

Abstract
Benign Prostate hyperplasia (BPH) and prostate cancer (PCa) are the most common prostatic disorders affecting elderly men. Multiple factors including hormonal imbalance, disruption of cell proliferation, apoptosis, chronic inflammation, and aging are thought to be responsible for the pathophysiology of these diseases. Both BPH and PCa are considered to be arisen from aberrant proliferation of prostate stem cells. Recent studies on BPH and PCa have provided significant evidence for the origin of these diseases from stem cells that share characteristics with normal prostate stem cells. Aberrant changes in prostate stem cell regulatory factors may contribute to the development of BPH or PCa. Understanding these regulatory factors may provide insight into the mechanisms that convert quiescent adult prostate cells into proliferating compartments and lead to BPH or carcinoma. Ultimately, the knowledge of the unique prostate stem or stem-like cells in the pathogenesis and development of hyperplasia will facilitate the development of new therapeutic targets for BPH and PCa. In this review, we address recent progress towards understanding the putative role and complexities of stem cells in the development of BPH and PCa.
1. Introduction
Prostate gland is a male accessory reproductive endocrine organ, which expels proteolytic solution in the urethra during ejaculation. In humans, the prostate is located immediately below the base of the bladder surrounding the neck region of the urethra. It is mainly associated with three types of disorders, namely, benign prostate hyperplasia (BPH), prostate cancer (PCa), and prostatitis. BPH and PCa are the most common pathophysiological conditions of prostate gland in elderly men. These diseases already represent significant challenges for health-care systems in most parts of the world. Epidemiologically, BPH is more prevalent in Asian population [1, 2]. Whereas, PCa is more common in the western world [3, 4]. Both the diseases are complex and multifactorial. Factors predisposing to the development of BPH or PCa include hormonal imbalance, oxidative stress, environmental pollutants, inflammation, hereditary, aging, and, more particularly, stromal to epithelial cells crosstalk [5–7]. So far, variety of growth factors and hormonal factors, including androgens and estrogens, has been described in the hyperplastic development of the prostate gland [8–10]. However, the cellular and molecular processes underlying the pathogenesis and development of BPH or PCa are poorly understood.
Stem cells have an extensive capacity to propagate themselves by self-renewal and to differentiate into tissue-specific progeny. It is well know that stem cells are required to maintain and repair tissues throughout the lifetime. The requirement to understand the biology of stem cells derived from the prostate is increasing, as new evidence suggests that BPH and PCa may arise from the stem or stem-like cell compartments [11–13]. This review summarises the biology of prostate stem or stem-like cells and their contribution in pathogenesis and development of BPH and PCa.
2. Prostatic Cellular Compartments
The prostate is a hormonally regulated glandular organ whose growth accelerates at sexual maturity due to androgen action on both stromal and epithelial cells [14, 15]. The human prostate is a complex ductal-acinar gland that is divided into three anatomically distinct zones: peripheral, transitional, and central zones, which are surrounded by a dense and continuous fibromuscular stroma [16–18]. BPH, a nonmalignant overgrowth found in older men, mainly, develops in the transitional zone, while PCa arises primarily in the peripheral zone [19].
At histological level, human prostate contains mainly two types of cells that are called epithelial and stromal cells. The stromal to epithelial ratio in normal prostate of human is 2 : 1 [18, 20]. The epithelial cell layer is composed of four differentiated cell types known as basal, secretory luminal, neuroendocrine (NE), and transit-amplifying (TA) cells that are identified by their morphology, location, and distinct marker expression (Figure 1). The basal cells form a layer of flattened to cuboidal shaped cells above the basement membrane and express p63 (a homolog of the tumor suppressor gene p53), Bc1-2 (an anti-apoptotic factor), Cluster designation (CD) 44, hepatocyte growth factor (HGF), and the high molecular weight cytokeratins (CK) 5 and 14. The expression of androgen receptor (AR) is low or undetectable in the basal cells, which makes the basal cells independent of androgens for their survival [21–23]. The luminal cells are the major cell type of the prostate that form a layer of columnar-shaped cells above the basal layer and constitute the exocrine compartment of the prostate, secreting prostate-specific antigen (PSA) and prostatic acid phosphatase (PAP) into the lumen. They are terminally differentiated, androgen dependent, and nonproliferating cells, expressing low molecular weight CK8 and 18, CD57 and p27Kip1 (a cell cycle inhibitor) [22–24] along with high levels of AR. NE cells are rare cells scattered in the basal and luminal layers of the prostate. They are terminally differentiated and androgen-insensitive cells, expressing chromogranin A, synaptophysin, and neuron-specific enolase (NSF) [23, 25, 26]. The NE cells also produce and secrete neuropeptides such as bombesin, calcitonin, and neurotensin that are believed to support epithelial cell growth and differentiation [19, 27, 28]. Additionally, there is a small group of intermediate cells referred to as TA cells that express both basal as well as luminal cell markers (CK5, CK8, CK14, CK18, AR, and PSA) [29–32]. The epithelial layer is surrounded by a stromal layer, which forms a peripheral boundary of the prostate gland. The stromal cell layer consists of several types of cells that include smooth muscle cells (the most abundant cell type in stroma), fibroblasts, and myofibroblasts. Stromal cells express mesenchymal markers like CD34, vimentin, CD44, CD117, and CD90 [33].
Figure 1.
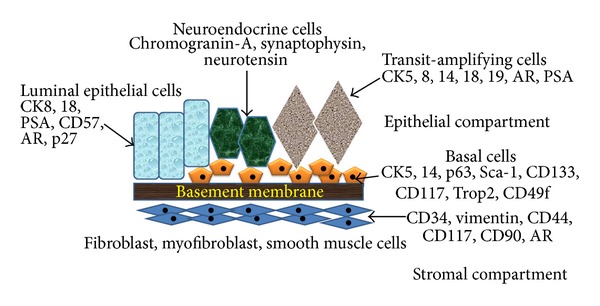
Prostatic cellular compartments and stem cell identity markers. Pictorial representation of different prostatic cells and their respective cellular markers.
3. Stem Cell in Normal Prostate
Prostatic epithelium is, structurally and functionally, a highly complex tissue composed of multiple differentiated cell types, including basal, luminal, and neuroendocrine cells, along with small population of relatively undifferentiated cells generally known as “stem cells” that are endowed with self-renewal and differentiation capacities [26]. If the stem cells are key target for mutagenic changes and tumourigenesis in human prostate, we need to understand more about stem cell status in normal prostate tissue.
As the adult prostate is relatively slow-growing organ with limited cycles of cell proliferation and apoptosis, the possible existence of adult prostate stem cells (PSCs) was controversial for many years. Several investigations based on stem cell models have elegantly defined role of stem cells in cellular turnover and morphogenesis of normal prostate [30, 34]. Evidence for the existence of the stem cells in normal prostate came from the studies which demonstrated that adult rodent prostate can undergo multiple rounds of castration-induced regression and testosterone-induced regrowth [35–37]. Adult PSCs were believed to reside within the basal cell layer because of the ability of the basal cells to survive and undergo regression and regeneration following repeated castration and androgen replacement [38–40]. Adult mouse prostate epithelial cells, when transplanted along with the urogenital sinus mesenchymal cells under the renal capsule, generated normal murine prostate like structures [41]. Prostate glands were also regenerated when dissociated cells were implanted in Matrigel subcutaneously into immunodeficient mice [42]. Studies, including 5-bromo-2-deoxyuridine (BrdU) retention analysis, showed that the enriched population of BrdU-labelled cells possessing stem cell features (quiescent, high proliferation potential) are localized at the proximal region of mouse prostate duct [43] and are programmed to regenerate proximal-distal ductal axis [44]. The proximal region of the prostatic duct is surrounded by a thick band of smooth muscle cells [45] that are known to produce high level of transforming growth factor-beta (TGF-β) [46], which is known to play a critical role in maintaining the relative dormancy of the PSCs [47]. Independent study by Burger et al. also identified a candidate population of PSCs in the proximal region of mouse prostatic ducts, using stem cell surface marker known as stem cell antigen 1 (Sca-1, also known as Ly6a) [48]. In addition to high expression of Sca-1, these cells were shown to coexpress integrin α6 (CD49f) and Bcl-2. The cells with these properties showed a higher efficiency to generate prostatic tissue in an in vivo reconstitution assay [48]. Lawson et al. showed that sorting prostatic cells for CD45(−)CD31(−)Ter119(−)Sca-1(+)CD49f(+) antigenic profile results in a 60-fold enrichment for colony and sphere-forming cells that can self-renew and expand to form spheres for many generations [49]. Leong and colleagues identified CD117 (c-Kit, stem cell factor receptor) as a new marker of a rare adult mouse PSC population that showed all the functional characteristics of stem cells including self-renewal and full differentiation potential. The CD117(+) single stem cell defined by the phenotype Lin(−)Sca-1(+)CD133(+)CD44(+)CD117(+) regenerated functional, secretion-producing prostate after transplantation in vivo. Moreover, CD117(+) PSCs showed long-term self renewal capacity after serial isolation and transplantation in vivo. CD117 expression was predominantly localized to the proximal region of the mouse prostate and was upregulated after castration-induced prostate involution, consistent with prostate stem cell identity and function [50].
Stem cells in the human prostate have been identified and isolated using the cell surface markers such as integrin α2β1 [51], CD133 (Prominin-1) [52], and CK6a (cytokeratin 6a) [53]. Based on high expression of α2β1 integrin, Collins and colleagues identified PSCs in the basal layer and showed that the α2β1high integrin cells represent ~1% of basal cell population in the human prostate [51]. This selected PSC population was enriched through rapid adherence to the type I collagen and showed higher colony-forming efficiency in vitro. Furthermore, when the α2β1high integrin cells were grafted subcutaneously together with stromal cells in Matrigel into nude mice, they formed prostatic gland structures in vivo. Nevertheless, these glandular-like structures, although containing basal cytokeratin positive as well as AR, PAP, and PSA positive cells, lack well-defined basal and luminal organizations [51]. However, recent studies by Missol-Kolka et al. have reported that the overall expression of CD133 in human prostate is not strictly limited to the rare basal stem and progenitor cells, but it is also expressed in some of the secretory luminal cells [54]. Furthermore, it has been shown that CD133 is downregulated in prostate cancer tissues and upregulated in the luminal cells in the vicinity of cancer area. In contrast to the human CD133, the mouse CD133 has been shown to express widely in prostate [54]. Several other surface markers, such as aldehyde dehydrogenase (ALDH), tumor-associated calcium signal transducer 2 (Trop-2), ATP-binding cassette transporter family membrane efflux pump (ABCG2), p63, and CD44, have also been reported for identification and isolation of the PSCs from the prostate tissues of human and mouse [49, 55–60]. Moreover, Trop2(+)CD44(+)CD49f(+) were used as the markers to identify basal stem cells with enhanced prostasphere-forming and tissue-regenerating abilities [61]. Unlike the murine PSCs, the human PSCs are randomly distributed within the basal epithelial layer throughout the acini and ductal regions of the prostate [51, 52]. In addition to the expression of stem-cell-specific markers, different studies have also shown that PSCs express both basal and luminal cell-specific markers in fetal and adult stages of prostate development [13, 22, 31, 62, 63]. Several studies have proposed the existence of different cell compartments based on stem-cell-driven differentiation hierarchical arrangements within the prostate epithelium [24, 29, 30, 64].
In addition to prostate epithelial stem cells, stromal stem cells (SSCs) have also been reported to exist in the prostate, where they are postulated to carry out function of replacing and regenerating local cells that are lost to normal tissue turnover, injury, or aging [65–67]. These subpopulation of SSCs expressed mesenchymal stem cell (MSC) markers such as CD34 and Sca-1, showed a high proliferative activity and ability to differentiate into fibroblastic, myogenic, adipogenic, and osteogenic lineages [68]. Of all these potential lineages, the most characteristic cell type derived from prostate stromal stem cell is fibroblast or smooth muscle cells [68, 69]. Growth factors that have regulatory effects on SSCs include members of TGF-β superfamily, the insulin-like growth factors, the fibroblast growth factors, the platelet-derived growth factor, and Wnts [70]. It is believed that the differentiation of stromal stem cells to smooth muscle cells is due to paracrine effects of prostrate epithelial cells, which permanently commit the stromal stem cells to mature into androgen receptor (AR) expressing smooth muscle cells [68].
4. Stem Cell in Benign Prostate Hyperplasia (BPH)
BPH is a slow progressive enlargement of the prostate gland which can lead to lower urinary tract symptoms (LUTS) in elderly men. It is characterized by hyperproliferation of epithelial and stromal cells in the transition zone of the prostate gland, which can be observed histopathologically [71]. Despite of its obvious importance as a major health problem, little is known in terms of biological processes that contribute to the development of BPH. To explain the etiology behind the pathogenesis of BPH, several theories, including stem cell, hormonal imbalance, apoptosis, epithelial-mesenchymal transition, embryonic awakening, and inflammation, have been proposed in recent years, and all of them seem to contribute together to some extent in the pathogenesis of BPH [12, 72]. According to stem cell theory, the stem cell population residing in the prostate gland is increased due to abnormal proliferation and apoptosis of stem cells, which may eventually contribute to BPH pathogenesis. Earlier, it was reported by Berry et al. that stem cell population is responsible for prostate gland maintenance [73]. Changes in tissue consistency and cellular hyperplasia are accompanied by downregulation of apoptotic factors and increased level of antiapoptotic factors that decrease the rate of prostatic cell death and, thus, contributing to hyperproliferation of prostatic tissue [74]. It has been reported that stromal to epithelial ratio is altered in BPH, where the ratio increases from 2 : 1 in normal glands to 5 : 1 in BPH [75]. Because stromal hyperproliferative activity is thought to promote the development of BPH, the existence of adult stem cells in the prostate stromal compartment is speculated to expand the stroma in response to stimuli during the pathogenesis of BPH [68]. Lin et al. showed that primary culture of prostate cells from BPH patients possessed many common stem cell markers, including CD30, CD44, CD54, neuronspecific enolase (NSE), CD34, vascular endothelial growth factor receptor-1 (Flt-1), and stem cell factor (SCF, also known as KIT ligand or steel factor) [68]. Compared to CD30, CD44, CD54, and NSE, the CD34, Flt-1, and SCF markers were expressed at low level. These stem cells were negative for CD11b, stem cell antigen-1 (SCA-1), SH2, AA4.1, and c-Kit. Furthermore, among this stem cell population only a fraction (5%) of the stem cells was positive for CD133 [68]. Although the origin of these stem cells is not known, the CD49(+)CD54(+)NSE(+)SCF(+) cell marker profile of these cells suggests that they are in a lineage closely related to MSCs. The stem cell population with the above profile possessed ability to differentiate or transdifferentiate into myogenic, adipogenic, and osteogenic lineages [68, 76]. Ceder et al. reported the possible existence of prostate stromal stem/progenitor cells in the adult human prostate [76]. This stromal population expressed vimentin (a mesenchymal marker), CD133, c-Kit, and SCF, with expression profiles similar to those observed in the Cajal cells of gastrointestinal tract, which represent a subset of stem cell-like cells. Several studies have identified c-Kit-expressing interstitial cells in the stromal compartment of human prostate [77–79]. Altered patterns of c-Kit expression have been reported in benign lesions of prostate and breast tissues [80, 81]. It has been shown that the c-Kit expression and number of c-Kit(+) interstitial cells were significantly higher in BPH than those of the normal prostate. Furthermore, it has been suggested that c-Kit regulates cell proliferation in prostate and plays a crucial role in the pathophysiology of BPH via altering the expression of JAK2 and STAT1 [77].
Stem cells from the BPH samples expressing CD49f, CD44, or CD133 markers have been shown to possess monolayer- and spheroid-colony-forming ability, where the highest (98%) recovery of colony-forming cells (CFCs) was achieved by CD49f(+) cells as compared to CD44(+) (17%) or CD133(+) (3%) cells [82]. These CFCs showed the capacity to undergo clonal proliferation, generates branching ductal structures, and they expressed both basal and luminal lineage markers. Further characterization of CD49f(+) cells revealed that they are comprised of two cell types: CK5(+) basal epithelial cells and CD31(+) endothelial cells [82]. Sca-1- and CD34-expressing cells isolated from BPH tissue showed a high proliferative capacity and increased plasticity, as these cells were able to differentiate into fibroblastic, myogenic, adipogenic and osteogenic lineages, similar to that of MSCs [68, 83]. Furthermore, Burger and colleagues found that cells with high Sca-1 expression had considerably more growth potential, and proliferative capabilities than cells expressing low or no Sca-1 antigen [48]. Expression of pluripotency markers such as Oct4A, Sox2, c-Myc, and Klf4 might represent a stemness-specific gene signature. A very recent study has demonstrated a relatively high expression of stemness-associated genes, including Oct4A, Sox2, c-Myc, Nanog, and Klf4, in BPH as compared to normal prostate tissue [84]. Thus, several studies have revealed the presence of stem cells that express pluripotency-associated markers and are hyperproliferative and capable of differentiation into different cell lineages within the hyperplastic prostate tissue. The presence of these high proliferative and plastic stem cells in the BPH tissue samples suggests that BPH could occur as a result of changes in the stem cell properties that could ultimately give rise to a clonal expansion of cell populations.
5. Stem Cell in Prostate Cancer (PCa)
PCa is the most prevalent and is the second most frequently diagnosed cancer and sixth leading cause of cancer-related deaths among men in the world [85]. Its etiology, although not clear, is partly attributed to multigenic and epigenetic mechanisms and the heterogeneous nature of this disease [4, 86–88]. Gleason and others described that when the transition of normal gland into adenocarcinoma of prostate takes place, its normal histological structure is disrupted and results in abnormal proliferation of the glandular structure, destruction of basement membrane, and progressive loss of basal cells (<1%) [87, 89]. In addition, AR(+) luminal cells increase and contribute in bulk of prostate mass (>99%) in PCa [90]. It is hypothesised that prostate cancer arises from AR(+) luminal cells and dramatic loss of basal cells. To support this hypothesis several investigations have been conducted [4, 91–93]. In addition, mouse basal population expressing Lin(−)Sca-1(+)CD49fhigh cells can differentiate into luminal cells in xenograft [49]. Lin(−)Sca-1(+)CD49fhigh cells from a Pten−/− mouse model display cancer stem cells phenotypes, which gave rise to adenocarcinoma after transplantation [94]. It has been reported that basal cells are the possible cells of prostate cancer origin [95]. When Goldstein et al., especially injected the mixture of urogenital sinus mesenchyme (UGSM) with human prostate basal (expressing CD49fhigh and Trop2high) or luminal cells (expressing CD49flow and Trop2high) into the subcutaneous space of immunodeficient NOD(−)SCID(−)IL(−)2Rg−/− mice, only basal cells formed prostatic duct after 16 week, whereas no prostatic duct or adenocarcinoma developed when using luminal cells [91, 95]. Luminal derived grafts lack epithelial structures and mimicked transplantation of UGSM cell alone [95]. Collins et al. reported basal cancer stem cells isolated from human prostate cancer biopsies expressing Cd44(+), α2β1high, and Cd133(+) and cell surface markers were of self renewal in vitro [96]. ALDHhigh is another marker used for cancer stem cells in human prostate cancer cell lines. Cells expressing ALDHhigh α2(+)/α6(+)/αv(+)-integrin CD44(+) showed increased tumourigenicity and metastasis in vivo and enhanced invasiveness in vitro [97]. Prostate cancer stem cells isolated from LNCaP and DU145 cell lines also showed expression of CD44(+), α2β1high, and CD133(+) markers [98, 99]. In addition, CD44(+) population isolated from xenograft human tumour and cell lines displayed high tumour initiating ability and metastasis in vitro [100]. Recently, Rajasekhar and his group isolated a small cell population expressing TRA-1-60(+)CD151(+)CD166(+) markers that displayed stem cell like features with increased NF-kB signalling along with basal cell markers, and this recapitulates the cellular hierarchy of the tumour origin from basal cells [101].
Over all data from several investigators indicated that origin of prostate cancer can be from basal stem cell population, which expresses CD44(+), α2β1high, CD133(+), ALDHhigh, and other normal basal stem cell markers.
6. Stem Cell Niche and Plasticity
Stem cells are localized in a defined microenvironment, which is known as their “niche.” The main function of a niche probably is to provide specific factors necessary for the maintenance of the stem cell properties via a combination of intracellular and intercellular signalling. These factors include a complex array of growth factors, cytokines, chemokines, and adhesive molecules known to be capable of altering the balance between proliferation, differentiation, and quiescence in stem cell populations [102, 103]. One can probably assume that this is equally true for prostatic stem cells as it is for other stem cell populations.
PSCs reside in niche areas within the basal layer of the epithelial compartment at a low percentage of approximately 0.5–1% [34]. PSCs population in the prostate undergoes a series of phenotype changes. Specifically, the basal SCs do not express the AR or the p63 protein. They have extended proliferative potential by slow cycling. According to these studies, it is postulated that, in addition to the reserve stem-cell population, there is a “TA” cell type, which is characterized by the expression of p63, as well as other basal markers such as CK5 and 14, Jagged-1, and Notch-1 [64, 104, 105]. A TA cell does not express AR protein and it is dependent, for proliferation, but not for survival, on andromedans secreted by stromal cells [105]. Under normal conditions a PSC is slow cycling in that it divides occasionally, undergoing asymmetric division to give rise to a new PSC along with a more differentiated TA daughter cell. TA cell undergoes a limited number of rapidly amplifying cell division cycles to increase the cell population derived from a single PSC before leaving the proliferative compartment to produce intermediate cell [106]. This intermediate cell expresses both epithelial specific (CK5 and 14) and luminal specific (CK8 and 18) cytokines, AR mRNA (but not protein), and prostate stem cell antigen (PSCA) [105, 107]. As an intermediate cell migrates through the basal layer, it differentiates into various terminally differentiated cell lineages of prostate epithelium.
7. Is BPH/PCa a Stem Cells Disease?
Numerous investigators demonstrated presence of stem cell in prostate tissue by using various high-end techniques that may contribute to local invasive to metastatic disease in human and research animals. In normal tissue-development, homeostasis is maintained by differentiation of stem cells and programmed cells death in regular cell cycle. This mechanism is established through interactions with tissue specific environmental factors such as growth factors and steroid hormones. Many signalling molecules and factors involvement have been reported in stem cell self-renewal and implication in cancer stem cells (CSCs) regulation (Figure 2).
Figure 2.
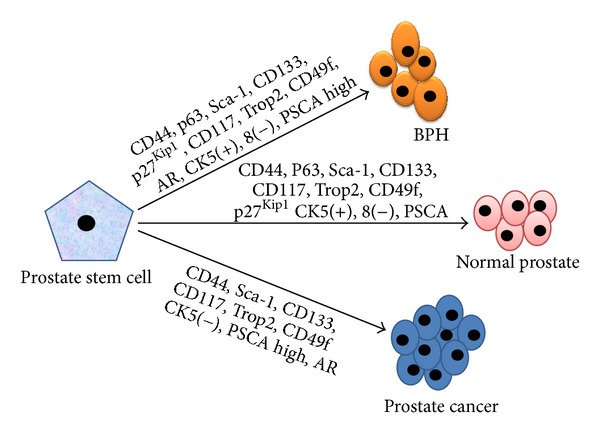
Cellular identity of stem cells in prostate. Stem cell model of normal tissue renewal, BPH and PCa.
Although the precise role of stem cells in tumourigenesis is still in debate, it is widely accepted that cancers can arise from normal stem cells which may accumulate mutation, genetic changes, and molecular pathway alterations that disrupt self-renewal control capacity (Table 1). It has been reported that, in prostate, putative stem/progenitor cells can reside in CK5(+) 8(−) basal cells. A diagnostic feature of human prostate cancer is the loss of basal cells [108], indicating cancer origin cells as basal cells. In BPH, CD133(+) cells expressed genes related to undifferentiated cells such as TDGF1 (teratocarcinoma-derived growth factor 1) and targets of the Wnt and Hedgehog developmental pathways, whereas CD133(−) cells showed upregulation of genes related to proliferation and metabolism. In cancer, CD133(+) cells specifically displayed more TA population phenotype with increased metabolic activity and proliferation, possibly explaining the transition from a relatively quiescent state to an active growing tumour phenotype. This reflects that CD133 isolates from benign and malignant tissues show biologically distinct characteristics [109]. CSCs exploit many of the signal pathways such as notch, hedgehog- and TGF-β, which play, important role in proliferation and differentiation in prostate stem cell [110, 111]. The sonic hedgehog signalling element receptor PTCH1 and glioma-associated oncogene homolog-1 (GL1) transcription factor were especially reported to be colocalized with p63 basal marker in BPH and PCa cells, expressing CD44/CK8/14. This suggests that hedgehog pathway may induce differentiation of prostate stem/progenitor cells into CD44(+)/P63(+/−) hyperplasia basal cells [112]. Other studies on DNA damage and proliferation markers p27Kip1, cyclin D3, and Ki-67, revealed interesting findings. It has been shown that p27Kip1 is significantly upregulated in BPH, whereas it is downregulated in PCa. In addition to downregulation of p27Kip1, there is also up regulation of Ki-67 and cyclin D3 in PCa [113].
Table 1.
Molecular alterations in BPH and PCa.
| Factors | Normal prostate | BPH | PCa |
|---|---|---|---|
| Prostate-specific factors | |||
| 5 α reductase | Normal | Upregulated | Upregulated |
| Androgen receptor (AR) | Normal | Upregulated | Upregulated |
| AR coactivator | Normal | Upregulated | Upregulated |
| Androgen corepressor | Normal | Upregulated | Upregulated |
| PSA level in serum | (0–4 ng/mL) | (2–8 ng/mL) | (4–10 ng/mL) |
|
| |||
| Growth factors | FGF-2,7,9 IGF 1,2 IGFBP-2 |
FGF 1,2,9 IGF-2 high IGFBP-3 |
FGF-1,2,6,8 IGF-1 high IGFBP-2 high IGFBP-3 high |
|
| |||
| NE cells | Normal | Number decrease | Number increase |
|
| |||
| Luminal cell factors | Vimentin | Vimentin increase | Vimentin over exp |
| Intracellular space normal | Intracellular space increase | Intracellular space decrease | |
| PMSA normal | PMSA decrease | PMSA increase | |
|
| |||
| Basal cells | Present | Present | Absent |
|
| |||
| Stromal cell factor | Fibroblast content normal | Fibroblast content increase | Fibroblast content increase |
| NMMHC | NMMHC increase | NMMHC | |
| Elastin | Elastin decrease | Elastin increase | |
| SMMHC | SMMHC decrease | SMMHC decrease | |
|
| |||
| Stem cell markers | CD44, P63, Sca-1, CD133, CD117, Trop2, CD49f, p27Kip1, CK5(+), 8(−), PSCA | CD44, p63, Sca-1, CD133, p27Kip1, CD117, Trop2, CD49f, AR, CK5(+), 8(−), PSCA high |
CD44, Sca-1, CD133, CD117, Trop2, CD49f, CK5(−), PSCA high, AR |
Several lines of evidence have been indicated that CSCs exhibit both stem cells and cancer cells characteristics. CSCs have the ability to form tumors when transplanted into an animal host. CSCs can be distinguished from other cells within the tumor by cell division and alterations in their gene expression profile [114].
Advanced prostate cancer is androgen independent and basal cells can be phenotypically identified in the majority of metastases [115]. Studies from several investigators revealed that tumor-initiating cells are negative for AR and p63 and expressed the stem cell markers Oct-4, Nanog, Sox-2, Nestin, CD44, CD133, and CD117. Moreover, Sca-1-positive cells having the ability with prostate-regeneration activity, showed evidence of a basal and luminal lineage [96, 100, 116, 117]. Gu et al. demonstrated human telomerase reverse transcriptase-(hTERT-) positive epithelial cells could regenerate tumor in mice that resembled the original tumor in patients [118]. These finding may be indicative of CSC role in prostate cancer.
The growing understanding of the prostate stem cell biology provides the rationale for acute approaches. But without a clear definition of stem cells in normal prostate and BPH/PCa, it is difficult to determine whether the cancer cell of origin in prostate is a stem cell, multipotent progenitor/TA cells, or a more differentiated progeny. Nonetheless, evidence exists that the cellular origin can include both basal and luminal cells.
8. Conclusion
The prostate stem cells are a key role player in prostate tumourigenesis and enlargement disorders. But their precise role in disease pathogenesis remains unknown. The prostate stromal and epithelial compartments and their reciprocal paracrine and autocrine interactions are crucial regulators of prostatic tissue homeostasis. The combination of the prostatic cell surface markers, such as Sca-1, CD133, p63, and CD49f, can aid in the identification of prostate stem cell populations. However, a prostate-specific stem cell marker has yet to be identified. The study of CSCs is still in its early stages. No standard treatments have yet been developed as a result of research on CSCs. The isolation and characterization of epithelial, stromal stem cells and cancer stem cells in the prostate will lead to understanding normal stem cells and CSCs activity to identify new strategies for the control of prostate diseases without harming normal cells milieu.
References
- 1.Gaynor ML. Isoflavones and the prevention and treatment of prostate disease: is there a role? Cleveland Clinic Journal of Medicine. 2003;70(3):203–204. doi: 10.3949/ccjm.70.3.203. [DOI] [PubMed] [Google Scholar]
- 2.Denis L, Morton MS, Griffiths K. Diet and its preventive role in prostatic disease. European Urology. 1999;35(5-6):377–387. doi: 10.1159/000019912. [DOI] [PubMed] [Google Scholar]
- 3.Jemal A, Siegel R, Ward E, Hao Y, Xu J, Thun MJ. Cancer statistics, 2009. CA Cancer Journal for Clinicians. 2009;59(4):225–249. doi: 10.3322/caac.20006. [DOI] [PubMed] [Google Scholar]
- 4.Shen MM, Abate-Shen C. Molecular genetics of prostate cancer: new prospects for old challenges. Genes and Development. 2010;24(18):1967–2000. doi: 10.1101/gad.1965810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Barclay WW, Woodruff RD, Hall MC, Cramer SD. A system for studying epithelial-stromal interactions reveals distinct inductive abilities of stromal cells from benign prostatic hyperplasia and prostate cancer. Endocrinology. 2005;146(1):13–18. doi: 10.1210/en.2004-1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Harman SM, Metter EJ, Tobin JD, Pearson J, Blackman MR. Longitudinal effects of aging on serum total and free testosterone levels in healthy men. Journal of Clinical Endocrinology and Metabolism. 2001;86(2):724–731. doi: 10.1210/jcem.86.2.7219. [DOI] [PubMed] [Google Scholar]
- 7.Nickel JC, Downey J, Young I, Boag S. Asymptomatic inflammation and/or infection in benign prostatic hyperplasia. BJU International. 1999;84(9):976–981. doi: 10.1046/j.1464-410x.1999.00352.x. [DOI] [PubMed] [Google Scholar]
- 8.McConnell JD. The pathophysiology of benign prostatic hyperplasia. Journal of Andrology. 1991;12(6):356–363. [PubMed] [Google Scholar]
- 9.Mimeault M, Batra SK. Recent advances on multiple tumorigenic cascades involved in prostatic cancer progression and targeting therapies. Carcinogenesis. 2006;27(1):1–22. doi: 10.1093/carcin/bgi229. [DOI] [PubMed] [Google Scholar]
- 10.Marcelli M, Cunningham GR. Hormonal signaling in prostatic hyperplasia and neoplasia. Journal of Clinical Endocrinology and Metabolism. 1999;84(10):3463–3468. doi: 10.1210/jcem.84.10.6083. [DOI] [PubMed] [Google Scholar]
- 11.Nikitin AY, Matoso A, Roy-Burman P. Prostate stem cells and cancer. Histology and histopathology. 2007;22(9):1043–1049. doi: 10.14670/HH-22.1043. [DOI] [PubMed] [Google Scholar]
- 12.Notara M, Ahmed A. Benign prostate hyperplasia and stem cells: a new therapeutic opportunity. Cell Biology and Toxicology. 2012;28(6):435–442. doi: 10.1007/s10565-012-9234-x. [DOI] [PubMed] [Google Scholar]
- 13.Powers GL, Marker PC. Recent advances in prostate development and links to prostatic diseases. Wiley Interdisciplinary Reviews. 2013;5(2):243–256. doi: 10.1002/wsbm.1208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Prins GS, Putz O. Molecular signaling pathways that regulate prostate gland development. Differentiation. 2008;76(6):641–659. doi: 10.1111/j.1432-0436.2008.00277.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Sugimura Y, Cunha GR, Donjacour AA. Morphogenesis of ductal networks in the mouse prostate. Biology of Reproduction. 1986;34(5):961–971. doi: 10.1095/biolreprod34.5.961. [DOI] [PubMed] [Google Scholar]
- 16.McNeal JE. Anatomy of the prostate and morphogenesis of BPH. Progress in Clinical and Biological Research. 1984;145:27–53. [PubMed] [Google Scholar]
- 17.McNeal JE, Bostwick DG. Anatomy of the prostatic urethra. Journal of the American Medical Association. 1984;251(7):890–891. [PubMed] [Google Scholar]
- 18.Timms BG. Prostate development: a historical perspective. Differentiation. 2008;76(6):565–577. doi: 10.1111/j.1432-0436.2008.00278.x. [DOI] [PubMed] [Google Scholar]
- 19.Abate-Shen C, Shen MM. Molecular genetics of prostate cancer. Genes and Development. 2000;14(19):2410–2434. doi: 10.1101/gad.819500. [DOI] [PubMed] [Google Scholar]
- 20.Bartsch G, Rohr HP. Comparative light and electron microscopic study of the human, dog and rat prostate: an approach to an experimental model for human benign prostatic hyperplasia (light and electron microscopic analysis): a review. Urologia Internationalis. 1980;35(2):91–104. doi: 10.1159/000280309. [DOI] [PubMed] [Google Scholar]
- 21.Bonkhoff H, Remberger K. Widespread distribution of nuclear androgen receptors in the basal cell layer of the normal and hyperplastic human prostate. Virchows Archiv. 1993;422(1):35–38. doi: 10.1007/BF01605130. [DOI] [PubMed] [Google Scholar]
- 22.Wang Y, Hayward SW, Cao M, Thayer KA, Cunha GR. Cell differentiation lineage in the prostate. Differentiation. 2001;68(4-5):270–279. doi: 10.1046/j.1432-0436.2001.680414.x. [DOI] [PubMed] [Google Scholar]
- 23.Long RM, Morrissey C, Fitzpatrick JM, Watson RWG. Prostate epithelial cell differentiation and its relevance to the understanding of prostate cancer therapies. Clinical Science. 2005;108(1):1–11. doi: 10.1042/CS20040241. [DOI] [PubMed] [Google Scholar]
- 24.De Marzo AM, Meeker AK, Epstein JI, Coffey DS. Prostate stem cell compartments: expression of the cell cycle inhibitor p27(Kip1) in normal, hyperplastic, and neoplastic cells. The American Journal of Pathology. 1998;153(3):911–919. doi: 10.1016/S0002-9440(10)65632-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bonkhoff H, Stein U, Remberger K. Endocrine-paracrine cell types in the prostate and prostatic adenocarcinoma are postmitotic cells. Human Pathology. 1995;26(2):167–170. doi: 10.1016/0046-8177(95)90033-0. [DOI] [PubMed] [Google Scholar]
- 26.Schalken JA, van Leenders G. Cellular and molecular biology of the prostate: stem cell biology. Urology. 2003;62(5) supplement 1:11–20. doi: 10.1016/s0090-4295(03)00758-1. [DOI] [PubMed] [Google Scholar]
- 27.Amorino GP, Parsons SJ. Neuroendocrine cells in prostate cancer. Critical Reviews in Eukaryotic Gene Expression. 2004;14(4):287–300. doi: 10.1615/critreveukaryotgeneexpr.v14.i4.40. [DOI] [PubMed] [Google Scholar]
- 28.Abrahamsson PA. Neuroendocrine differentiation in prostatic carcinoma. Prostate. 1999;39(2):135–148. doi: 10.1002/(sici)1097-0045(19990501)39:2<135::aid-pros9>3.0.co;2-s. [DOI] [PubMed] [Google Scholar]
- 29.Bonkhoff H, Stein U, Remberger K. Multidirectional differentiation in the normal, hyperplastic, and neoplastic human prostate: simultaneous demonstration of cell-specific epithelial markers. Human Pathology. 1994;25(1):42–46. doi: 10.1016/0046-8177(94)90169-4. [DOI] [PubMed] [Google Scholar]
- 30.Bonkhoff H, Remberger K. Differentiation pathways and histogenetic aspects of normal and abnormal prostatic growth: a stem cell model. Prostate. 1996;28(2):98–106. doi: 10.1002/(SICI)1097-0045(199602)28:2<98::AID-PROS4>3.0.CO;2-J. [DOI] [PubMed] [Google Scholar]
- 31.Xue Y, Smedts F, Debruyne FM, de la Rosette JJ, Schalken JA. Identification of intermediate cell types by keratin expression in the developing human prostate. Prostate. 1998;34(4):292–301. doi: 10.1002/(sici)1097-0045(19980301)34:4<292::aid-pros7>3.0.co;2-j. [DOI] [PubMed] [Google Scholar]
- 32.Hudson DL, O’Hare M, Watt FM, Masters JRW. Proliferative heterogeneity in the human prostate: evidence for epithelial stem cells. Laboratory Investigation. 2000;80(8):1243–1250. doi: 10.1038/labinvest.3780132. [DOI] [PubMed] [Google Scholar]
- 33.Takao T, Tsujimura A. Prostate stem cells: the niche and cell markers. International Journal of Urology. 2008;15(4):289–294. doi: 10.1111/j.1442-2042.2008.02047.x. [DOI] [PubMed] [Google Scholar]
- 34.Isaacs JT, Coffey DS. Etiology and disease process of benign prostatic hyperplasia. Prostate. 1989;2:33–50. doi: 10.1002/pros.2990150506. [DOI] [PubMed] [Google Scholar]
- 35.English HF, Santen RJ, Isaacs JT. Response of glandular versus basal rat ventral prostatic epithelial cells to androgen withdrawal and replacement. Prostate. 1987;11(3):229–242. doi: 10.1002/pros.2990110304. [DOI] [PubMed] [Google Scholar]
- 36.Evans GS, Chandler JA. Cell proliferation studies in the rat prostate: II. The effects of castration and androgen-induced regeneration upon basal and secretory cell proliferation. Prostate. 1987;11(4):339–351. doi: 10.1002/pros.2990110406. [DOI] [PubMed] [Google Scholar]
- 37.Verhagen APM, Aalders TW, Ramaekers FCS, Debruyne FMJ, Schalken JA. Differential expression of keratins in the basal and luminal compartments of rat prostatic epithelium during degeneration and regeneration. Prostate. 1988;13(1):25–38. doi: 10.1002/pros.2990130104. [DOI] [PubMed] [Google Scholar]
- 38.DeKlerk DP, Coffey DS. Quantitative determination of prostatic epithelial and stromal hyperplasia by a new technique. Biomorphometrics. Investigative Urology. 1978;16(3):240–245. [PubMed] [Google Scholar]
- 39.Kyprianou N, Isaacs JT. Identification of a cellular receptor for transforming growth factor-β in rat ventral prostate and its negative regulation by androgens. Endocrinology. 1988;123(4):2124–2131. doi: 10.1210/endo-123-4-2124. [DOI] [PubMed] [Google Scholar]
- 40.Montpetit M, Abrahams P, Clark AF, Tenniswood M. Androgen-independent epithelial cells of the rat ventral prostate. Prostate. 1988;12(1):13–28. doi: 10.1002/pros.2990120104. [DOI] [PubMed] [Google Scholar]
- 41.Xin L, Ide H, Kim Y, Dubey P, Witte ON. In vivo regeneration of murine prostate from dissociated cell populations of postnatal epithelia and urogenital sinus mesenchyme. Proceedings of the National Academy of Sciences of the United States of America. 2003;100(supplement 1):11896–11903. doi: 10.1073/pnas.1734139100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Azuma M, Hirao A, Takubo K, Hamaguchi I, Kitamura T, Suda T. A quantitative matrigel assay for assessing repopulating capacity of prostate stem cells. Biochemical and Biophysical Research Communications. 2005;338(2):1164–1170. doi: 10.1016/j.bbrc.2005.10.055. [DOI] [PubMed] [Google Scholar]
- 43.Tsujimura A, Koikawa Y, Salm S, et al. Proximal location of mouse prostate epithelial stem cells: a model of prostatic homeostasis. Journal of Cell Biology. 2002;157(7):1257–1265. doi: 10.1083/jcb.200202067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Goto K, Salm SN, Coetzee S, et al. Proximal prostatic stem cells are programmed to regenerate a proximal-distal ductal axis. Stem Cells. 2006;24(8):1859–1868. doi: 10.1634/stemcells.2005-0585. [DOI] [PubMed] [Google Scholar]
- 45.Nemeth JA, Lee C. Prostatic ductal system in rats: regional variation in stromal organization. Prostate. 1996;28(2):124–128. doi: 10.1002/(SICI)1097-0045(199602)28:2<124::AID-PROS8>3.0.CO;2-G. [DOI] [PubMed] [Google Scholar]
- 46.Nemeth JA, Sensibar JA, White RR, Zelner DJ, Kim IY, Lee C. Prostatic ductal system in rats: tissue-specific expression and regional variation in stromal distribution of transforming growth factor-beta 1. Prostate. 1997;33(1):64–71. doi: 10.1002/(sici)1097-0045(19970915)33:1<64::aid-pros11>3.0.co;2-j. [DOI] [PubMed] [Google Scholar]
- 47.Salm SN, Burger PE, Coetzee S, Goto K, Moscatelli D, Wilson EL. TGF-β maintains dormancy of prostatic stem cells in the proximal region of ducts. Journal of Cell Biology. 2005;170(1):81–90. doi: 10.1083/jcb.200412015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Burger PE, Xiong X, Coetzee S, et al. Sca-1 expression identifies stem cells in the proximal region of prostatic ducts with high capacity to reconstitute prostatic tissue. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(20):7180–7185. doi: 10.1073/pnas.0502761102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lawson DA, Xin L, Lukacs RU, Cheng D, Witte ON. Isolation and functional characterization of murine prostate stem cells. Proceedings of the National Academy of Sciences of the United States of America. 2007;104(1):181–186. doi: 10.1073/pnas.0609684104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Leong KG, Wang B-E, Johnson L, Gao W-Q. Generation of a prostate from a single adult stem cell. Nature. 2008;456(7223):804–810. doi: 10.1038/nature07427. [DOI] [PubMed] [Google Scholar]
- 51.Collins AT, Habib FK, Maitland NJ, Neal DE. Identification and isolation of human prostate epithelial stem cells based on α2β1-integrin expression. Journal of Cell Science. 2001;114(21):3865–3872. doi: 10.1242/jcs.114.21.3865. [DOI] [PubMed] [Google Scholar]
- 52.Richardson GD, Robson CN, Lang SH, Neal DE, Maitland NJ, Collins AT. CD133, a novel marker for human prostatic epithelial stem cells. Journal of Cell Science. 2004;117(16):3539–3545. doi: 10.1242/jcs.01222. [DOI] [PubMed] [Google Scholar]
- 53.Schmelz M, Moll R, Hesse U, et al. Identification of a stem cell candidate in the normal human prostate gland. European Journal of Cell Biology. 2005;84(2-3):341–354. doi: 10.1016/j.ejcb.2004.12.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Missol-Kolka E, Karbanová J, Janich P, et al. Prominin-1 (CD133) is not restricted to stem cells located in the basal compartment of murine and human prostate. Prostate. 2011;71(3):254–267. doi: 10.1002/pros.21239. [DOI] [PubMed] [Google Scholar]
- 55.Bhatt RI, Brown MD, Hart CA, et al. Novel method for the isolation and characterisation of the putative prostatic stem cell. Cytometry A. 2003;54(2):89–99. doi: 10.1002/cyto.a.10058. [DOI] [PubMed] [Google Scholar]
- 56.Burger PE, Gupta R, Xiong X, et al. High aldehyde dehydrogenase activity: a novel functional marker of murine prostate stem/progenitor cells. Stem Cells. 2009;27(9):2220–2228. doi: 10.1002/stem.135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Goldstein AS, Lawson DA, Cheng D, Sun W, Garraway IP, Witte ON. Trop2 identifies a subpopulation of murine and human prostate basal cells with stem cell characteristics. Proceedings of the National Academy of Sciences of the United States of America. 2008;105(52):20882–20887. doi: 10.1073/pnas.0811411106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Liu AY, True LD, Latray L, et al. Cell-cell interaction in prostate gene regulation and cytodifferentiation. Proceedings of the National Academy of Sciences of the United States of America. 1997;94(20):10705–10710. doi: 10.1073/pnas.94.20.10705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Pignon JC, Grisanzio C, Geng Y, Song J, Shivdasani RA, Signoretti S. p63-expressing cells are the stem cells of developing prostate, bladder, and colorectal epithelia. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(20):8105–8110. doi: 10.1073/pnas.1221216110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Yao M, Taylor RA, Richards MG, et al. Prostate-regenerating capacity of cultured human adult prostate epithelial cells. Cells Tissues Organs. 2010;191(3):203–212. doi: 10.1159/000240244. [DOI] [PubMed] [Google Scholar]
- 61.Garraway IP, Sun W, Tran CP, et al. Human prostate sphere-forming cells represent a subset of basal epithelial cells capable of glandular regeneration in vivo. Prostate. 2010;70(5):491–501. doi: 10.1002/pros.21083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.van Leenders G, Dijkman H, Hulsbergen-van de Kaa C, Ruiter D, Schalken J. Demonstration of intermediate cells during human prostate epithelial differentiation in situ and in vitro using triple-staining confocal scanning microscopy. Laboratory Investigation. 2000;80(8):1251–1258. doi: 10.1038/labinvest.3780133. [DOI] [PubMed] [Google Scholar]
- 63.Barclay WW, Axanova LS, Chen W, et al. Characterization of adult prostatic progenitor/stem cells exhibiting self-renewal and multilineage differentiation. Stem Cells. 2008;26(3):600–610. doi: 10.1634/stemcells.2007-0309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Taylor RA, Toivanen R, Risbridger GP. Stem cells in prostate cancer: treating the root of the problem. Endocrine-Related Cancer. 2010;17(4):R273–R285. doi: 10.1677/ERC-10-0145. [DOI] [PubMed] [Google Scholar]
- 65.Roehrborn CG, McConnell JD, Lieber M, et al. Serum prostate-specific antigen concentration is a powerful predictor of acute urinary retention and need for surgery in men with clinical benign prostatic hyperplasia. Urology. 1999;53(3):473–480. doi: 10.1016/s0090-4295(98)00654-2. [DOI] [PubMed] [Google Scholar]
- 66.Naslund MJ, Miner M. A review of the clinical efficacy and safety of 5α-reductase inhibitors for the enlarged prostate. Clinical Therapeutics. 2007;29(1):17–25. doi: 10.1016/j.clinthera.2007.01.018. [DOI] [PubMed] [Google Scholar]
- 67.Boyle P, Roehrborn C, Harkaway R, Logie J, de La Rosette J, Emberton M. 5-alpha reductase inhibition provides superior benefits to alpha blockade by preventing AUR and BPH-related surgery. European Urology. 2004;45(5):620–627. doi: 10.1016/j.eururo.2003.09.012. [DOI] [PubMed] [Google Scholar]
- 68.Lin VK, Wang S-Y, Vazquez DV, Xu CC, Zhang S, Tang L. Prostatic stromal cells derived from benign prostatic hyperplasia specimens possess stem cell like property. Prostate. 2007;67(12):1265–1276. doi: 10.1002/pros.20599. [DOI] [PubMed] [Google Scholar]
- 69.Cunha GR, Hayward SW, Dahiya R, Foster BA. Smooth muscle-epithelial interactions in normal and neoplastic prostatic development. Acta Anatomica. 1996;155(1):63–72. doi: 10.1159/000147791. [DOI] [PubMed] [Google Scholar]
- 70.Richard C, Kim G, Koikawa Y, et al. Androgens modulate the balance between VEGF and angiopoietin expression in prostate epithelial and smooth muscle cells. Prostate. 2002;50(2):83–91. doi: 10.1002/pros.10035. [DOI] [PubMed] [Google Scholar]
- 71.Schuster GA, Schuster TG. The relative amount of epithelium, muscle, connective tissue and lumen in prostatic hyperplasia as a function of the mass of tissue resected. Journal of Urology. 1999;161(4):1168–1173. [PubMed] [Google Scholar]
- 72.Tang J, Yang J. Etiopathogenesis of benign prostatic hypeprlasia. Indian Journal of Urology. 2009;25(3):312–317. doi: 10.4103/0970-1591.56179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Berry SJ, Coffey DS, Strandberg JD, Ewing LL. Effect of age, castration, and testosterone replacement on the development and restoration of canine benign prostatic hyperplasia. Prostate. 1986;9(3):295–302. doi: 10.1002/pros.2990090308. [DOI] [PubMed] [Google Scholar]
- 74.Kyprianou N, Tu H, Jacobs SC. Apoptotic versus proliferative activities in human benign prostatic hyperplasia. Human Pathology. 1996;27(7):668–675. doi: 10.1016/s0046-8177(96)90396-2. [DOI] [PubMed] [Google Scholar]
- 75.Shapiro E, Becich MJ, Hartanto V, Lepor H. The relative proportion of stromal and epithelial hyperplasia is related to the development of symptomatic benign prostate hyperplasia. Journal of Urology. 1992;147(5):1293–1297. doi: 10.1016/s0022-5347(17)37546-8. [DOI] [PubMed] [Google Scholar]
- 76.Ceder JA, Jansson L, Ehrnström RA, Rönnstrand L, Abrahamsson P-A. The characterization of epithelial and stromal subsets of candidate stem/progenitor cells in the human adult prostate. European Urology. 2008;53(3):524–532. doi: 10.1016/j.eururo.2007.11.028. [DOI] [PubMed] [Google Scholar]
- 77.Imura M, Kojima Y, Kubota Y, et al. Regulation of cell proliferation through a KIT-mediated mechanism in benign prostatic hyperplasia. Prostate. 2012;72(14):1506–1513. doi: 10.1002/pros.22500. [DOI] [PubMed] [Google Scholar]
- 78.Lammie A, Drobnjak M, Gerald W, Saad A, Cote R, Cordon-Cardo C. Expression of c-kit and kit ligand proteins in normal human tissues. Journal of Histochemistry and Cytochemistry. 1994;42(11):1417–1425. doi: 10.1177/42.11.7523489. [DOI] [PubMed] [Google Scholar]
- 79.Shafik A, Shafik I, El-Sibai O. Identification of c-kit-positive cells in the human prostate: the interstitial cells of Cajal. Archives of Andrology. 2005;51(5):345–351. doi: 10.1080/014850190944456. [DOI] [PubMed] [Google Scholar]
- 80.Kondi-Pafiti A, Arkadopoulos N, Gennatas C, Michalaki V, Frangou-Plegmenou M, Chatzipantelis P. Expression of c-kit in common benign and malignant breast lesions. Tumori. 2010;96(6):978–984. [PubMed] [Google Scholar]
- 81.Simak R, Capodieci P, Cohen DW, et al. Expression of c-kit and kit-ligand in benign and malignant prostatic tissues. Histology and Histopathology. 2000;15(2):365–374. doi: 10.14670/HH-15.365. [DOI] [PubMed] [Google Scholar]
- 82.Yamamoto H, Masters JR, Dasgupta P, et al. CD49f is an efficient marker of monolayer- and spheroid colony-forming cells of the benign and malignant human prostate. PLoS ONE. 2012;7(10) doi: 10.1371/journal.pone.0046979.e46979 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Pittenger MF, Mackay AM, Beck SC, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–147. doi: 10.1126/science.284.5411.143. [DOI] [PubMed] [Google Scholar]
- 84.Le Magnen C, Bubendorf L, Ruiz C, et al. Klf4 transcription factor is expressed in the cytoplasm of prostate cancer cells. European Journal of Cancer. 2013;49(4):955–963. doi: 10.1016/j.ejca.2012.09.023. [DOI] [PubMed] [Google Scholar]
- 85.Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer Journal for Clinicians. 2011;61(2):69–90. doi: 10.3322/caac.20107. [DOI] [PubMed] [Google Scholar]
- 86.Maitland NJ, Collins A. A tumour stem cell hypothesis for the origins of prostate cancer. BJU International. 2005;96(9):1219–1223. doi: 10.1111/j.1464-410X.2005.05744.x. [DOI] [PubMed] [Google Scholar]
- 87.Maitland NJ, Frame FM, Polson ES, Lewis JL, Collins AT. Prostate cancer stem cells: do they have a basal or luminal phenotype? Hormones and Cancer. 2011;2(1):47–61. doi: 10.1007/s12672-010-0058-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Oldridge EE, Pellacani D, Collins AT, Maitland NJ. Prostate cancer stem cells: are they androgen-responsive? Molecular and Cellular Endocrinology. 2011;360(1-2):14–24. doi: 10.1016/j.mce.2011.07.008. [DOI] [PubMed] [Google Scholar]
- 89.Gleason DF. Classification of prostatic carcinomas. Cancer Chemotherapy Reports. 1966;50(3):125–128. [PubMed] [Google Scholar]
- 90.Grisanzio C, Signoretti S. p63 in prostate biology and pathology. Journal of Cellular Biochemistry. 2008;103(5):1354–1368. doi: 10.1002/jcb.21555. [DOI] [PubMed] [Google Scholar]
- 91.Wang ZA, Shen MM. Revisiting the concept of cancer stem cells in prostate cancer. Oncogene. 2011;30(11):1261–1271. doi: 10.1038/onc.2010.530. [DOI] [PubMed] [Google Scholar]
- 92.Korsten H, Ziel-van der Made A, Ma X, van der Kwast T, Trapman J. Accumulating progenitor cells in the luminal epithelial cell layer are candidate tumor initiating cells in a Pten knockout mouse prostate cancer model. PLoS ONE. 2009;4(5) doi: 10.1371/journal.pone.0005662.e5662 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Ma X, Ziel-van der Made AC, Autar B, et al. Targeted biallelic inactivation of Pten in the mouse prostate leads to prostate cancer accompanied by increased epithelial cell proliferation but not by reduced apoptosis. Cancer Research. 2005;65(13):5730–5739. doi: 10.1158/0008-5472.CAN-04-4519. [DOI] [PubMed] [Google Scholar]
- 94.Mulholland DJ, Xin L, Morim A, Lawson D, Witte O, Wu H. Lin-Sca-1+CD49fhigh stem/progenitors are tumor-initiating cells in the Pten-null prostate cancer model. Cancer Research. 2009;69(22):8555–8562. doi: 10.1158/0008-5472.CAN-08-4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Goldstein AS, Huang J, Guo C, Garraway IP, Witte ON. Identification of a cell of origin for human prostate cancer. Science. 2010;329(5991):568–571. doi: 10.1126/science.1189992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Research. 2005;65(23):10946–10951. doi: 10.1158/0008-5472.CAN-05-2018. [DOI] [PubMed] [Google Scholar]
- 97.van den Hoogen C, van der Horst G, Cheung H, et al. High aldehyde dehydrogenase activity identifies tumor-initiating and metastasis-initiating cells in human prostate cancer. Cancer Research. 2010;70(12):5163–5173. doi: 10.1158/0008-5472.CAN-09-3806. [DOI] [PubMed] [Google Scholar]
- 98.Hurt EM, Kawasaki BT, Klarmann GJ, Thomas SB, Farrar WL. CD44+CD24- prostate cells are early cancer progenitor/stem cells that provide a model for patients with poor prognosis. British Journal of Cancer. 2008;98(4):756–765. doi: 10.1038/sj.bjc.6604242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wei C, Guomin W, Yujun L, Ruizhe Q. Cancer stem-like cells in human prostate carcinoma cells DU145: the seeds of the cell line? Cancer Biology and Therapy. 2007;6(5):763–768. doi: 10.4161/cbt.6.5.3996. [DOI] [PubMed] [Google Scholar]
- 100.Patrawala L, Calhoun T, Schneider-Broussard R, et al. Highly purified CD44+ prostate cancer cells from xenograft human tumors are enriched in tumorigenic and metastatic progenitor cells. Oncogene. 2006;25(12):1696–1708. doi: 10.1038/sj.onc.1209327. [DOI] [PubMed] [Google Scholar]
- 101.Rajasekhar VK, Studer L, Gerald W, Socci ND, Scher HI. Tumour-initiating stem-like cells in human prostate cancer exhibit increased NF-κB signalling. Nature Communications. 2011;2, article 162 doi: 10.1038/ncomms1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Whetton AD, Graham GJ. Homing and mobilization in the stem cell niche. Trends in Cell Biology. 1999;9(6):233–238. doi: 10.1016/s0962-8924(99)01559-7. [DOI] [PubMed] [Google Scholar]
- 103.Spradling A, Drummond-Barbosa D, Kai T. Stem cells find their niche. Nature. 2001;414(6859):98–104. doi: 10.1038/35102160. [DOI] [PubMed] [Google Scholar]
- 104.Litvinov IV, Vander Griend DJ, Xu Y, Antony L, Dalrymple SL, Isaacs JT. Low-calcium serum-free defined medium selects for growth of normal prostatic epithelial stem cells. Cancer Research. 2006;66(17):8598–8607. doi: 10.1158/0008-5472.CAN-06-1228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Isaacs JT. Prostate stem cells and benign prostatic hyperplasia. Prostate. 2008;68(9):1025–1034. doi: 10.1002/pros.20763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Hudson DL. Epithelial stem cells in human prostate growth and disease. Prostate Cancer and Prostatic Diseases. 2004;7(3):188–194. doi: 10.1038/sj.pcan.4500745. [DOI] [PubMed] [Google Scholar]
- 107.Tran CP, Lin C, Yamashiro J, Reiter RE. Prostate stem cell antigen is a marker of late intermediate prostate epithelial cells. Molecular Cancer Research. 2002;1(2):113–121. [PubMed] [Google Scholar]
- 108.Humphrey PA. Diagnosis of adenocarcinoma in prostate needle biopsy tissue. Journal of Clinical Pathology. 2007;60(1):35–42. doi: 10.1136/jcp.2005.036442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Shepherd CJ, Rizzo S, Ledaki I, et al. Expression profiling of CD133+ and CD133- epithelial cells from human prostate. Prostate. 2008;68(9):1007–1024. doi: 10.1002/pros.20765. [DOI] [PubMed] [Google Scholar]
- 110.Collins AT, Maitland NJ. Prostate cancer stem cells. European Journal of Cancer. 2006;42(9):1213–1218. doi: 10.1016/j.ejca.2006.01.037. [DOI] [PubMed] [Google Scholar]
- 111.Blum R, Gupta R, Burger PE, et al. Molecular signatures of prostate stem cells reveal novel signaling pathways and provide insights into prostate cancer. PLoS ONE. 2009;4(5) doi: 10.1371/journal.pone.0005722.e5722 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Chen B-Y, Liu J-Y, Chang H-H, et al. Hedgehog is involved in prostate basal cell hyperplasia formation and its progressing towards tumorigenesis. Biochemical and Biophysical Research Communications. 2007;357(4):1084–1089. doi: 10.1016/j.bbrc.2007.04.091. [DOI] [PubMed] [Google Scholar]
- 113.Nikoleishvili D, Pertia A, Trsintsadze O, Gogokhia N, Managadze L, Chkhotua A. Expression of p27(Kip1), cyclin D3 and Ki67 in BPH, prostate cancer and hormone-treated prostate cancer cells. International Urology and Nephrology. 2008;40(4):953–959. doi: 10.1007/s11255-008-9350-y. [DOI] [PubMed] [Google Scholar]
- 114.Rosen JM, Jordan CT. The increasing complexity of the cancer stem cell paradigm. Science. 2009;324(5935):1670–1673. doi: 10.1126/science.1171837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Liu AY, Nelson PS, van Engh GD, Hood L. Human prostate epithelial cell-type cDNA libraries and prostate expression patterns. Prostate. 2002;50(2):92–103. doi: 10.1002/pros.10036. [DOI] [PubMed] [Google Scholar]
- 116.van Leenders GJLH, Schalken JA. Stem cell differentiation within the human prostate epithelium: implications for prostate carcinogenesis. BJU International. 2001;88(2) Supplement:35–42. doi: 10.1046/j.1464-410x.2001.00117.x. [DOI] [PubMed] [Google Scholar]
- 117.Craft N, Chhor C, Tran C, et al. Evidence for clonal outgrowth of androgen-independent prostate cancer cells from androgen-dependent tumors through a two-step process. Cancer Research. 1999;59(19):5030–5036. [PubMed] [Google Scholar]
- 118.Gu A, Yuan J, Wills M, Kasper S. Prostate cancer cells with stem cell characteristics reconstitute the original human tumor in vivo. Cancer Research. 2007;67(10):4807–4815. doi: 10.1158/0008-5472.CAN-06-4608. [DOI] [PubMed] [Google Scholar]