provide physical support for stem cell attachment, feeder cells also affect stem cells by producing various undefined factors through their interactions with stem cells (8). Sadri et al. cultured human testicular cells overnight in uncoated dishes in supplemented MEM (Minimum Essential Medium) containing 10% FCS at 37°C and 5% CO2. After overnight incubation, floating cells were collected and cultured in uncoated dishes with supplemented StemPro-34 and human growth factors (9).
He et al. isolated human spermatogonial cells by magnetic activated cell sorting (MACS), using GPR125 as a specific marker for spermatogonial cells in mouse. They studied more than six markers that have been identified for SSCs and progenitors in other species to characterize the phenotypes of human spermatogonia and more differentiated germ cells. They indicated that human spermatogonia share some but not all phenotypes with spermatogonial stem cells and progenitors from other species such as rodent, pig and nonhuman primates (10).
The isolation of SSCs is commonly performed through digestion of testicular tissue with a multi-step enzymatic process by using four proteolytic enzymes (11). In this study, we developed two enzyme digestion processes for SSC separation. Results demonstrate that two-step enzymatic digestion is a good method to isolate testicular cells. Digestion developed in our experiment was a modified one pursuant to the procedures described by previous researchers on the separation and culture of SSCs.
In the current study, we chose to examine the expression of ubiquitin carboxyl-terminal esterase L1 (UCHL1); also known as protein gene product 9.5, because it is expressed in spermatogonia of many species including bovine (2), pigs (12), mice (13), monkeys (14), etc. This marker may be involved in the asymmetric divisions (12). UCHL1 is a specific marker identified for spermatogonial and progenitors cells, but not for differentiated spermatogonia or sertoli cells. UCHL1 is expressed in type A (As, Apr, and Aal) spermatogonia located at the basement membrane (BM) of seminiferous tubules at high and low concentrations, but not in differentiated germ cells distant from the BM (12). We demonstrated the proliferation of human spermatogonia by in vitro culture for a long-term and identified them by UCHL1 as their specific marker.
Methods
Preparation of testicular cell suspension
Testicular tissues were obtained by biopsy from 10 patients with non-obstructive azoospermia at Avicenna Infertility Clinic after they signed an informed consent form. The volume of each biopsy was 25–35 mm 3. The biopsies were immediately transferred to Avicenna Research Institute in αMEM solution supplemented by penicillin 100 U/ml and streptomycin 100 mg/ml at 37°C. The testicular tissues were placed in culture plates containing PBS. Tissues were minced into small pieces digested with 4 mg/ml type IV collagenase (type IV, Sigma-Aldrich, USA) at 37°C with up and down for 30 min followed by 0.25% trypsin/1 mM EDTA (Invitrogen, USA) for 10 min with up and down. Trypsin activation was stopped by adding 10% fetal bovine serum. A nylon mesh with 100 µm pore sizes (BD falcon Cat No. 8343702) was used for removing cell clots or disturbing particles. We approximately collected 8×105 cells per each biopsy by this procedure. We calculated viabile cells by trypan blue for the estimation of their percentage after enzymatic digestion was performed. Cell suspention was centrifugated at 1200 rpm for 10 min at 25°C. Supernatant was discarded and pellet was suspended in DMED/F12 supplemented by nonessential amino acids, 15mM 4-[2-hydroxyethyl]-1 pi-perazineethanesulfonic acid [HEPES], 0.12% sodium bicarbonate, 4 mM L-glutamine (all from Invitrogen), 100 IU/ml penicillin, 100 µg/ ml streptomycin (Sigma-Aldrich), 40 µg/ml fangizone and 10% FBS.
Differential plating of testicular cells
Testicular cells were cultured in uncoated plates at 37°C and 5% CO2 for 16 hr. After the collection of floating cells and their culture in uncoated dishes with StemPro-34 SFM, supplemented with StemPro-34 supplement, 1-nonessential amino acids, 15 mM 4-[2-hydroxyethyl]-1 pi-perazineethanesulfonic acid [HEPES], 0.12% sodium bicarbonate, 4 mM L-glutamine (all from Invitrogen), 100 IU/ml penicillin, 100 µg/ml streptomycin, 40 µg/ml fangi-zone, 25 mg/ml insulin, 100 mg/ml transferrin, 60 mM putrescine, 30 nM sodiumselenite, 6 mg/ml D-(1)-glucose, 30 mg/ml pyruvic acid, 1 mg/ml DL-lactic acid (all from Sigma-Aldrich), 5 mg/ml bovine albumin (ICN Biomedical, USA), 2 mM L-glutamine, 5×10−5 M 2-mercaptoethanol, minimal essential medium (MEM) vitamin solution, MEM nonessential amino acid solution (all form Invitrogen), 10−4 M ascorbic acid, 10 mg/ml d-biotin, 30 ng/ml β-estradiol, 60 ng/ml progesterone, (20 ng/ml) recombinant human EGF, (10 ng/ml) recombinant human GDNF (all from Sigma-Aldri-ch), (10 ng/ml) recombinant human LIF (Chemicon International Inc., USA) and 5% fetal bovine serum, they were incubated at 37°C in a humidified atmosphere with 5% CO2. The number of dead cells was generally less than 5% as assessed by trypan blue staining. Germ line stem cell colonies were transferred on laminin-coated dishes (20 µg/ml) (Sigma-Aldrich) in supplemented Stem-Pro-34 after 14 days from initiating the cultures by pipet mouth or trypsination. Germ stem cell clusters were passaged by short-term trypsination (3 min) every 2 weeks.
Gene Expression
In this study, we specifically applied UCHL1 for the identification of human germ stem cells. Presence of spermatogonial stem cell clusters during the entire culture, was proved by studying the expression of spermatogonial genes (12, 13). Total RNA from cultured testicular cells, sub- cultured germ line stem cells and testicular tissue, as a positive control, were extracted. Reverse transcriptase polymerase chain reaction (PCR) was carried out by first-strand cDNA synthesized by random hexamers and the Superscript II preamplification system (Invitrogen). RT-PCR reaction was proceeded with specific primers for UCHL1 (PGP9.5) (Forward: GGC TGGAAGAGGAGTCTCTG, Reverse: CACAGG AATTCCCAATGGTC) with master mix kit (Taq DNA polymerase master mix RED Amplicon).
Results
Cell viability after enzymatic digestion of human testicular tissue
As we isolated SSCs through a simple digesting protocol, viable cells were counted in the cell suspension after two-step enzymatic digestion. Results demonstrated that the mean percentage of viable cells was approximately 89.6%. This result suggested viability of isolated cells after enzymatic process (Table 1).
Table 1.
Cell viability after tow-step enzymatic digestion
| Number of Counts | Total count (106) | Viable cells (106) | Cell viability (%) |
|---|---|---|---|
| 1 | 3.56 | 3.25 | 91.29 |
| 2 | 4.85 | 4.42 | 91.13 |
| 3 | 4.71 | 4.25 | 90.23 |
| 4 | 3.78 | 3.32 | 87.83 |
| 5 | 2.89 | 2.53 | 87.54 |
| Average | 3.95 | 3.55 | 89.60 |
Morphology of spermatogonial clusters during the culture
After culturing for 16 hr, a great number of testicular cells, mostly including somatic cells, adhered to culture plates. We isolated floating cells, containing germ, cells and transferred them into a new dish with StemPro34 supplement and 5% FBS. Under inverted microscope, the SSCs were seen as round or oval in shape with a large nucleus and little cytoplasm after one week of culture. The isolated SSCs tended to aggregate and form small cell clusters (Figure 1A) and attach to the remaining somatic cells. After 2 weeks of culture, SSCs began to form cell clusters (Figure 1B). These clusters were transferred into laminincoated plates containing the same media and 2% FBS. The medium was changed every 3–5 days. Under feeder-free conditions in laminincoated plates, extension of SSCs was observed for up to 6 weeks (Figure 1C and D).
Figure 1.

Spermatogonial Stem Cell (SSC) Clusters in vitro culture condition. A: cultured human testicular cells (arrows) before colony formation (7 days after digestion); B: Germ stem cell colonies (arrow) after 2 weeks; C and D: germ stem cell clusters (arrows) after 4 and 8 weeks of culture after being transferred into laminin. Scale bar 50 µm
The number of spermatogonial clusters in second, fourth and sixth weeks were 12.83±13.92, 69.95±24.79 and 178.15±65.65, respectively; therefore, the proliferative increase in the number of spermatogonial clusters from the second week until sixth week of culture was statistically significant (p < 0.001) and demonstrated the colony growth under culture conditions (Figure 3).
Figure 3.

Number of spermatogonial cluster proliferation during culture on laminin-coated plates, p < 0.001
Gene expression
RT-PCR was performed to confirm the expression of spermatogonial specific gene UCHL1 (ubiquitin carboxyl-terminal esterase L1; also known as protein gene product 9.5) in adult human testis and throughout the entire culture period at passages 2, 4 and 8, relative to a normal human testis sample. GAPDH primer proved cell presentation during RT-PCR process. Results demonstrated that the during culture hTS Cs express spermatogonial specific gene UCHL1 in the testis as shown in Figure 2.
Figure 2.
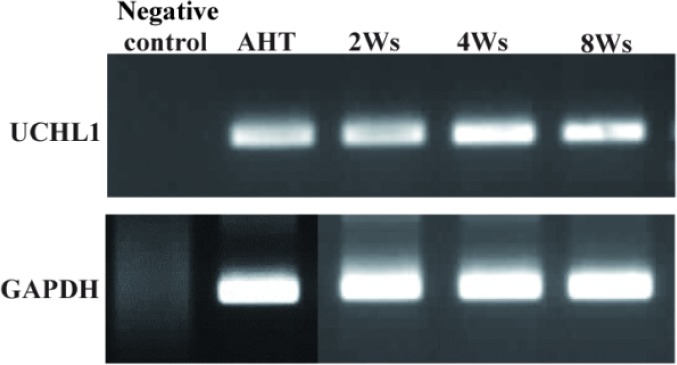
RT-PCR representation for spermatogonial stem cell marker UCHL1 in adult human testis (AHT) tissue and throughout the entire culture. GAPDH was used for cell presence
Discussion
SSCs are special types of stem cells that can transfer genetic information to the next generation (15). Studying the biology of SSCs provides better understanding of stem cell self-renewal and differentiation in the testis and a novel target for therapy in male infertility and testicular cancer (16). Spermatogenesis is a complex process that begins from SSC differentiation. In seminiferous epithelium during the fetal period, spermatogenic cells are hardly seen at different levels except for SSCs and supporting cells (7).
There are different methods for the digestion of testicular tissue. Complex digestion processes and time-consuming protocols cause injury cells and increase the possibility of cell infection (15, 17). Therefore, a simple digestion method is important for preparing uni-cell suspensions. In our study, we performed a two-step enzymatic digestion with less contamination through which we obtained a high rate of cell viability (89.6%) without reducing the total cell count. These results are similar to the results of He et al. in testicular tissue digestion protocol (10). They used 4 enzymes including collagenase, trypsin, DNase and hyaluronidase. In our study, digestion method was more simple and feasible in comparison to those of others. This study provided the first step to reproduce the self-renewal division of spermatogonial stem cells in vitro, and clearly showed the necessity for an improved culture system to allow more analysis of self-renewal division and its related niche.
Although, laminin was able to replace somatic cells as feeder layer, it is also important to establish culture conditions that direct a specific type of self-renewal division (18). In the current study, spermatogonial cells were transferred to laminincoated dishes after cluster formation for better stability and proliferation. The identification of crucial factors will increase our knowledge about the regulation of the self-renewal division of spermatogonial stem cells and lead us to develop more improved techniques for male germline modification (6). Currently, the conditions needed for a long-term stabilization of SSCs usually consists of adding growth factors such as bFGF, LIF, EGF and GDNF to the media. Kanatsu-Shinohara et al. established a long-term in vitro culture system of SSCs by a combination of growth factors such as GDNF that induced long-term survival and sustainable differentiation of SSCs (6).
In our culture condition, human spermatogonial stem cells increased in number by self-renewal in vitro after transfer to laminin. In this culture condition, germ stem cell (GSC) clusters presented as clumps of individually visible cells up to 8 weeks. We could characterize them by the expression of a specific marker, UCHL1, that had been proved earlier as a germ stem cell marker in many species (12).
According to previous studies, there are two strategies to continue SSC culture. The first is transferring colonies to plates that are coated with mytomycin C-inactivated somatic cells as feeder layer, thus, SSCs could maintain and proliferate for a long time (9). The second is to culture SSCs in feeder free plates or laminin-coated plates. Laminin is secreted by sertoli cells as an adhesion molecule and bind to ITGα6/β1 receptors on the spermatogonial cell surface. Therefore, laminin provides the best condition for SSC maintenance and proliferation for a long time (19).
Ubiquitin-dependent proteolysis has played an important role in the control of mammalian gametogenesis. UCHL1 is a deubiquitinating enzyme that regenerates monoubiquitin from ubiquitin-protein complexes. In the testis, UCHL1 is expressed in spermatogonia among male germ cells in many species (12). In mice, the specific role of UCHL1 has been proved in apoptosis, meiosis, and mitotic proliferation of germ cells. Lack of UCHL1 decreased apoptosis rate and increased the number of premeiotic germ cells in immature mice (13), but in adult mice spermatogenesis was damaged by spermatogonial reduction and primary spermatocytes which mainly undergo apoptosis (13). Overexpression of UCHL1 was associated with the limitation of apoptosis in spermatogonia and primary spermatocytes undergoing apoptosis. According to previous studies, co-expression of UCHL1 and promyelocyticleukaemia zinc finger protein PLZF in gonocytes and spermatogonia induced self-renewal and prevented differentiation (20). Ubiquitin-proteasome system regulates essential processes in mammalian stem and progenitor cells of embryonic, neural, hematopoietic, and mesenchymal origin (12). Establishment of human SSC culture from testicular biopsies might allow individual cell-based therapy for fertility preservation in cancer patient after completion of the treatment by SSC transplantation without ethical violations or immunological problems. Thus, SSCs will provide new possibilities in fertility preservation, biotechnology and medicine (21).
Conclusion
In conclusion, these results show that long-term culture and propagation of human spermatogonial stem cells in vitro is achievable. Propagation of these cells could be approved by specific marker, UCHL1, during the culture period.
Acknowledgement
Authors would like to express their gratitude to Avicenna Research Institute for supporting this study.
To cite this article: Piravar Z, Jeddi-Tehrani M, Sadeghi MR, Mohazzab A, Eidi A, Akhondi MM. In vitro Culture of Human Testicular Stem Cells on Feeder-Free Condition. J Reprod Infertil. 2013;14(1):17-22.
Conflict of Interest
The author declares no conflict of interest.
References
- 1.Clermont Y. Renewal of spermatogonia in man. Am J Anat. 1966;118(2):509–24. doi: 10.1002/aja.1001180211. [DOI] [PubMed] [Google Scholar]
- 2.Zhang ZJ, Burgunder JM, An XK, Wu Y, Chen WJ, Zhang JH, et al. Lack of evidence for association of a UCH-L1 S18Y polymorphism with Parkinson's disease in a Han-Chinese population. Neurosci Lett. 2008;442(3):200–2. doi: 10.1016/j.neulet.2008.07.012. [DOI] [PubMed] [Google Scholar]
- 3.Hofmann MC, Braydich-Stolle L, Dym M. Isolation of male germ-line stem cells; influence of GDNF. Dev Biol. 2005;279(1):114–24. doi: 10.1016/j.ydbio.2004.12.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kubota H, Avarbock MR, Brinster RL. Growth factors essential for self-renewal and expansion of mouse spermatogonial stem cells. Proc Natl Acad Sci U S A. 2004;101(47):16489–94. doi: 10.1073/pnas.0407063101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Clermont Y. Kinetics of spermatogenesis in mammals: seminiferous epithelium cycle and spermatogonial renewal. Physiol Rev. 1972;52(1):198–236. doi: 10.1152/physrev.1972.52.1.198. [DOI] [PubMed] [Google Scholar]
- 6.Kanatsu-Shinohara M, Miki H, Inoue K, Ogonuki N, Toyokuni S, Ogura A, et al. Long-term culture of mouse male germline stem cells under serum-or feeder-free conditions. Biol Reprod. 2005;72(4):985–91. doi: 10.1095/biolreprod.104.036400. [DOI] [PubMed] [Google Scholar]
- 7.Dym M, Kokkinaki M, He Z. Spermatogonial stem cells: mouse and human comparisons. Birth Defects Res Part C Embryo Today. 2009;87(1):27–34. doi: 10.1002/bdrc.20141. [DOI] [PubMed] [Google Scholar]
- 8.Clermont Y. The cycle of the seminiferous epithelium in man. Am J Anat. 1963;112:35–51. doi: 10.1002/aja.1001120103. [DOI] [PubMed] [Google Scholar]
- 9.Sadri-Ardekani H, Akhondi MA, van der Veen F, Repping S, van Pelt AM. In vitro propagation of human prepubertal spermatogonial stem cells. JAMA. 2011;305(23):2416–8. doi: 10.1001/jama.2011.791. [DOI] [PubMed] [Google Scholar]
- 10.He Z, Kokkinaki M, Jiang J, Dobrinski I, Dym M. Isolation, characterization, and culture of human spermatogonia. Biol Reprod. 2010;82(2):363–72. doi: 10.1095/biolreprod.109.078550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kanatsu-Shinohara M, Inoue K, Ogonuki N, Mori-moto H, Ogura A, Shinohara T. Serum- and feeder-free culture of mouse germline stem cells. Biol Reprod. 2011;84(1):97–105. doi: 10.1095/biolreprod.110.086462. [DOI] [PubMed] [Google Scholar]
- 12.Luo J, Megee S, Rathi R, Dobrinski I. Protein gene product 9.5 is a spermatogonia-specific marker in the pig testis: application to enrichment and culture of porcine spermatogonia. Mol Reprod Dev. 2006;73(12):1531–40. doi: 10.1002/mrd.20529. [DOI] [PubMed] [Google Scholar]
- 13.Rodriguez-Sosa JR, Foster RA, Hahnel A. Development of strips of ovine testes after xenografting under the skin of mice and co-transplantation of exogenous spermatogonia with grafts. Reproduction. 2010;139(1):227–35. doi: 10.1530/REP-09-0176. [DOI] [PubMed] [Google Scholar]
- 14.Tokunaga Y, Imai S, Torii R, Maeda T. Cytoplasmic liberation of protein gene product 9.5 during the seasonal regulation of spermatogenesis in the monkey (Macaca fuscata) Endocrinology. 1999;140(4):1875–83. doi: 10.1210/endo.140.4.6615. [DOI] [PubMed] [Google Scholar]
- 15.Dann CT, Alvarado AL, Molyneux LA, Denard BS, Garbers DL, Porteus MH. Spermatogonial stem cell self-renewal requires OCT4, a factor down-regulated during retinoic acid-induced differentiation. Stem Cells. 2008;26(11):2928–37. doi: 10.1634/stemcells.2008-0134. [DOI] [PubMed] [Google Scholar]
- 16.Liu S, Tang Z, Xiong T, Tang W. Isolation and characterization of human spermatogonial stem cells. Reprod Biol Endocrinol. 2011;9:141. doi: 10.1186/1477-7827-9-141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hermann BP, Sukhwani M, Lin CC, Sheng Y, Tomko J, Rodriguez M, et al. Characterization, cryopreservation, and ablation of spermatogonial stem cells in adult rhesus macaques. Stem Cells. 2007;25(9):2330–8. doi: 10.1634/stemcells.2007-0143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Wu X, Goodyear SM, Tobias JW, Avarbock MR, Brinster RL. Spermatogonial stem cell self-renewal requires ETV5-mediated downstream activation of Brachyury in mice. Biol Reprod. 2011;85(6):1114–23. doi: 10.1095/biolreprod.111.091793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kanatsu-Shinohara M, Ogonuki N, Iwano T, Lee J, Kazuki Y, Inoue K, et al. Genetic and epigenetic properties of mouse male germline stem cells during long-term culture. Development. 2005;132(18):4155–63. doi: 10.1242/dev.02004. [DOI] [PubMed] [Google Scholar]
- 20.Costoya JA, Hobbs RM, Barna M, Cattoretti G, Manova K, Sukhwani M, et al. Essential role of Plzf in maintenance of spermatogonial stem cells. Nat Genet. 2004;36(6):653–9. doi: 10.1038/ng1367. [DOI] [PubMed] [Google Scholar]
- 21.Meng X, Lindahl M, Hyvönen ME, Parvinen M, de Rooij DG, Hess MW, et al. Regulation of cell fate decision of undifferentiated spermatogonia by GD NF. Science. 2000;287(5457):1489–93. doi: 10.1126/science.287.5457.1489. [DOI] [PubMed] [Google Scholar]